
Polymodal Transient Receptor Potential Vanilloid (TRPV) Ion Channels in Chondrogenic Cells

 ,
,
Abstract
:1. Introduction
2. Results
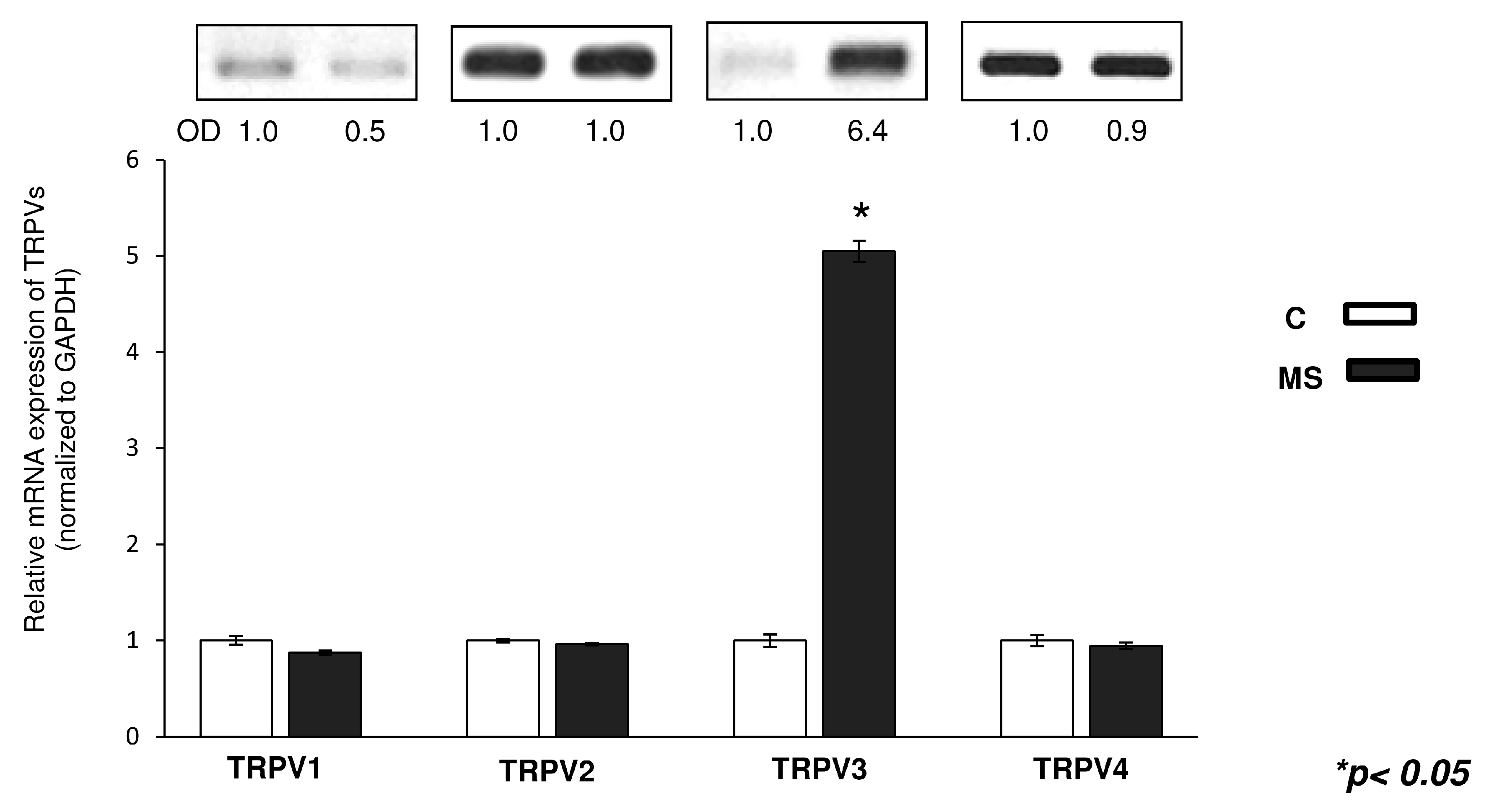
2.1. mRNA Expression Profiling of TRPV Ion Channels in Chicken and Mouse Tissue Samples
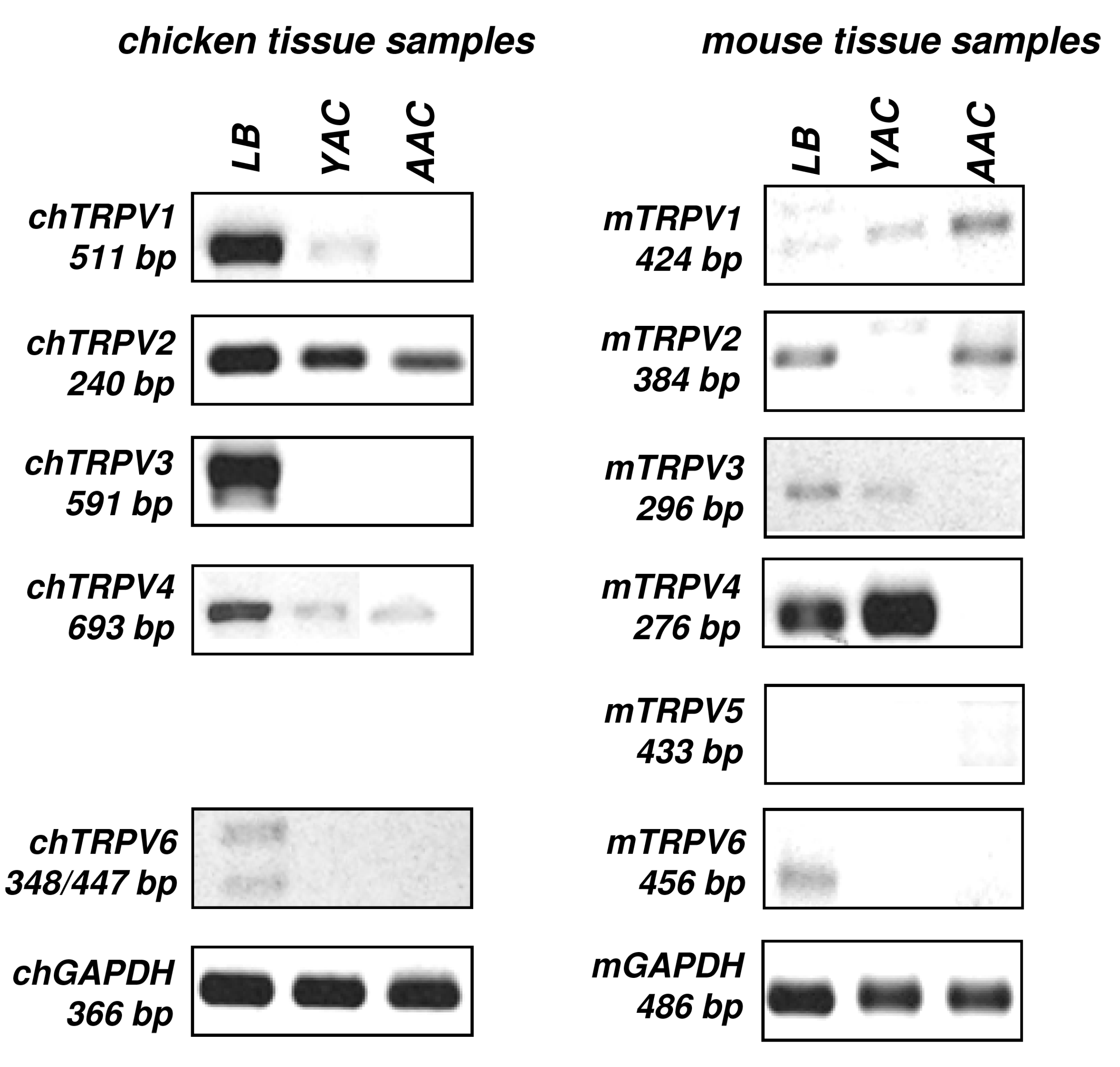

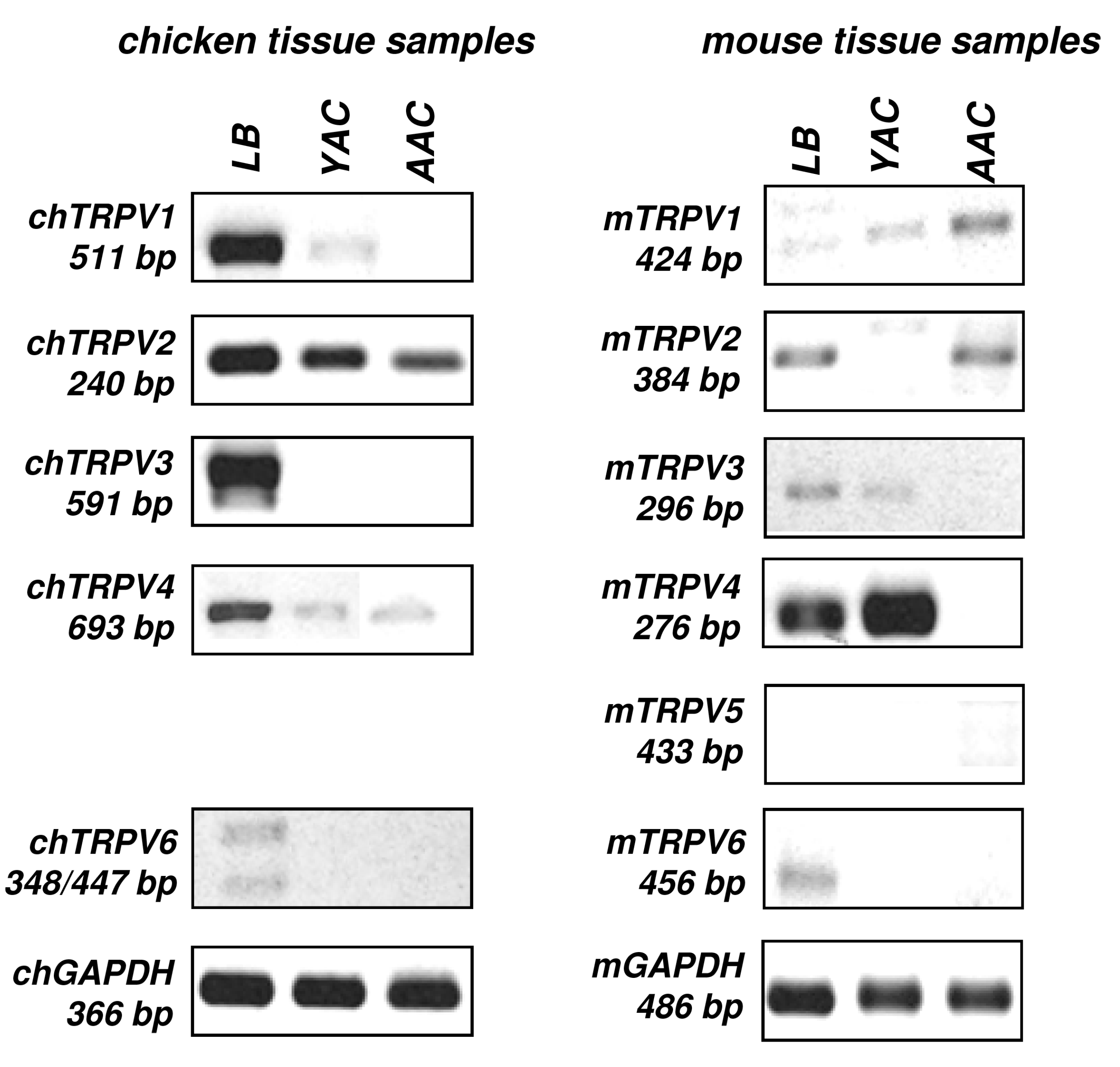{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chicken Gene | Primer | Primer Sequences (5′→3′) | GenBank Accession No. | * | Amplimer Size (bp) |
|---|---|---|---|---|---|
| Transient Receptor Potential Vanilloid type 1 (TRPV1_chick) | forward | TTCGTTCACTCTTTGCTCCTC (1633–1653) | NM_204572.1 | 58 | 511 |
| reverse | TGCTCACAGTTTCTCCCATCA (2143–2123) | ||||
| Transient Receptor Potential Vanilloid type 2 (TRPV2_chick) | forward | CCCTTGGAGTCACCTTACC (547–565) | XM_004946687.1 | 54 | 240 |
| reverse | CTTCCCAGTCTTTGCATCT (786–768) | ||||
| Transient Receptor Potential Vanilloid type 3 (TRPV3_chick) | forward | CCCCTCAATTCACTCCTGC (2794–2812) | XM_004946676.1 | 60 | 591 |
| reverse | GGAAAGGCATTCACCACCA (3384–3366) | ||||
| Transient Receptor Potential Vanilloid type 4 (TRPV4_chick) | forward | TCGCCGAGAAGACGGGAAAC (733–752) | NM_204692.1 | 60 | 693 |
| reverse | GGCGGTTCTCAATCTTGCTGTT (1425–1404) | ||||
| Transient Receptor Potential Vanilloid type 6 (TRPV6_chick) | forward | CATGTAGCTGCCTTGTATGA (806–825) or (429–448) | XM_004938143.1 | 52 | 348 |
| reverse | TGATCTTGGTCCCTCTTTG (1153–1135) or (875–857) | XM_004938142.1 | 447 | ||
| Aggrecan core protein (ACAN_chick) | forward | CAATGCAGAGTACAGAGA (435–452) | NM_204955.2 | 54 | 429 |
| reverse | TCTGTCTCACGGACACCG (863–846) | ||||
| Collagen II (COL2A1_chick) | forward | GGACCCAAAGGACAGACGG (1191–1209) | NM_204426.1 | 59 | 401 |
| reverse | TCGCCAGGAGCACCAGTT (1591–1574) | ||||
| SRY (sex determining region Y)-box 9 (SOX9_chick) | forward | CCCCAACGCCATCTTCAA (713–730) | NM_204281.1 | 54 | 381 |
| reverse | CTGCTGATGCCGTAGGTA (1093–1076) | ||||
| Glyceraldehyde 3-phosphate dehydrogenase (GAPDH_chick) | forward | CTGCCCAGAACATCATCCCA (656–675) | NM_204305.1 | 58 | 366 |
| reverse | CACGGTTGCTGTATCCAAACTCAT (1021–998) |
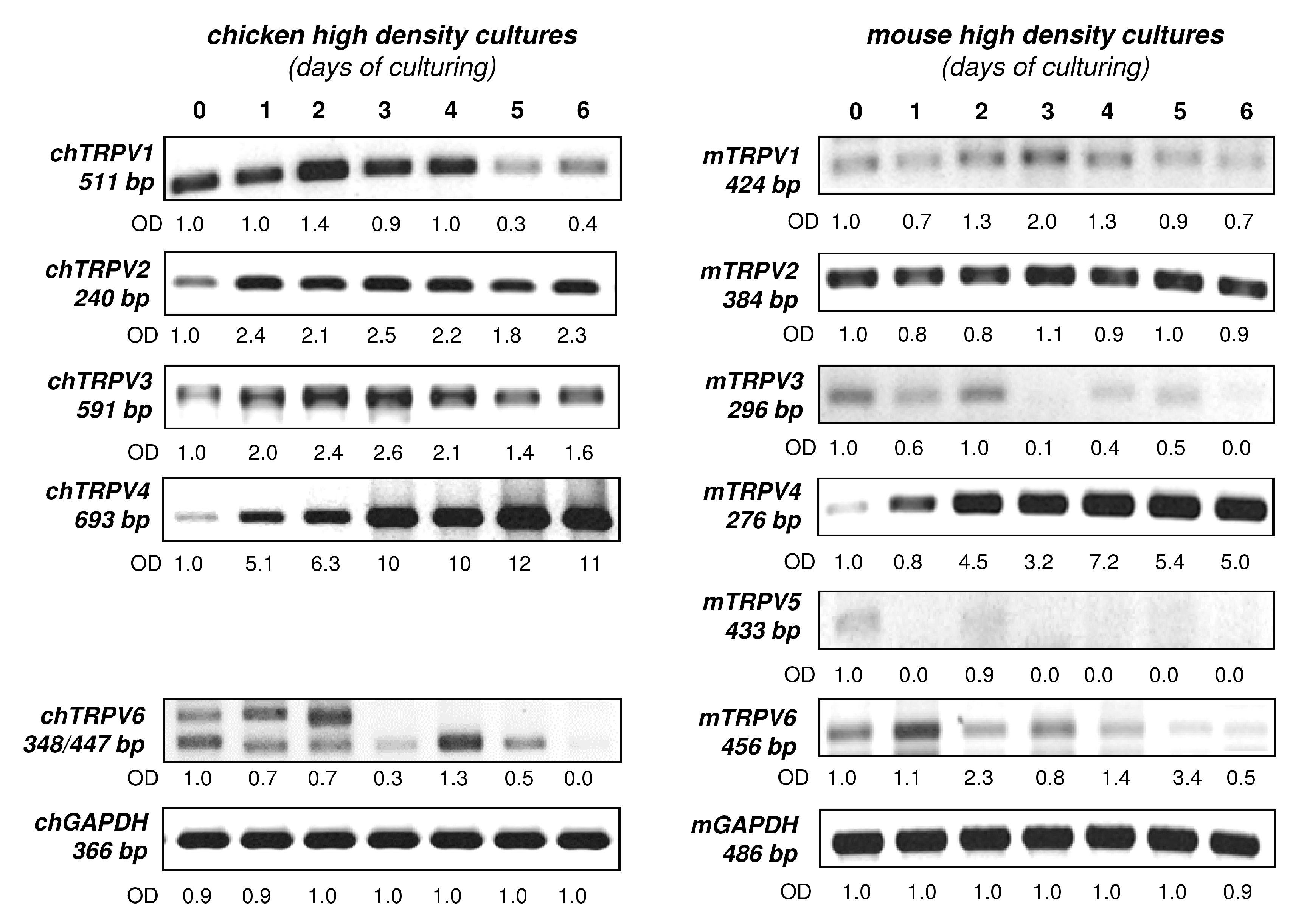
2.2. mRNA Expression Profiling of TRPV in Embryonic Limb Bud-Derived Micromass Cultures

| Mouse Gene | Primer | Primer Sequences (5′→3′) | GenBank Accession No. | * | Amplimer Size (bp) |
|---|---|---|---|---|---|
| Transient Receptor Potential Vanilloid type 1 (TRPV1_mouse) | forward | CTCTTACAACAGCCTGTATTCC (2130–2151) | NM_001001445.2 | 59 | 424 |
| reverse | ACAGTTGCCTGGGTCCTC (2553–2536) | ||||
| Transient Receptor Potential Vanilloid type 2 (TRPV2_mouse) | forward | CTTTGCTGTAGCCCTAGTAAGC (2007–2028) | NM_011706.2 | 59 | 384 |
| reverse | CACCACCAGTAACCATTCTCC (2390–2370) | ||||
| Transient Receptor Potential Vanilloid type 3 (TRPV3_mouse) | forward | AGCAGAACTCCACCTACCC (1998–2016) | NM_145099.2 | 58 | 296 |
| reverse | TTTCCATTCCGTCCACTT (2293–2276) | ||||
| Transient Receptor Potential Vanilloid type 4 (TRPV4_mouse) | forward | TCTGTCTCGCAAGTTCAAGG (1314–1333) | NM_022017.3 | 59 | 276 |
| reverse | GGCTGATAGTAGGCGGTGA (1589–1571) | ||||
| Transient Receptor Potential Vanilloid type 5 (TRPV5_mouse) | forward | TCCGAGATGCCAACCGTAC (1108–1126) | NM_001007572.2 | 59 | 433 |
| reverse | GCCATTAGCCAGCAGAAGC (1540–1522) | ||||
| Transient Receptor Potential Vanilloid type 6 (TRPV6_mouse) | forward | GCTGGCTGATGGCTGTGGT (1773–1791) | NM_022413.4 | 63 | 456 |
