
CrWSKP1, an SKP1-like Gene, Is Involved in the Self-Incompatibility Reaction of “Wuzishatangju” (Citrus reticulata Blanco)

Abstract
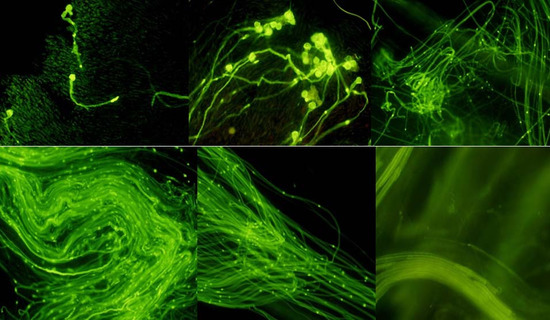
:
1. Introduction
2. Results
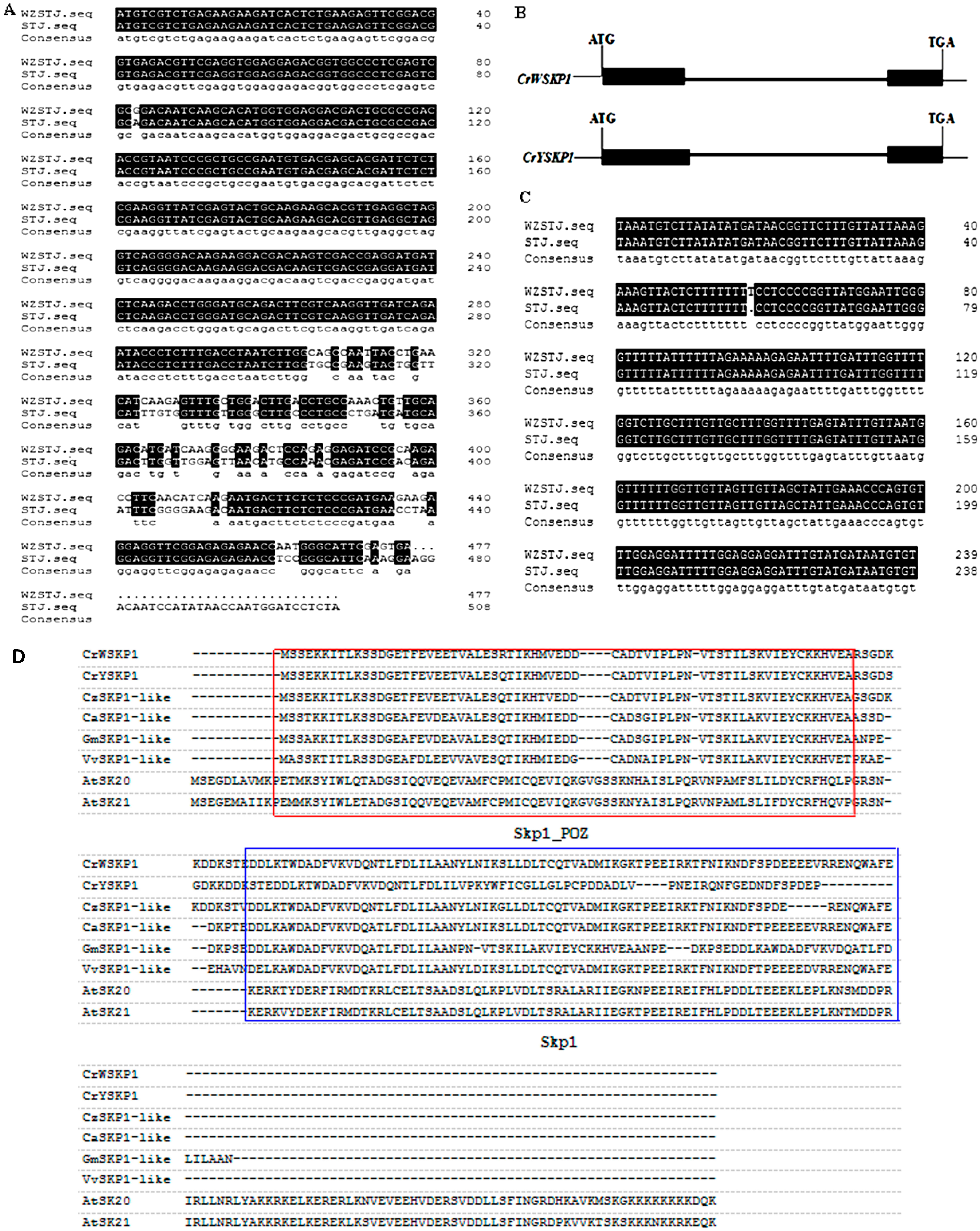
2.1. Cloning and Sequence Analysis of SKP1-like Gene
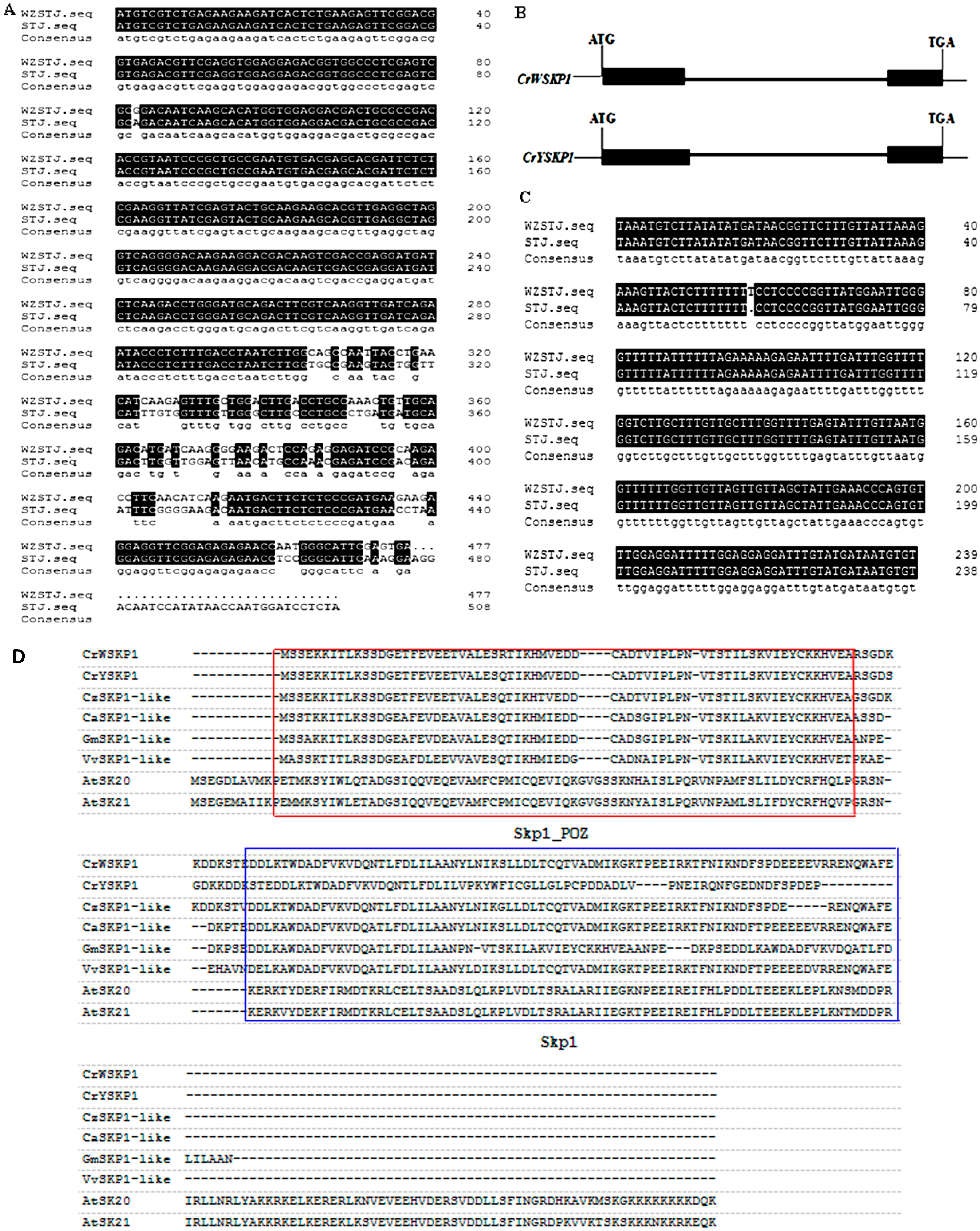
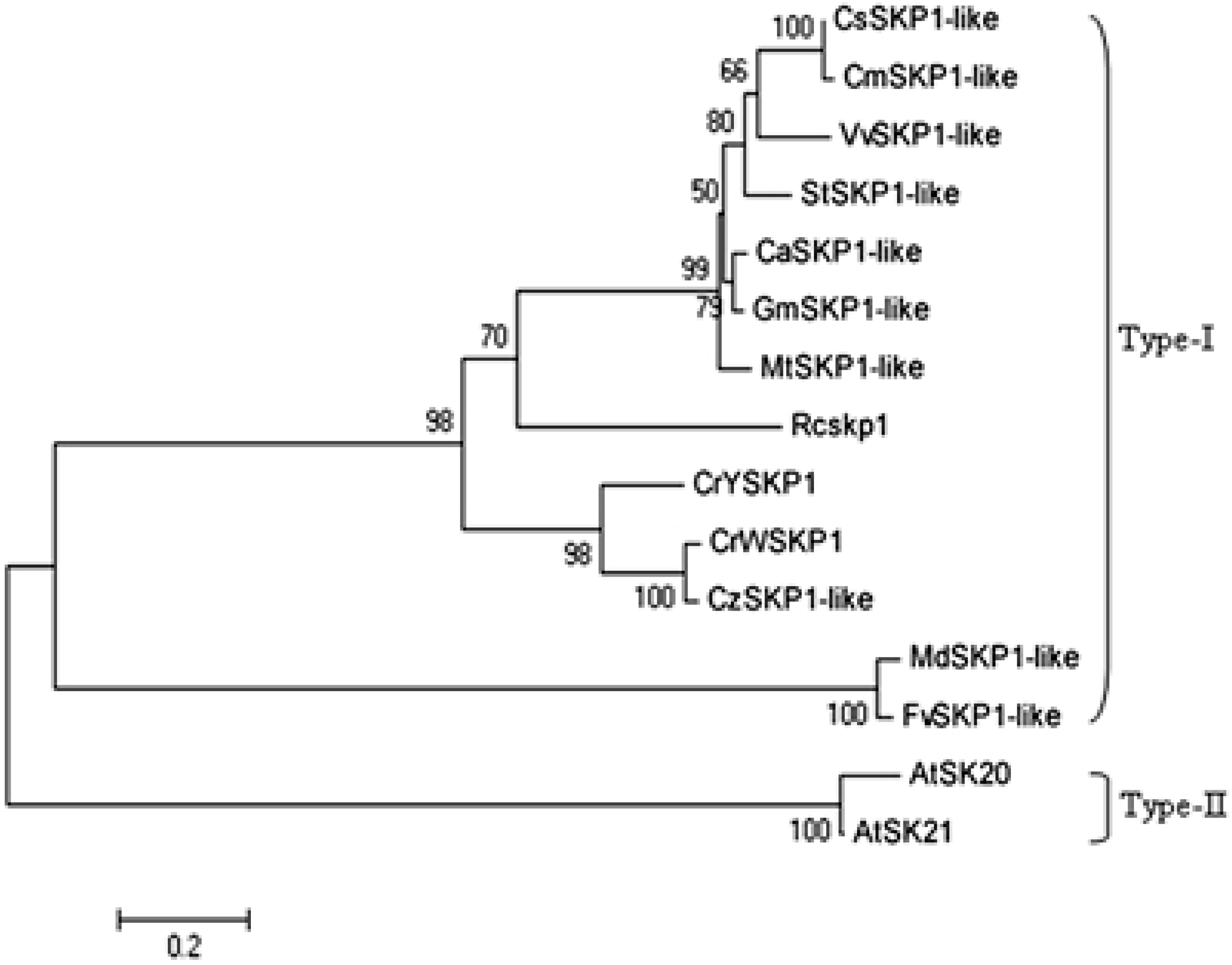
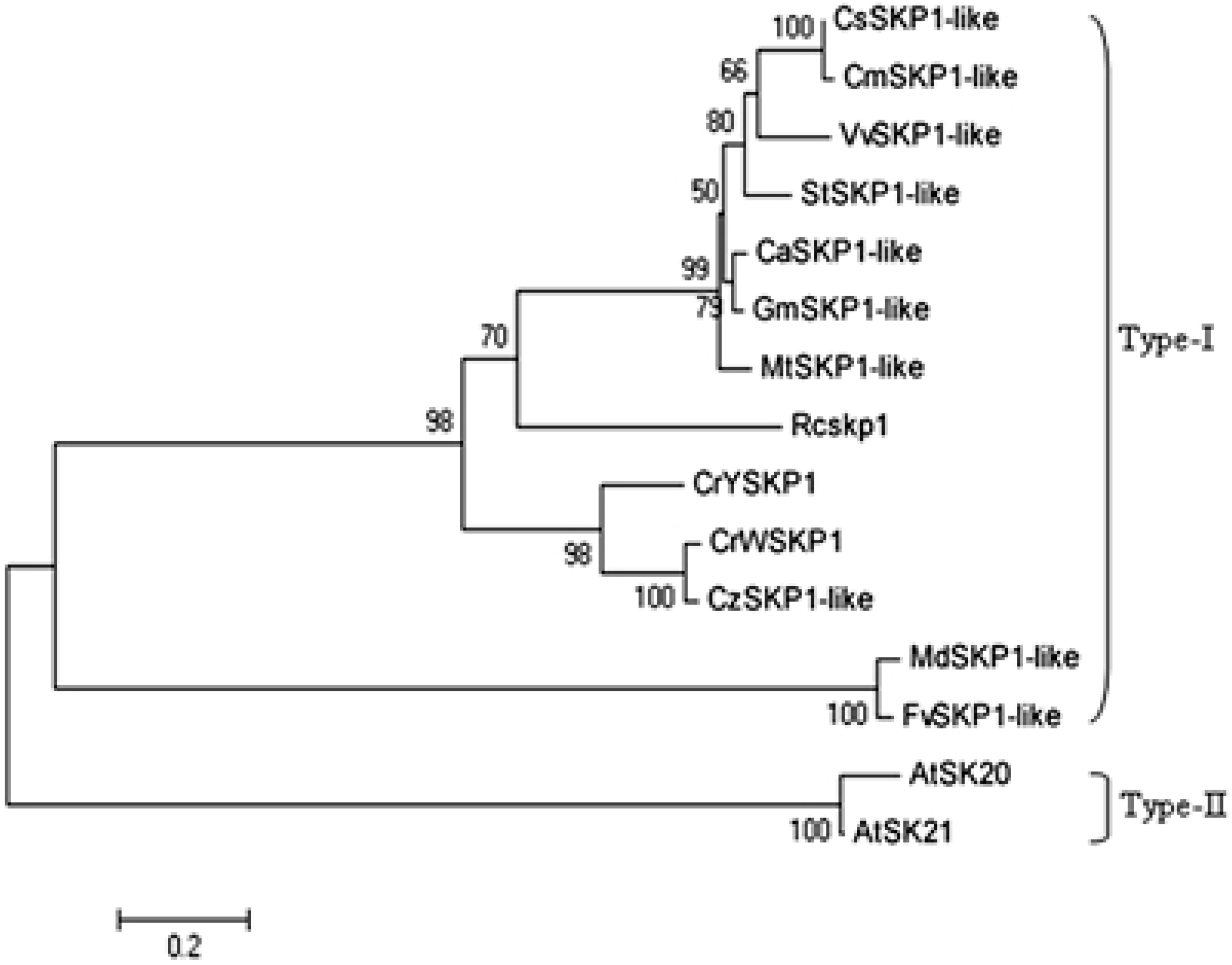
2.2. Phylogenetic Analysis of SKP1-like Genes
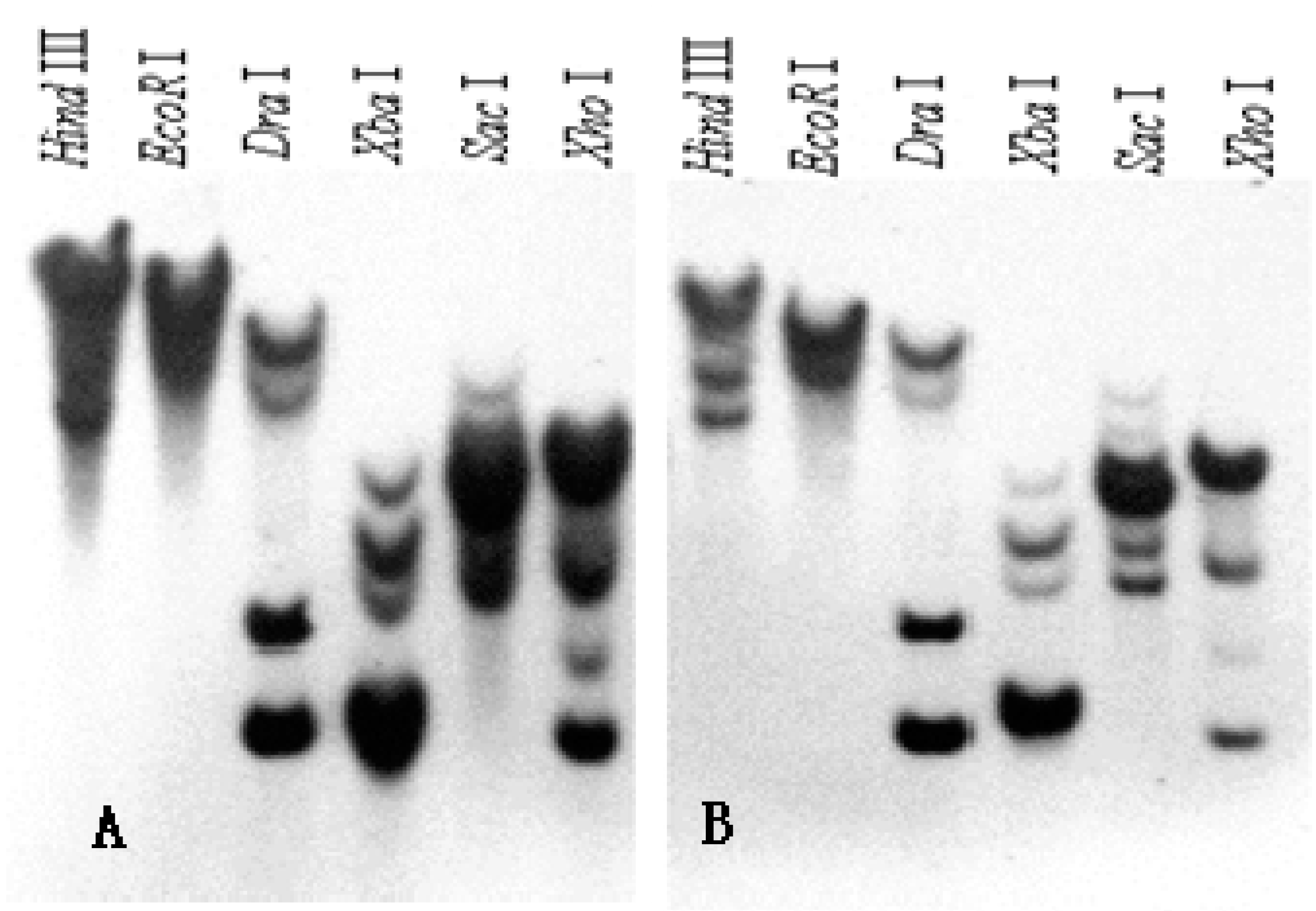
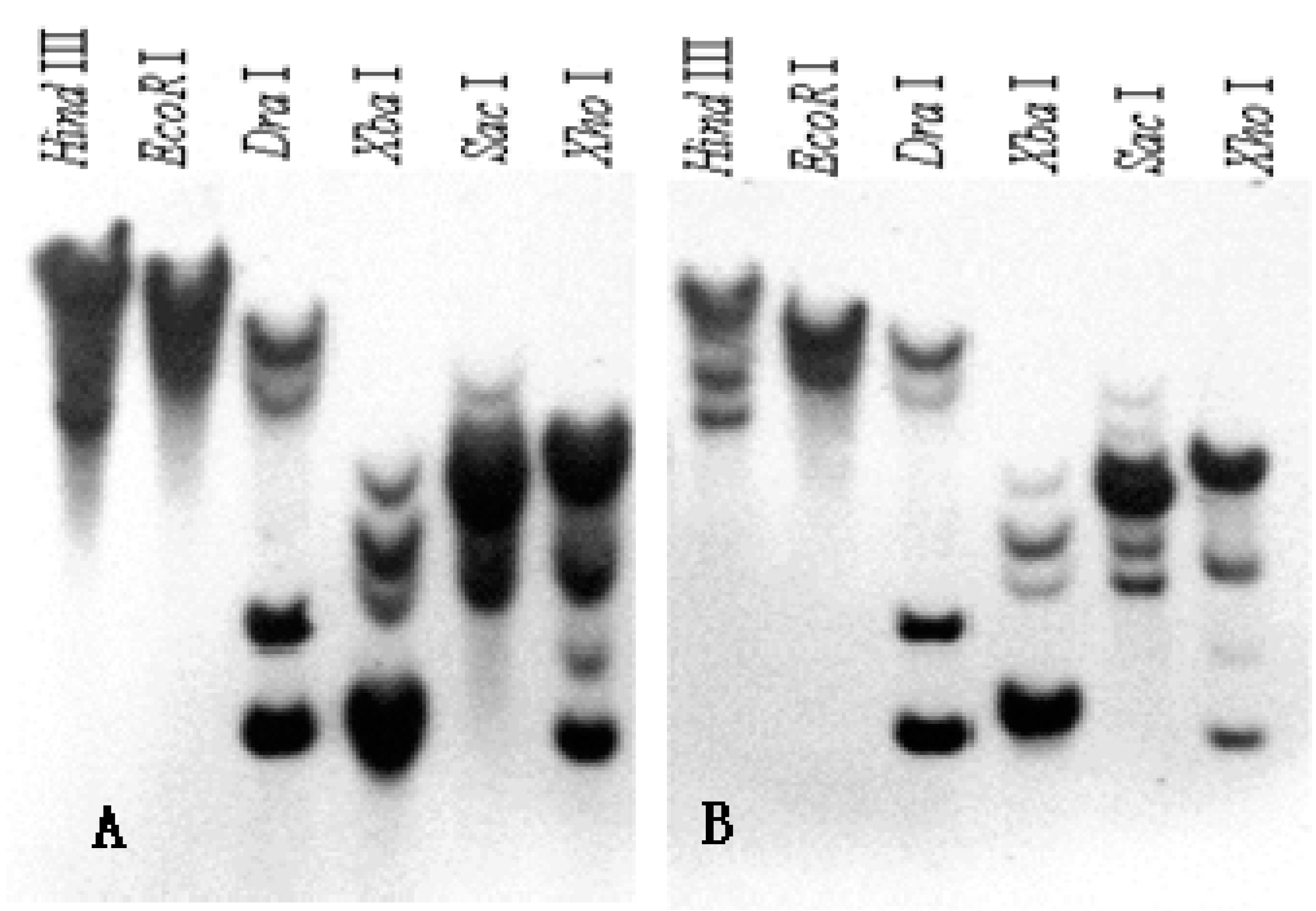
2.3. Southern Blot Analysis of SKP1-like Genes
2.4. Molecular Analyses of CrWSKP1 Transgenic Tobacco
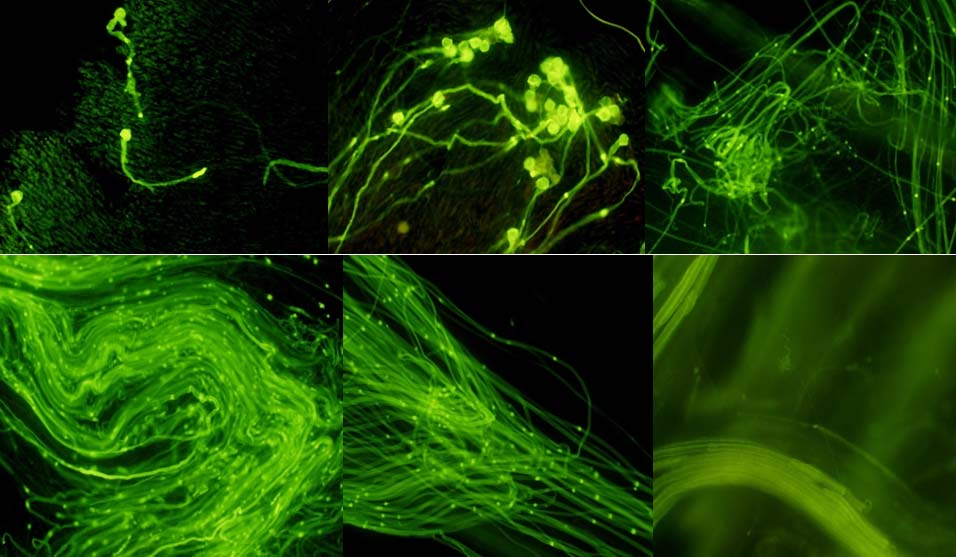
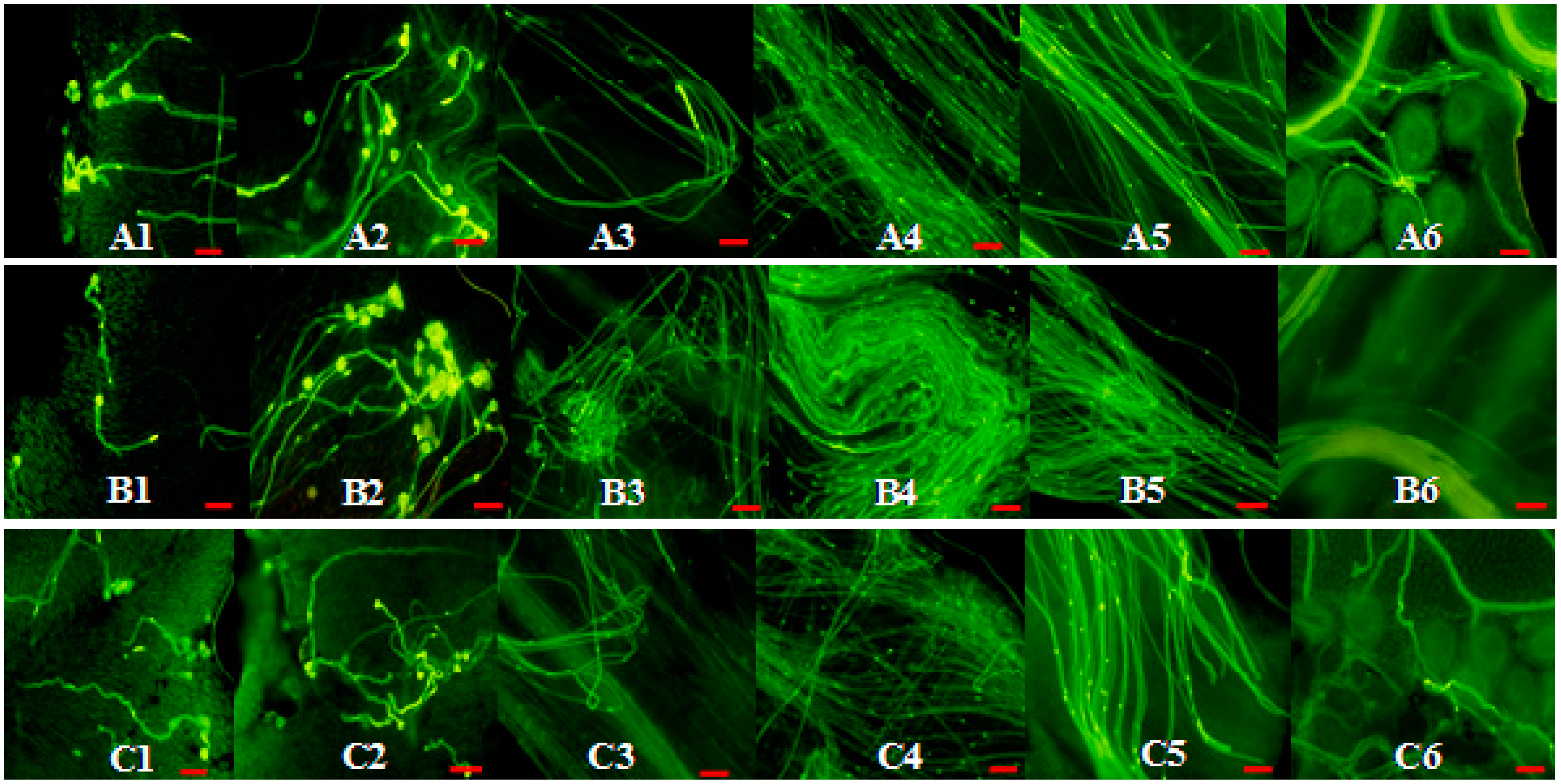
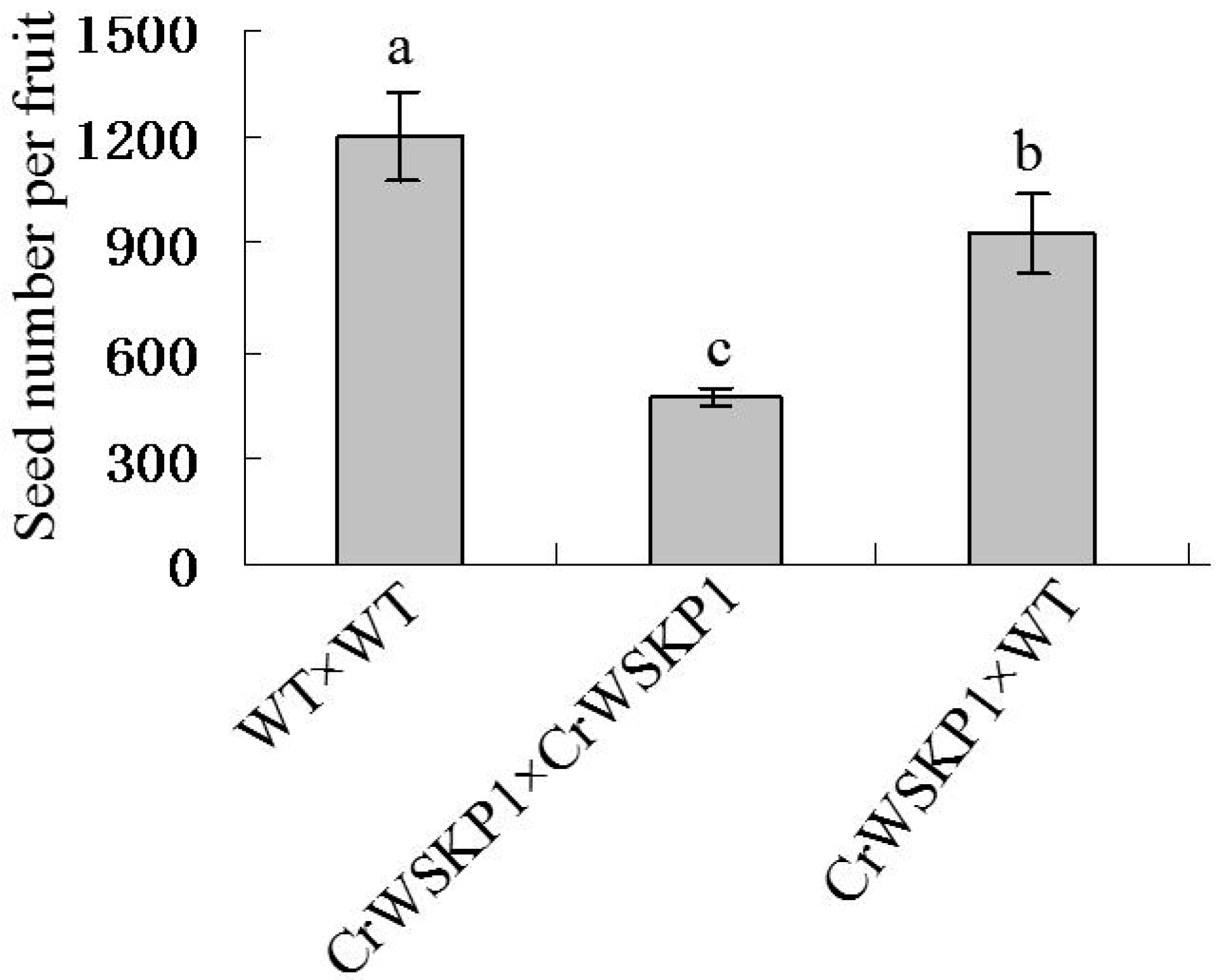
2.5. Phenotype Analyses of CrWSKP1 Transgenic Tobacco
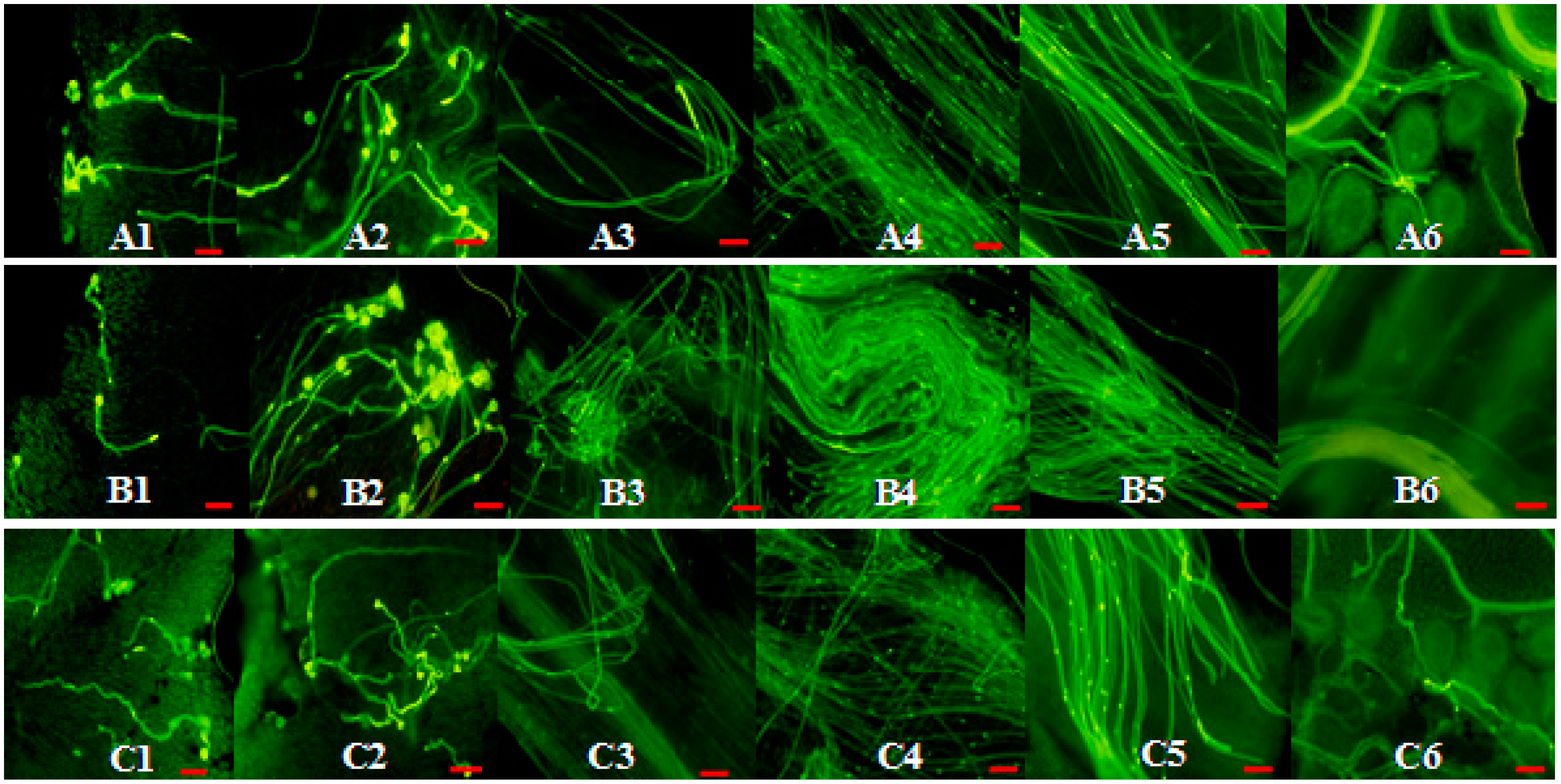
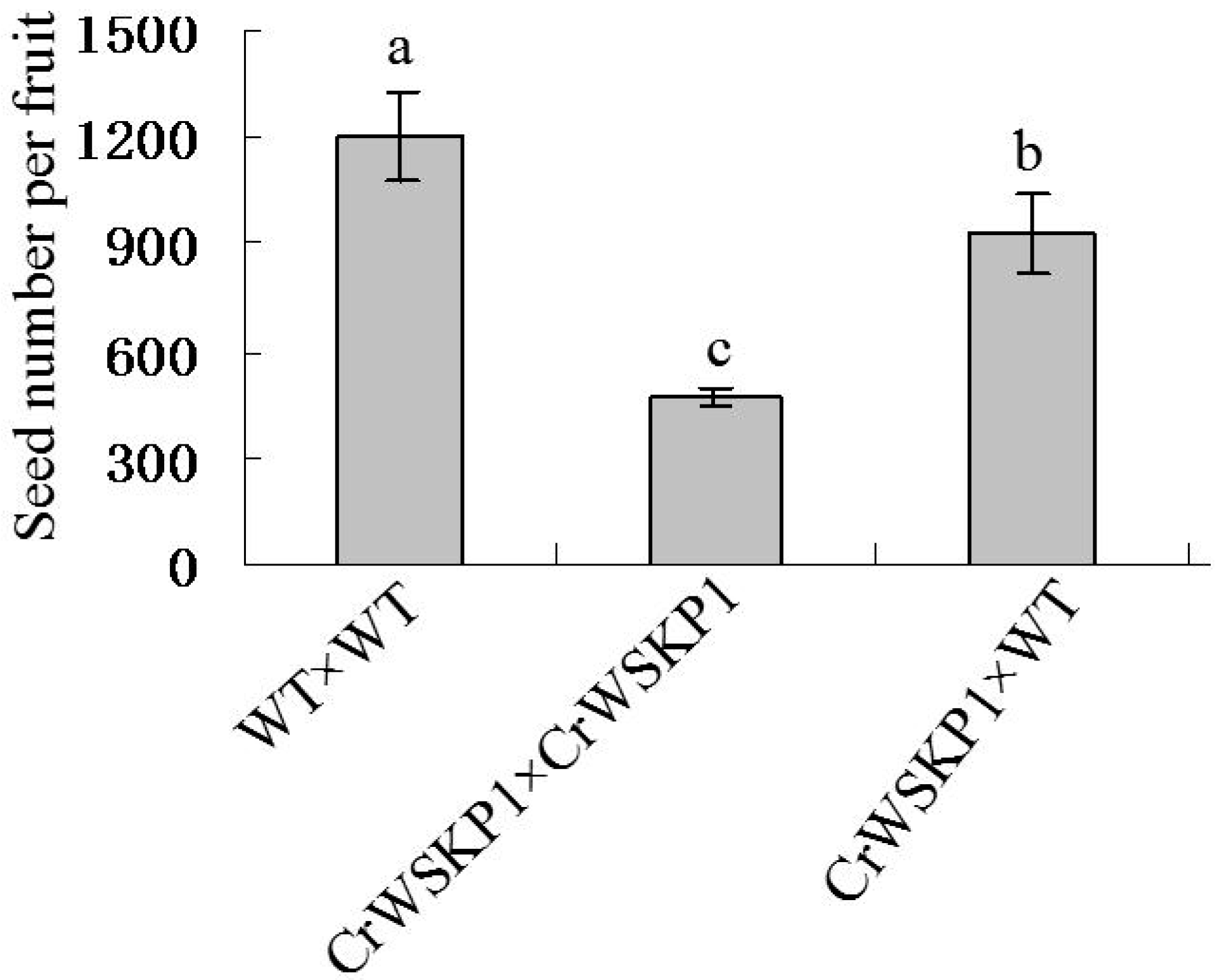
3. Discussions
4. Experimental Section
4.1. Plant Materials
4.2. Genomic DNA and Total RNA Extraction
4.3. Cloning and Sequence Analysis of Skp1-like Gene

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequences (5′–3′) | Procedures of PCR |
|---|---|---|
| SKP1-F | GAAACGATGTCGTCTGAGAAGAAGAT | 94 °C 4 min; 94 °C 30 s, 59 °C 30 s, 72 °C 60 s, 35 cycles; 72 °C 10 min |
| SKP1-R | GTCCTTCACTCGAATGCCCATTGGTT | |
| TZSKP1-F | ATAAAATGAGAACTTAATTTAC | 94 °C 4 min; 94 °C 30 s, 57 °C 30 s, 72 °C 60 s, 35 cycles; 72 °C 10 min |
| TZSKP1-R | TCACTCGAATGCCCATTGGTT | |
| NPTII-F | GTTCTTTTTGTCAAGACCGACC | 94 °C 4 min; 94 °C 30 s, 55 °C 30 s, 72 °C 60 s, 35 cycles; 72 °C 10 min |
| NPTII-R | CAAGCTCTTCAGCAATATCACG | |
| 35S-F | GAGGACCTAACAGAACTCG | 94 °C 4 min; 94 °C 30 s, 57 °C 30 s, 72 °C 60 s, 35 cycles; 72 °C 10 min |
| 35S-R | GTCTTGCGAAGGATAGTGG | |
| RTSKP1-F | ATGTCGTCTGAGAAGAAGATC | 94 °C 5 min; 94 °C 30 s, 59 °C 30 s, 72 °C 1 min, 35 cycles; 72 °C 10 min |
| RTSKP1-R | CTCGAATGCCCATTGGTTCTC | |
| ChvA-F | TCCATCAGCAACGTGTCGGTGCT | 94 °C 4 min; 94 °C 30 s, 60 °C 30 s, 72 °C 90 s, 35 cycles; 72 °C 10 min |
