
Tetramethylpyrazine Protects Retinal Capillary Endothelial Cells (TR-iBRB2) against IL-1β-Induced Nitrative/Oxidative Stress

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
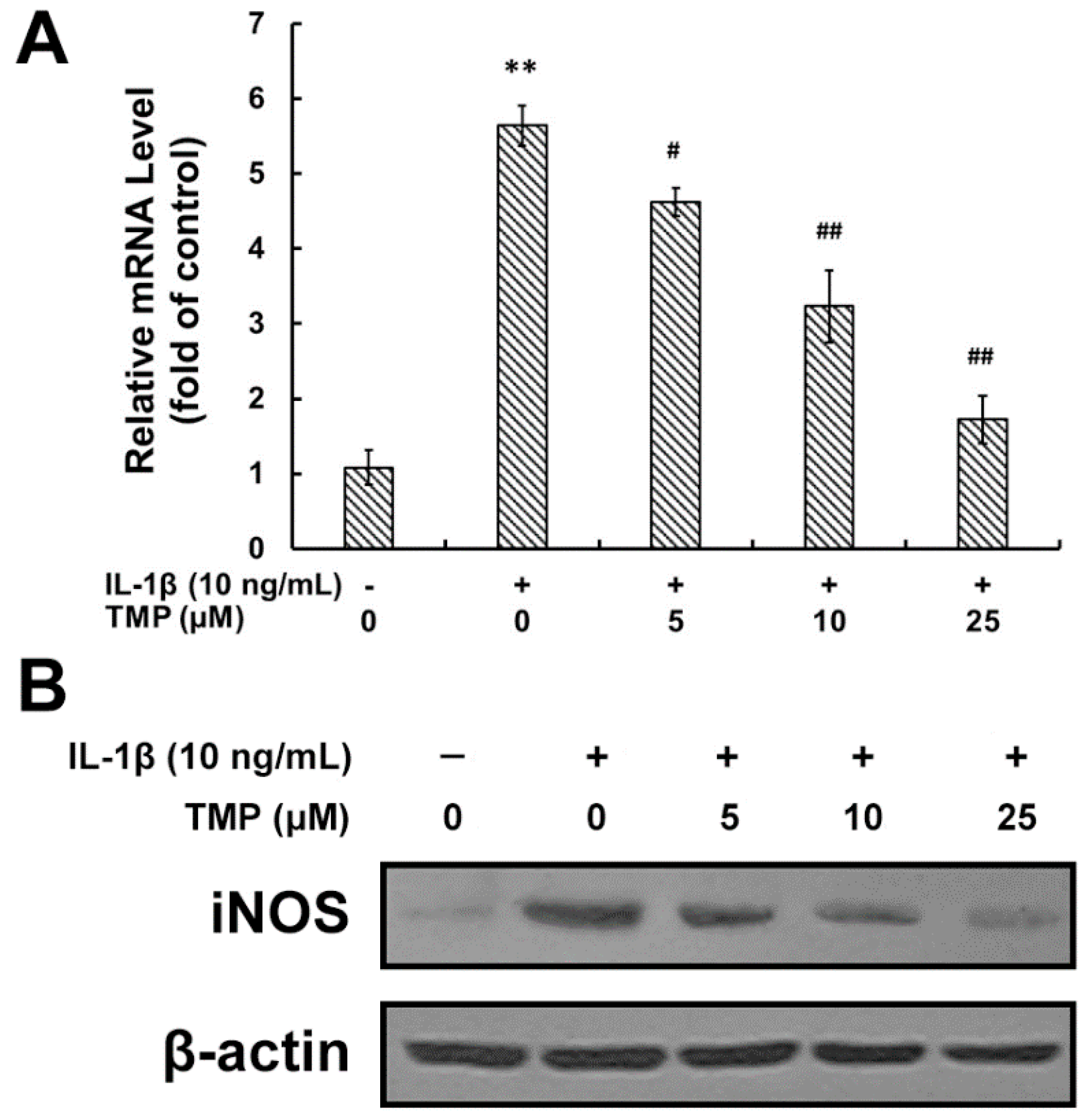
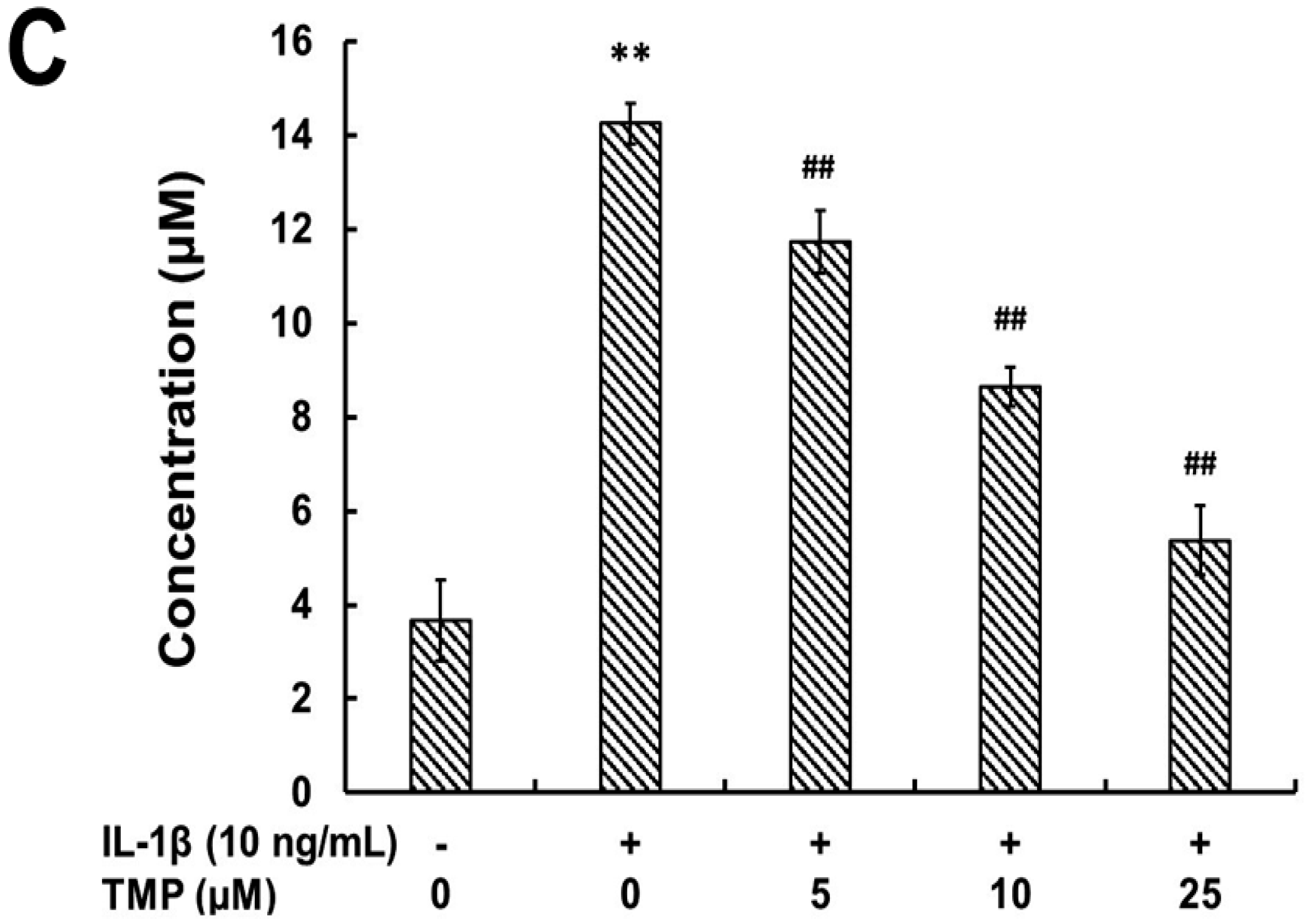
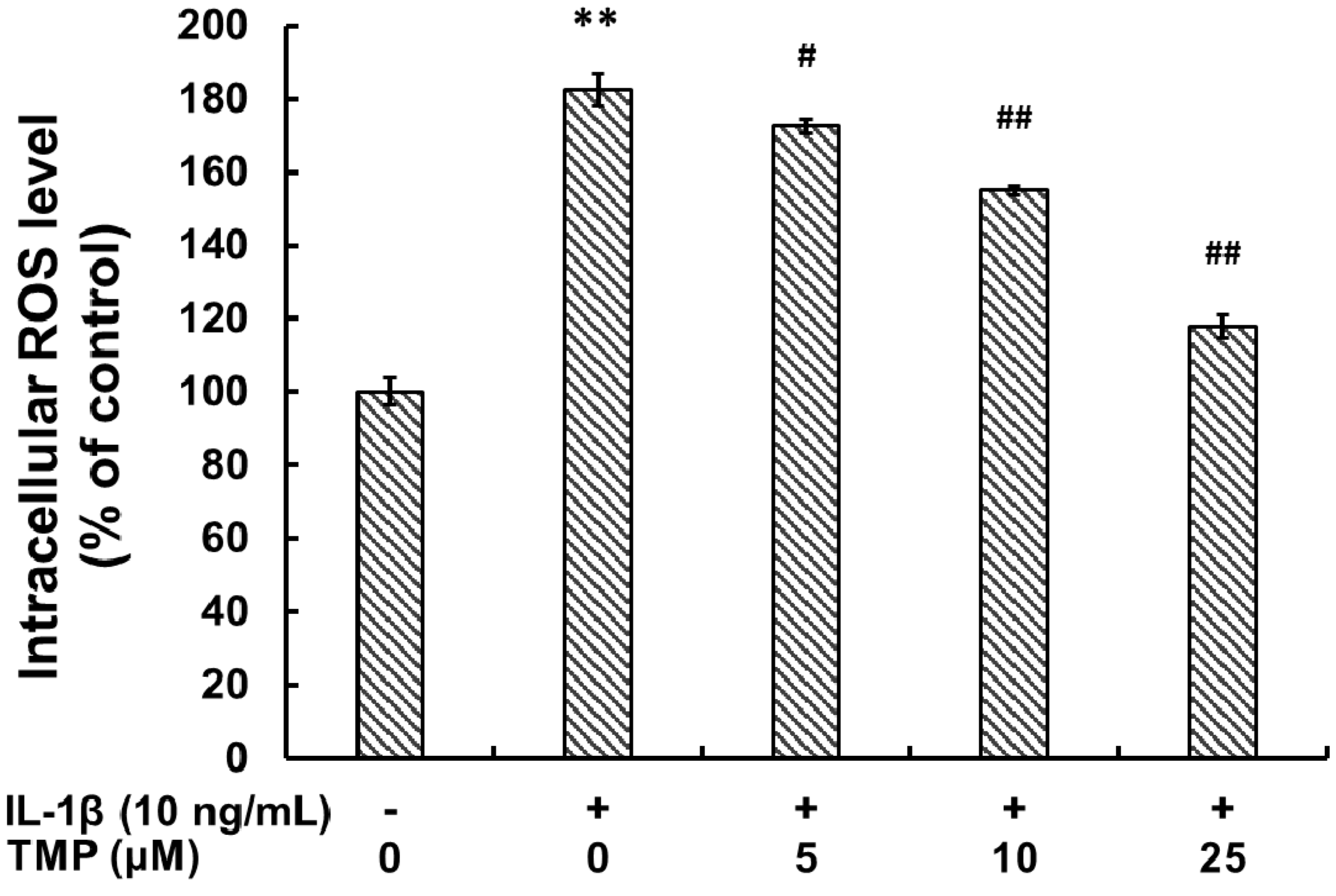
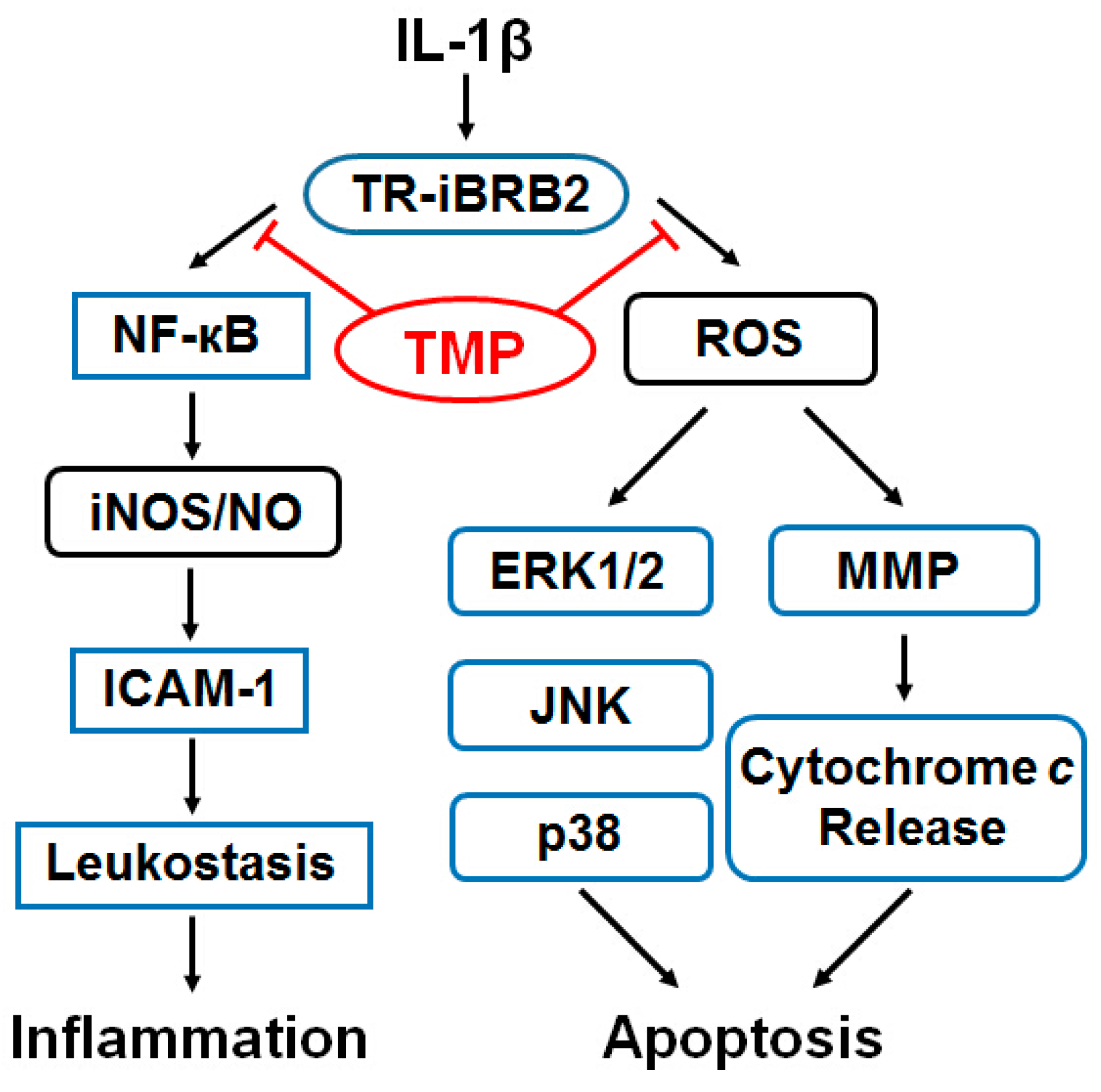
2.1.1. Tetramethylpyrazine (TMP) Reduced IL-1β-Induced Inducible Nitric Oxide Synthase (iNOS) Expression and Reactive Oxygen Species (ROS) Generation
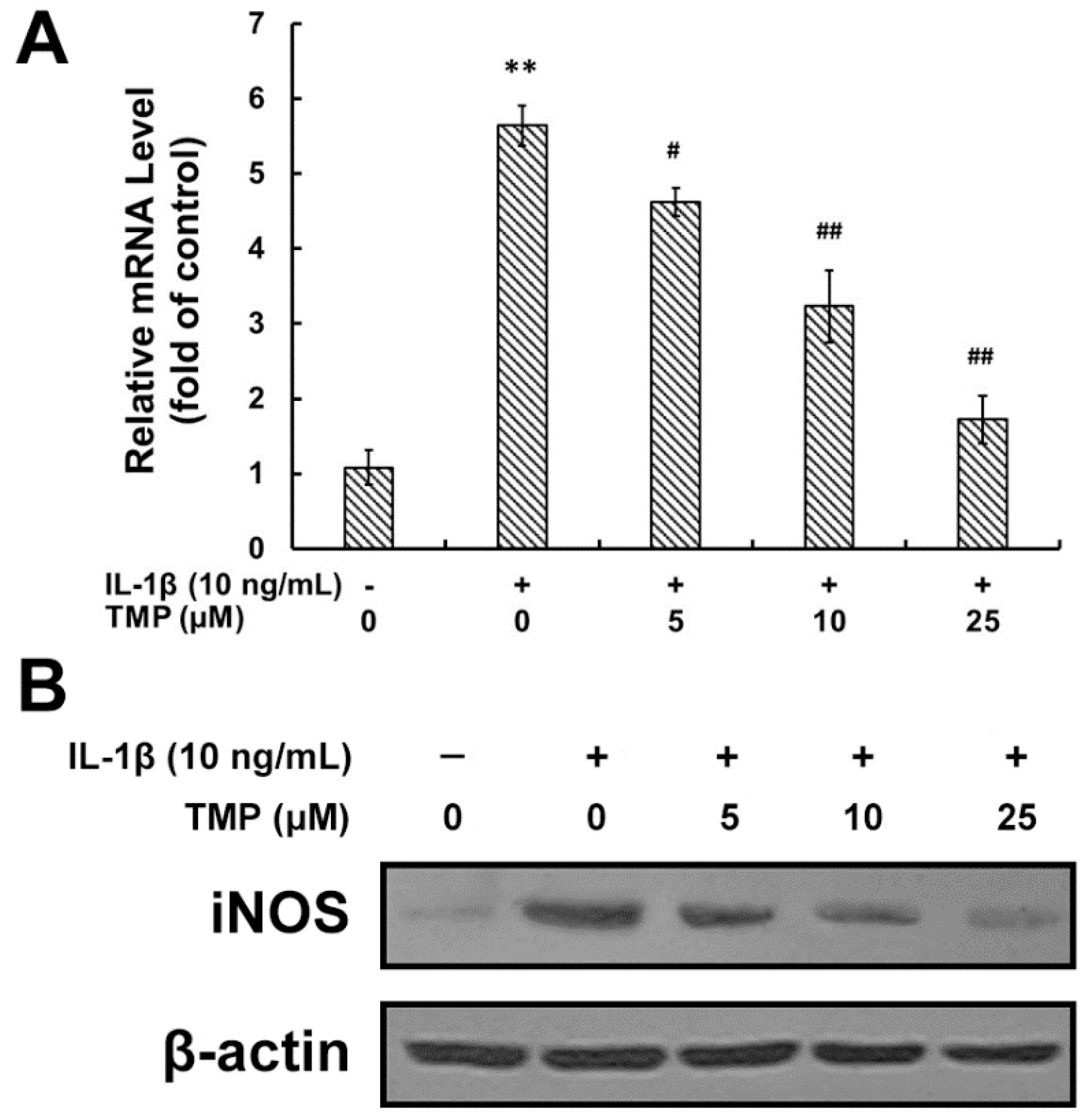
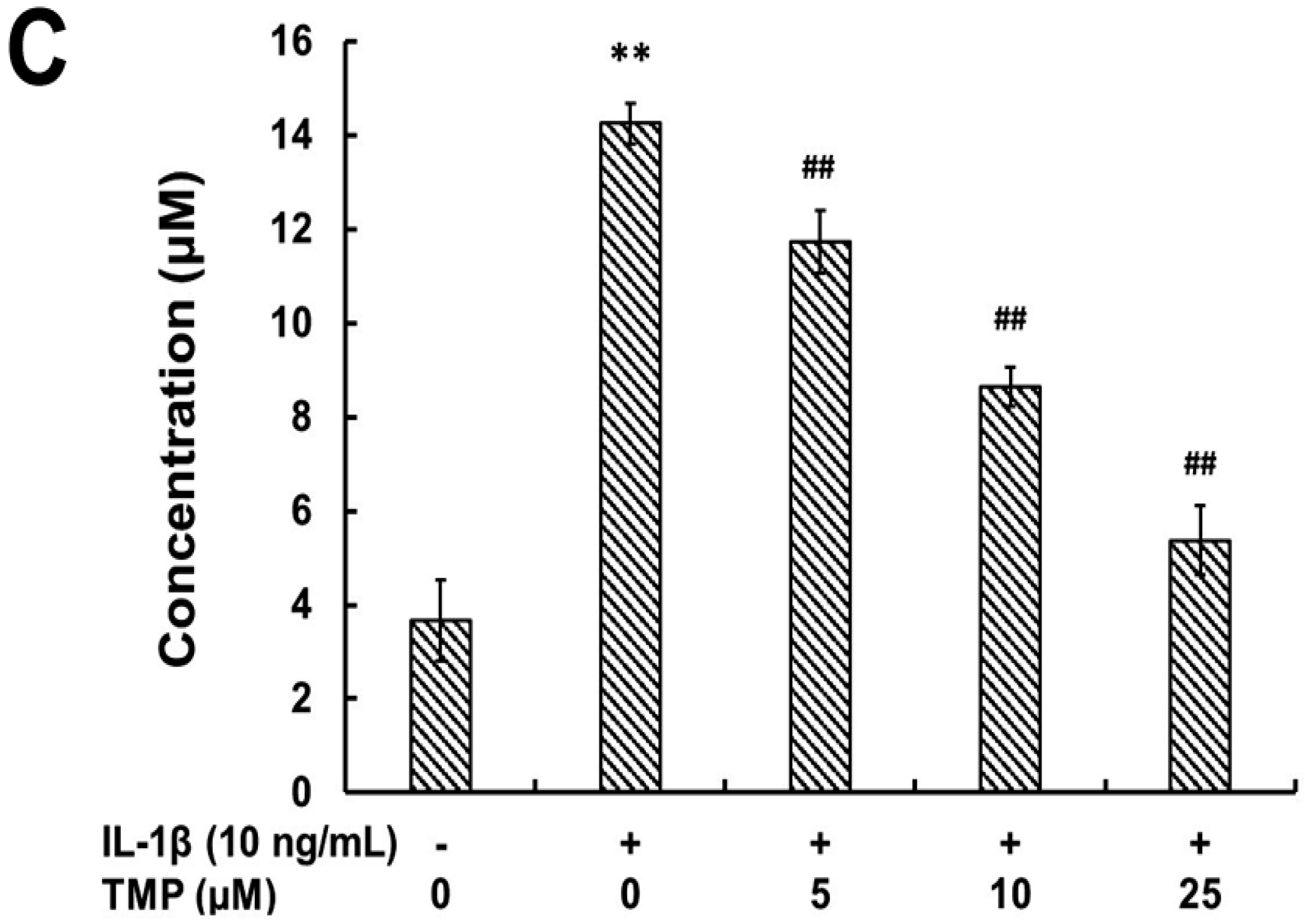
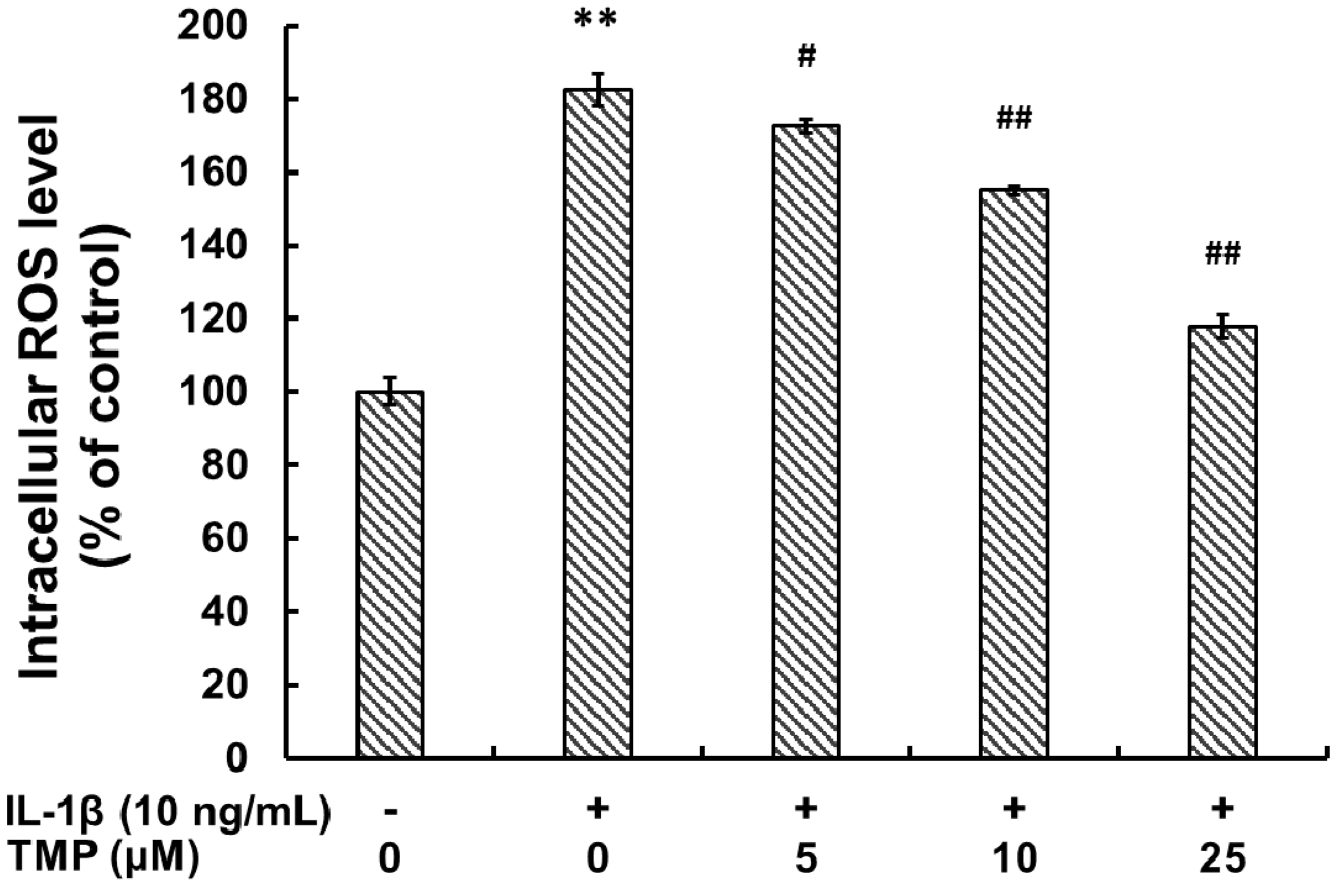
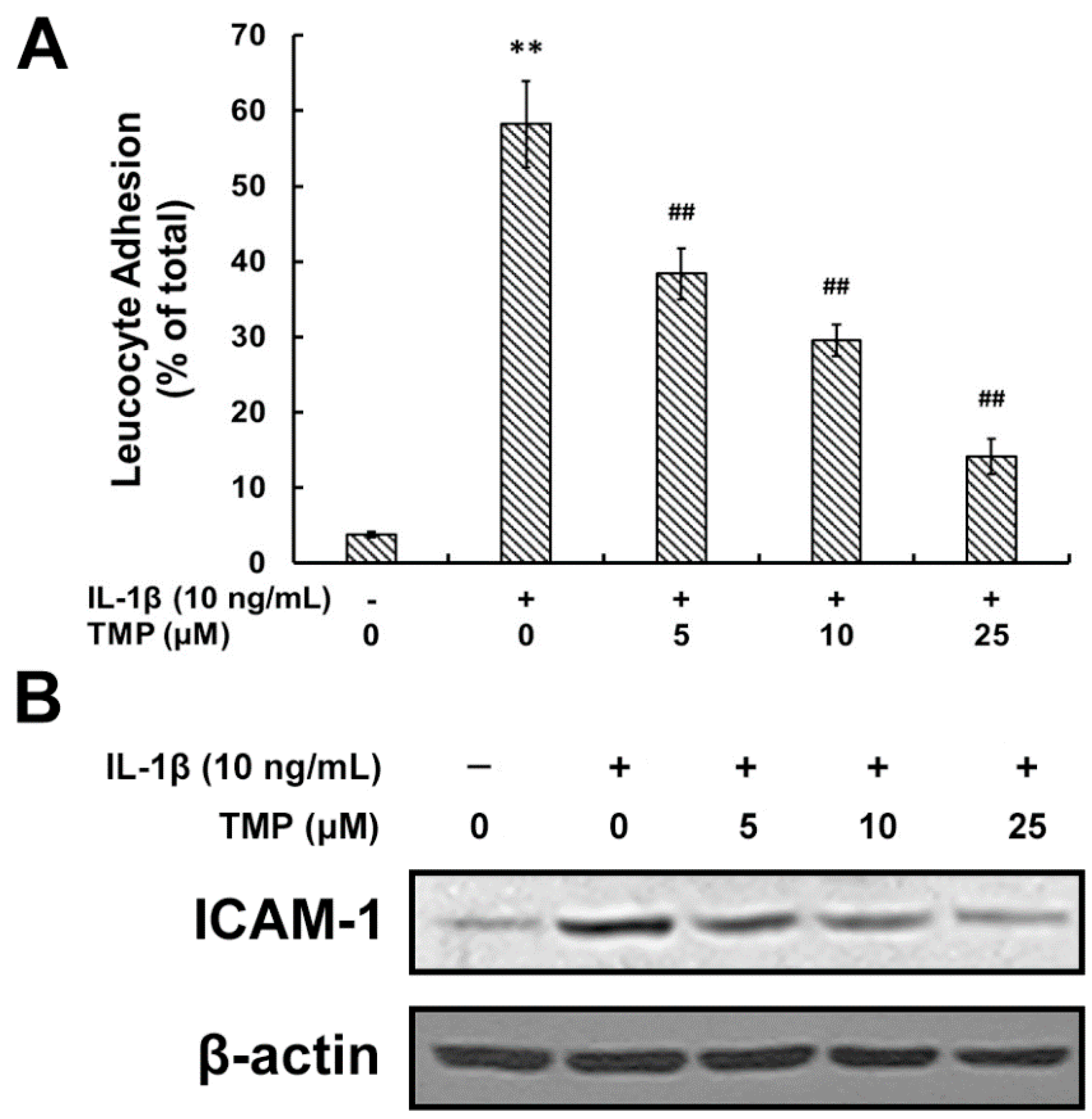
2.1.2. TMP Attenuated IL-1β-Induced Leukocyte Adhesion to TR-iBRB2 Cells and ICAM-1 Expression

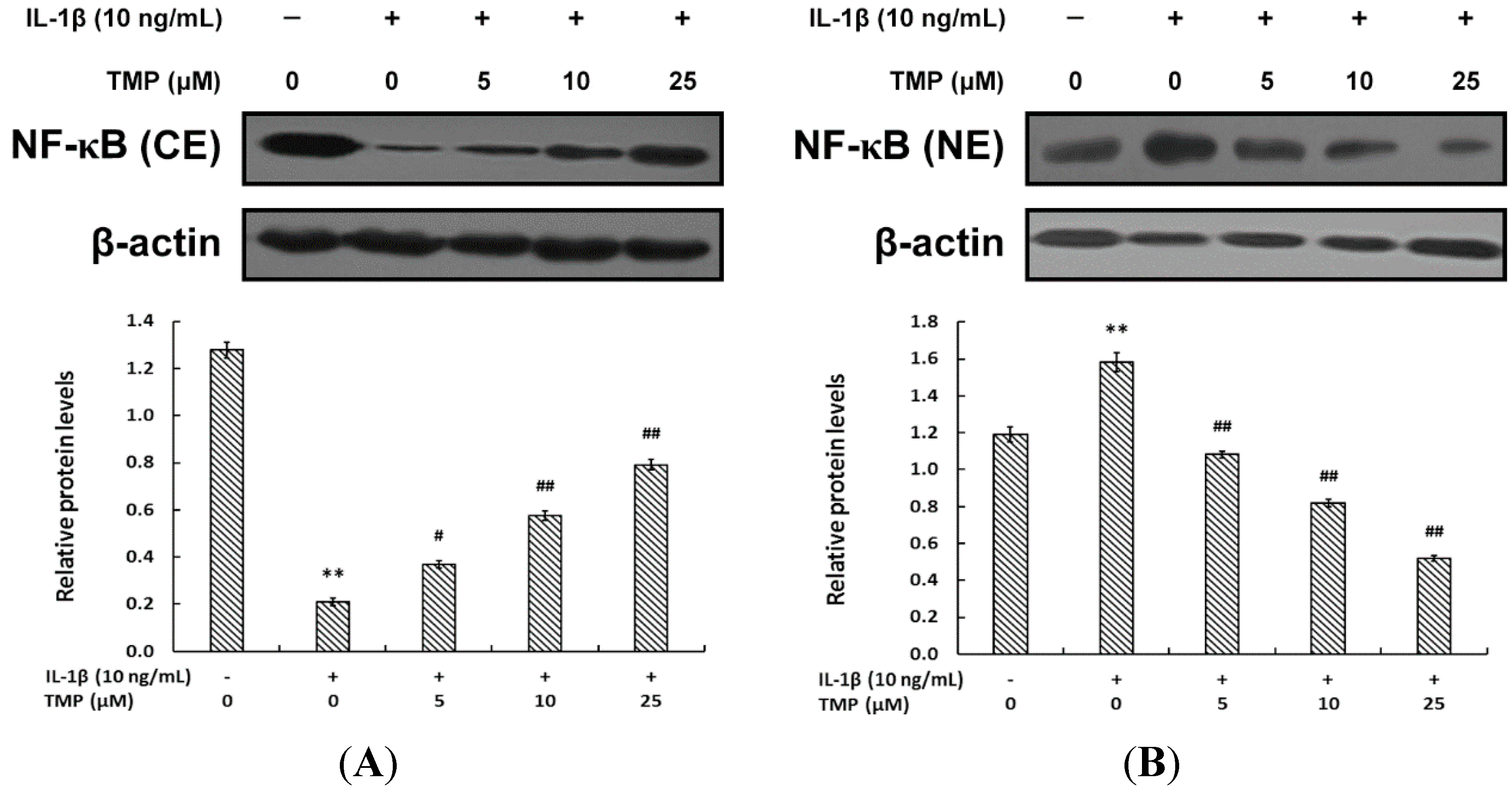
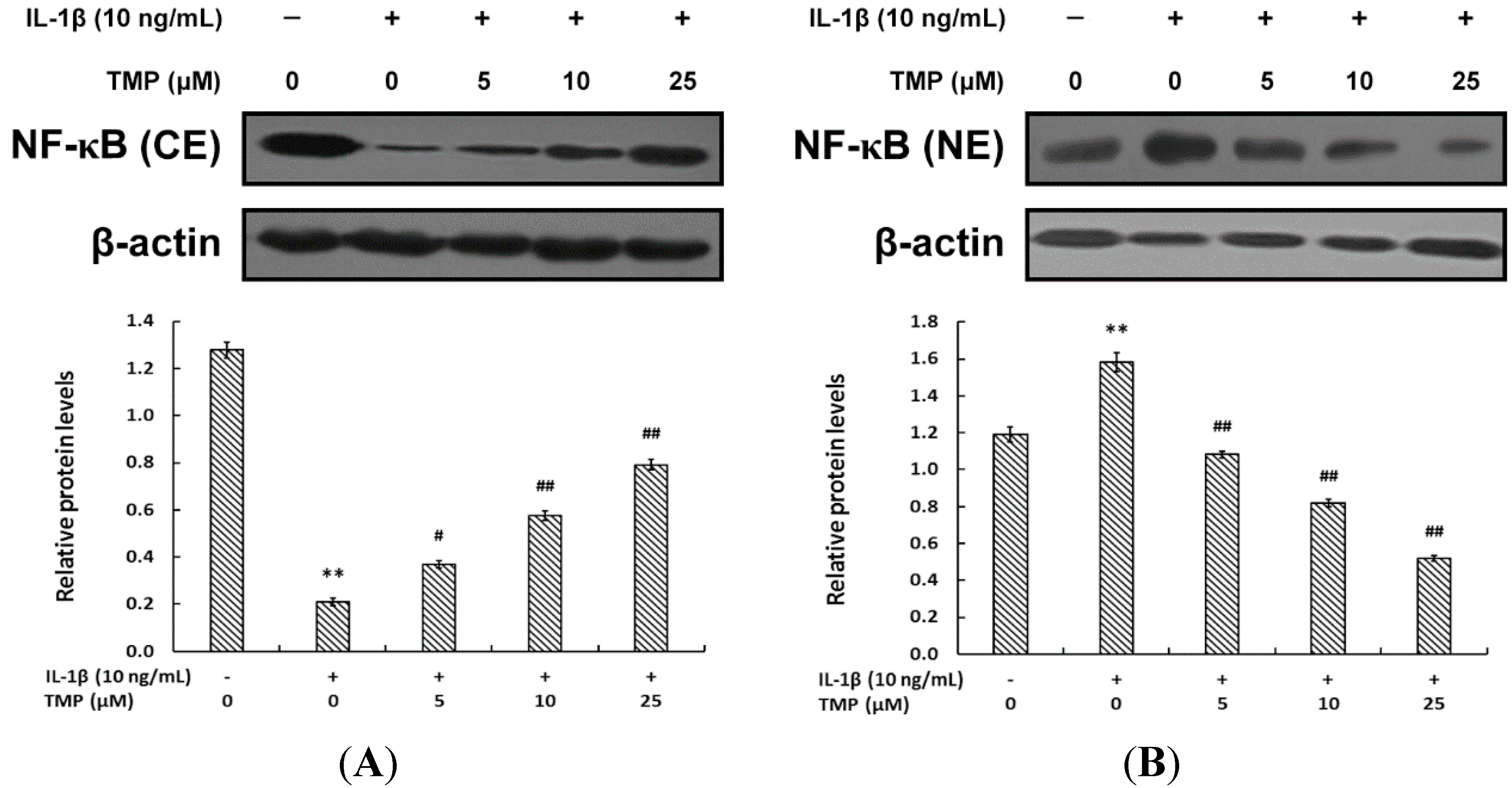
2.1.3. TMP Blocked IL-1β-Induced NF-κB Translocation into the Nucleus in TR-iBRB2 Cells
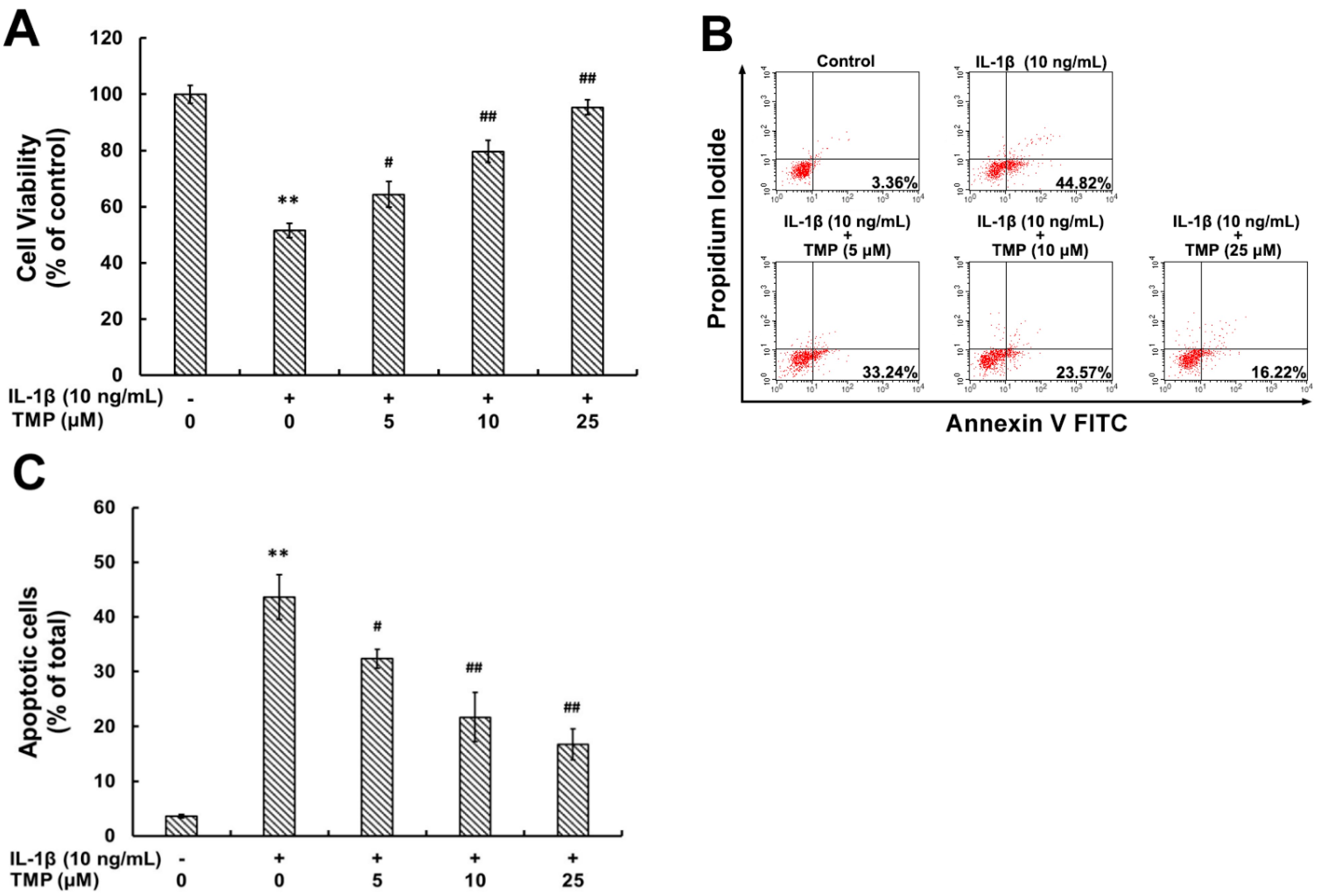
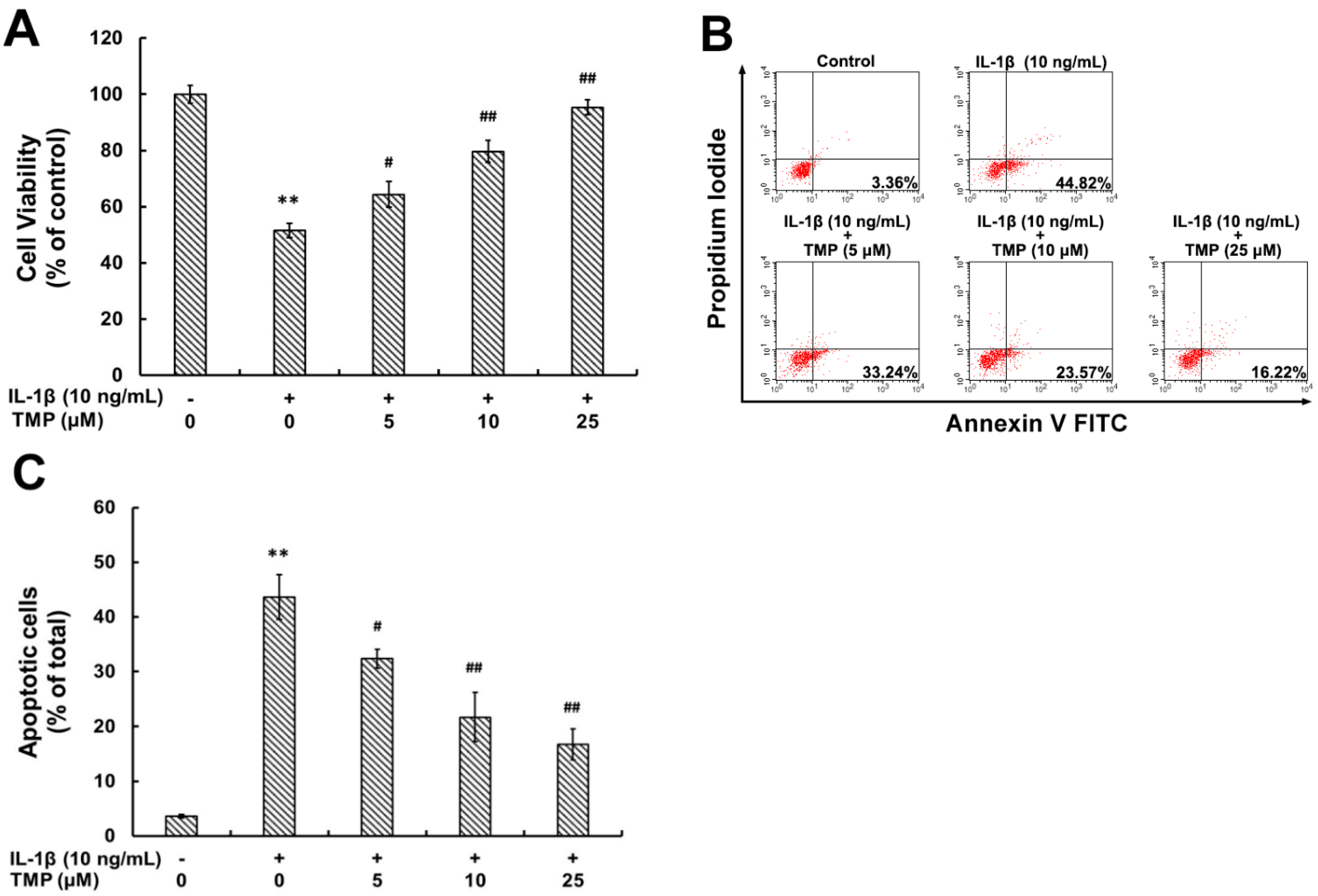
2.1.4. TMP Attenuated IL-1β-Induced Cellular Injury and Apoptosis in TR-iBRB2 Cells
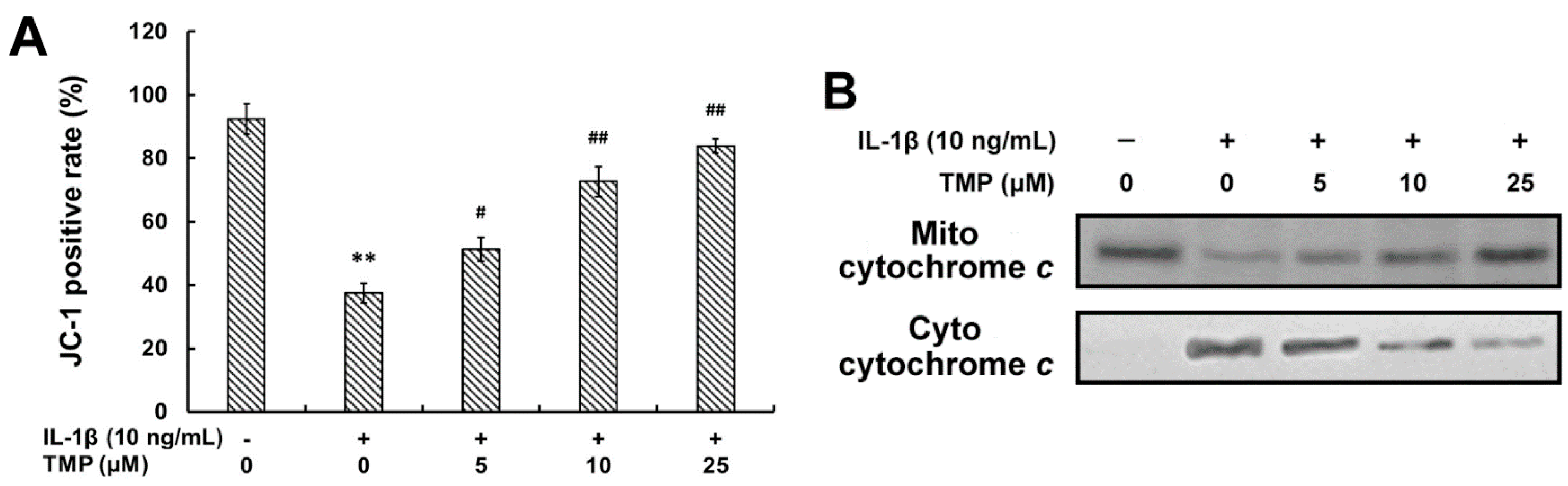
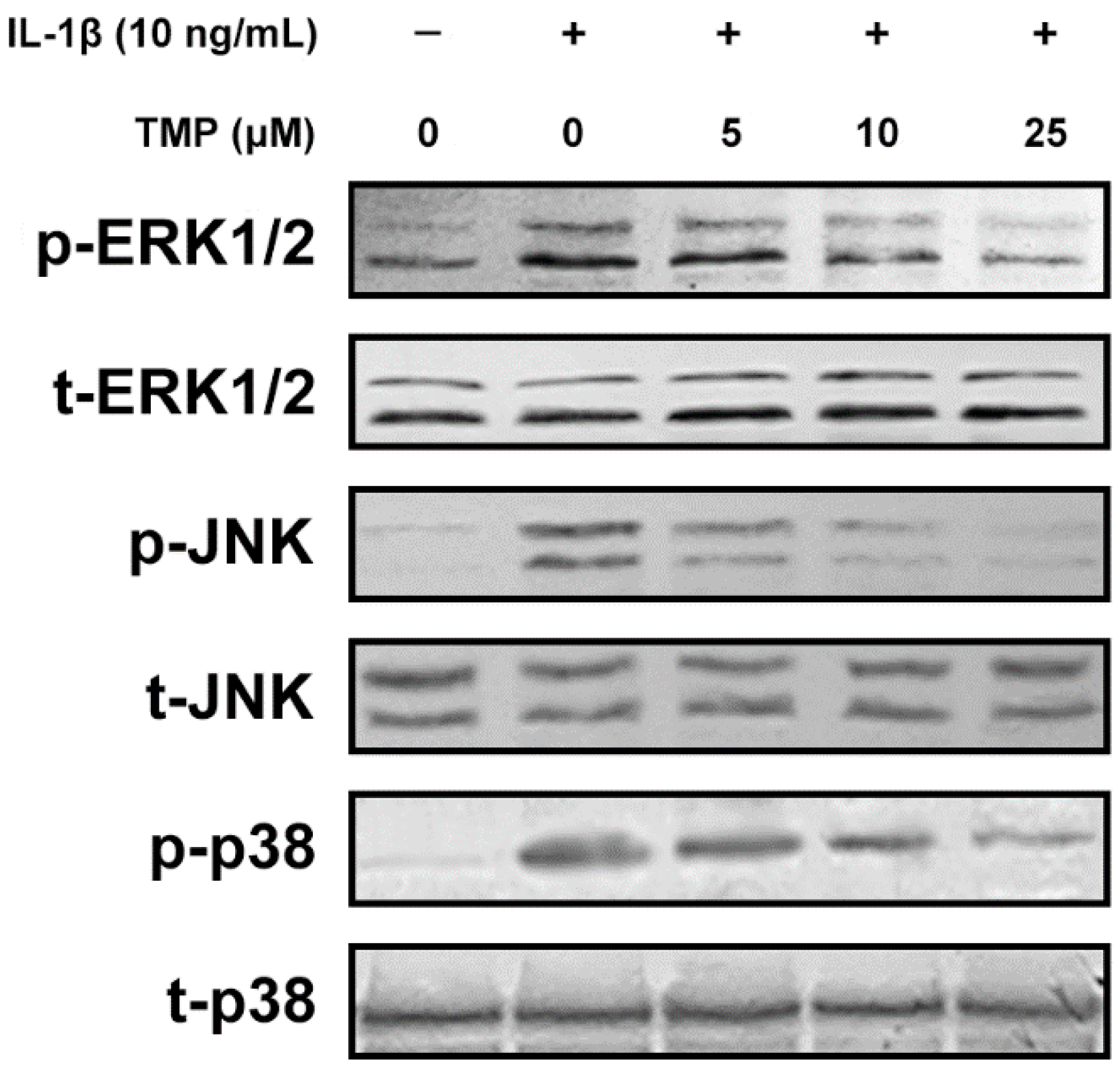
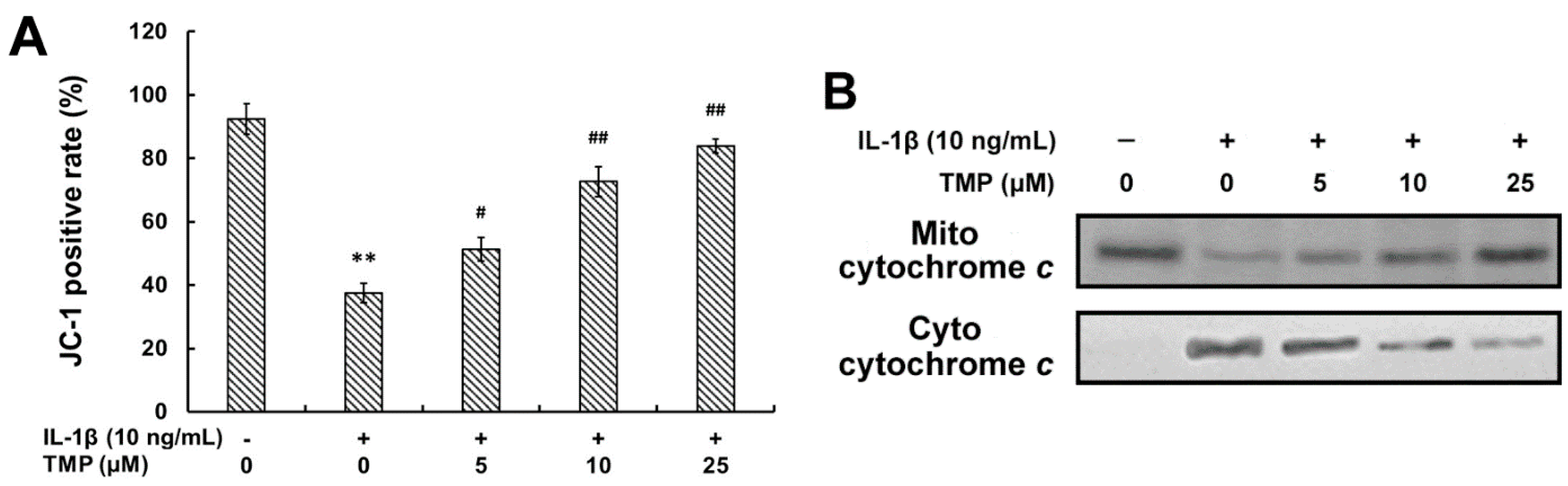
2.1.5. TMP Decreased IL-1β-Induced Mitochondrial Dysfunction and Phosphorylation of MAPKs in TR-iBRB2 Cells

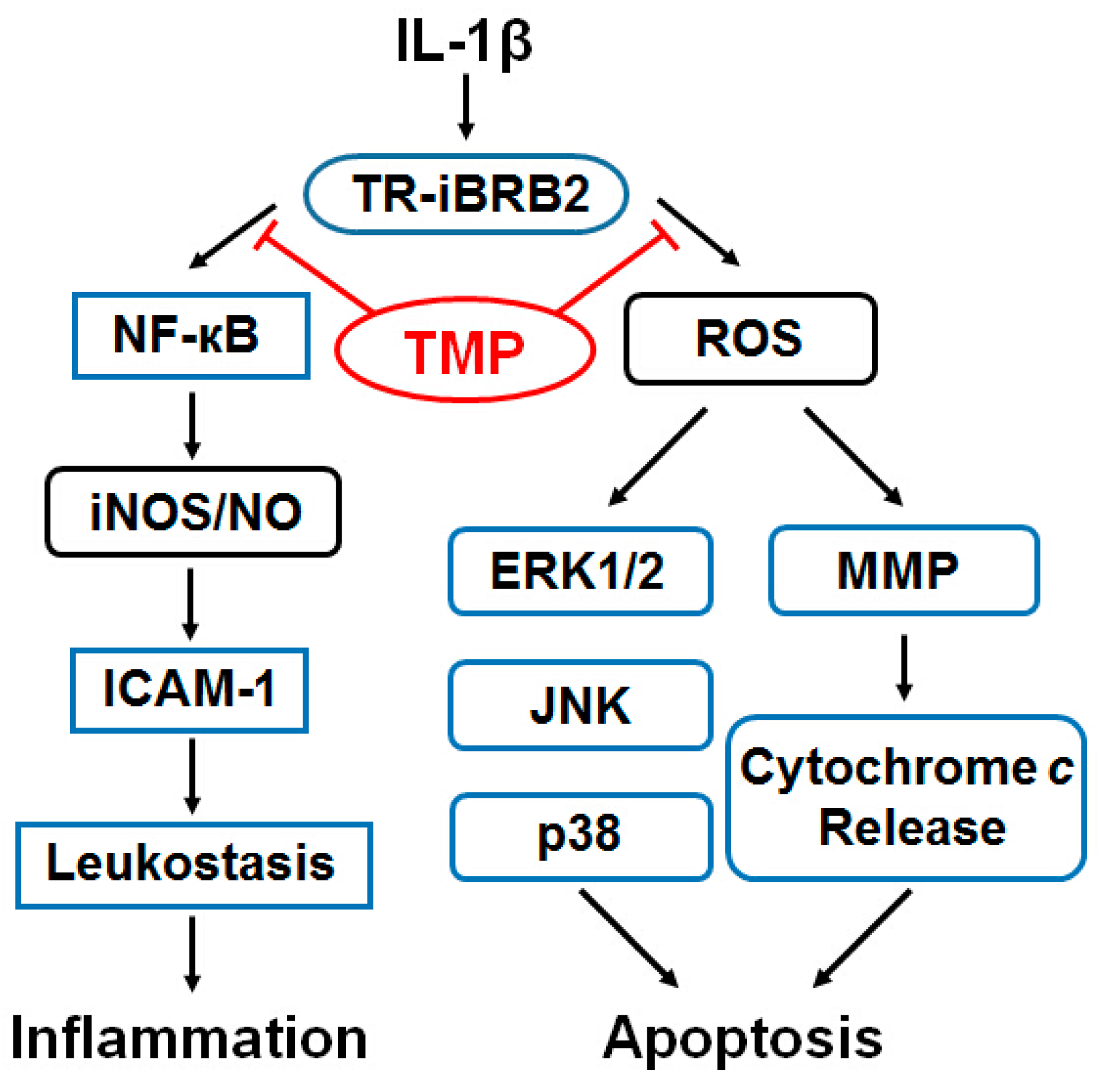