| reverse | GGCGGATGCGTTGTCTGTT (2228–2210) | ||||
| Glyceraldehyde 3-phosphate dehydrogenase (GAPDH_mouse) | forward | TGGCAAAGTGGAGATTGTTG (161–180) | NM_001289726.1 | 58 | 486 |
| reverse | GTCTTCTGGGTGGCAGTGAT (646–627) |
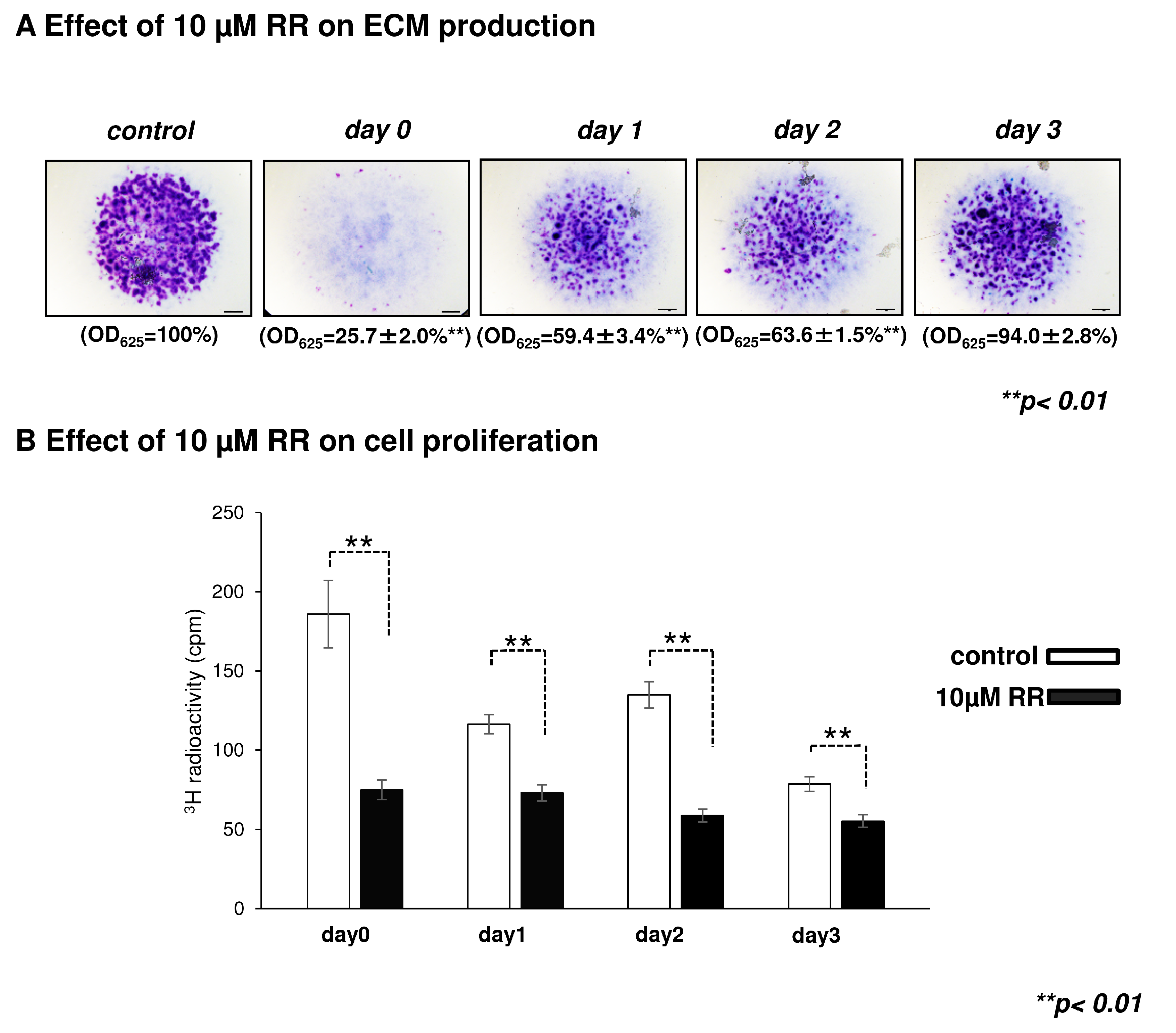
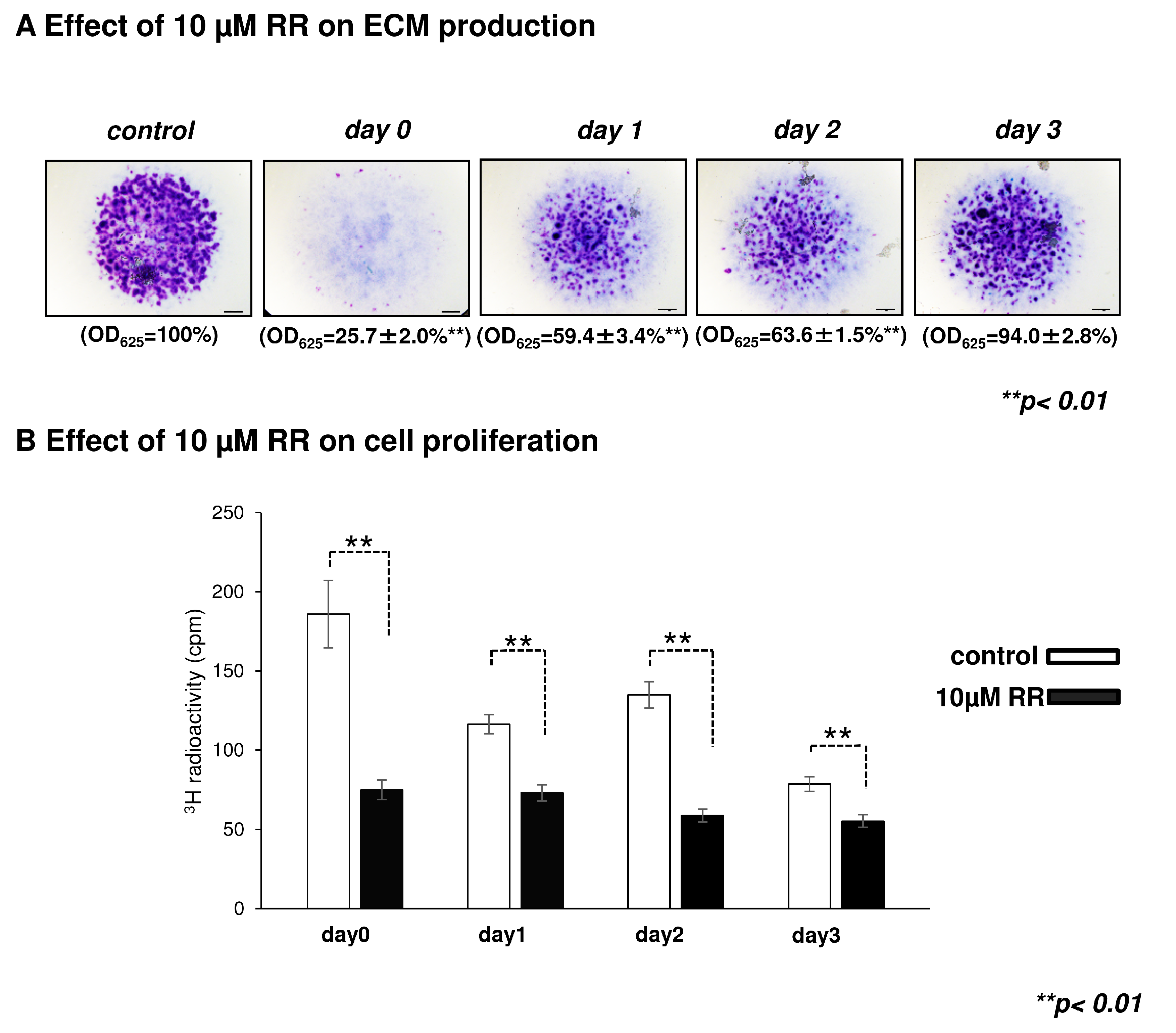
2.3. Effect of Ruthenium Red, a General TRPV Inhibitor, on Chondrogenesis
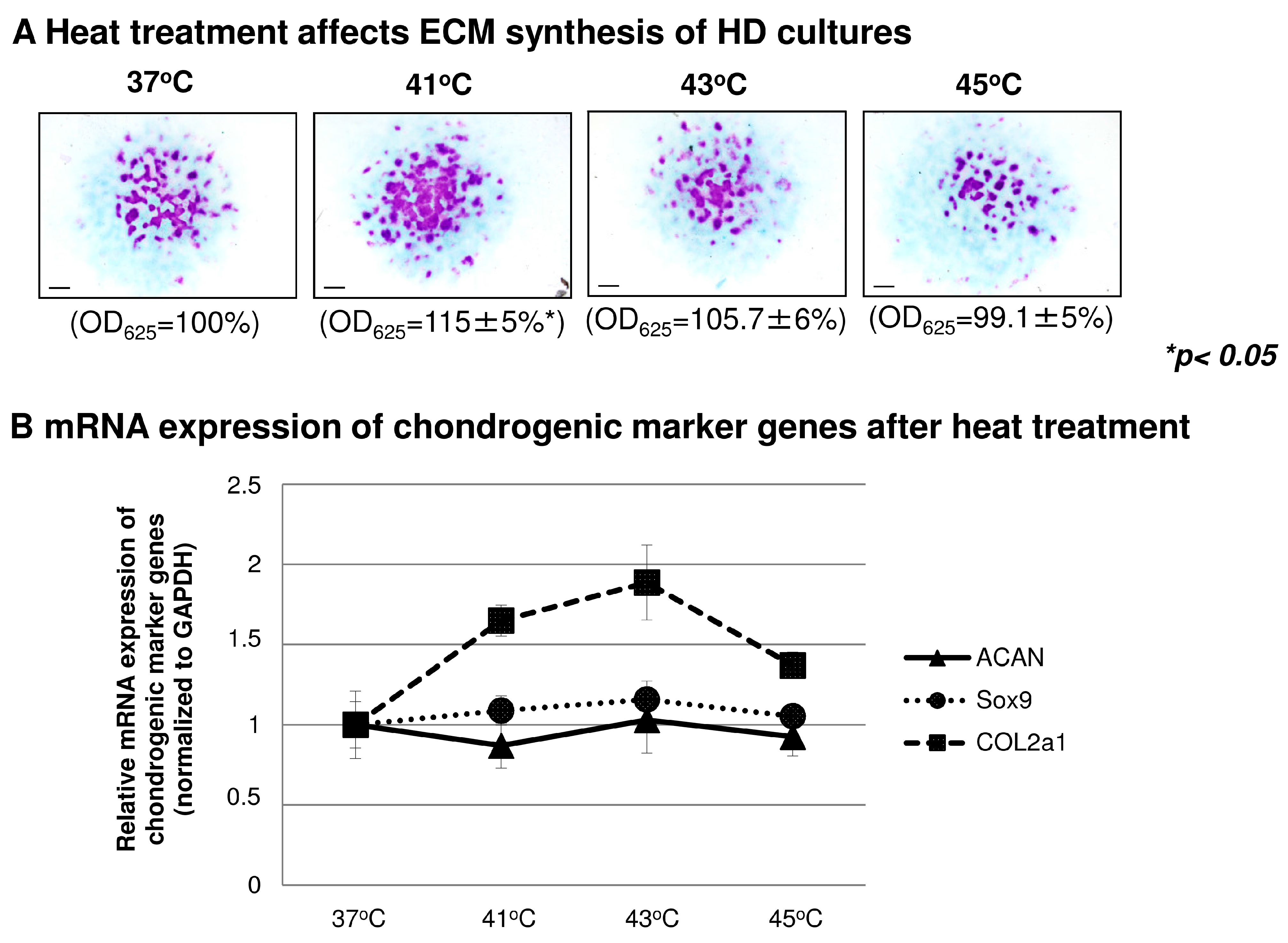
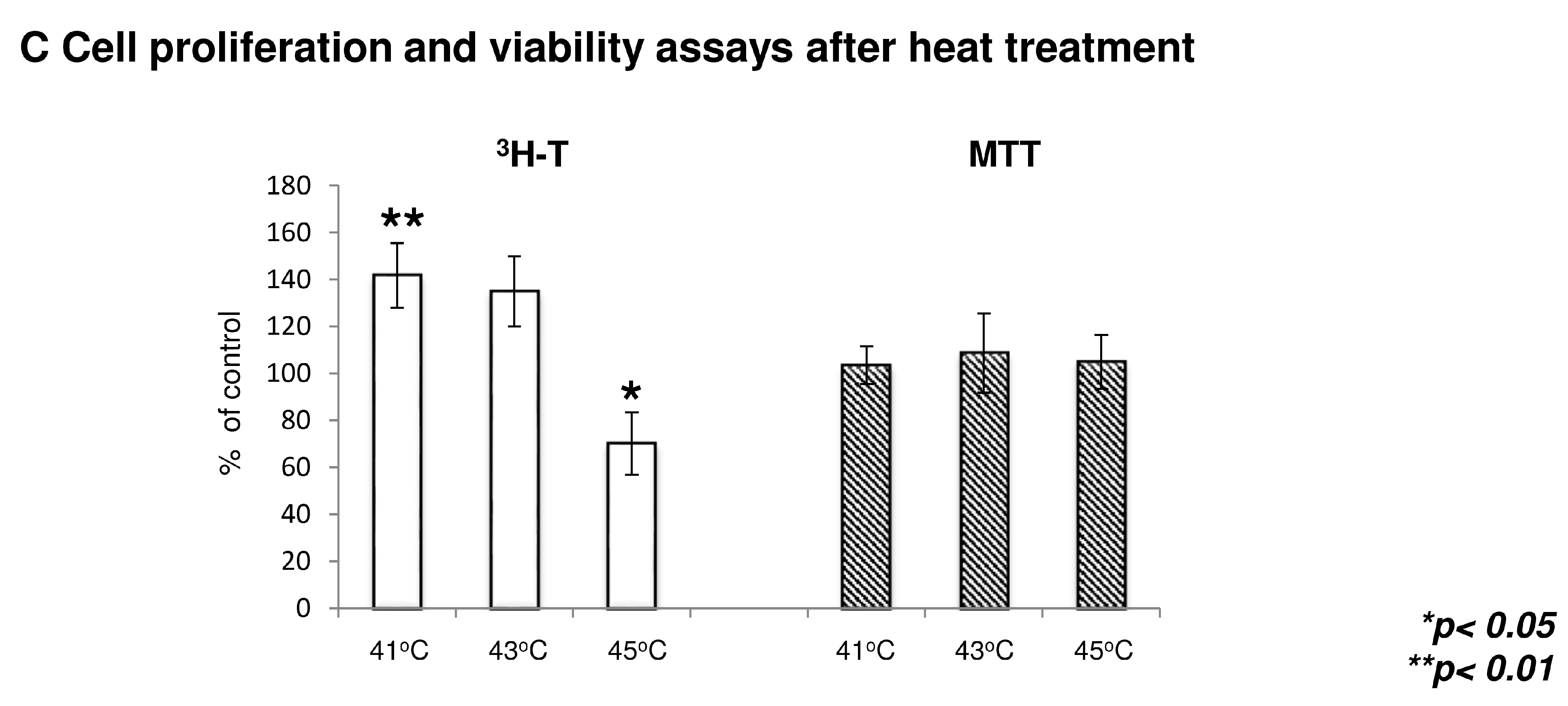
2.4. The Effect of Heat Treatment on Chondrogenesis
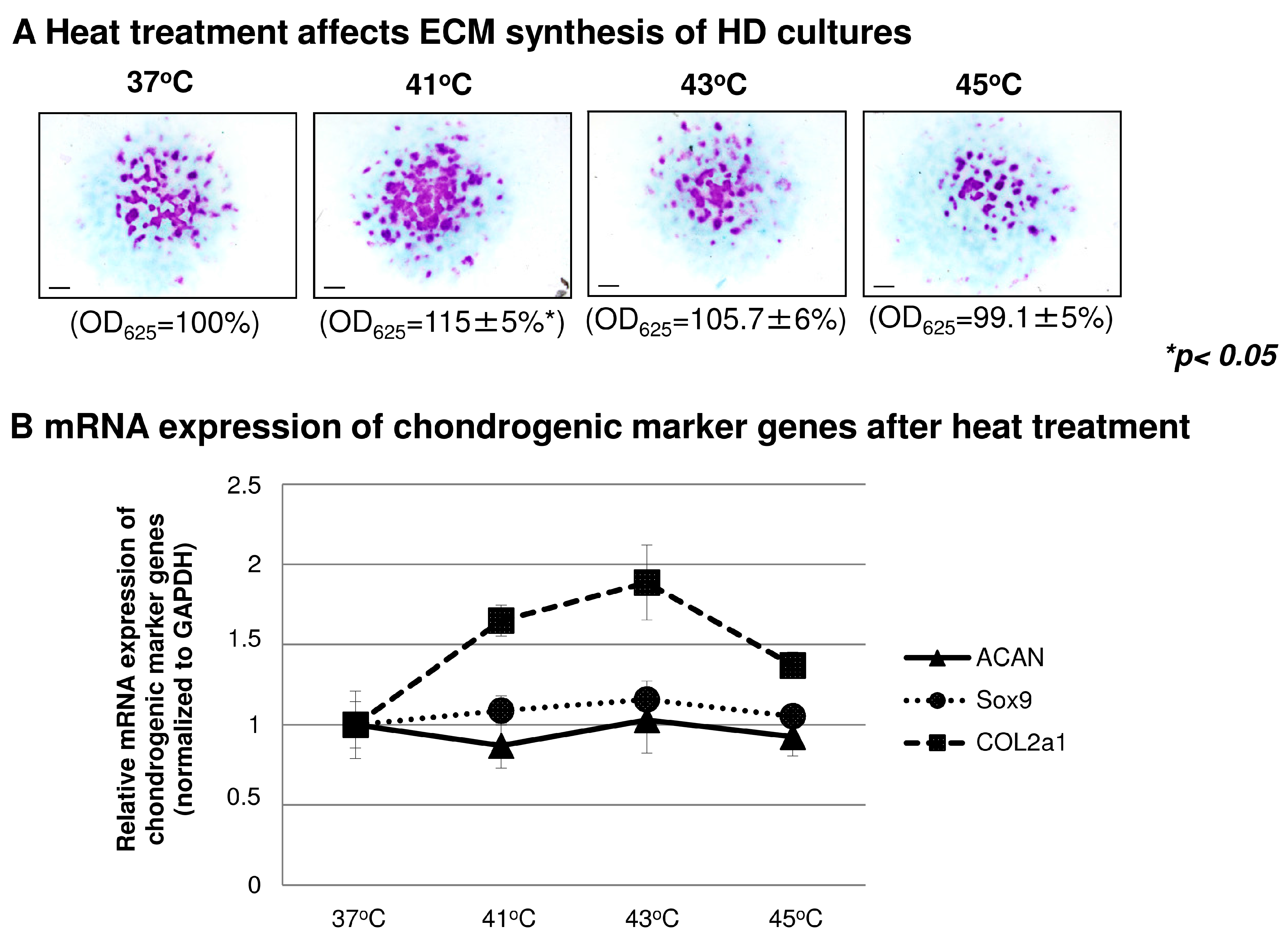
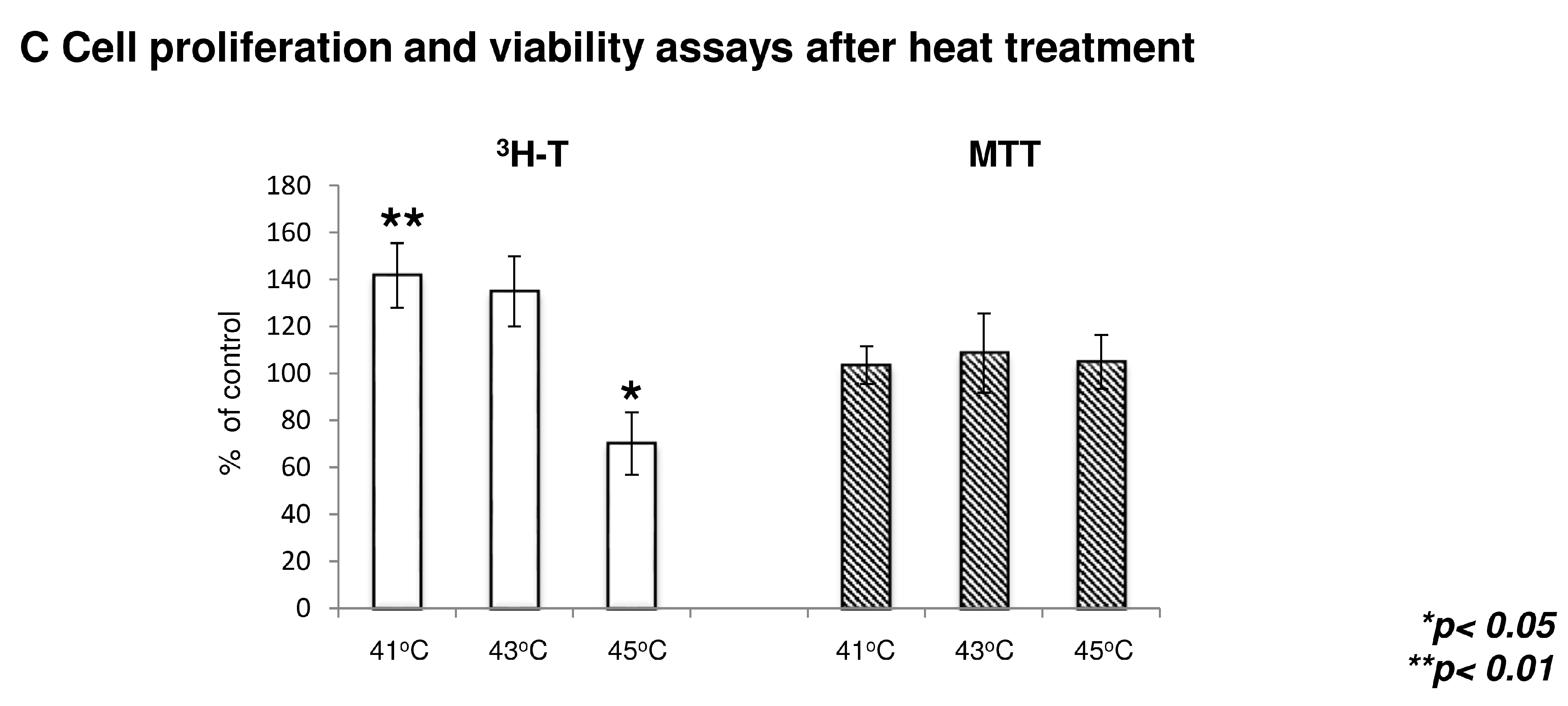
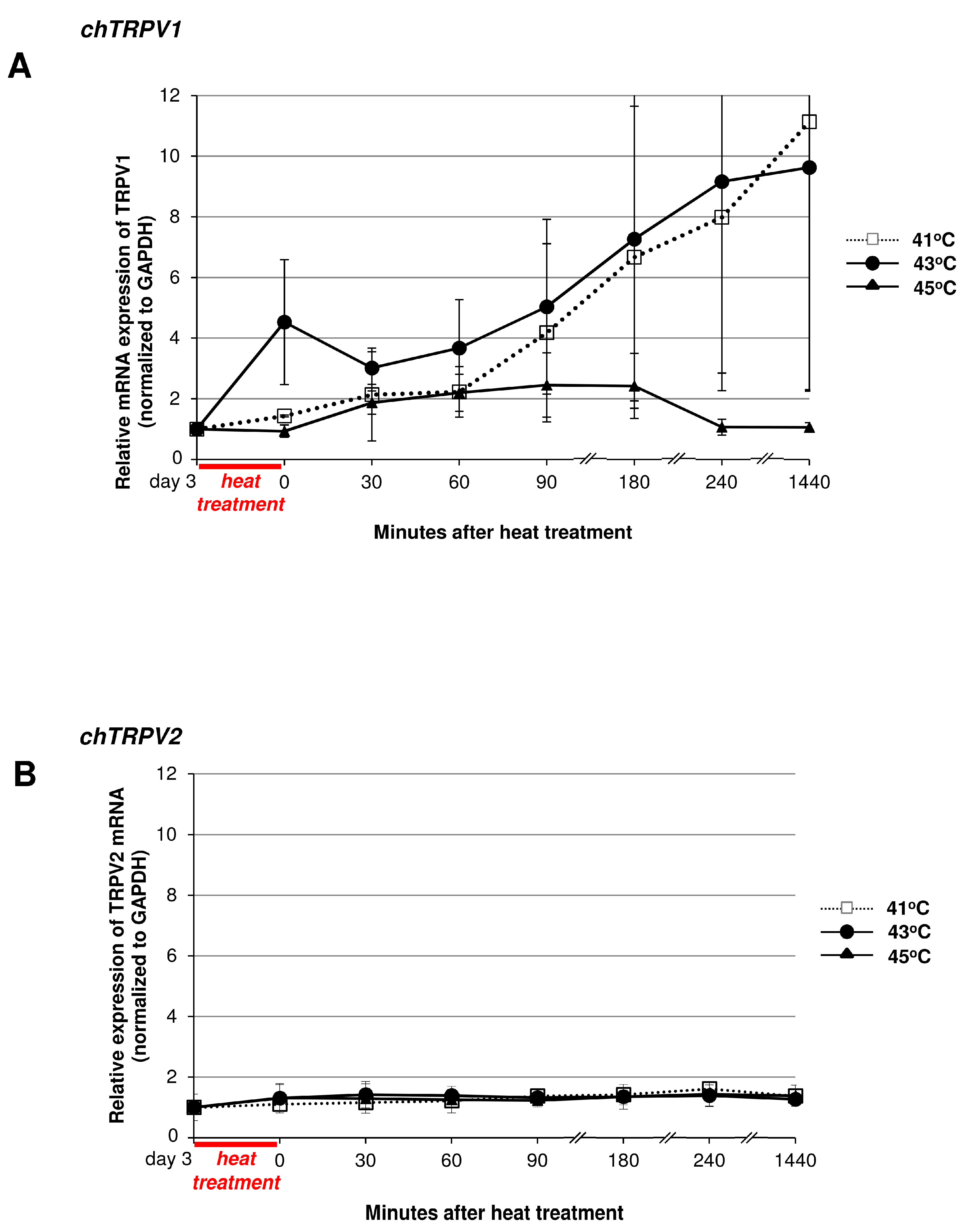
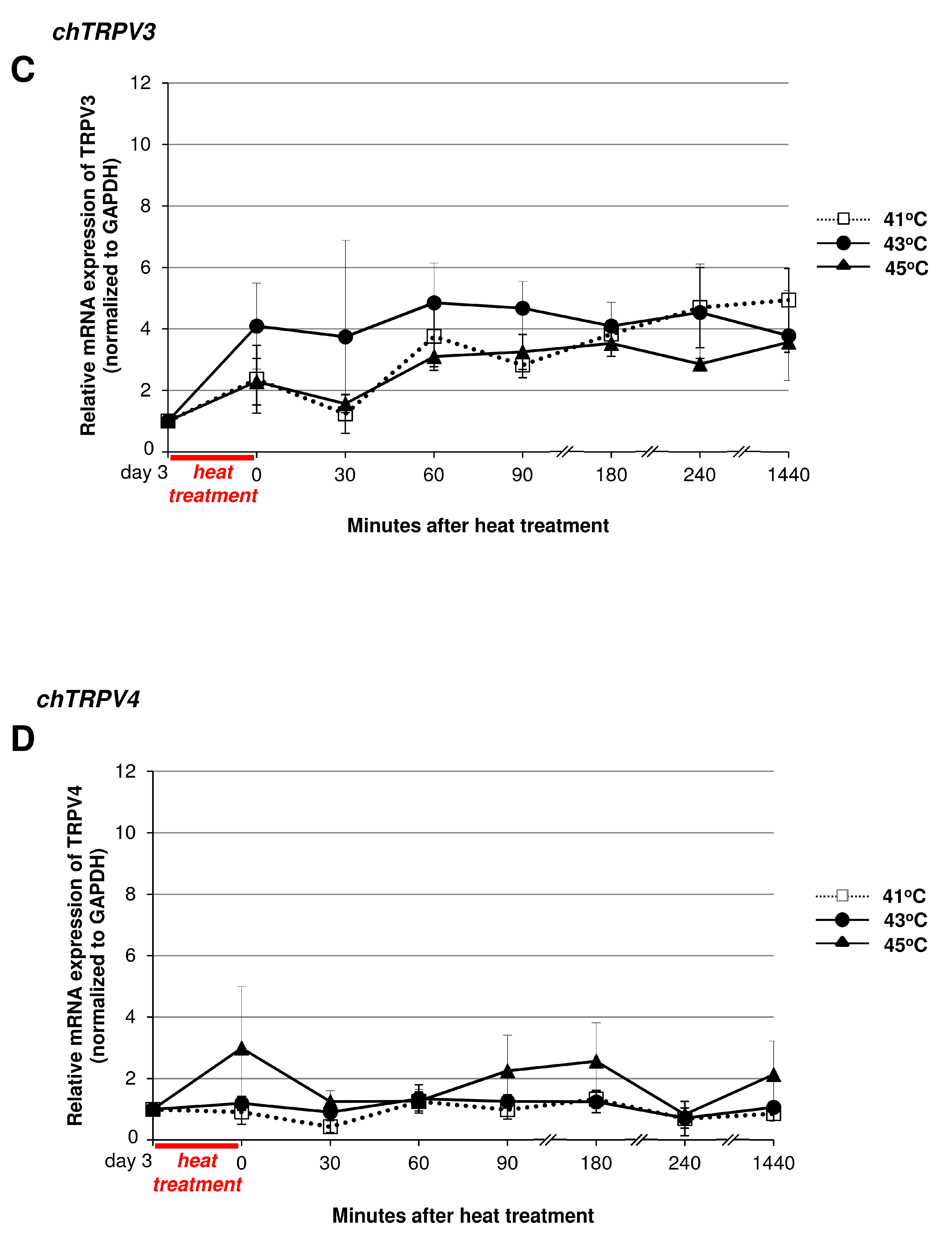
2.5. Heat Treatment and its Effect on High Density (HD) Cultures in the Context of TRPVs
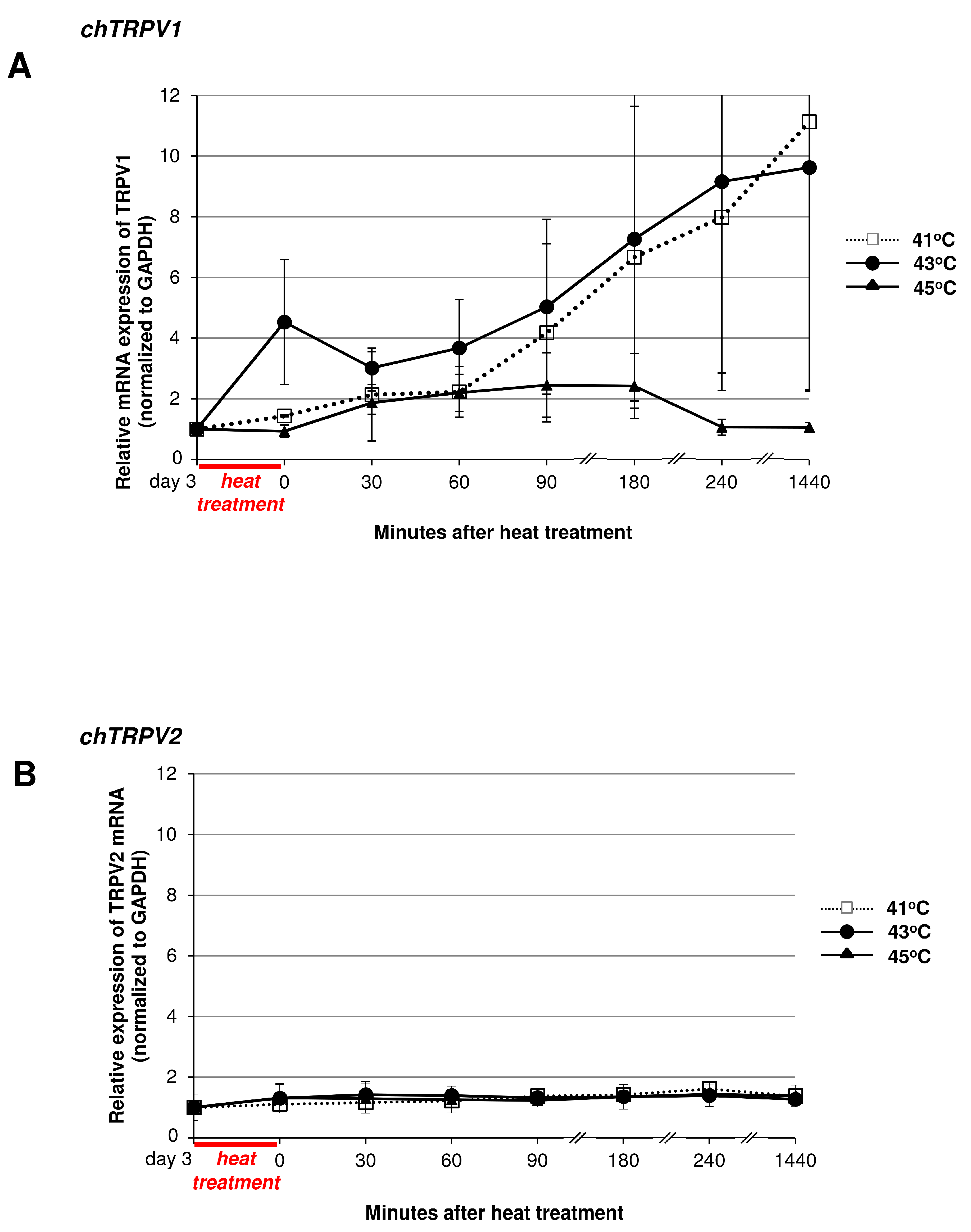
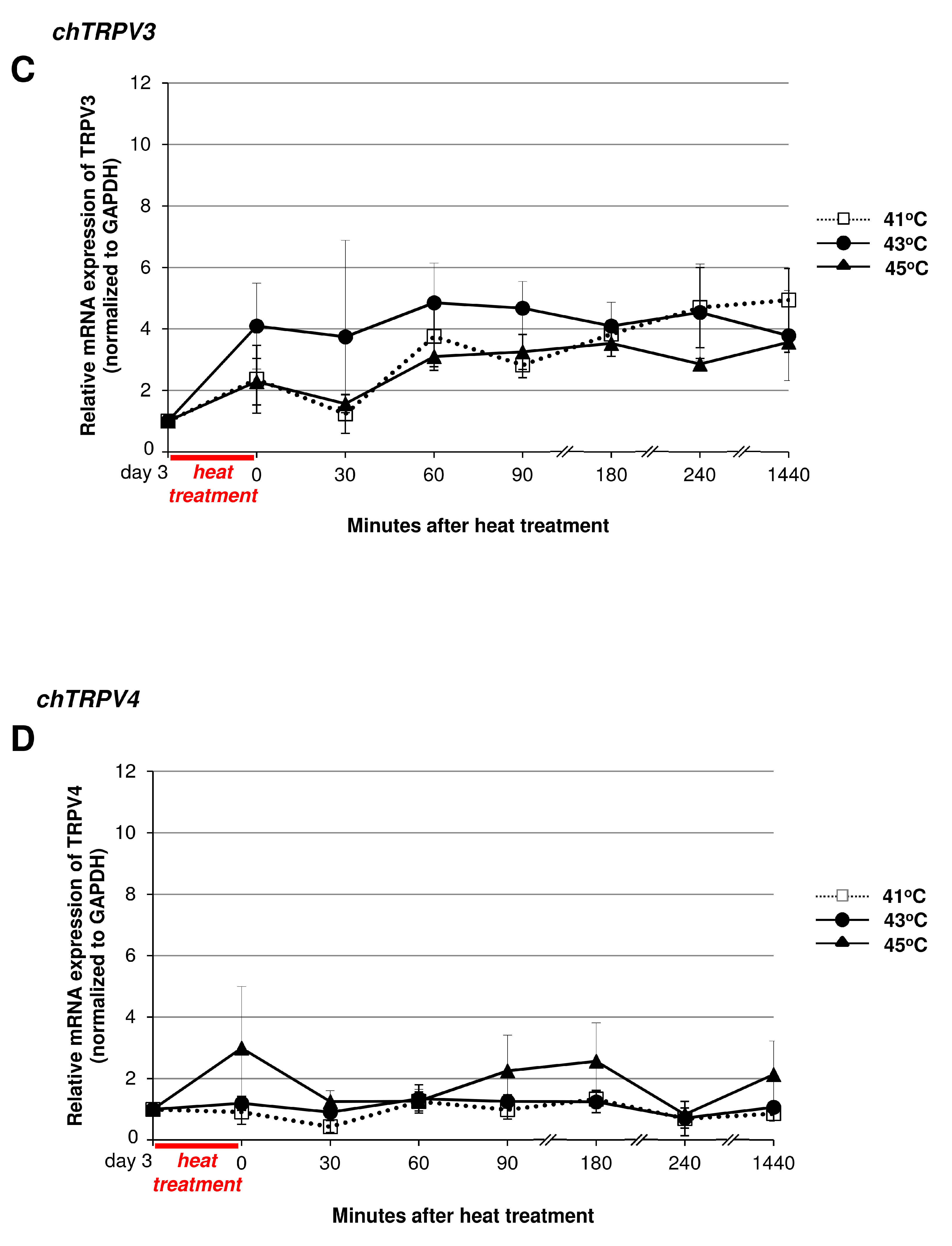
2.6. Mechanical Load in the Context of TRPVs

3. Discussion
3.1. TRPV Ion Channels Are Expressed during in Vitro Chondrogenesis
3.2. Thermal and Mechanical Stimuli Alter Polymodal TRPV Ion Channel Expression in Micromass Cultures
4. Experimental Section
4.1. Tissue Sample Collection
4.2. Chicken and Mouse HD Primer Cell Cultures
4.3. mRNA Expression Analysis Followed by Reverse Transcription Polymerase Chain Reaction
4.4. Heat Treatment of Chicken HD Cultures
4.5. Assessment of Chondrogenic Differentiation by Metachromatic Staining
4.6. Measuring Cell Proliferation with 3H-Thymidine Labelling and Mitochondrial Activity with MTT Assay
4.7. Mechanical Loading of HD Cultures
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AAC | adult articular cartilage |
| YAC | young articular cartilage |
| CREB | cAMP response element-binding protein |
| CMF-PBS | calcium and magnesium-free phosphate buffered saline |
| DMMB | dimethyl-methylene blue |
| ECM | extracellular matrix |
| FBS | foetal bovine serum |
| GAG | glycosaminoglycan |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| HSP | heat shock protein |
| HD | high density |
| LB | limb bud |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NMRI | Naval Medical Research Institute |
| PBS | phosphate buffered saline |
| NMDA | N-methyl-d-aspartate |
| PG | proteoglycan |
| PKA | protein kinase A |
| RR | ruthenium red |
| TB | toluidine blue |
| TRPV | transient receptor potential receptor vanilloid |
| TM | transmembrane |
References
- Barrett-Jolley, R.; Lewis, R.; Fallman, R.; Mobasheri, A. The emerging chondrocyte channelome. Front. Physiol. 2010, 1, 135. [Google Scholar]
- Lee, R.B.; Urban, J.P. Evidence for a negative pasteur effect in articular cartilage. Biochem. J. 1997, 321, 95–102. [Google Scholar] [PubMed]
- Brucker, P.U.; Izzo, N.J.; Chu, C.R. Tonic activation of hypoxia-inducible factor 1α in avascular articular cartilage and implications for metabolic homeostasis. Arthritis Rheum. 2005, 52, 3181–3191. [Google Scholar] [PubMed]
- Oldershaw, R.A. Cell sources for the regeneration of articular cartilage: The past, the horizon and the future. Int. J. Exp. Pathol. 2012, 93, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Grad, S.; Eglin, D.; Alini, M.; Stoddart, M.J. Physical stimulation of chondrogenic cells in vitro: A review. Clin. Orthop. Relat. Res. 2011, 469, 2764–2772. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.A.; Saunders, K.; Urban, J.P.; Wilkins, R.J. The influence and interactions of hydrostatic and osmotic pressures on the intracellular milieu of chondrocytes. Biorheology 2004, 41, 299–308. [Google Scholar] [PubMed]