| ChvA-R | GTGGAAAGGCGGTGAGCGATGAT | |
| QSKP1-F | CTCTTTGACCTAATCTTGGCAG | 95 °C 1 min; 95 °C 15 s, 55 °C 20 s, 72 °C 30 s, 40 cycles |
| QSKP1-R | CTTGCGGATCTCCTCTGGAGTC | |
| Q-actin-F | CTGGCATTGCAGATCGTATGA | 95 °C 1 min; 95 °C 15 s, 55 °C 20 s, 72 °C 30 s, 40 cycles |
| Q-actin-R | GCGCCACCACCTTGATCTT |
4.4. Southern Blot Analysis
4.5. Genetic Transformation of Tobacco
4.6. Pollination and Fertilization of Transgenic Tobacco
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ye, W.J.; Qin, Y.H.; Ye, Z.X.; Teixeira da Silva, J.A.; Zhang, L.X.; Wu, X.Y.; Lin, S.Q.; Hu, G.B. Seedless mechanism of a new mandarin cultivar “Wuzishatangju” (Citrus reticulata Blanco). Plant Sci. 2009, 177, 19–27. [Google Scholar] [CrossRef]
- Distefano, G.; Caruso, M.; Malfa, S.L.; Gentile, A.; Tribulato, E. Histological and molecular analysis of pollen-pistil interaction in clementine. Plant Cell Rep. 2009, 28, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- Ngo, B.X.; Wakana, A.; Kim, J.H.; Mori, T.; Sakai, K. Estimation of self-incompatibility S genotypes of Citrus cultivars and plants based on controlled pollination with restricted number of pollen grains. J. Fac. Agric. Kyushu Univ. 2010, 55, 67–72. [Google Scholar]
- Caruso, M.; Merelo, P.; Distefano, G.; La Malfa, S.; Lo Piero, A.R.; Tadeo, F.R.; Talon, M.; Gentile, A. Comparative transcriptome analysis of stylar canal cells identifies novel candidate genes implicated in the self-incompatibility response of Citrus clementina. BMC Plant Biol. 2012, 1. [Google Scholar] [CrossRef] [PubMed]
- Mesejo, C.; Yuste, R.; Martínez-Fuentes, A.; Reig, C.; Iglesias, D.J.; Primo-Millo, E.; Agustí, M. Self-pollination and parthenocarpic ability in developing ovaries of self-incompatible Clementine mandarins (Citrus clementina). Physiol. Plant. 