2.2. Discussion
3. Experimental Section
3.1. Materials
3.2. Cell Culture
3.3. Real-Time PCR Analysis
3.4. Western Blot Analysis
3.5. NO Measurement
3.6. Measurement of ROS
3.7. Leukocyte-Endothelial Cell Adhesion Assay
3.8. Cell Viability Assay
3.9. Cell Apoptosis Assay
3.10. Measurement of MMP
3.11. Measurement of Cytochrome c Release
3.12. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cunha-Vaz, J.; de Abreu, J.R.F.; Campos, A.J. Early breakdown of the blood-retinal barrier in diabetes. Br. J. Ophthalmol. 1975, 59, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Hafezi-Moghadam, A. Mechanisms of blood-retinal barrier breakdown in diabetic retinopathy. In Visual Dysfunction in Diabetes, 2nd ed.; Tombran-Tink, J., Barnstable, C.J., et al., Eds.; Springer: New York, NY, USA, 2012; pp. 105–122. [Google Scholar]
- Klaassen, I.; van Noorden, C.J.F.; Schlingemann, R.O. Molecular basis of the inner blood-retinal barrier and its breakdown in diabetic macular edema and other pathological conditions. Prog. Retin. Eye Res. 2013, 34, 19–48. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Gandhi, J.K.; Zhong, X.; Wei, Y.; Gong, J.; Duh, E.J.; Vinores, S.A. TNFα is required for late BRB breakdown in diabetic retinopathy, and its inhibition prevents leukostasis and protects vessels and neurons from apoptosis. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1336–1344. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; Odenbach, S. Role of interleukin-1β in the pathogenesis of diabetic retinopathy. Br. J. Ophthalmol. 2004, 88, 1343–1347. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Xiong, X.; Fang, Z.; Lu, H.; Wang, Z. Protective effect of tetramethylpyrazine on myocardial ischemia-reperfusion injury. Evid. Based Complement. Altern. Med. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Kao, T.K.; Chang, C.Y.; Ou, Y.C.; Chen, W.Y.; Kuan, Y.H.; Pan, H.C.; Liao, S.L.; Li, G.Z.; Chen, C.J. Tetramethylpyrazine reduces cellular inflammatory response following permanent focal cerebral ischemia in rats. Exp. Neurol. 2013, 247, 188–201. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.J.; Chen, K. Ischemic stroke treated with Ligusticum chuanxiong. Chin. Med. J. 1992, 105, 870–873. [Google Scholar] [PubMed]
- Gong, X.; Ivanov, V.N.; Davidson, M.M.; Hei, T.K. Tetramethylpyrazine (TMP) protects against sodium arsenite-induced nephrotoxicity by suppressing ROS production, mitochondrial dysfunction, pro-inflammatory signaling pathways and programed cell death. Arch. Toxicol. 2015, 89, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhou, H.; Ding, Y.; Li, J.; Yang, C.; Luo, Y.; Li, S.; Sun, G.; Liao, X.; Min, W. TMP prevents retinal neovascularization and imparts neuroprotection in an oxygen-induced retinopathy model. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2157–2169. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, Q.; Ge, J.; Tan, Z. Protective effects of tetramethylpyrazine on rat retinal cell cultures. Neurochem. Int. 2008, 52, 1176–1187. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Dong, X.; Liu, X.Y.; Cheng, X.C.; Cheng, Y.N.; Yu, L.G.; Guo, X.L. Mechanism of tetramethylpyrazine analogue CXC195 inhibition of hydrogen peroxide-induced apoptosis in human endothelial cells. Biol. Pharm. Bull. 2010, 33, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.N.; Chen, J.M.; Luo, L.; Lin, S.C.; Li, D.; Hu, S.X. Tetramethylpyrazine protected photoreceptor cells of rats by modulating nuclear translocation of NF-κB. Acta Pharmacol. Sin. 2005, 26, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, K.I.; Tomi, M.; Ohtsuki, S.; Takanaga, H.; Ueda, M.; Yanai, N.; Obinata, M.; Terasaki, T. Conditionally immortalized retinal capillary endothelial cell lines (TR-iBRB) expressing differentiated endothelial cell functions derived from a transgenic rat. Exp. Eye Res. 2001, 72, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Kern, T.S. Role of nitric oxide, superoxide, peroxynitrite and PARP in diabetic retinopathy. Front. Biosci. 2009, 14, 3974–3987. [Google Scholar] [CrossRef]