- Tepic, S.; Macirowski, T.; Mann, R.W. Experimental temperature rise in human hip joint in vitro in simulated walking. J. Orthop. Res. 1985, 3, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Pritchett, J. Heat generated by hip resurfacing prostheses: An in vivo pilot study. J. Long Term. Eff. Med. Implants 2011, 21, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Fialho, J.C.; Fernandes, P.R.; Eca, L.; Folgado, J. Computational hip joint simulator for wear and heat generation. J. Biomech. 2007, 40, 2358–2366. [Google Scholar] [CrossRef] [PubMed]
- Usuba, M.; Miyanaga, Y.; Miyakawa, S.; Maeshima, T.; Shirasaki, Y. Effect of heat in increasing the range of knee motion after the development of a joint contracture: An experiment with an animal model. Arch. Phys. Med. Rehabil. 2006, 87, 247–253. [Google Scholar] [CrossRef] [PubMed]
- June, R.K.; Fyhrie, D.P. Temperature effects in articular cartilage biomechanics. J. Exp. Biol. 2010, 213, 3934–3940. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Aoyama, T.; Iijima, H.; Nagai, M.; Yamaguchi, S.; Tajino, J.; Zhang, X.; Akiyama, H.; Kuroki, H. Optimum temperature for extracellular matrix production by articular chondrocytes. Int. J. Hyperth. 2014, 30, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Matta, C.; Fodor, J.; Szijgyarto, Z.; Juhasz, T.; Gergely, P.; Csernoch, L.; Zakany, R. Cytosolic free Ca2+ concentration exhibits a characteristic temporal pattern during in vitro cartilage differentiation: A possible regulatory role of calcineurin in Ca-signalling of chondrogenic cells. Cell Calcium 2008, 44, 310–323. [Google Scholar] [CrossRef] [PubMed]
- OʼConor, C.J.; Case, N.; Guilak, F. Mechanical regulation of chondrogenesis. Stem Cell Res. Ther. 2013, 4, 61. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, T.; Matta, C.; Somogyi, C.; Katona, E.; Takacs, R.; Soha, R.F.; Szabo, I.A.; Cserhati, C.; Szody, R.; Karacsonyi, Z.; et al. Mechanical loading stimulates chondrogenesis via the PKA/CREB-Sox9 and PP2A pathways in chicken micromass cultures. Cell Signal. 2014, 26, 468–482. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Appendino, G.; Nilius, B. Pharmacology of vanilloid transient receptor potential cation channels. Mol. Pharmacol. 2009, 75, 1262–1279. [Google Scholar] [CrossRef] [PubMed]
- Vennekens, R.; Owsianik, G.; Nilius, B. Vanilloid transient receptor potential cation channels: An overview. Curr. Pharm. Des. 2008, 14, 18–31. [Google Scholar] [PubMed]
- Szabo, A.; Helyes, Z.; Sandor, K.; Bite, A.; Pinter, E.; Nemeth, J.; Banvolgyi, A.; Bolcskei, K.; Elekes, K.; Szolcsanyi, J. Role of transient receptor potential vanilloid 1 receptors in adjuvant-induced chronic arthritis: In vivo study using gene-deficient mice. J. Pharmacol. Exp. Ther. 2005, 314, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Gavenis, K.; Schumacher, C.; Schneider, U.; Eisfeld, J.; Mollenhauer, J.; Schmidt-Rohlfing, B. Expression of ion channels of the TRP family in articular chondrocytes from osteoarthritic patients: Changes between native and in vitro propagated chondrocytes. Mol. Cell. Biochem. 2009, 321, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Hdud, I.M.; El-Shafei, A.A.; Loughna, P.; Barrett-Jolley, R.; Mobasheri, A. Expression of transient receptor potential vanilloid (TRPV) channels in different passages of articular chondrocytes. Int. J. Mol. Sci. 2012, 13, 4433–4445. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, S.; Wakabayashi, M.; Ohno, T.; Amano, K.; Ooishi, R.; Sugahara, T.; Shiojiri, S.; Tashiro, K.; Suzuki, Y.; Nishimura, R.; et al. Functional gene screening system identified TRPV4 as a regulator of chondrogenic differentiation. J. Biol. Chem. 2007, 282, 32158–32167. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.S.; Shin, S.H.; Auh, C.K.; Chun, J. Human skeletal dysplasia caused by a constitutive activated transient receptor potential vanilloid 4 (TRPV4) cation channel mutation. Exp. Mol. Med. 2012, 44, 707–722. [Google Scholar] [CrossRef] [PubMed]