2013, 148, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.W.; Ding, F.; He, X.H.; Luo, C.; Huang, G.X.; Hu, Y. Characterization of the ‘Xiangshui’ lemon transcriptome by de novo assembly to discover genes associated with self-incompatibility. Mol. Genet. Genom. 2015, 290, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.W.; Huang, G.X.; Ding, F.; He, X.H.; Pan, J.C. Mechanism of seedlessness in a new lemon cultivar “Xiangshui” (Citrus limon (L.) Burm. F.). Sex. Plant Reprod. 2012, 25, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Takayama, S.; Shimosato, H.; Shiba, H.; Funato, M.; Che, F.S.; Watanabe, M.; Iwano, M.; Isogai, A. Direct ligand-receptor complex interaction controls Brassica self-incompatibility. Nature 2001, 413, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Sassa, H.; Kakui, H.; Minamikawa, M. Pollen-expressed F-box gene family and mechanism of S-RNase-based gametophytic self-incompatibility (GSI) in Rosaceae. Sex. Plant Reprod. 2010, 23, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Kao, T.H. Self-incompatibility in Petunia inflata: The relationship between a self-incompatibility locus F-box protein and its non-self S-RNases. Plant Cell 2013, 25, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, L.; Yang, Q.Y.; Xue, Y.B. AhSSK1, a novel SKP1-like protein that interacts with the S-locus F-box protein SLF. Plant J. 2006, 46, 780–793. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Huang, J.; Zhao, Z.H.; Li, Q.; Sims, T.L.; Xue, Y.B. The Skp1-like protein SSK1 is required for cross-pollen compatibility in S-RNase-based self-incompatibility. Plant J. 2010, 62, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.X.; Ye, Z.X.; Qin, Y.H.; Hu, G.B. Molecular characterization and expression analysis of S1 self-incompatibility locus-linked pollen 3.15 gene in Citrus reticulata. J. Integr. Plant Biol. 2013, 55, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.X.; Ye, Z.X.; Qin, Y.H.; Hu, G.B. Molecular characterization and expression analysis of ubiqutitn-activating enzyme E1 gene in Citrus reticulata. Gene 2013, 513, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.X.; Ye, Z.X.; Qin, Y.H.; Hu, G.B. Identifying differentially expressed genes in pollen from self-incompatible “Wuzishatangju” and self-compatible “Shatangju” mandarins. Int. J. Mol. Sci. 2013, 14, 8538–8555. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Meng, D.; Gu, Z.Y.; Li, W.; Wang, A.D.; Yang, Q.; Zhu, Y.D.; Li, T.Z. A novel gene, MdSSK1, as a component of the SCF complex rather than MdSBP1 can mediate the ubiquitination of S-RNase in apple. J. Exp. Bot. 2014, 65, 3121–3131. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Gu, Z.Y.; Li, W.; Wang, A.D.; Yuan, H.; Yang, Q.; Li, T.Z. Apple MdABCF assists in the transportation of S-RNase into pollen tubes. Plant J. 2014, 78, 990–1002. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Gu, Z.Y.; Wang, A.D.; Yuan, H.; Li, W.; Yang, Q.; Duan, X.W.; Li, T.Z. Screening and characterization of apple Rho-like GTPase (MdROPs) genes related to S-RNase mediated self-incompatibility. Plant Cell Tissue Organ Cult. 2014, 117, 465–476. [Google Scholar] [CrossRef]
- Wheeler, M.J.; Armstrong, S.A.; Franklin-Tong, V.E.; Franklin, F.C.H. Genomic organization of the Papaver rhoeas self-incompatibility S1 locus. J. Exp. Bot. 2003, 54, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.G.; Franklin-Tong, V.E. Self-incompatibility triggers programmed cell death in Papaver pollen. Nature 2004, 429, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Porat, R.; Lu, P.; O’Neill, S.D. Arabidopsis SKP1, a homologue of a cell cycle regulator gene, is predominantly expressed in meristematic cells. Planta 1998, 204, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Kuroda, H.; Kuromori, T.; Hirayama, T.; Seki, M.; Shinozaki, K.; Shimada, H.; Matsui, M. Expression and interaction analysis of Arabidopsis Skp1-related genes. Plant Cell Physiol. 