
Morphogenesis, Flowering, and Gene Expression of Dendranthema grandiflorum in Response to Shift in Light Quality of Night Interruption



Abstract
:1. Introduction
2. Results
2.1. Morphogenesis
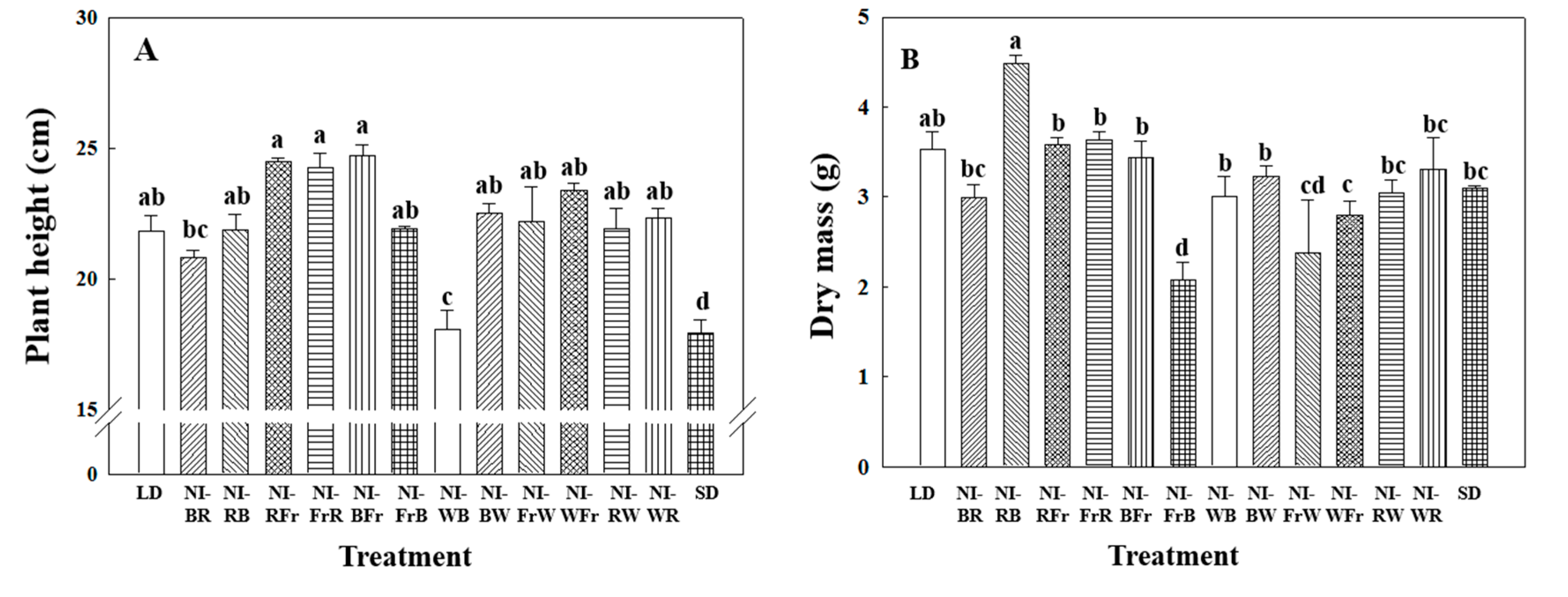
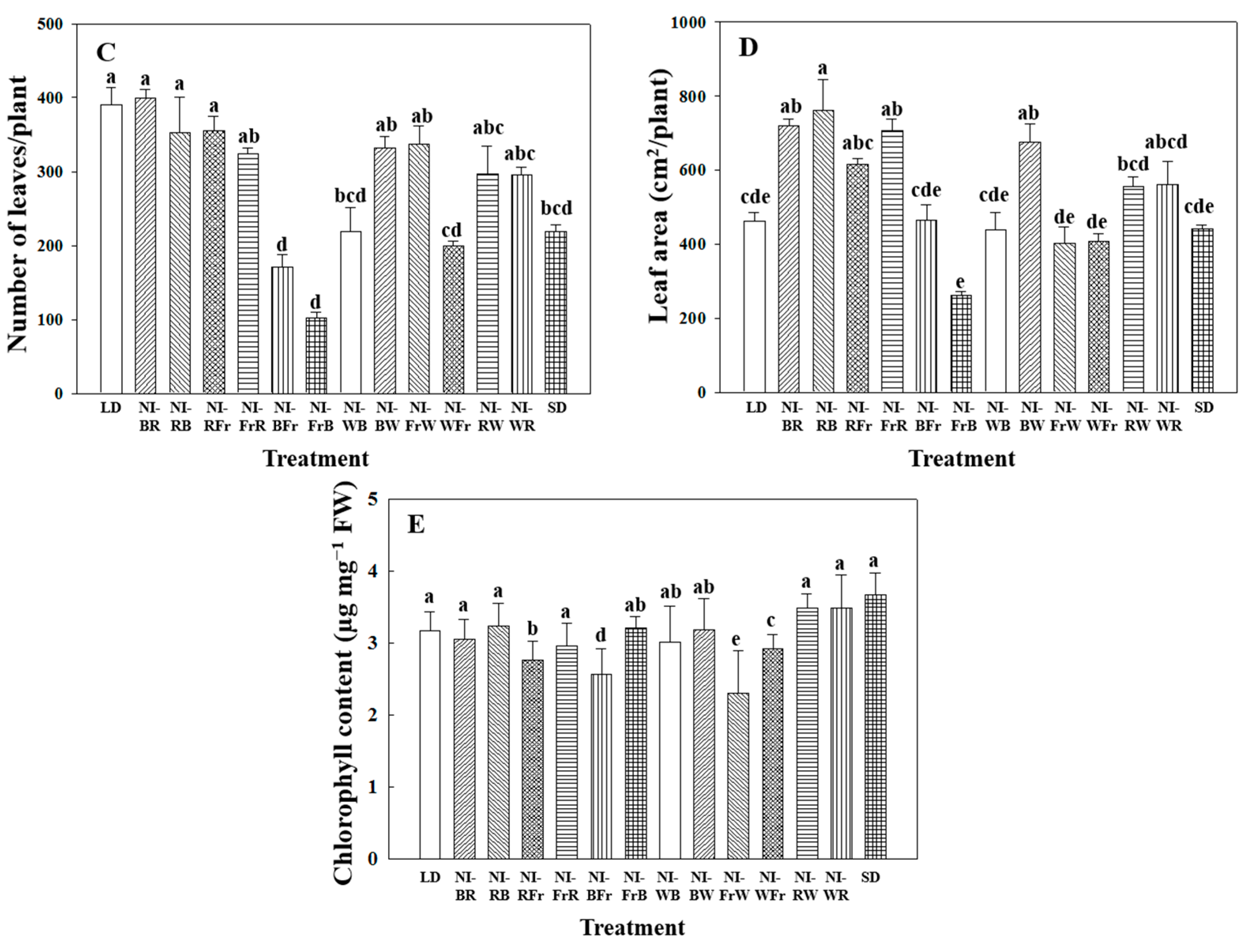
2.2. Flowering



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | DVB y (Day) | Number of Flowers/Plant |
|---|---|---|
| LD | – z | – |
| NI-BR | – | – |
| NI-RB | 44.0 | 0.3 e w |
| NI-RFr | – | – |
| NI-FrR | 44.0 | – |
| NI-BFr | 14.8 | 27.3 a w |
| NI-FrB | 12.2 | 20.0 b w |
| NI-WB | 26.4 | 9.3 d w |
| NI-BW | – | – |
| NI-FrW | 33.7 | 12.3 c w |
| NI-WFr | 17.6 | 15.0 c w |
| NI-RW | – | – |
| NI-WR | 41.6 | 5.3 e w |
| SD | 13.6 | 20.6 b w |
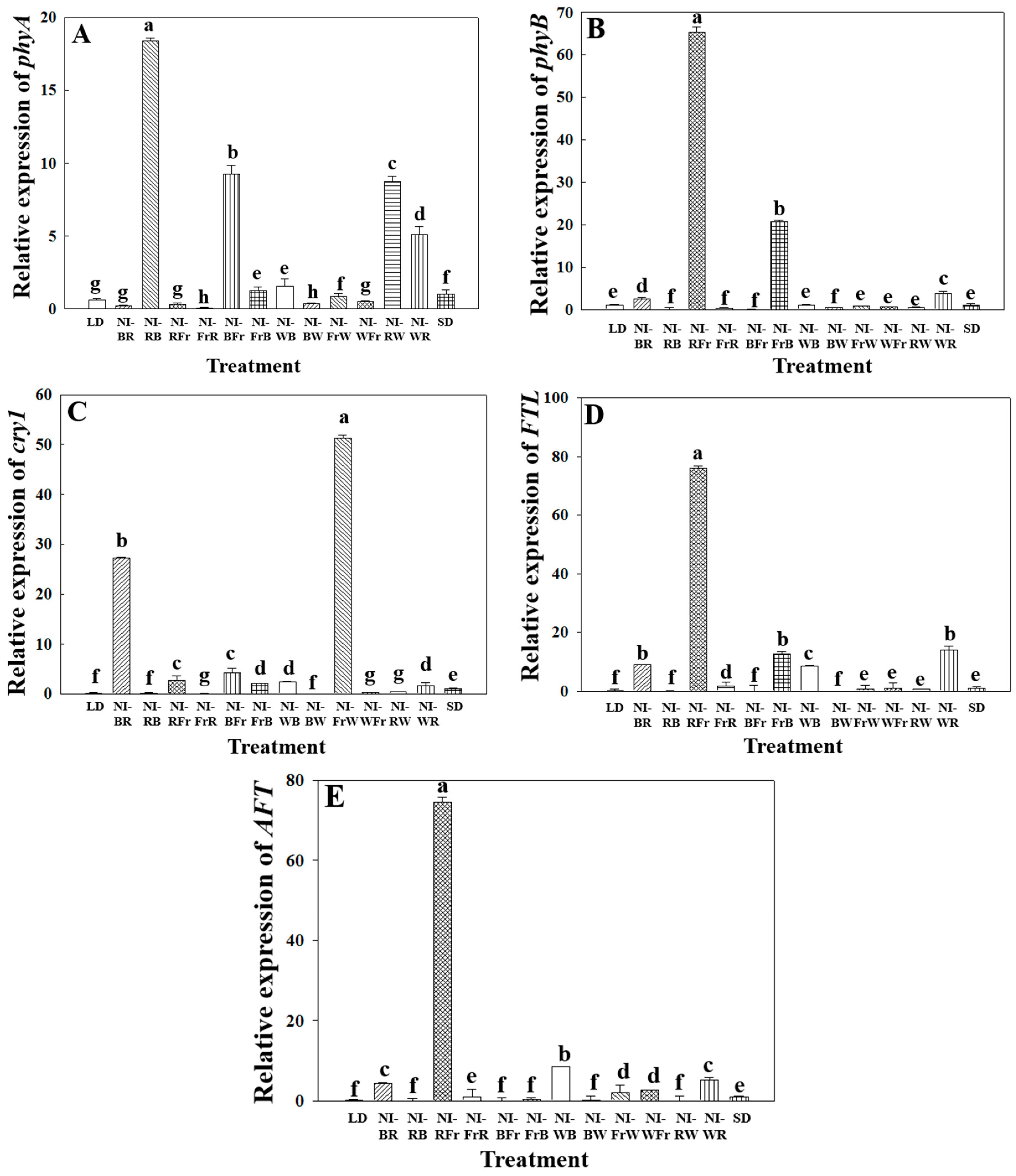
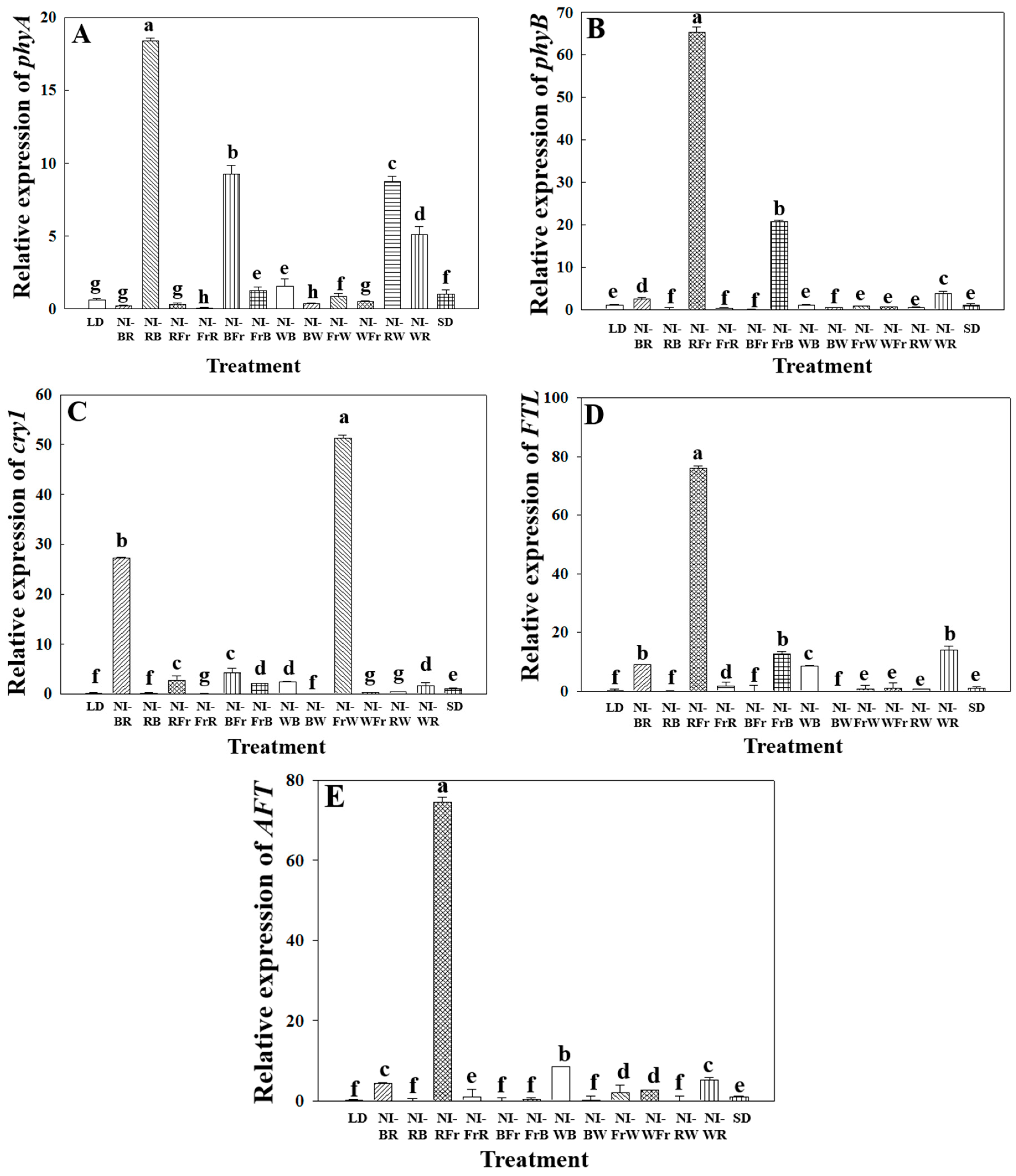
2.3. Photoreceptor Gene Expression Analysis
3. Discussion
3.1. Morphogenesis
3.2. Flowering
3.3. Photoreceptor Gene Expression Analysis
4. Experimental Section
4.1. Plant Material and Growth Conditions
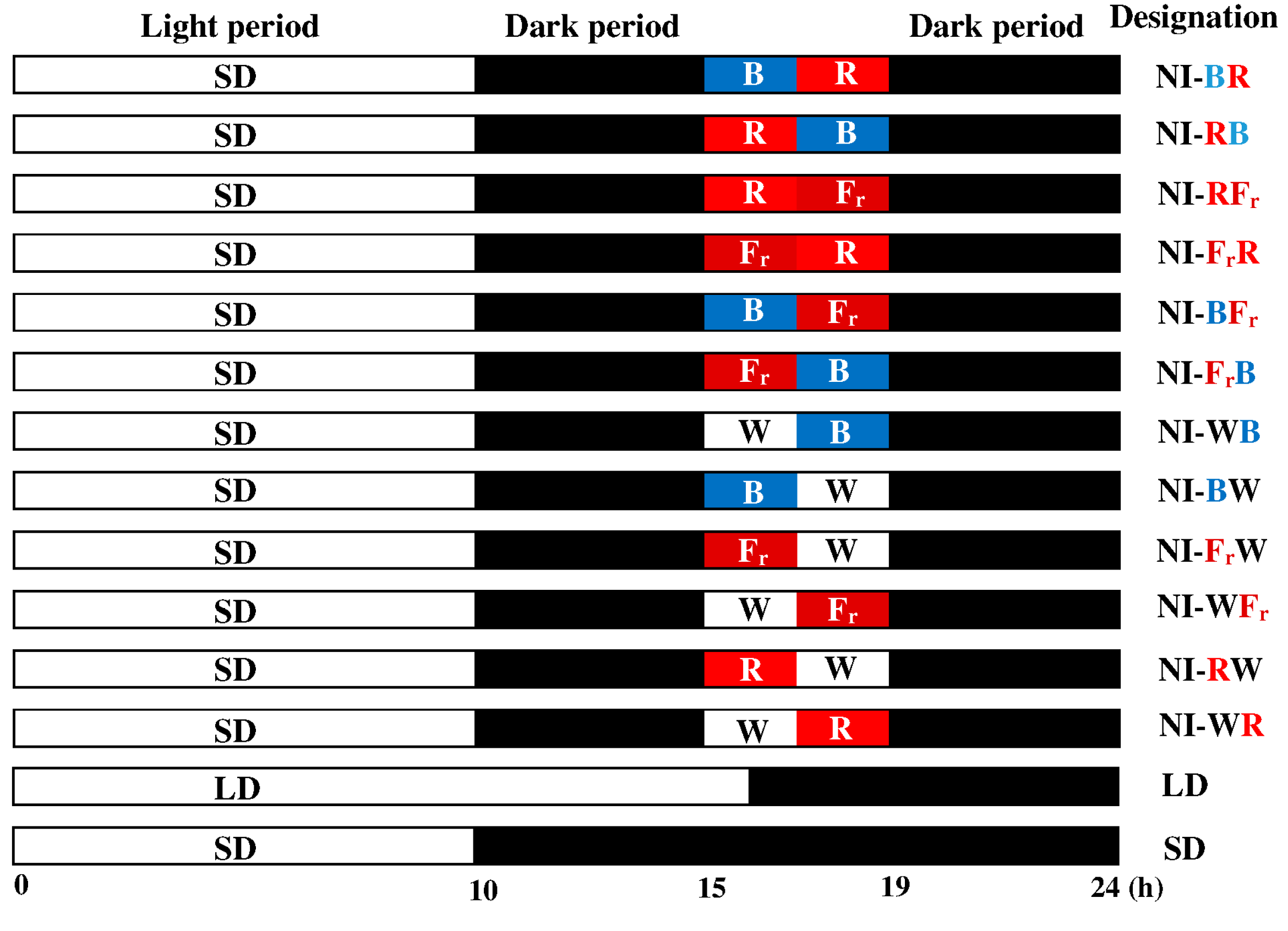
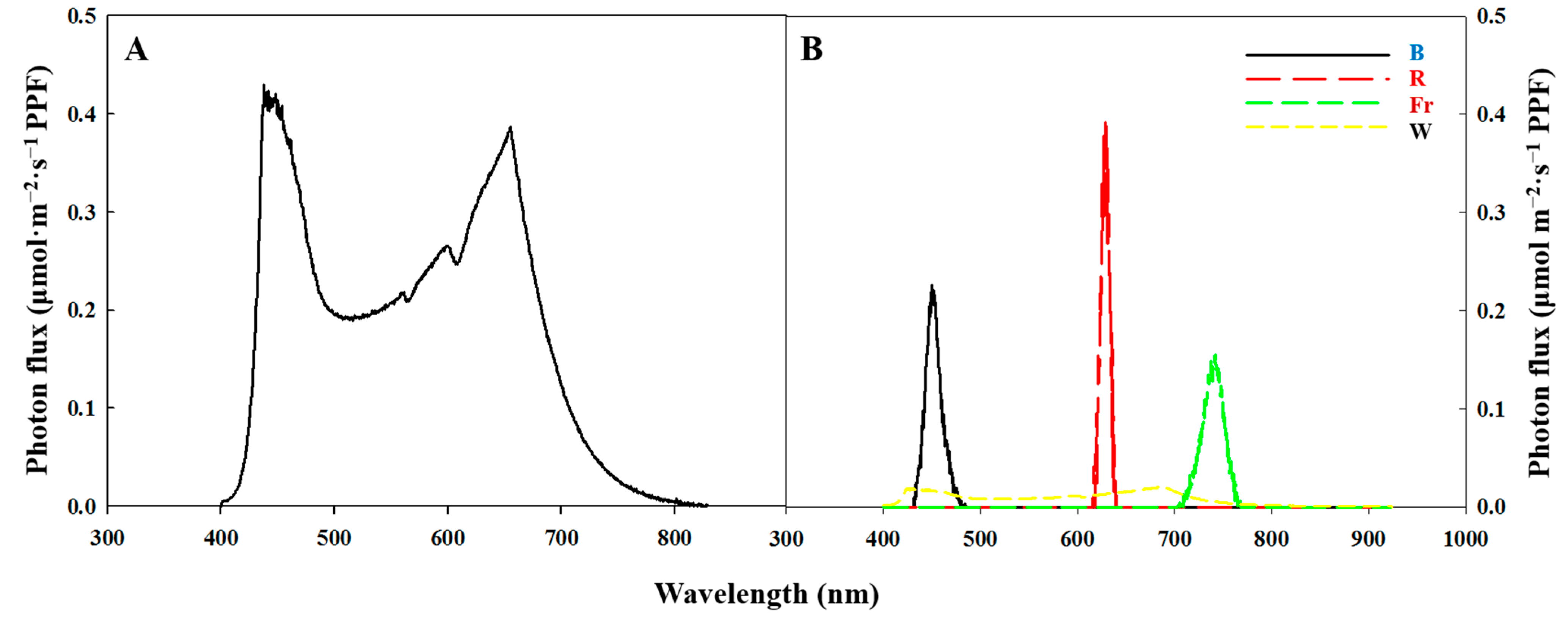
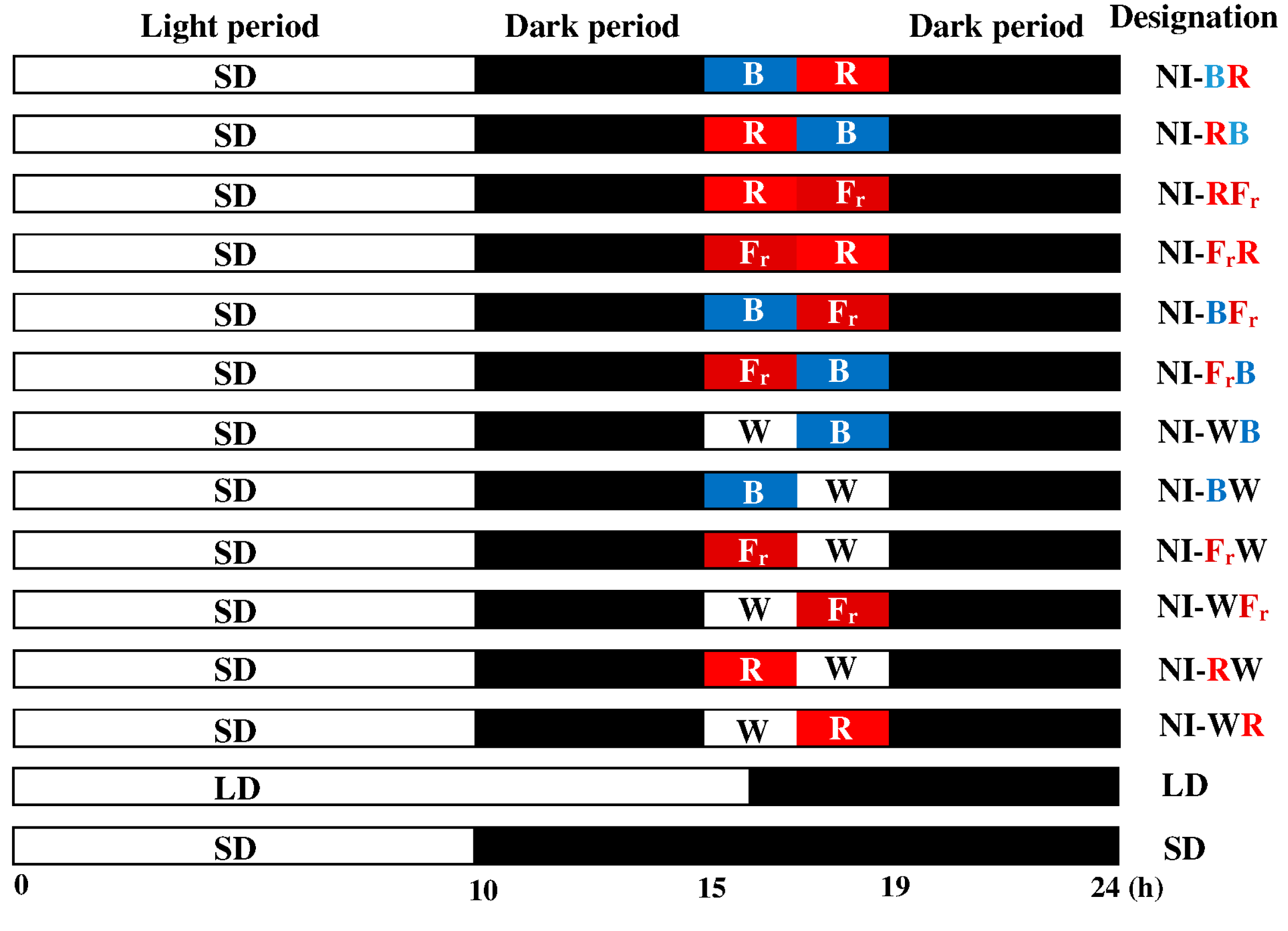
4.2. Photoperiodic Light Treatments
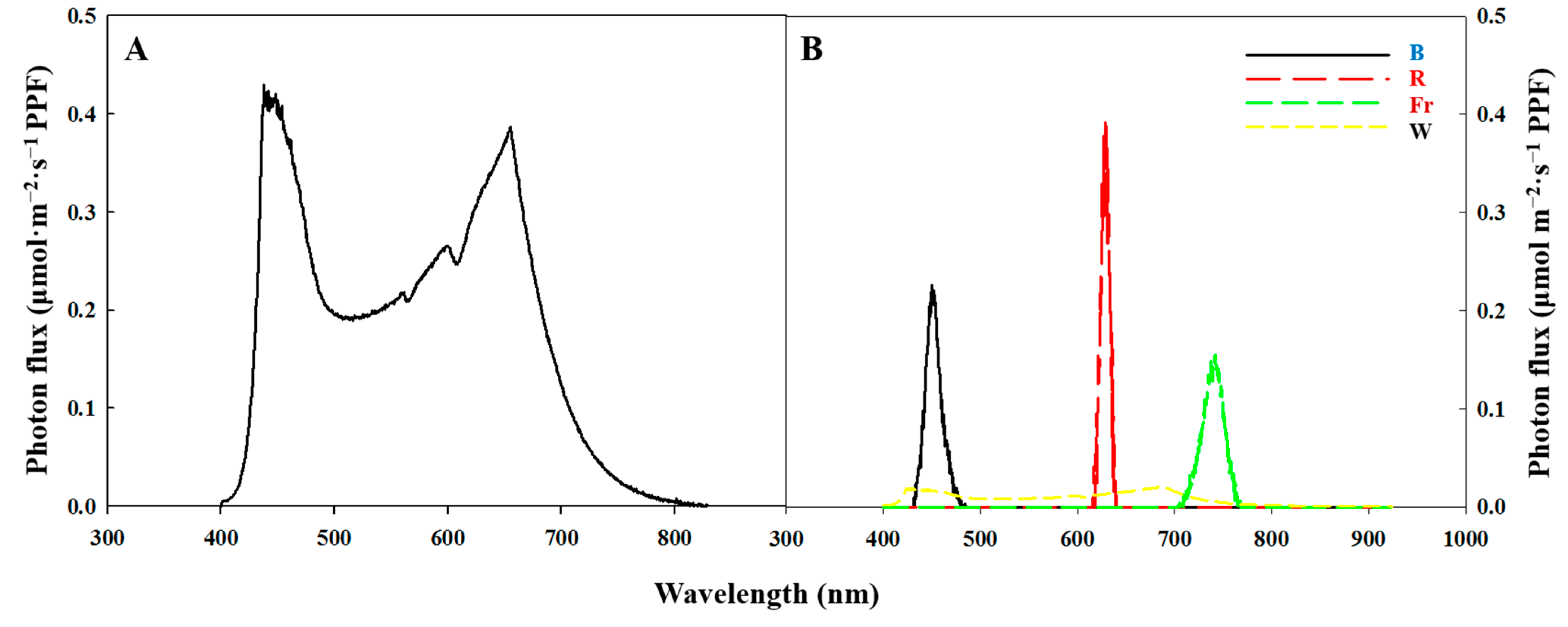
4.3. Data Collection and Analysis
4.4. Total RNA Isolation, cDNA Synthesis, and Real-Time Polymerase Chain Reaction (PCR) of Selected Genes
| Gene | Accession Number | Forward Primer | Reverse Primer |
|---|---|---|---|
| Ch-phyA | EU915082 | 5′-GACAGTGTCAGGCTTCAACAAG-3′ | 5′-ACCACCAGTGTGTGTTATCCTG-3′ |
| Ch-phyB | NM_127435 | 5′-GTGCTAGGGAGATTACGCTTTC-3′ | 5′-CCAGCTTCTGAGACTGAACAGA-3′ |
| Ch-cry1 | NM_116961 | 5′-CGTAAGGGATCACCGAGTAAAG-3′ | 5′-CTTTTAGGTGGGAGTTGTGGAG-3′ |
| Ch-FTL | AB839767 | 5′-ACAACGGACTCCTCATTTGG-3′ | 5′-CGCGAAACTACGAGTGTTGA-3′ |
| Ch-AFT | AB839766 | 5′-AGAACACCTCCATTGGATCG-3′ | 5′-CTGGAACTAGGTGGCCTCAC-3′ |
