
TNFSF15 Inhibits Blood Retinal Barrier Breakdown Induced by Diabetes

 ,
,
Abstract
:1. Introduction
2. Results
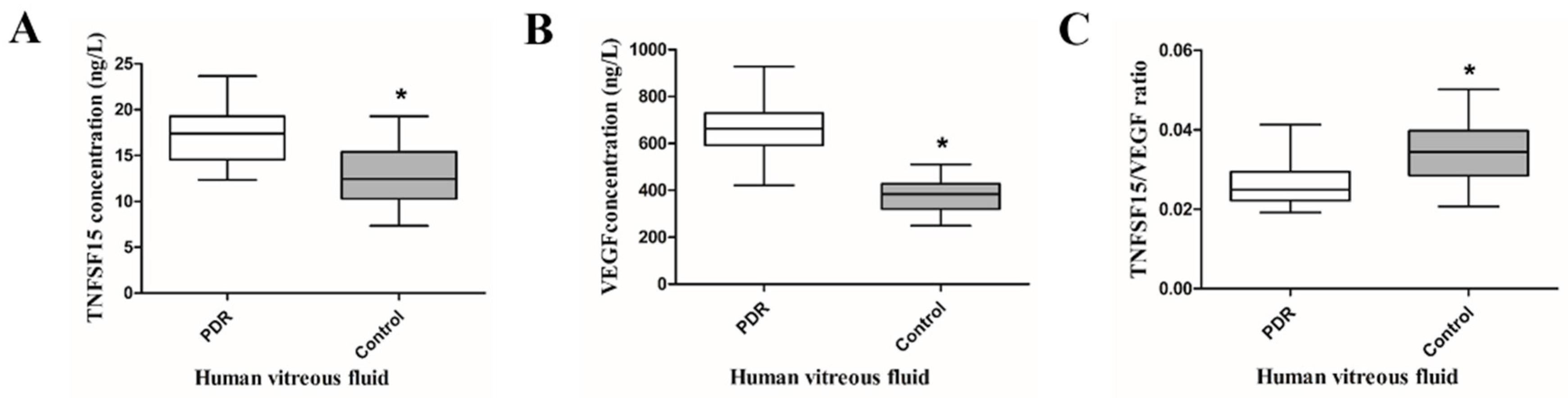
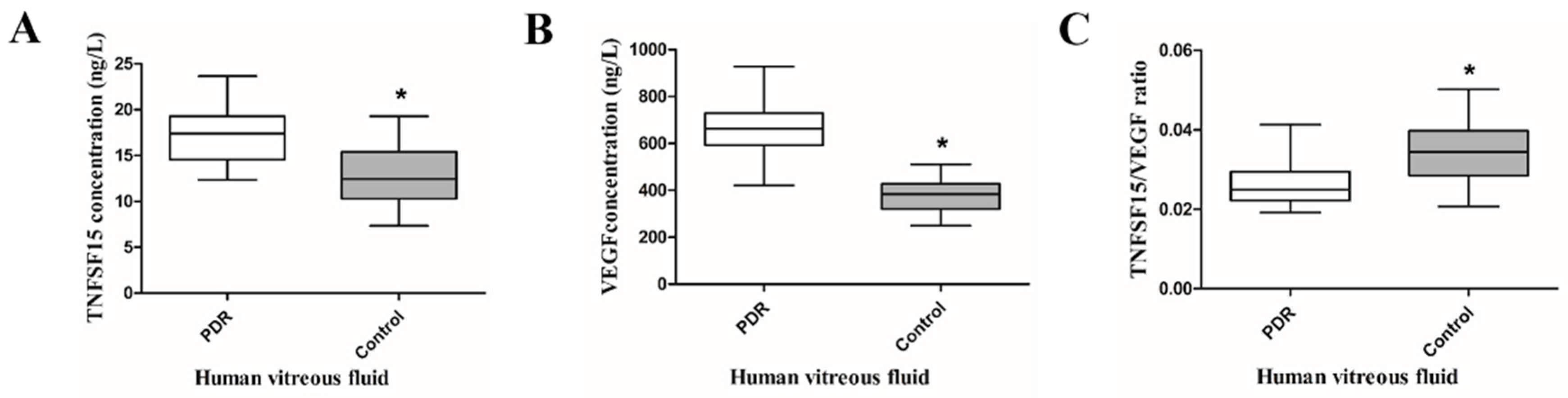
2.1. TNFSF15 and VEGF Increased, but the TNFSF15/VEGF Ratio Decreased in the Vitreous Fluid of PDR Patients
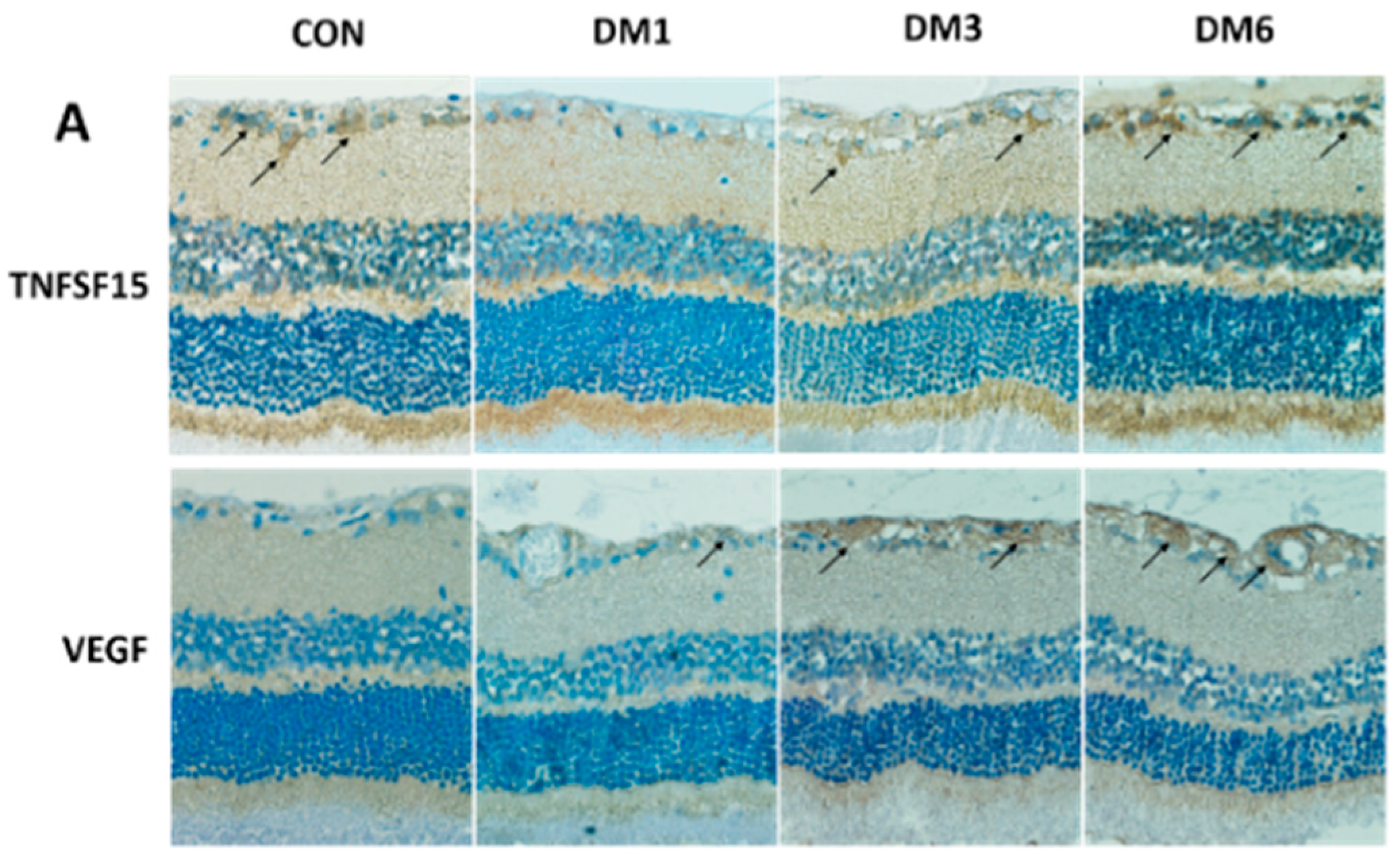
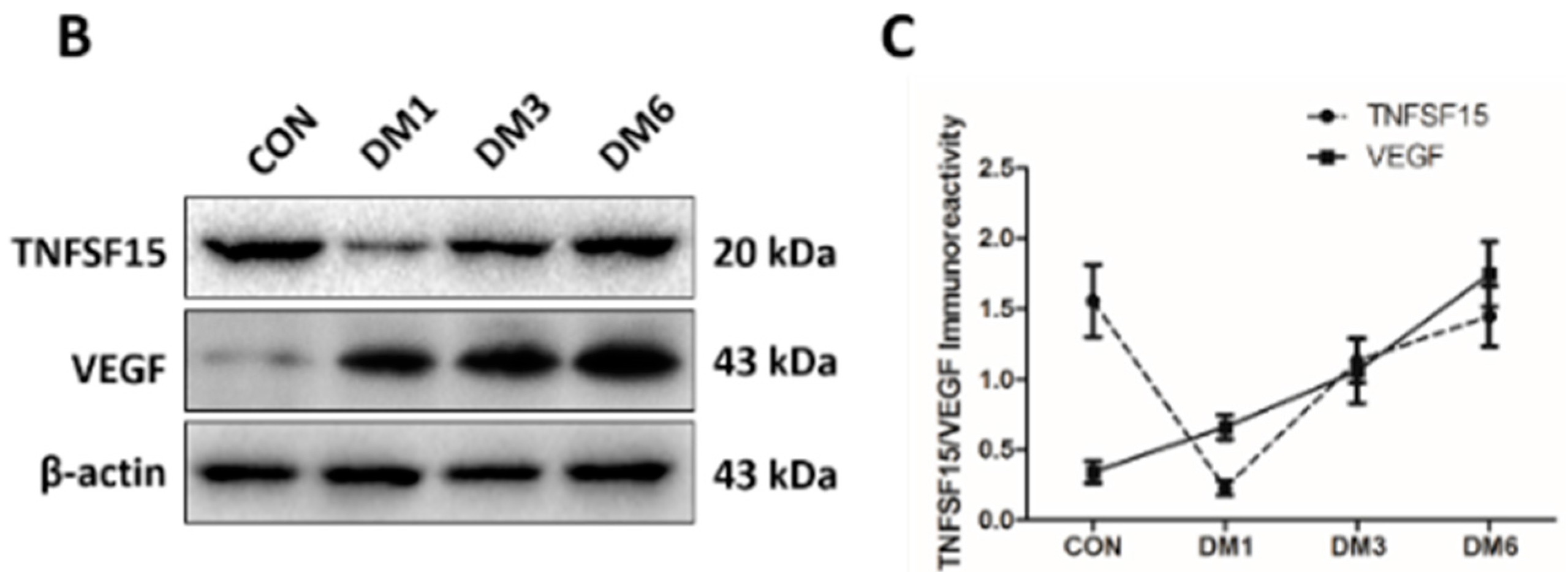
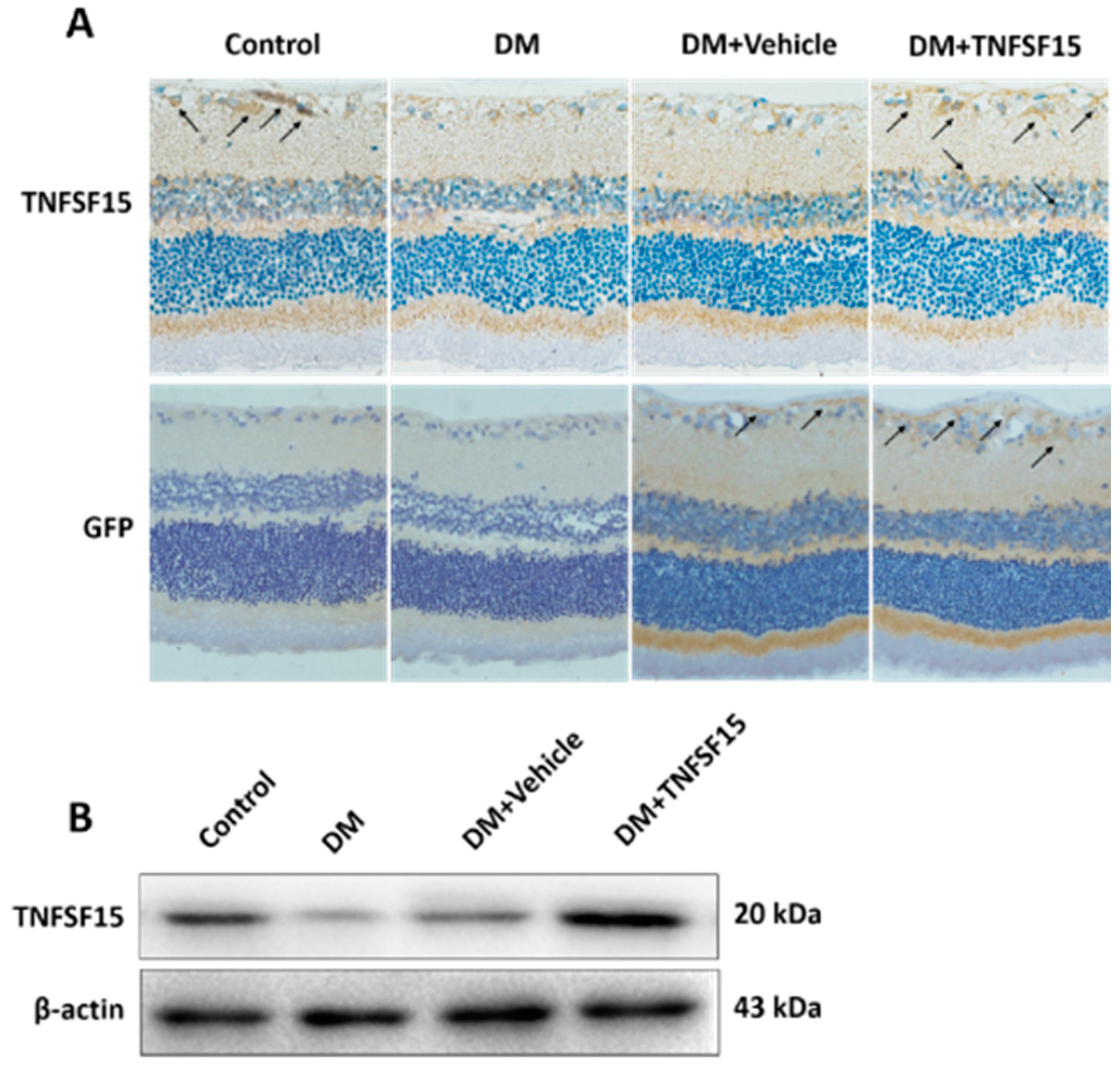
2.2. The Expression of TNFSF15 and VEGF Changed in the Retina of Diabetic Rats
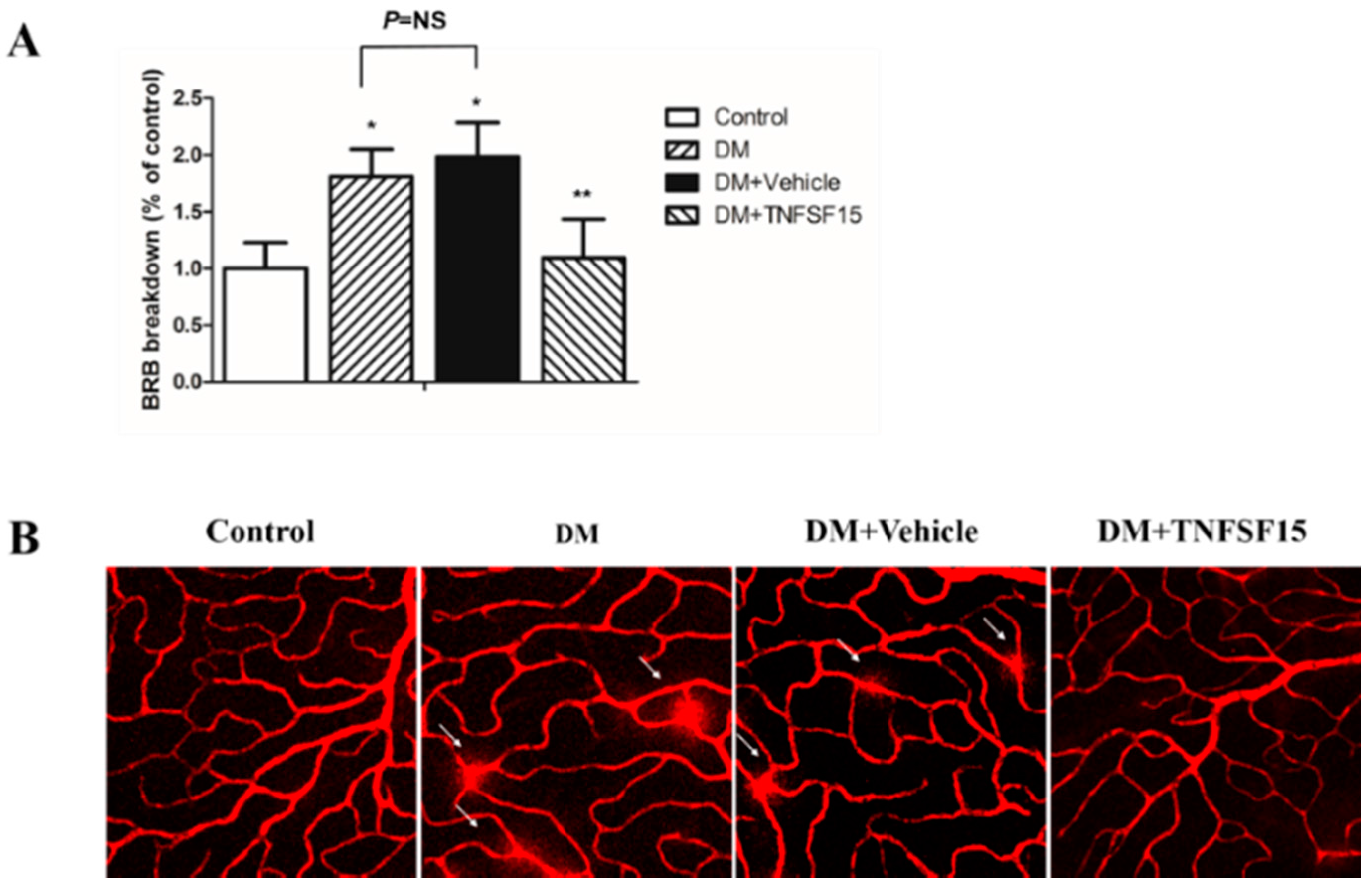
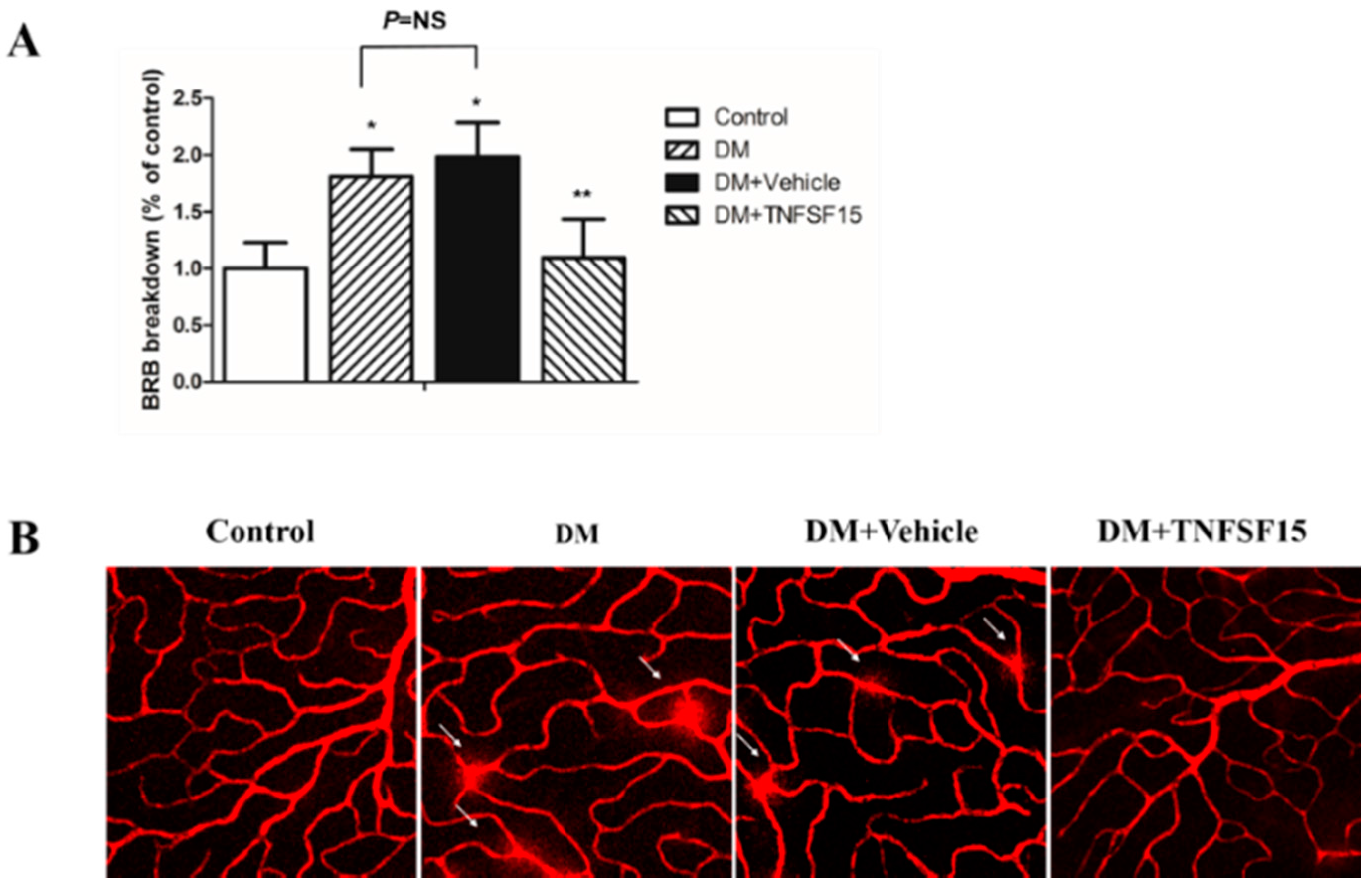
2.3. TNFSF15 Inhibits Increased BRB Permeability Induced by Diabetes in Rats
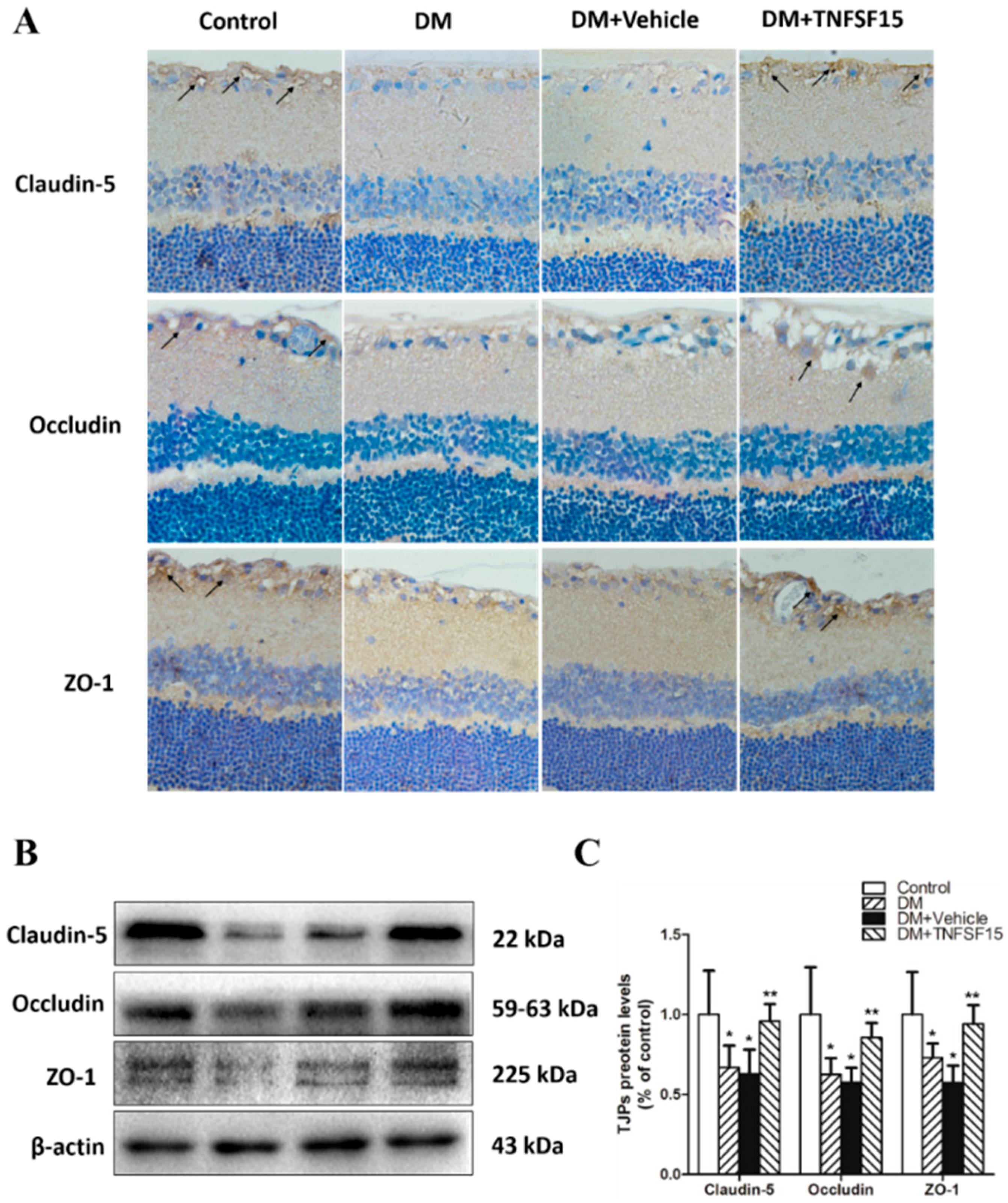
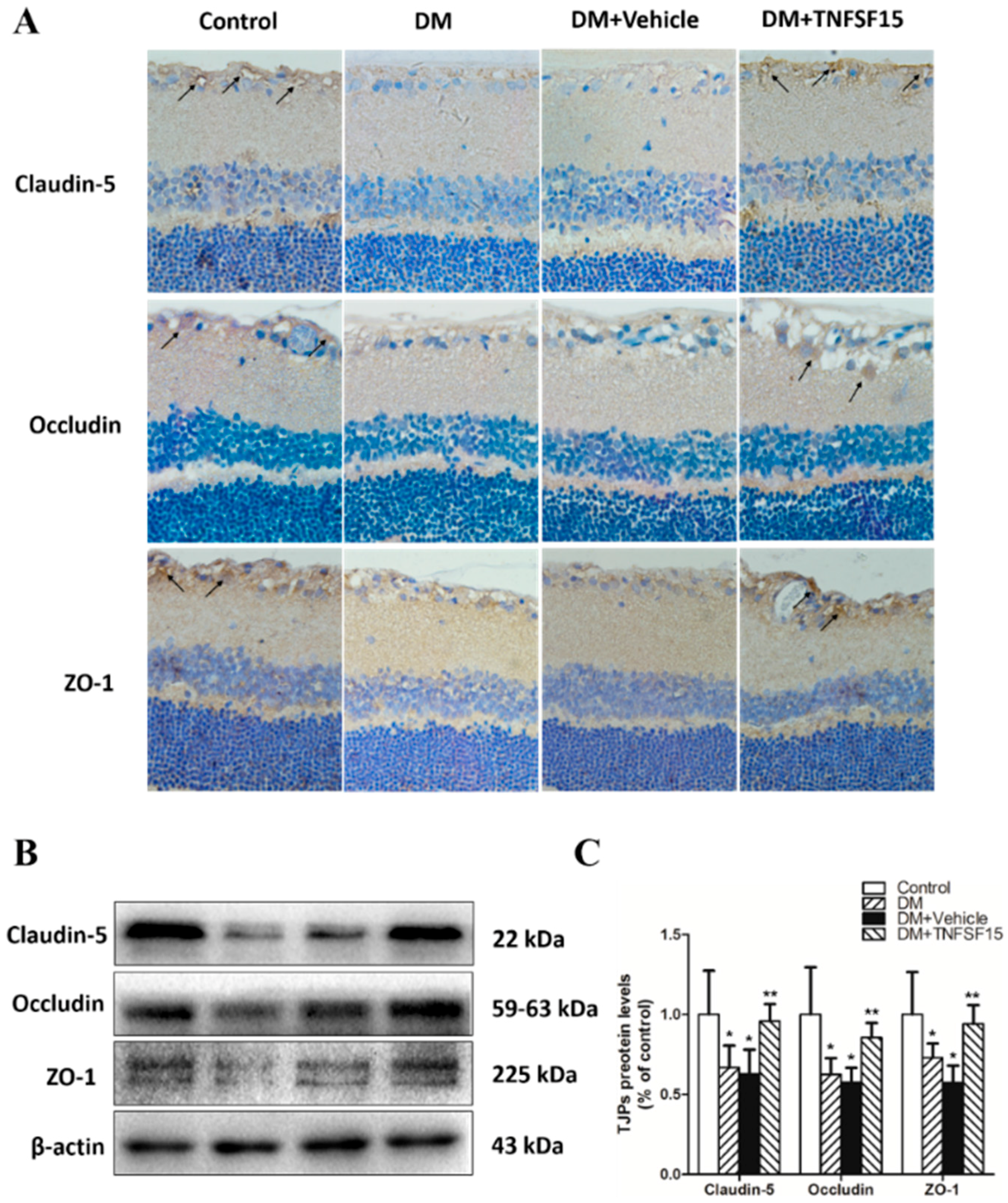
2.4. TNFSF15 Treatment Induced the Expression of TJPs in Retinal Vessels
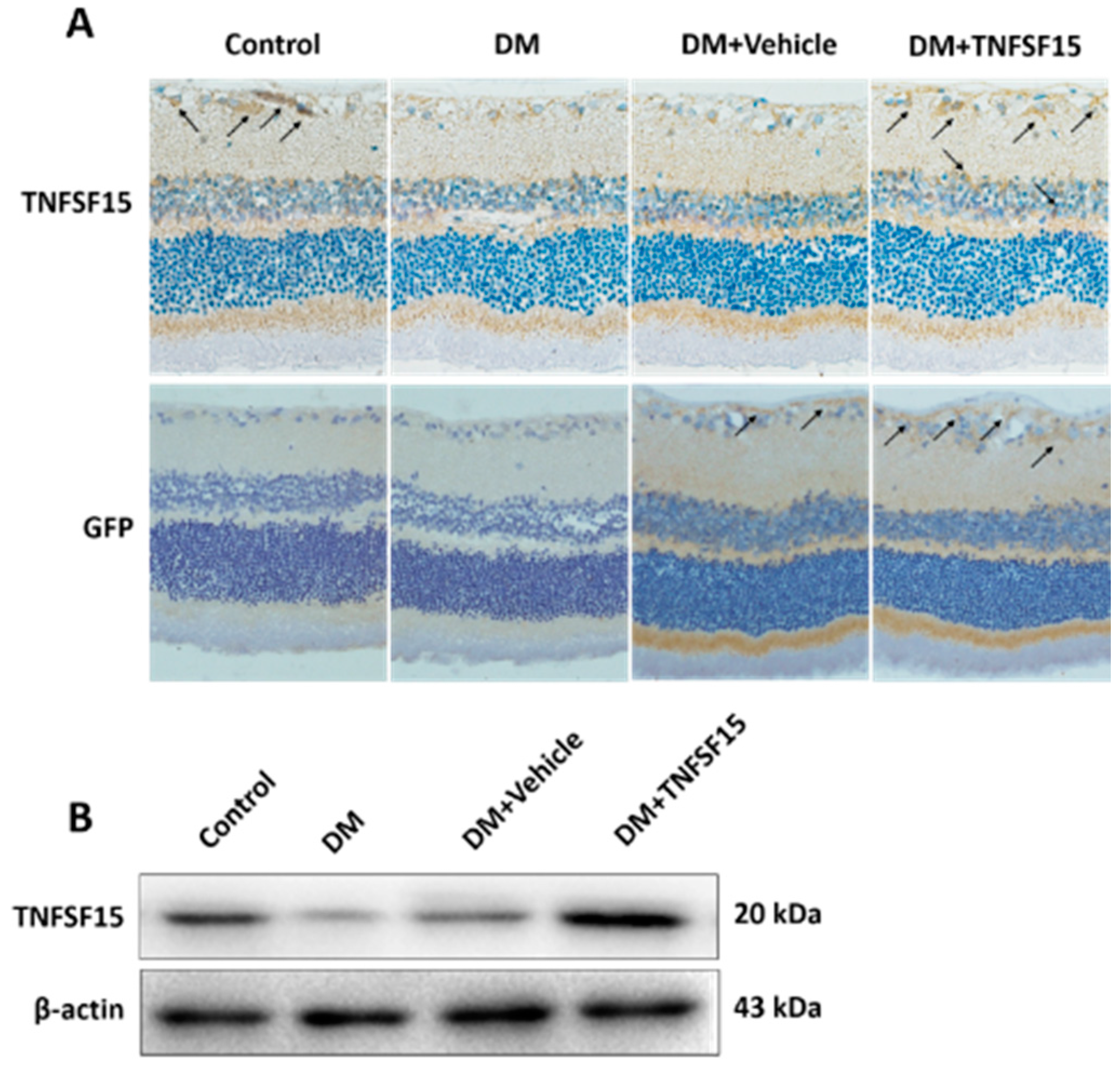
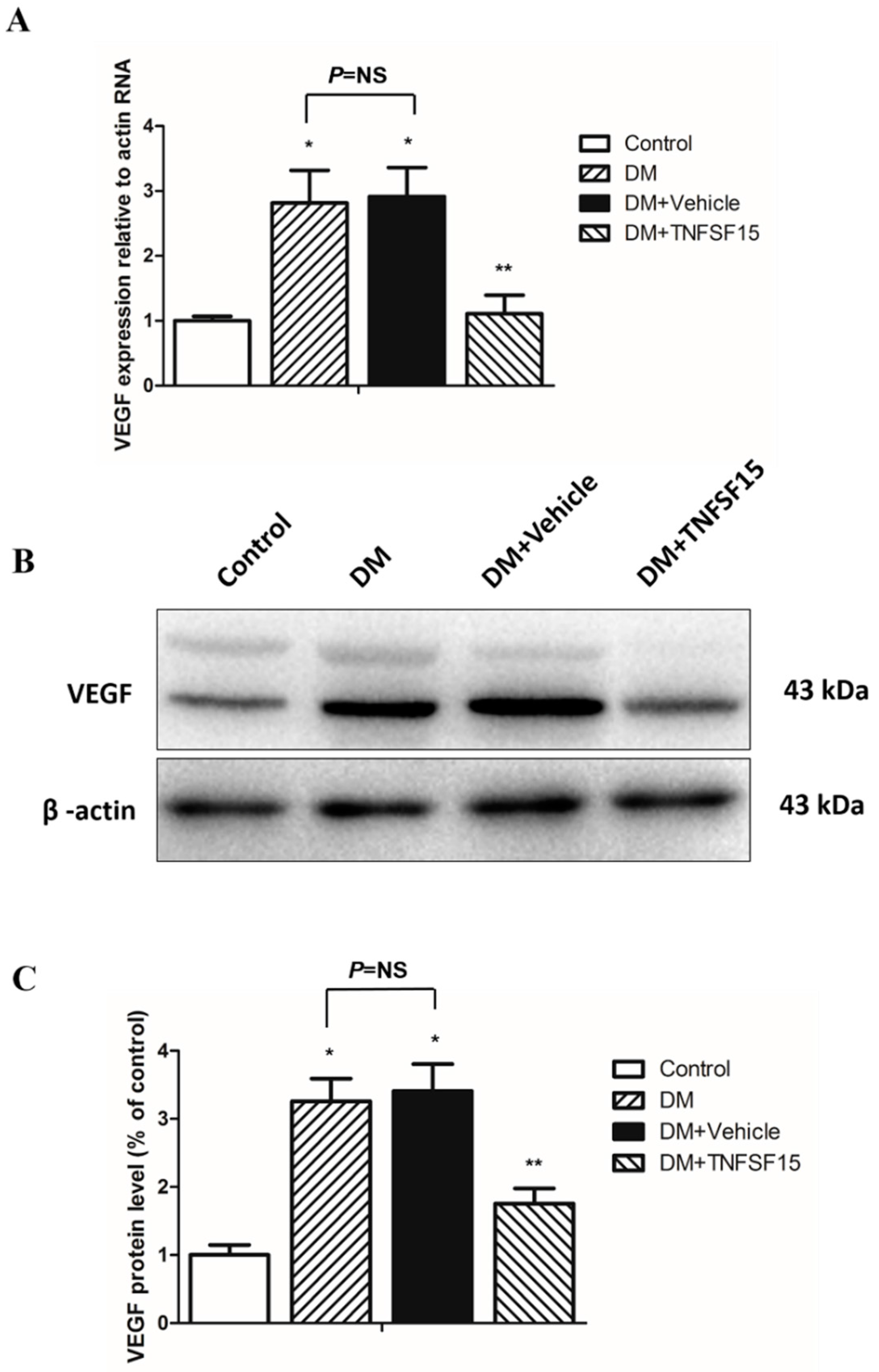
2.5. TNFSF15 Downregulated VEGF Expression in the Retina of Diabetic Rats
3. Discussion
4. Materials and Methods
4.1. Human Vitreous Fluid Samples
4.2. Measurement of TNFSF15 and VEGF Levels
4.3. Animals
4.4. Intravitreal Injection
4.5. Immunohistochemical Staining of TNFSF15, VEGF, Claudin-5, Occludin, and ZO-1
4.6. Blood–Retinal Barrier Quantitation
4.7. Visualization of Retinal Vessel Leakage
4.8. Western Blot Analysis
4.9. Quantitative Real-Time Polymerase Chain Reaction
4.10. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| TNFSF15 | tumor necrosis factor superfamily 15 |
| VEGF | vascular endothelial growth factor |
| PDR | proliferative diabetic retinopathy |
| TJPs | tight junction proteins |
| BRB | blood retinal barrier |
| EB | Evans Blue |
| DR | diabetic retinopathy |
| ECs | endothelial cells |
| EPC | endothelial progenitor cell |
| mFlt1 | membrane-bound VEGF receptor 1 |
| sFlt1 | soluble Flt1 |
| BBB | blood brain barrier |
| CAM | chorioallantoic membrane |
| STZ | streptozotocin |
| NFL | nerve fibre layer |
| GCL | ganglion cell layer |
| INL | inner nuclear layer |
| MCP-1 | monocyte chemoattractant protein-1 |
| TNF | tumor necrosis factor |