2004, 45, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Dezfulian, M.H.; Soulliere, D.M.; Dhaliwal, R.K.; Sareen, M.; Crosby, W.L. The SKP1-like gene family of Arabidopsis exhibits a high degree of differential gene expression and gene product interaction during development. PLoS ONE 2012, 7, e50984. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.Z.; Ni, W.M.; Feng, B.M.; Han, T.F.; Petrasek, M.G.; Ma, H. Members of the Arabidopsis-SKP1-like gene family exhibit a variety of expression patterns and may play diverse roles in Arabidopsis. Plant Physiol. 2003, 133, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Chai, L.J.; Biswas, M.K.; Ge, X.X.; Deng, X.X. Isolation, characterization, and expression analysis of an SKP1-like gene from “Shatian” Pummelo (Citrus grandis Osbeck). Plant Mol. Biol. Rep. 2010, 28, 569–577. [Google Scholar] [CrossRef]
- Hong, M.J.; Kim, D.Y.; Seo, Y.W. SKP1-like-related genes interact with various F-box proteins and may form SCF complexes with Cullin-F-box proteins in wheat. Mol. Biol. Rep. 2013, 40, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Hu, Y.; Lodhi, M.; McCombie, W.R.; Ma, H. The Arabidopsis SKP1-LIKE1 gene is essential for male meiosis and may control homologue separation. Proc. Natl. Acad. Sci. USA 1999, 96, 11416–11421. [Google Scholar] [CrossRef] [PubMed]
- Sijacic, P.; Wang, X.; Skirpan, A.L.; Wang, Y.; Dowd, P.E.; McCubbin, A.G.; Huang, S.; Kao, T.H. Identification of the pollen determinant of S-RNase-mediated self-incompatibility. Nature 2004, 429, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.C.; Guo, C.L.; Lin, Y.S.; Fu, H.; Wang, C.S.; Jauh, G.Y. Pollen-specific SKP1-like proteins are components of functional SCF complexes and essential for lily pollen tube elongation. Plant Cell Physiol. 2009, 50, 1558–1572. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, D.; Yamane, H.; Abe, K.; Tao, R. Identification of a Skp1-like protein interaction with SFB, the pollen S determinant of the gametophytic self-incompatibility in Prunus. Plant Physiol. 2012, 159, 1252–1262. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.X.; Zeng, T.; Xu, J.K.; Luo, Z.D.; Hu, G.B.; Zhang, Z.Q.; Ji, Z.L.; Chen, Y.C.; Chen, G.L.; Chen, L.L.; et al. Wuzishatangju, a new mandarin cultivar. (in Chinese). J. Fruit Sci. 2006, 23, 149–150. [Google Scholar]