| Ch-Actin | AB205087 | 5′-CGTTTGGATCTTGCTGGTCG-3′ | 5′-CAGGACATCTGAAACGCTCA-3′ |
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Casal, J.J.; Fankhauser, C.; Coupland, G.; Blázquez, M.A. Signalling for developmental plasticity. Trends Plant Sci. 2004, 9, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Kutschera, U.; Briggs, W.R. Seedling development in buckwheat and the discovery of the photomorphogenic shade-avoidance response. Plant Biol. 2013, 15, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Lumsden, P.J.; Millar, A.J. Biological Rhythms and Photoperiodism in Plants, 1st ed.; Bios Scientific Publishers: Oxford, UK, 1998. [Google Scholar]
- Somers, D.E.; Devlin, P.F.; Kay, S.A. Phytochromes and cryptochromes in the entertainment of the Arabidopsis circadian clock. Science 1998, 282, 1488–1490. [Google Scholar] [CrossRef] [PubMed]
- Somers, D.E.; Kim, W.Y.; Geng, R. The F-Box protein ZEITLUPE confers dosage-dependent control on the circadian clock, photomorphogenesis, and flowering time. Plant Cell 2004, 16, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.Y.; Hicks, K.A.; Somers, D.E. Independent roles for EARLY FLOWERING 3 and ZEITLUPE in the control of circadian timing, hypocotyl length, and flowering time. Plant Physiol. 2005, 139, 1557–1569. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, S.; Wang, L.; Han, L.; Suh, S.S.; Salome, P.A.; McClung, C.R.; Somers, D.E. Post-translational regulation of the Arabidopsis circadian clock through selective proteolysis and phosphorylation of pseudo-response regulator proteins. J. Biol. Chem. 2008, 34, 23073–23083. [Google Scholar] [CrossRef] [PubMed]
- Warner, R.M.; Erwin, J.E. Effect of photoperiod and daily light integral on flowering of five Hibiscus sp. Sci. Hortic. 2003, 97, 341–351. [Google Scholar] [CrossRef]