- Rock, M.J.; Prenen, J.; Funari, V.A.; Funari, T.L.; Merriman, B.; Nelson, S.F.; Lachman, R.S.; Wilcox, W.R.; Reyno, S.; Quadrelli, R.; et al. Gain-of-function mutations in TRPV4 cause autosomal dominant brachyolmia. Nat. Genet. 2008, 40, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Eleswarapu, S.V.; Athanasiou, K.A. TRPV4 channel activation improves the tensile properties of self-assembled articular cartilage constructs. Acta Biomater. 2013, 9, 5554–5561. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, R.K.; Julkunen, P.; Wilson, W.; Herzog, W. Importance of collagen orientation and depth-dependent fixed charge densities of cartilage on mechanical behavior of chondrocytes. J. Biomech. Eng. 2008, 130, 021003. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Hermanns, I.; Bittinger, F.; Kirkpatrick, C.J. Induction of stress proteins in human endothelial cells by heavy metal ions and heat shock. Am. J. Physiol. 1999, 277, L1026–L1033. [Google Scholar] [PubMed]
- Kaarniranta, K.; Elo, M.; Sironen, R.; Lammi, M.J.; Goldring, M.B.; Eriksson, J.E.; Sistonen, L.; Helminen, H.J. Hsp70 accumulation in chondrocytic cells exposed to high continuous hydrostatic pressure coincides with mRNA stabilization rather than transcriptional activation. Proc. Natl. Acad. Sci. USA 1998, 95, 2319–2324. [Google Scholar] [CrossRef] [PubMed]
- Cao, E.; Cordero-Morales, J.F.; Liu, B.; Qin, F.; Julius, D. TRPV1 channels are intrinsically heat sensitive and negatively regulated by phosphoinositide lipids. Neuron 2013, 77, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [PubMed]
- Gracheva, E.O.; Cordero-Morales, J.F.; Gonzalez-Carcacia, J.A.; Ingolia, N.T.; Manno, C.; Aranguren, C.I.; Weissman, J.S.; Julius, D. Ganglion-specific splicing of TRPV1 underlies infrared sensation in vampire bats. Nature 2011, 476, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Peralvarez-Marin, A.; Donate-Macian, P.; Gaudet, R. What do we know about the transient receptor potential vanilloid 2 (TRPV2) ion channel? FEBS J. 2013, 280, 5471–5487. [Google Scholar] [CrossRef] [PubMed]
- Sharif-Naeini, R.; Dedman, A.; Folgering, J.H.; Duprat, F.; Patel, A.; Nilius, B.; Honore, E. TRP channels and mechanosensory transduction: Insights into the arterial myogenic response. Pflug. Arch. 2008, 456, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, P.B.; Solursh, M.; Reiter, R.S. Stage-related capacity for limb chondrogenesis in cell culture. Dev. Biol. 1977, 60, 69–82. [Google Scholar] [CrossRef]
- San Antonio, J.D.; Tuan, R.S. Chondrogenesis of limb bud mesenchyme in vitro: Stimulation by cations. Dev. Biol. 1986, 115, 313–324. [Google Scholar] [CrossRef]
- Jordt, S.E.; Julius, D. Molecular basis for species-specific sensitivity to “hot” chili peppers. Cell 2002, 108, 421–430. [Google Scholar] [CrossRef]
- Wach, J.; Marin-Burgin, A.; Klusch, A.; Forster, C.; Engert, S.; Schwab, A.; Petersen, M. Low-threshold heat receptor in chick sensory neurons is upregulated independently of nerve growth factor after nerve injury. Neuroscience 2003, 117, 513–519. [Google Scholar] [CrossRef]