- Miao, H.X.; Ye, Z.X.; Qin, Y.H.; Hu, G.B. Identification of differentially expressed genes in 72 h styles from self-incompatible Citrus reticulata. Sci. Hortic. 2013, 161, 278–285. [Google Scholar] [CrossRef]
- Miao, H.X.; Ye, Z.X.; Hu, G.B.; Qin, Y.H. Comparative transcript profiling of gene expression between self-incompatibility and self-compatibility mandarins by suppression subtractive hybridization and cDNA microarray. Mol. Breed. 2015, 35. [Google Scholar] [CrossRef]
- Risseeuw, E.P.; Daskalchuk, T.E.; Banks, T.W.; Liu, E.; Cotelesage, J.L.; Hellmann, H.; Estelle, M.; Somers, D.E.; William, L. Protein interaction analysis of SCF ubiquitin E3 ligase subunits from Arabidopsis. Plant J. 2003, 34, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Timofejeva, L.; Ma, H.; Makaroff, C. The Arabidopsis SKP1 homolog ASK1 controls meiotic chromosome remodeling and release of chromatin from the nuclear membrane and nucleolus. J. Cell Sci. 2006, 119, 3754–3763. [Google Scholar] [CrossRef] [PubMed]
- Li, C.J.; Liang, Y.; Chen, C.B.; Li, J.H.; Xu, Y.Y.; Xu, Z.H.; Ma, H.; Chong, K. Cloning and expression analysis of TSK1, a wheat SKP1 homologue,and functional comparison with Arabidopsis ASK1 in male meiosis and auxin signaling. Funct. Plant Biol. 2006, 33, 381–390. [Google Scholar] [CrossRef]
- Kong, H.; Landherr, L.L.; Frohlich, M.W.; Leebens-Mack, J.; Ma, H.; de Pamphilis, C.W. Patterns of gene duplication in the plant SKP1 gene family in angiosperms: Evidence for multiple mechanisms of rapid gene birth. Plant J. 2007, 50, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Kahloul, E.; HajSalah, E.B.; Boulaflous, A.; Ferchichi, A.; Kong, H.; Mouzeyar, S.; Bouzidi, M.F. Structural, expression and interaction analysis of rice