- Runkle, E.S.; Heins, R.D. Specific functions of red, far red, and blue light in flowering and stem extension of long-day plants. J. Am. Soc. Hortic. Sci. 2001, 126, 275–282. [Google Scholar]
- Casal, J.J.; Deregibus, V.A.; Sanchez, R.A. Variations in tiller dynamics and morphology in Lolium multiflorum Lam. vegetative and reproductive plants as affected by differences in red/far-red irradiation. Ann. Bot. 1985, 56, 553–559. [Google Scholar]
- Leopold, A.C. Photoperiodism in plants. Q. Rev. Biol. 1951, 26, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Lin, C. Photoreception and regulation of flowering time. Plant Physiol. 2000, 123, 139–150. [Google Scholar] [CrossRef]
- Blanchard, M.G.; Runkle, E.S. Intermittent light from a rotating high-pressure sodium lamp promotes flowering of long-day plants. HortScience 2010, 45, 236–241. [Google Scholar]
- Yamada, A.T.; Tanigawa, T.; Suyama, T.; Matsuno, T.; Kunitake, T. Night break treatment using different light sources promotes or delays growth and flowering of Eustoma grandiflorum (Raf.) Shinn. J. Jpn. Soc. Hortic. Sci. 2008, 77, 69–74. [Google Scholar] [CrossRef]
- Vince-Prue, D.; Canham, A.E. Horticultural significance of photomorphogenesis. In Encyclopedia of Plant Physiology; Shropshire, W., Mohr, H., Eds.; Springer-Verlag: Berlin, Germany, 1983; pp. 518–544. [Google Scholar]
- Kim, Y.J.; Lee, H.J.; Kim, K.S. Night interruption promotes vegetative growth and flowering of Cymbidium. Sci. Hortic. 2011, 130, 887–893. [Google Scholar] [CrossRef]
- Runkle, E.S.; Heins, R.D.; Cameron, A.C.; Carlson, W.H. Flowering of herbaceous perennials under various night interruption and cyclic lighting treatments. HortScience 1998, 33, 672–677. [Google Scholar]