| DM | diabetes mellitus |
| PPV | pars plana vitrectomy |
| TRD | tractional retinal detachment |
| NCVH | non-clearing vitreous haemorrhage |
| IMH | idiopathic macular holes |
| ERM | idiopathic epiretinal membranes |
| RVO | retinal vein occlusion |
| RAO | retinal artery occlusion |
| RRD | rhegmatogenous retinal detachment |
| ELISA | enzyme-linked immunosorbent assay |
| ARVO | the Association for Research in Vision and Ophthalmology |
| LV-TNFSF15-GFP | lentivirus vector-TNFSF-15-green fluorescent protein |
| LV-NC-GFP | lentivirus vector-noncontain-GFP |
| PMSF | phenylmethylsulfonyl fluoride |
| DTT | dithiothreitol |
| PVDF | polyvinylidene fluoride |
References
- Klein, B.E. Overview of epidemiologic studies of diabetic retinopathy. Ophthalmic Epidemiol. 2007, 14, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Yau, J.W.; Rogers, S.L.; Kawasaki, R.; Lamoureux, E.L.; Kowalski, J.W.; Bek, T.; Chen, S.J.; Dekker, J.M.; Fletcher, A.; Grauslund, J.; et al. Global prevalence and major risk factors of diabetic retinopathy. Diabetes Care 2012, 35, 556–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Wu, X.; Liu, L.; Geng, J.; Yuan, Z.; Shan, Z.; Chen, L. Prevalence of diabetic retinopathy in mainland China: A meta-analysis. PLoS ONE 2012, 7, e45264. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; Chan, P.S. Oxidative stress and diabetic retinopathy. Exp. Diabetes Res. 2007, 2007, 43603. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Kern, T.S. Inflammation in diabetic retinopathy. Prog. Retin. Eye Res. 2011, 30, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, H.; Rojas, M.; Caldwell, R.W.; Caldwell, R.B. Anti-inflammatory therapy for diabetic retinopathy. Immunotherapy 2011, 3, 609–628. [Google Scholar] [CrossRef] [PubMed]
- Zong, H.; Ward, M.; Stitt, A.W. AGEs, RAGE, and diabetic retinopathy. Curr. Diabetes Rep. 2011, 11, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Amadio, M.; Bucolo, C.; Leggio, G.M.; Drago, F.; Govoni, S.; Pascale, A. The PKCbeta/HuR/VEGF pathway in diabetic retinopathy. Biochem. Pharmacol. 2010, 80, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Lupo, G.; Motta, C.; Giurdanella, G.; Anfuso, C.D.; Alberghina, M.; Drago, F.; Salomone, S.; Bucolo, C. Role of phospholipases A2 in diabetic retinopathy: In vitro and in vivo studies. Biochem. Pharmacol. 2013, 86, 1603–1613. [Google Scholar] [CrossRef] [PubMed]