SKP1-like genes. DNA Res. 2013, 20, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Minamikawa, M.F.; Koyano, R.; Kikuchi, S.; Koba, T.; Sassa, H. Identification of SFBB-containing canonical and noncanonical SCF complexes in pollen of apple (Malus×domestica). PLoS ONE 2014, 9, e97642. [Google Scholar] [CrossRef] [PubMed]
- Schulman, B.A.; Carrano, A.C.; Jeffrey, P.D.; Bowen, Z.; Kinnucan, E.R.; Finnin, M.S.; Elledge, S.J.; Harper, J.W.; Pagano, M.; Pavletich, N.P. Insights into SCF ubiquitin ligases from the structure of the Skp1-Skp2 complex. Nature 2000, 408, 381–386. [Google Scholar] [PubMed]
- Kong, H.; Leebens-Mack, J.; Ni, W.; de Pamphilis, C.W.; Ma, H. Highly heterogeneous rates of evolution in the SKP1 gene family in plants and animals: Functional and evolutionary implications. Mol. Biol. Evol. 2004, 21, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.; Parry, G.; Estelle, M. The ubiquitin-proteasome pathway and plant development. Plant Cell 2004, 16, 3181–3195. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Wang, H.; Zhao, L.; Zhou, J.L.; Huang, J.; Zhang, Y.S.; Xue, Y.B. The F-box protein AhSLF-S physically interacts with S-RNases that may be inhibited by the ubiquitin/26S proteasome pathway of protein degradation during compatible pollination in Antirrhinum. Plant Cell 2004, 16, 582–595. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.M.; Lan, X.G.; Li, Y.H. The Ub/26S proteasome pathway and self-incompatible responses in flowering plants. (in Chinese). Chin. Bull. Bot. 2006, 23, 197–206. [Google Scholar]
- Zhang, Y.J.; Zhao, Z.H.; Xue, Y.B. Roles of proteolysis in plant self-incompatibility. Annu. Rev. Plant Biol. 2009, 60, 21–42. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhang, B.; Liu, L.J.; Li, Q.; Zhang, Y.E.; Xie, Q.; Xue, Y.B. Identification of a ubiquitin-binding structure in the S-locus F-Box protein controlling S-RNase-based self-incompatibility. J. Genet. Genom. 2012, 39, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Song, G.L.; Cui, R.X.; Wang, K.B.; Guo, L.P.; Li, S.H. A rapid improved CTAB method for extraction of cotton genomic DNA. (in Chinese). Acta Gossypii Sin. 1998, 10, 273–275. [Google Scholar]