- El-Remessy, A.B.; Behzadian, M.A.; Abou-Mohamed, G.; Franklin, T.; Caldwell, R.W.; Caldwell, R.B. Experimental diabetes causes breakdown of the blood-retina barrier by a mechanism involving tyrosine nitration and increases in expression of vascular endothelial growth factor and urokinase plasminogen activator receptor. Am. J. Pathol. 2003, 162, 1995–2004. [Google Scholar] [CrossRef]
- Joussen, A.M.; Poulaki, V.; Qin, W.; Kirchhof, B.; Mitsiades, N.; Wiegand, S.J.; Rudge, J.; Yancopoulos, G.D.; Adamis, A. P. Retinal vascular endothelial growth factor induces intercellular adhesion molecule-1 and endothelial nitric oxide synthase expression and initiates early diabetic retinal leukocyte adhesion in vivo. Am. J. Pathol. 2002, 160, 501–509. [Google Scholar] [CrossRef]
- Sinha, K.; Das, J.; Pal, P.; Sil, P. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef] [PubMed]
- Caroppi, P.; Sinibaldi, F.; Fiorucci, L.; Santucci, R. Apoptosis and human diseases: mitochondrion damage and lethal role of released cytochrome c as proapoptotic protein. Curr. Med. Chem. 2009, 16, 4058–4065. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Kim, S.; Chung, H.T.; Pae, H.O. Reactive oxygen species in the activation of MAP kinases. Methods Enzymol. 2013, 528, 27–48. [Google Scholar] [PubMed]