- Giurdanella, G.; Anfuso, C.D.; Olivieri, M.; Lupo, G.; Caporarello, N.; Eandi, C.M.; Drago, F.; Bucolo, C.; Salomone, S. Aflibercept, bevacizumab and ranibizumab prevent glucose-induced damage in human retinal pericytes in vitro, through a PLA2/COX-2/VEGF-A pathway. Biochem. Pharmacol. 2015, 96, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, H.; Nie, J.; Wang, F. Protective factors in diabetic retinopathy: Focus on blood-retinal barrier. Discov. Med. 2014, 18, 105–112. [Google Scholar] [PubMed]
- Mohan, N.; Monickaraj, F.; Balasubramanyam, M.; Rema, M.; Mohan, V. Imbalanced levels of angiogenic and angiostatic factors in vitreous, plasma and postmortem retinal tissue of patients with proliferative diabetic retinopathy. J. Diabetes Complicat. 2012, 26, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Zhang, S.X.; Lu, K.; Chen, Y.; Mott, R.; Sato, S.; Ma, J.X. Decreased expression of pigment epithelium-derived factor is involved in the pathogenesis of diabetic nephropathy. Diabetes 2005, 54, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.X.; Wang, J.J.; Gao, G.; Parke, K.; Ma, J.X. Pigment epithelium-derived factor downregulates vascular endothelial growth factor (VEGF) expression and inhibits VEGF-VEGF receptor 2 binding in diabetic retinopathy. J. Mol. Endocrinol. 2006, 37, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rask-Madsen, C.; King, G.L. Kidney complications: Factors that protect the diabetic vasculature. Nat. Med. 2010, 16, 40–41. [Google Scholar] [CrossRef] [PubMed]