- The National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov/ (accessed on 25 January 2014).
- Sambrook, J.F.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; pp. 474–490. [Google Scholar]
- Miao, H.X.; Qin, Y.H.; Teixeira da Silva, J.A.; Ye, Z.X.; Hu, G.B. Cloning and expression analysis of S-RNase homologous gene in Citrus reticulate Blanco cv. Wuzishatangju. Plant Sci. 2011, 180, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.J.; Hou, X.L. Measurement of self-incompatible by fluoroscope observation in non-heading Chinese cabbage. (in Chinese). J. Wuhan Bot. Res. 2004, 22, 197–200. [Google Scholar]
- Tao, S.T.; Zhang, S.L.; Chen, D.X.; Wei, B.Y. Study on characteristics of in situ pollen germination and tube growth of Prunus mume. (in Chinese). J. Fruit Sci. 2004, 21, 338–340. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Miao, H.; Ma, Y.; Wang, L.; Hu, G.; Ye, Z.; Zhao, J.; Qin, Y. CrWSKP1, an SKP1-like Gene, Is Involved in the Self-Incompatibility Reaction of “Wuzishatangju” (Citrus reticulata Blanco). Int. J. Mol. Sci. 2015, 16, 21695-21710. https://doi.org/10.3390/ijms160921695
Li P, Miao H, Ma Y, Wang L, Hu G, Ye Z, Zhao J, Qin Y. CrWSKP1, an SKP1-like Gene, Is Involved in the Self-Incompatibility Reaction of “Wuzishatangju” (Citrus reticulata Blanco). International Journal of Molecular Sciences. 2015; 16(9):21695-21710. https://doi.org/10.3390/ijms160921695
Chicago/Turabian StyleLi, Peng, Hongxia Miao, Yuewen Ma, Lu Wang, Guibing Hu, Zixing Ye, Jietang Zhao, and Yonghua Qin. 2015. "CrWSKP1, an SKP1-like Gene, Is Involved in the Self-Incompatibility Reaction of “Wuzishatangju” (Citrus reticulata Blanco)" International Journal of Molecular Sciences 16, no. 9: 21695-21710. https://doi.org/10.3390/ijms160921695