- Xu, H.; Blair, N.T.; Clapham, D.E. Camphor activates and strongly desensitizes the transient receptor potential vanilloid subtype 1 channel in a vanilloid-independent mechanism. J. Neurosci. 2005, 25, 8924–8937. [Google Scholar] [CrossRef] [PubMed]
- Donate-Macian, P.; Peralvarez-Marin, A. Dissecting domain-specific evolutionary pressure profiles of transient receptor potential vanilloid subfamily members 1 to 4. PLoS ONE 2014, 9, e110715. [Google Scholar] [CrossRef] [PubMed]
- Rutter, A.R.; Ma, Q.P.; Leveridge, M.; Bonnert, T.P. Heteromerization and colocalization of TRPV1 and TRPV2 in mammalian cell lines and rat dorsal root ganglia. Neuroreport 2005, 16, 1735–1739. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, N.; Albrecht, N.; Harteneck, C.; Schultz, G.; Schaefer, M. Homo- and heteromeric assembly of trpv channel subunits. J. Cell Sci. 2005, 118, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Ma, X.; Shen, B.; Huang, Y.; Birnbaumer, L.; Yao, X. TRPV4, TRPC1, and TRPP2 assemble to form a flow-sensitive heteromeric channel. FASEB J. 2014, 28, 4677–4685. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.D.; Gunthorpe, M.J.; Kelsell, R.E.; Hayes, P.D.; Reilly, P.; Facer, P.; Wright, J.E.; Jerman, J.C.; Walhin, J.P.; Ooi, L.; et al. TRPV3 is a Temperature-sensitive vanilloid receptor-like protein. Nature 2002, 418, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Cameron, T.L.; Belluoccio, D.; Farlie, P.G.; Brachvogel, B.; Bateman, J.F. Global comparative transcriptome analysis of cartilage formation in vivo. BMC Dev. Biol. 2009, 9, 20. [Google Scholar] [CrossRef] [PubMed]
- Camacho, N.; Krakow, D.; Johnykutty, S.; Katzman, P.J.; Pepkowitz, S.; Vriens, J.; Nilius, B.; Boyce, B.F.; Cohn, D.H. Dominant TRPV4 mutations in nonlethal and lethal metatropic dysplasia. Am. J. Med. Genet. A 2010, 152A, 1169–1177. [Google Scholar] [CrossRef] [PubMed]
- Lamande, S.R.; Yuan, Y.; Gresshoff, I.L.; Rowley, L.; Belluoccio, D.; Kaluarachchi, K.; Little, C.B.; Botzenhart, E.; Zerres, K.; Amor, D.J.; et al. Mutations in TRPV4 cause an inherited arthropathy of hands and feet. Nat. Genet. 2011, 43, 1142–1146. [Google Scholar] [CrossRef] [PubMed]
- Phan, M.N.; Leddy, H.A.; Votta, B.J.; Kumar, S.; Levy, D.S.; Lipshutz, D.B.; Lee, S.H.; Liedtke, W.; Guilak, F. Functional characterization of TRPV4 as an osmotically sensitive ion channel in porcine articular chondrocytes. Arthritis Rheum. 2009, 60, 3028–3037. [Google Scholar] [CrossRef] [PubMed]
- Shibasaki, K.; Murayama, N.; Ono, K.; Ishizaki, Y.; Tominaga, M. TRPV2 enhances axon outgrowth through its activation by membrane stretch in developing sensory and motor neurons. J. Neurosci. 2010, 30, 4601–4612. [Google Scholar] [CrossRef] [PubMed]
- Morelli, M.B.; Amantini, C.; Liberati, S.; Santoni, M.; Nabissi, M. TRP channels: New potential therapeutic approaches in cns neuropathies. CNS Neurol. Disord. Drug Targets 2013, 12, 274–293. [Google Scholar] [CrossRef] [PubMed]
- Levine, J.D.; Alessandri-Haber, N. TRP channels: Targets for the relief of pain. Biochim. Biophys. Acta 2007, 1772, 989–1003. [Google Scholar] [CrossRef] [PubMed]
- Numazaki, M.; Tominaga, M. Nociception and TRP channels. Curr. Drug Targets CNS Neurol. Disord. 2004, 3, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Gautier, M.; Dhennin-Duthille, I.; Ay, A.S.; Rybarczyk, P.; Korichneva, I.; Ouadid-Ahidouch, H. New insights into pharmacological tools to TR(I)P cancer up. Br. J. Pharmacol. 2014, 171, 2582–2592. [Google Scholar] [CrossRef] [PubMed]