- Jeong, I.K.; King, G.L. New perspectives on diabetic vascular complications: The loss of endogenous protective factors induced by hyperglycemia. Diabetes Metab. J. 2011, 35, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Weis, S.M.; Cheresh, D.A. Pathophysiological consequences of VEGF-induced vascular permeability. Nature 2005, 437, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Qaum, T.; Xu, Q.; Joussen, A.M.; Clemens, M.W.; Qin, W.; Miyamoto, K.; Hassessian, H.; Wiegand, S.J.; Rudge, J.; Yancopoulos, G.D.; et al. VEGF-initiated blood-retinal barrier breakdown in early diabetes. Investig Ophthalmol. Vis. Sci. 2001, 42, 2408–2413. [Google Scholar]
- Murata, T.; Nakagawa, K.; Khalil, A.; Ishibashi, T.; Inomata, H.; Sueishi, K. The relation between expression of vascular endothelial growth factor and breakdown of the blood-retinal barrier in diabetic rat retinas. Lab. Investig. 1996, 74, 819–825. [Google Scholar] [PubMed]
- Gilbert, R.E.; Vranes, D.; Berka, J.L.; Kelly, D.J.; Cox, A.; Wu, L.L.; Stacker, S.A.; Cooper, M.E. Vascular endothelial growth factor and its receptors in control and diabetic rat eyes. Lab. Investig. 1998, 78, 1017–1027. [Google Scholar] [PubMed]
- Wang, J.; Chen, S.; Jiang, F.; You, C.; Mao, C.; Yu, J.; Han, J.; Zhang, Z.; Yan, H. Vitreous and plasma VEGF levels as predictive factors in the progression of proliferative diabetic retinopathy after vitrectomy. PLoS ONE 2014, 9, e110531. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, K.; Ogata, N.; Jo, N.; Shima, C.; Matsuoka, M.; Matsumura, M. Levels of vascular endothelial growth factor and pigment epithelium-derived factor in eyes before and after intravitreal injection of bevacizumab. Jpn. J. Ophthalmol. 2009, 53, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Costagliola, C.; Daniele, A.; dell'Omo, R.; Romano, M.R.; Aceto, F.; Agnifili, L.; Semeraro, F.; Porcellini, A. Aqueous humor levels of vascular endothelial growth factor and adiponectin in patients with type 2 diabetes before and after intravitreal bevacizumab injection. Exp. Eye Res. 2013, 110, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Sawada, O.; Kawamura, H.; Kakinoki, M.; Sawada, T.; Ohji, M. Vascular endothelial growth factor in aqueous humor before and after intravitreal injection of bevacizumab in eyes with diabetic retinopathy. Arch. Ophthalmol. 2007, 125, 1363–1366. [Google Scholar] [CrossRef] [PubMed]