- Zheng, Z.; Li, Z.; Chen, S.; Pan, J.; Ma, X. Tetramethylpyrazine attenuates TNF-α-induced iNOS expression in human endothelial cells: Involvement of Syk-mediated activation of PI3K-IKK-IκB signaling pathways. Exp. Cell Res. 2013, 319, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.J.; Hao, J.; Wang, S.Q.; Jin, B.L.; Chen, X.B. Protective effects of ligustrazine on TNF-α-induced endothelial dysfunction. Eur. J. Pharmacol. 2012, 674, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Leal, E.C.; Manivannan, A.; Hosoya, K.; Terasaki, T.; Cunha-Vaz, J.; Ambrosio, A. F.; Forrester, J.V. Inducible nitric oxide synthase isoform is a key mediator of leukostasis and blood-retinal barrier breakdown in diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5257–5265. [Google Scholar] [CrossRef] [PubMed]
- Sahnoun, Z.; Jamoussi, K.; Zeghal, K.M. Free radicals and antioxidants: Physiology, human pathology and therapeutic aspects (part II). Therapie 1998, 53, 315–339. [Google Scholar] [PubMed]
- Rajan, S.; Ye, J.; Bai, S.; Huang, F.; Guo, Y.L. NF-κB, but not p38 MAP kinase, is required for TNF-α-induced expression of cell adhesion molecules in endothelial cells. J. Cell. Biochem. 2008, 105, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Leal, E.C.; Martins, J.; Voabil, P.; Liberal, J.; Chiavaroli, C.; Bauer, J.; Cunha-Vaz, J.; Ambrosio, A.F. Calcium dobesilate inhibits the alterations in tight junction proteins and leukocyte adhesion to retinal endothelial cells induced by diabetes. Diabetes 2010, 59, 2637–2645. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; Chan, P.S. Oxidative stress and diabetic retinopathy. Exp. Diabetes Res. 2007, 2007. [Google Scholar] [CrossRef] [PubMed]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Lung Cell Mol. Physiol. 2000, 279, 1005–1028. [Google Scholar]
- Davis, W.; Ronai, Z.E.; Tew, K.D. Cellular thiols and reactive oxygen species in drug-induced apoptosis. J. Pharmacol. Exp. Ther. 2001, 296, 1–6. [Google Scholar] [PubMed]
- Anuradha, C.D.; Kanno, S.; Hirano, S. Oxidative damage to mitochondria is a preliminary step to caspase-3 activation in fluoride-induced apoptosis in HL-60 cells. Free Radic. Biol. Med. 2001, 31, 367–373. [Google Scholar] [CrossRef]
- Pan, J.S.; Hong, M.Z.; Ren, J.L. Reactive oxygen species: A double-edged sword in oncogenesis. World J. Gastroenterol. 2009, 15, 1702–1707. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef] [PubMed]
- Gerhardinger, C.; Dagher, Z.; Sebastiani, P.; Park, Y.S.; Lorenzi, M. The transforming growth factor-β pathway is a common target of drugs that prevent experimental diabetic retinopathy. Diabetes 2009, 58, 1659–1667. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lu, H.F.; Sue, C.C.; Yu, C.S.; Chen, S.C.; Chen, G.W.; Chung, J. G. Diallyl disulfide (DADS) induced apoptosis undergo caspase-3 activity in human bladder cancer T24 cells. Food Chem. Toxicol. 2004, 42, 1543–1552. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, X.; Bhalla, K.; Kim, C.N.; Ibrado, A.M.; Cai, J.; Peng, T.I.; Jones, D.P.; Wang, X. Prevention of apoptosis by Bcl-2: Release of cytochrome c from mitochondria blocked. Science 1997, 275, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, X.; Wang, K.; Zhang, K.; Tan, X.; Wu, Z.; Sun, S.; Zhou, F.; Zhu, L. Tetramethylpyrazine Protects Retinal Capillary Endothelial Cells (TR-iBRB2) against IL-1β-Induced Nitrative/Oxidative Stress. Int. J. Mol. Sci. 2015, 16, 21775-21790. https://doi.org/10.3390/ijms160921775
Zhu X, Wang K, Zhang K, Tan X, Wu Z, Sun S, Zhou F, Zhu L. Tetramethylpyrazine Protects Retinal Capillary Endothelial Cells (TR-iBRB2) against IL-1β-Induced Nitrative/Oxidative Stress. International Journal of Molecular Sciences. 2015; 16(9):21775-21790. https://doi.org/10.3390/ijms160921775
Chicago/Turabian StyleZhu, Xue, Ke Wang, Kai Zhang, Xuhua Tan, Zhifeng Wu, Song Sun, Fanfan Zhou, and Ling Zhu. 2015. "Tetramethylpyrazine Protects Retinal Capillary Endothelial Cells (TR-iBRB2) against IL-1β-Induced Nitrative/Oxidative Stress" International Journal of Molecular Sciences 16, no. 9: 21775-21790. https://doi.org/10.3390/ijms160921775