- Ishikawa, R.; Tamaki, S.; Yokoi, S.; Inagaki, N.; Shinomura, T.; Takano, M.; Shimamoto, K. Suppression of the floral activator Hd3a is the principal cause of the night break effect in rice. Plant Cell 2005, 17, 3326–3336. [Google Scholar] [CrossRef] [PubMed]
- Goto, N.; Kumagai, T.; Koornneef, M. Flowering responses to light-breaks in photomorphogenic mutants of Arabidopsis thaliana, a long-day plant. Physiol. Plant. 2006, 83, 209–215. [Google Scholar] [CrossRef]
- Oh, W.; Rhie, Y.H.; Park, J.H.; Runkle, E.S.; Kim, K.S. Flowering of cyclamen is accelerated by an increase in temperature, photoperiod and daily light integral. J. Hortic. Sci. Biotechnol. 2008, 83, 559–562. [Google Scholar]
- Kim, Y.J.; Lee, H.J.; Kim, K.S. Carbohydrate changes in Cymbidium “Red Fire” in response to night interruption. Sci. Hortic. 2013, 162, 82–89. [Google Scholar] [CrossRef]
- Park, Y.J.; Kim, Y.J.; Kim, K.S. Vegetative growth and flowering of Dianthus, Zinnia, and Pelargonium as affected by night interruption at different timings. Hortic. Environ. Biotechnol. 2013, 54, 236–242. [Google Scholar] [CrossRef]
- Liao, Y.; Suzuki, K.; Yu, W.; Zhuang, D.; Takai, Y.; Ogasawara, R.; Shimazu, T.; Fukui, H. Night-break effect of LED light with different wavelengths on shoot elongation of Chrysanthemum morifolium Ramat “Jimba” and “Iwa no hakusen”. Environ. Control Biol. 2014, 52, 51–55. [Google Scholar] [CrossRef]
- Higuchi, Y.; Sumitomo, K.; Oda, A.; Shimizu, H.; Hisamatsu, T. Day light quality affects the night-break response in the short-day plant chrysanthemum, suggesting differential phytochrome-mediated regulation of flowering. J. Plant Physiol. 2012, 169, 1789–1796. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Runkle, E.S. Low-intensity blue light in night-interruption lighting does not influence flowering of herbaceous ornamentals. Sci. Hortic. 2015, 186, 230–238. [Google Scholar] [CrossRef]