- Simo, R.; Sundstrom, J.M.; Antonetti, D.A. Ocular Anti-VEGF therapy for diabetic retinopathy: The role of VEGF in the pathogenesis of diabetic retinopathy. Diabetes Care 2014, 37, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Ip, M.S.; Domalpally, A.; Hopkins, J.J.; Wong, P.; Ehrlich, J.S. Long-term effects of ranibizumab on diabetic retinopathy severity and progression. Arch. Ophthalmol. 2012, 130, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Agard, E.; Elchehab, H.; Ract-Madoux, G.; Russo, A.; Lagenaite, C.; Dot, C. Repeated intravitreal anti-vascular endothelial growth factor injections can induce iatrogenic ocular hypertension, especially in patients with open-angle glaucoma. Can. J. Ophthalmol. 2015, 50, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Cheungpasitporn, W.; Chebib, F.T.; Cornell, L.D.; Brodin, M.L.; Nasr, S.H.; Schinstock, C.A.; Stegall, M.D.; Amer, H. Intravitreal Antivascular Endothelial Growth Factor Therapy May Induce Proteinuria and Antibody Mediated Injury in Renal Allografts. Transplantation 2015, 99, 2382–2386. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.B.; Harrop, J.; Reddy, M.; Young, P.; Terrett, J.; Emery, J.; Moore, G.; Truneh, A. Characterization of a novel TNF-like ligand and recently described TNF ligand and TNF receptor superfamily genes and their constitutive and inducible expression in hematopoietic and non-hematopoietic cells. Gene 1997, 204, 35–46. [Google Scholar] [CrossRef]
- Chew, L.J.; Pan, H.; Yu, J.; Tian, S.; Huang, W.Q.; Zhang, J.Y.; Pang, S.; Li, L.Y. A novel secreted splice variant of vascular endothelial cell growth inhibitor. FASEB J. 2002, 16, 742–744. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Ni, J.; Jiang, G.W.; Lu, J.; Xing, L.; Lincoln, C.; Carter, K.C.; Janat, F.; Kozak, D.; Xu, S.; et al. VEGI, a novel cytokine of the tumor necrosis factor family, is an angiogenesis inhibitor that suppresses the growth of colon carcinomas in vivo. FASEB J. 1999, 13, 181–189. [Google Scholar] [PubMed]