- Waning, J.; Vriens, J.; Owsianik, G.; Stuwe, L.; Mally, S.; Fabian, A.; Frippiat, C.; Nilius, B.; Schwab, A. A Novel function of capsaicin-sensitive TRPV1 channels: Involvement in cell migration. Cell Calcium 2007, 42, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.; Dahan, D.; Cardouat, G.; Gillibert-Duplantier, J.; Marthan, R.; Savineau, J.P.; Ducret, T. Involvement of TRPV1 and TRPV4 channels in migration of rat pulmonary arterial smooth muscle cells. Pflug. Arch. 2012, 464, 261–272. [Google Scholar] [CrossRef] [PubMed]
- El Andaloussi-Lilja, J.; Lundqvist, J.; Forsby, A. TRPV1 expression and activity during retinoic acid-induced neuronal differentiation. Neurochem. Int. 2009, 55, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Morgan, P.J.; Hubner, R.; Rolfs, A.; Frech, M.J. Spontaneous calcium transients in human neural progenitor cells mediated by transient receptor potential channels. Stem Cells Dev. 2013, 22, 2477–2486. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Hu, H.Z.; Colton, C.K.; Wood, J.D.; Zhu, M.X. An alternative splicing product of the murine TRPV1 gene dominant negatively modulates the activity of TRPV1 channels. J. Biol. Chem. 2004, 279, 37423–37430. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Fu, Y.; Wang, D.H.; Cohen, D.M. Regulation of TRPV1 by a novel renally expressed rat TRPV1 splice variant. Am. J. Physiol. Ren. Physiol. 2006, 290, F117–F126. [Google Scholar] [CrossRef] [PubMed]
- Eilers, H.; Lee, S.Y.; Hau, C.W.; Logvinova, A.; Schumacher, M.A. The rat vanilloid receptor splice variant VR.5′sv blocks TRPV1 activation. Neuroreport 2007, 18, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.A.; Eilers, H. TRPV1 splice variants: Structure and function. Front. Biosci. (Landmark Ed.) 2010, 15, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Mistry, S.; Paule, C.C.; Varga, A.; Photiou, A.; Jenes, A.; Avelino, A.; Buluwela, L.; Nagy, I. Prolonged exposure to bradykinin and prostaglandin E2 increases TRPV1 mRNA but does not alter TRPV1 and TRPV1b protein expression in cultured rat primary sensory neurons. Neurosci. Lett. 2014, 564, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Kedei, N.; Szabo, T.; Lile, J.D.; Treanor, J.J.; Olah, Z.; Iadarola, M.J.; Blumberg, P.M. Analysis of the native quaternary structure of vanilloid receptor 1. J. Biol. Chem. 2001, 276, 28613–28619. [Google Scholar] [CrossRef] [PubMed]
- Veldhuis, N.A.; Lew, M.J.; Abogadie, F.C.; Poole, D.P.; Jennings, E.A.; Ivanusic, J.J.; Eilers, H.; Bunnett, N.W.; McIntyre, P. N-Glycosylation determines ionic permeability and desensitization of the TRPV1 capsaicin receptor. J. Biol. Chem. 2012, 287, 21765–21772. [Google Scholar] [CrossRef] [PubMed]
- Winter, Z.; Buhala, A.; Otvos, F.; Josvay, K.; Vizler, C.; Dombi, G.; Szakonyi, G.; Olah, Z. Functionally important amino acid residues in the transient receptor potential vanilloid 1 (TRPV1) ion channel—An overview of the current mutational data. Mol. Pain 2013, 9, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arniges, M.; Fernandez-Fernandez, J.M.; Albrecht, N.; Schaefer, M.; Valverde, M.A. Human TRPV4 channel splice variants revealed a key role of ankyrin domains in multimerization and trafficking. J. Biol. Chem. 2006, 281, 1580–1586. [Google Scholar] [CrossRef] [PubMed]