- Downs, R.J.; Thomas, J.F. Phytochrome regulation of flowering in the long-day plant Hyoscyamus niger. Plant Physiol. 1982, 70, 898–900. [Google Scholar] [CrossRef] [PubMed]
- Lane, H.C.; Cathey, H.M.; Evans, L.T. Dependence of flowering in several long-day plants on spectral composition of light extending photoperiod. Am. J. Bot. 1965, 52, 1006–1014. [Google Scholar] [CrossRef]
- Cathey, H.M.; Borthwick, H.A. Photoreversibility of floral initiation in chrysanthemum. Bot. Gaz. 1957, 119, 71–76. [Google Scholar] [CrossRef]
- Craig, D.S.; Runkle, E.S. Using LEDs to quantify the effect of the red to far-red ratio of night interruption lighting on flowering of photoperiodic crops. Acta Hortic. 2012, 956, 179–185. [Google Scholar]
- Izawa, T.; Oikawa, T.; Sugiyama, N.; Tanisaka, T.; Yano, M.; Shimamoto, K. Phytochrome mediates the external light signal to repress FT orthologs in photoperiodic flowering of rice. Genes Dev. 2002, 16, 2006–2020. [Google Scholar] [CrossRef] [PubMed]
- Takano, M.; Inagaki, N.; Xie, X.; Kiyota, S.; Baba-Kasai, A.; Tanabata, T.; Shinomura, T. Phytochromes are the sole photoreceptors for perceiving red/far-red light in rice. Proc. Natl. Acad. Sci. USA 2009, 106, 14705–14710. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.G.; Muneer, S.; Jeong, B.R. Quality and positioning of night interruption light affects morphogenesis and flowering in chrysanthemum. Plant Growth Regul. 2015. manuscript in preparation. [Google Scholar]
- Folta, K.M.; Spalding, E.P. Unexpected roles for cryptochrome 2 and phototropin revealed by high-resolution analysis of blue light-mediated hypocotyl growth inhibition. Plant J. 2001, 26, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Devlin, P.F.; Patel, S.R.; Whitelam, G.C. Phytochrome E influences internode elongation and flowering time in Arabidopsis. Plant Cell 1998, 10, 1479–1488. [Google Scholar] [CrossRef] [PubMed]