- Zhai, Y.; Yu, J.; Iruela-Arispe, L.; Huang, W.Q.; Wang, Z.; Hayes, A.J.; Lu, J.; Jiang, G.; Rojas, L.; Lippman, M.E.; et al. Inhibition of angiogenesis and breast cancer xenograft tumor growth by VEGI, a novel cytokine of the TNF superfamily. Int. J. Cancer 1999, 82, 131–136. [Google Scholar] [CrossRef]
- Deng, W.; Gu, X.; Lu, Y.; Gu, C.; Zheng, Y.; Zhang, Z.; Chen, L.; Yao, Z.; Li, L.Y. Down-modulation of TNFSF15 in ovarian cancer by VEGF and MCP-1 is a pre-requisite for tumor neovascularization. Angiogenesis 2012, 15, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Parr, C.; Gan, C.H.; Watkins, G.; Jiang, W.G. Reduced vascular endothelial growth inhibitor (VEGI) expression is associated with poor prognosis in breast cancer patients. Angiogenesis 2006, 9, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yang, Z.; Tsuji, T.; Gong, J.; Xie, J.; Chen, C.; Li, W.; Amar, S.; Luo, Z. LITAF and TNFSF15, two downstream targets of AMPK, exert inhibitory effects on tumor growth. Oncogene 2011, 30, 1892–1900. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, L.Y. TNFSF15 Modulates Neovascularization and Inflammation. Cancer Microenviron. 2012, 5, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Sanders, A.J.; Ye, L.; Kynaston, H.G.; Jiang, W.G. Expression of vascular endothelial growth inhibitor (VEGI) in human urothelial cancer of the bladder and its effects on the adhesion and migration of bladder cancer cells in vitro. Anticancer Res. 2010, 30, 87–95. [Google Scholar] [PubMed]
- Zhang, N.; Sanders, A.J.; Ye, L.; Jiang, W.G. Vascular endothelial growth inhibitor in human cancer (Review). Int. J. Mol. Med. 2009, 24, 3–8. [Google Scholar] [PubMed]
- Conway, K.P.; Price, P.; Harding, K.G.; Jiang, W.G. The role of vascular endothelial growth inhibitor in wound healing. Int. Wound J. 2007, 4, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.W.; Qin, T.T.; Xu, L.X.; Zhang, K.; Yang, G.L.; Li, J.; Xiao, H.Y.; Zhang, Z.S.; Li, L.Y. TNFSF15 inhibits vasculogenesis by regulating relative levels of membrane-bound and soluble isoforms of VEGF receptor 1. Proc. Natl. Acad. Sci. USA 2013, 110, 13863–13868. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Zhao, Z.; Yu, G.; Zhou, Z.; Zhou, Y.; Hu, T.; Jiang, R.; Zhang, J. VEGI attenuates the inflammatory injury and disruption of blood-brain barrier partly by suppressing the TLR4/NF-kappaB signaling pathway in experimental traumatic brain injury. Brain Res. 2015, 1622, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Wang, Y.; Zhang, M.; Pan, W.; Qi, Z.T.; Cao, G.W. Effects of endostatin-vascular endothelial growth inhibitor chimeric recombinant adenoviruses on antiangiogenesis. World J. Gastroenterol. 2004, 10, 1409–1414. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, B. Inhibition of corneal neovascularization by vascular endothelia growth inhibitor gene. Int. J. Ophthalmol. 2010, 3, 295–298. [Google Scholar] [PubMed]
- Abu El-Asrar, A.M.; De Hertogh, G.; Nawaz, M.I.; Siddiquei, M.M.; van den Eynde, K.; Mohammad, G.; Opdenakker, G.; Geboes, K. The Tumor Necrosis Factor Superfamily Members TWEAK, TNFSF15 and Fibroblast Growth Factor-Inducible Protein 14 Are Upregulated in Proliferative Diabetic Retinopathy. Ophthalmic Res. 2015, 53, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Bussolino, F.; Mantovani, A.; Persico, G. Molecular mechanisms of blood vessel formation. Trends Biochem. Sci. 1997, 22, 251–256. [Google Scholar] [CrossRef]
- Huang, H.; Gandhi, J.K.; Zhong, X.; Wei, Y.; Gong, J.; Duh, E.J.; Vinores, S.A. TNFalpha is required for late BRB breakdown in diabetic retinopathy, and its inhibition prevents leukostasis and protects vessels and neurons from apoptosis. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1336–1344. [Google Scholar] [CrossRef] [PubMed]
- Vinores, S.A.; Xiao, W.H.; Shen, J.; Campochiaro, P.A. TNF-α is critical for ischemia-induced leukostasis, but not retinal neovascularization nor VEGF-induced leakage. J. Neuroimmunol. 2007, 182, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.X.; Grimaldo, S.; Qi, J.W.; Yang, G.L.; Qin, T.T.; Xiao, H.Y.; Xiang, R.; Xiao, Z.; Li, L.Y.; Zhang, Z.S. Death receptor 3 mediates TNFSF15- and TNFalpha-induced endothelial cell apoptosis. Int. J. Biochem. Cell Biol. 2014, 55c, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Qaum, T.; Adamis, A.P. Sensitive blood-retinal barrier breakdown quantitation using Evans blue. Investig. Ophthalmol. Vis. Sci. 2001, 42, 789–794. [Google Scholar]