- Hammond, C.L.; Simbi, B.H.; Stickland, N.C. In ovo temperature manipulation influences embryonic motility and growth of limb tissues in the chick (Gallus gallus). J. Exp. Biol. 2007, 210, 2667–2675. [Google Scholar] [CrossRef] [PubMed]
- Serrat, M.A.; King, D.; Lovejoy, C.O. Temperature regulates limb length in homeotherms by directly modulating cartilage growth. Proc. Natl. Acad. Sci. USA 2008, 105, 19348–19353. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, C.; Wang, S. Periodic heat shock accelerated the chondrogenic differentiation of human mesenchymal stem cells in pellet culture. PLoS ONE 2014, 9, e91561. [Google Scholar] [CrossRef] [PubMed]
- Hojo, T.; Fujioka, M.; Otsuka, G.; Inoue, S.; Kim, U.; Kubo, T. Effect of heat stimulation on viability and proteoglycan metabolism of cultured chondrocytes: Preliminary report. J. Orthop. Sci. 2003, 8, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.A.; Tonomura, H.; Arai, Y.; Terauchi, R.; Honjo, K.; Hiraoka, N.; Hojo, T.; Kunitomo, T.; Kubo, T. Hyperthermia for the treatment of articular cartilage with osteoarthritis. Int. J. Hyperth. 2009, 25, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Kopakkala-Tani, M.; Leskinen, J.J.; Karjalainen, H.M.; Karjalainen, T.; Hynynen, K.; Toyras, J.; Jurvelin, J.S.; Lammi, M.J. Ultrasound stimulates proteoglycan synthesis in bovine primary chondrocytes. Biorheology 2006, 43, 271–282. [Google Scholar] [PubMed]
- Fan-xin, M.; Li-mei, S.; Bei, S.; Xin, Q.; Yu, Y.; Yu, C. Heat shock factor 1 regulates the expression of the TRPV1 gene in the rat preoptic-anterior hypothalamus area during lipopolysaccharide-induced fever. Exp. Physiol. 2012, 97, 730–740. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Biro, T. TRPV3: A more than skinny channel. Exp. Dermatol. 2013, 22, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Morelli, M.B.; Nabissi, M.; Amantini, C.; Farfariello, V.; Ricci-Vitiani, L.; di Martino, S.; Pallini, R.; Larocca, L.M.; Caprodossi, S.; Santoni, M.; et al. The transient receptor potential vanilloid-2 cation channel impairs glioblastoma stem-like cell proliferation and promotes differentiation. Int. J. Cancer 2012, 131, E1067–E1077. [Google Scholar] [CrossRef] [PubMed]
- Hatano, N.; Suzuki, H.; Itoh, Y.; Muraki, K. TRPV4 partially participates in proliferation of human brain capillary endothelial cells. Life Sci. 2013, 92, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.M.; Kuang, H.Y.; Zhang, L.X.; Wu, J.Z.; Chen, X.L.; Zhang, X.Y.; Ma, L.J. Effect of TRPV1 channel on proliferation and apoptosis of airway smooth muscle cells of rats. J. Huazhong Univ. Sci. Technol. Med. Sci. 2014, 34, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Stock, K.; Garthe, A.; de Almeida Sassi, F.; Glass, R.; Wolf, S.A.; Kettenmann, H. The capsaicin receptor TRPV1 as a novel modulator of neural precursor cell proliferation. Stem Cells 2014, 32, 3183–3195. [Google Scholar] [CrossRef] [PubMed]
- Ho, K.W.; Ward, N.J.; Calkins, D.J. TRPV1: A stress response protein in the central nervous system. Am. J. Neurodegener. Dis. 2012, 1, 1–14. [Google Scholar] [PubMed]
- Lamb, K.J.; Lewthwaite, J.C.; Bastow, E.R.; Pitsillides, A.A. Defining boundaries during joint cavity formation: Going out on a limb. Int. J. Exp. Pathol. 2003, 84, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Guilak, F. Biomechanical factors in osteoarthritis. Best Pract. Res. Clin. Rheumatol. 2011, 25, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Han, S.K.; Wouters, W.; Clark, A.; Herzog, W. Mechanically induced calcium signaling in chondrocytes in situ. J. Orthop. Res. 2012, 30, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Plant, T.D. TRPs in mechanosensing and volume regulation. Handb. Exp. Pharmacol. 2014, 223, 743–766. [Google Scholar] [PubMed]