- Keiller, D.; Smith, H. Control of carbon portioning by light quality mediated by phytochrome. Plant Sci. 1989, 63, 25–29. [Google Scholar] [CrossRef]
- Robson, P.R.H.; Whitelam, G.C.; Smith, H. Selected components of the shade-avoidance syndrome are displayed in a normal manner in mutants of Arabidopsis thaliana and Brassica rapa deficient in phytochrome B. Plant Physiol. 1993, 102, 1179–1184. [Google Scholar] [PubMed]
- Kang, K.J.; Oh, W.; Shin, J.H.; Kim, K.S. Night interruption and cyclic lighting promote flowering of Cyclamen persicum under low temperature regime. Hortic. Environ. Biotechnol. 2008, 49, 72–77. [Google Scholar]
- Runkle, E.S.; Heins, R.D. Photocontrol of flowering and extension growth in the long-day plant pansy. J. Am. Soc. Hortic. Sci. 2003, 128, 479–485. [Google Scholar]
- Smorenburg, K.; Courreges-Lacoste, G.B.; Berger, M.; Buschman, C.; del Bello, U.; Langsdorf, G.; Lichtenthaler, H.K.; Sioris, C.; Stoll, M.; Visser, H. Remote sensing of solar induced fluorescence of vegetation. Int. Soc. Opt. Photonics 2002, 4542, 178–190. [Google Scholar]
- Purohit, S.S.; Ranjan, R. Phytochrome and flowering. In Flowering, 1st ed.; Purohit, S.S., Ranjan, R., Eds.; Agrobios: Jodhpur, India, 2002; pp. 52–61. [Google Scholar]
- Smith, H.; Whitelam, G.C. The shade avoidance syndrome: Multiple responses mediated by multiple phytochromes. Plant Cell Environ. 1997, 20, 840–844. [Google Scholar] [CrossRef]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef] [PubMed]
- Vandenbussche, F.; Pierik, R.; Millenaar, F.F.; Voesenek, L.A.; van Der Straeten, D. Reaching out of the shade. Curr. Opin. Plant Biol. 2005, 8, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Hersch, M.; Lorrain, S.; Wit, M.; Trevisan, M.; Ljung, K.; Bergmann, S.; Fankhause, C. Light intensity modulates the regulatory network of the shade avoidance responses in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 6515–6520. [Google Scholar] [CrossRef] [PubMed]