- Klaassen, I.; Van Noorden, C.J.; Schlingemann, R.O. Molecular basis of the inner blood-retinal barrier and its breakdown in diabetic macular edema and other pathological conditions. Prog. Retin. Eye Res. 2013, 34, 19–48. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | PDR (n = 35) | Control (n = 30) | p Value |
|---|---|---|---|
| Age (years) | 59.8 ± 9.15 | 62.60 ± 9.46 | 0.230 |
| Gender/male (%) | 16 (45.7) | 14 (46.7) | 0.939 |
| Duration of diabetes (years) | 14.74 ± 8.07 | - | - |
| HbA1c (%) | 8.06 ± 1.76 | 4.60 ± 0.69 | 0.000 |
| Glucose (mmol/L) | 7.29 ± 1.76 | 4.97 ± 0.94 | 0.000 |
| Weight (g) | Glycaemia (mg/dL) | N | |
|---|---|---|---|
| Control | 293.9 ± 12.9 | 82.08 ± 6.3 | 15 |
| DM | 249.9 ± 13.4 | 456.48 ± 57.6 | 15 |
| DM+vehicle | 250.4 ± 12.8 | 461.34 ± 52.74 | 30 |
| DM+TNFSF15 | 251.8 ± 12.9 | 461.52 ± 55.62 | 30 |
| F value | 75.583 | 376.721 | - |
| p value | 0.000 | 0.000 | - |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, F.; Chen, Q.; Huang, L.; Wang, Y.; Zhang, Z.; Meng, X.; Liu, Y.; Mao, C.; Zheng, F.; Zhang, J.; et al. TNFSF15 Inhibits Blood Retinal Barrier Breakdown Induced by Diabetes. Int. J. Mol. Sci. 2016, 17, 615. https://doi.org/10.3390/ijms17050615
Jiang F, Chen Q, Huang L, Wang Y, Zhang Z, Meng X, Liu Y, Mao C, Zheng F, Zhang J, et al. TNFSF15 Inhibits Blood Retinal Barrier Breakdown Induced by Diabetes. International Journal of Molecular Sciences. 2016; 17(5):615. https://doi.org/10.3390/ijms17050615
Chicago/Turabian StyleJiang, Feng, Qingzhong Chen, Liming Huang, Ying Wang, Zhuhong Zhang, Xiangda Meng, Yuanyuan Liu, Chunjie Mao, Fang Zheng, Jingkai Zhang, and et al. 2016. "TNFSF15 Inhibits Blood Retinal Barrier Breakdown Induced by Diabetes" International Journal of Molecular Sciences 17, no. 5: 615. https://doi.org/10.3390/ijms17050615