- Clark, A.L.; Votta, B.J.; Kumar, S.; Liedtke, W.; Guilak, F. Chondroprotective role of the osmotically sensitive ion channel transient receptor potential vanilloid 4: Age- and sex-dependent progression of osteoarthritis in TRPV4-deficient mice. Arthritis Rheum. 2010, 62, 2973–2983. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Cai, X.; Wang, J.; Tang, H.; Yuan, Q.; Gong, P.; Lin, Y. Mechanical stretch inhibits adipogenesis and stimulates osteogenesis of adipose stem cells. Cell Prolif. 2012, 45, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Liang, L.; Dou, Y.; Huang, Z.; Mo, H.; Wang, Y.; Yu, B. Mechanical strain regulates osteogenic and adipogenic differentiation of bone marrow mesenchymal stem cells. Biomed. Res. Int. 2015, 2015, 873251. [Google Scholar] [CrossRef] [PubMed]
- Cheung, S.Y.; Huang, Y.; Kwan, H.Y.; Chung, H.Y.; Yao, X. Activation of transient receptor potential vanilloid 3 channel suppresses adipogenesis. Endocrinology 2015, 156, 2074–2086. [Google Scholar] [CrossRef] [PubMed]
- Takacs, R.; Matta, C.; Somogyi, C.; Juhasz, T.; Zakany, R. Comparative analysis of osteogenic/chondrogenic differentiation potential in primary limb bud-derived and c3h10t1/2 cell line-based mouse micromass cultures. Int. J. Mol. Sci. 2013, 14, 16141–16167. [Google Scholar] [CrossRef] [PubMed]
- Juhász, T.; Matta, C.; Mészár, Z.; Nagy, G.; Szíjgyártó, Z.; Molnár, Z.; Kolozsvári, B.; Bakó, E.; Zákány, R. Optimalized transient transfection of chondrogenic primary cell cultures. Cent. Eur. J. Biol. 2010, 5, 572–584. [Google Scholar] [CrossRef]
- Niswander, L.; Martin, G.R. FGF-4 regulates expression of EVX-1 in the developing mouse limb. Development 1993, 119, 287–294. [Google Scholar] [PubMed]
- Vogel, A.; Tickle, C. FGF-4 Maintains polarizing activity of posterior limb bud cells in vivo and in vitro. Development 1993, 119, 199–206. [Google Scholar] [PubMed]
- Varga, Z.; Juhasz, T.; Matta, C.; Fodor, J.; Katona, E.; Bartok, A.; Olah, T.; Sebe, A.; Csernoch, L.; Panyi, G.; et al. Switch of voltage-gated K+ channel expression in the plasma membrane of chondrogenic cells affects cytosolic Ca2+-oscillations and cartilage formation. PLoS ONE 2011, 6, e27957. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Somogyi, C.S.; Matta, C.; Foldvari, Z.; Juhász, T.; Katona, É.; Takács, Á.R.; Hajdú, T.; Dobrosi, N.; Gergely, P.; Zákány, R. Polymodal Transient Receptor Potential Vanilloid (TRPV) Ion Channels in Chondrogenic Cells. Int. J. Mol. Sci. 2015, 16, 18412-18438. https://doi.org/10.3390/ijms160818412
Somogyi CS, Matta C, Foldvari Z, Juhász T, Katona É, Takács ÁR, Hajdú T, Dobrosi N, Gergely P, Zákány R. Polymodal Transient Receptor Potential Vanilloid (TRPV) Ion Channels in Chondrogenic Cells. International Journal of Molecular Sciences. 2015; 16(8):18412-18438. https://doi.org/10.3390/ijms160818412
Chicago/Turabian StyleSomogyi, Csilla Szűcs, Csaba Matta, Zsofia Foldvari, Tamás Juhász, Éva Katona, Ádám Roland Takács, Tibor Hajdú, Nóra Dobrosi, Pál Gergely, and Róza Zákány. 2015. "Polymodal Transient Receptor Potential Vanilloid (TRPV) Ion Channels in Chondrogenic Cells" International Journal of Molecular Sciences 16, no. 8: 18412-18438. https://doi.org/10.3390/ijms160818412