- Fankhauser, C.; Ulm, R. Light-regulated interactions with SPA proteins underlie cryptochrome-mediated gene expression. Genes Dev. 2011, 2, 1004–1009. [Google Scholar] [CrossRef] [PubMed]
- Hirose, F.; Shinomura, T.; Tanabata, T.; Shimada, H.; Takano, M. Involvement of rice cryptochromes in de-etiolation responses and flowering. Plant Cell Physiol. 2006, 47, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.; Bradley, M.; Harberd, N.P.; Whitelam, G.C. Photoresponses of light-grown phyA mutants of Arabidopsis (phytochrome A is required for the perception of day length extensions). Plant Physiol. 1994, 105, 141–149. [Google Scholar] [PubMed]
- Mockler, T.; Yang, H.; Yu, W.; Parikh, D.; Cheng, Y.; Dolan, S.; Lin, C. Regulation of photoperiodic flowering by Arabidopsis photoreceptors. Proc. Natl. Acad. Sci. USA 2003, 100, 2140–2145. [Google Scholar] [CrossRef] [PubMed]
- Devlin, P.F.; Robson, P.R.; Patel, S.R.; Goosey, L.; Sharrock, R.A.; Whitelam, G.C. Phytochrome D acts in the shade avoidance syndrome in Arabidopsis by controlling elongation growth and flowering time. Plant Physiol. 1999, 119, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.A.; Praekelt, U.; Stoddart, W.M.; Billingham, O.E.; Halliday, K.J.; Whitelam, G.C. Phytochromes B, D, and E act redundantly to control multiple physiological responses in Arabidopsis. Plant Physiol. 2003, 131, 1340–1346. [Google Scholar] [CrossRef] [PubMed]
- Weller, J.L.; Beauchamp, N.; Kerckhoffs, L.H.J.; Platten, J.D.; Reid, J.B. Interaction of phytochromes A and B in the control of de-etiolation and flowering in pea. Plant J. 2001, 26, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Mockler, T.C.; Guo, H.; Yang, H.; Duong, H.; Lin, C. Antagonistic actions of Arabidopsis cryptochromes and phytochrome B in the regulation of floral induction. Development 1999, 126, 2073–2082. [Google Scholar] [PubMed]
- Guo, H.; Yang, H.; Mockler, T.C.; Lin, C. Regulation of flowering time by Arabidopsis photoreceptors. Science 1998, 279, 1360–1363. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, Y.; Narumi, T.; Oda, A.; Nakano, Y.; Sumitomo, K.; Fukai, S.; Hisamatsu, T. The gated induction system of a systemic floral inhibitor, antiflorigen, determines obligate short-day flowering in chrysanthemums. Proc. Natl. Acad. Sci. USA 2013, 110, 17137–17142. [Google Scholar] [CrossRef] [PubMed]
- Dere, S.; Gunes, T.; Sivaci, R. Spectrophotometric determination of chlorophyll—A, B and total carotenoid contents of some algae species using different solvents. Turk. J. Bot. 1998, 22, 13–17. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.G.; Muneer, S.; Jeong, B.R. Morphogenesis, Flowering, and Gene Expression of Dendranthema grandiflorum in Response to Shift in Light Quality of Night Interruption. Int. J. Mol. Sci. 2015, 16, 16497-16513. https://doi.org/10.3390/ijms160716497
Park YG, Muneer S, Jeong BR. Morphogenesis, Flowering, and Gene Expression of Dendranthema grandiflorum in Response to Shift in Light Quality of Night Interruption. International Journal of Molecular Sciences. 2015; 16(7):16497-16513. https://doi.org/10.3390/ijms160716497
Chicago/Turabian StylePark, Yoo Gyeong, Sowbiya Muneer, and Byoung Ryong Jeong. 2015. "Morphogenesis, Flowering, and Gene Expression of Dendranthema grandiflorum in Response to Shift in Light Quality of Night Interruption" International Journal of Molecular Sciences 16, no. 7: 16497-16513. https://doi.org/10.3390/ijms160716497