
Nitrogen Removal Characteristics of a Newly Isolated Indigenous Aerobic Denitrifier from Oligotrophic Drinking Water Reservoir, Zoogloea sp. N299

Abstract
:1. Introduction
2. Results
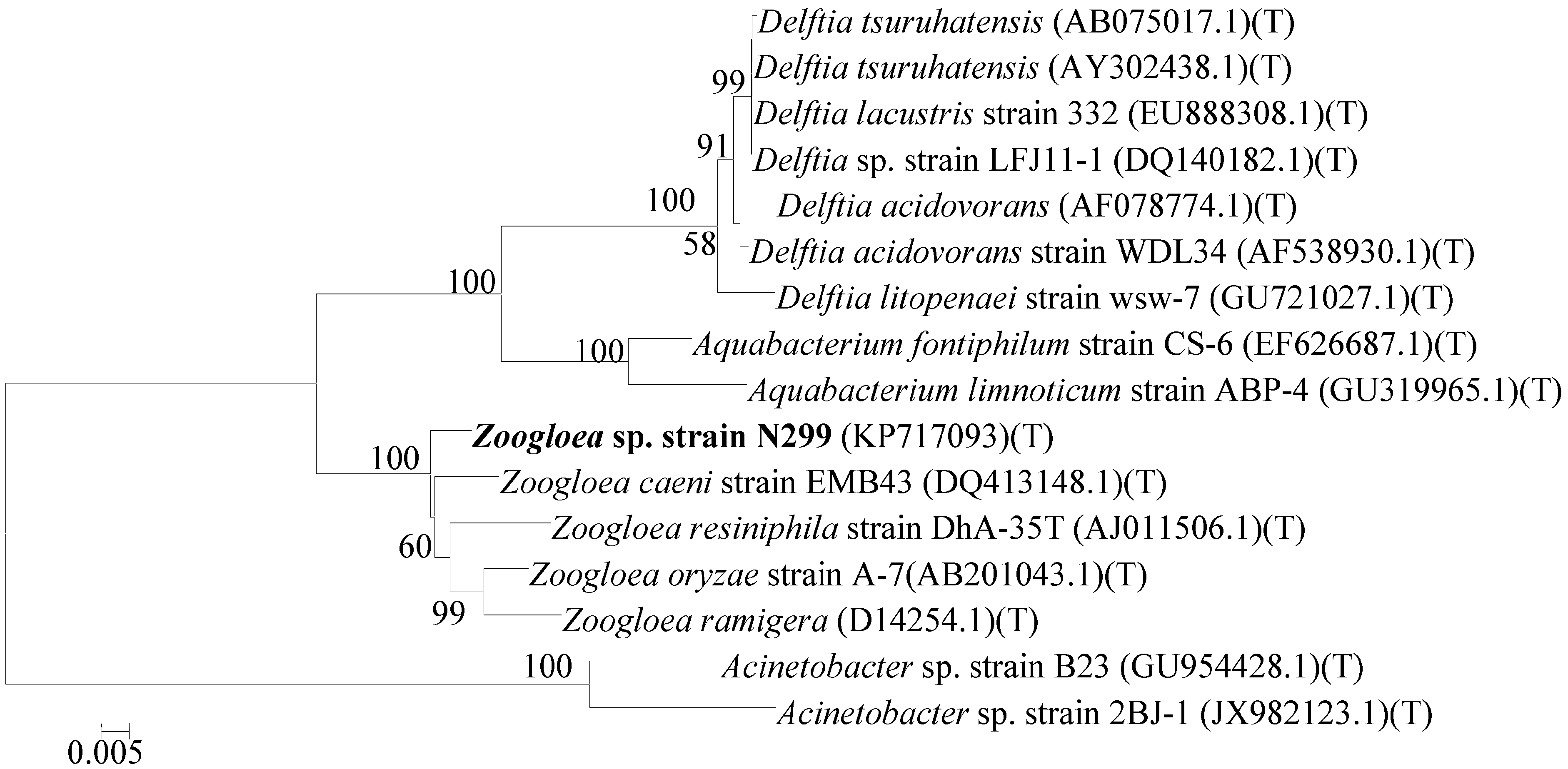
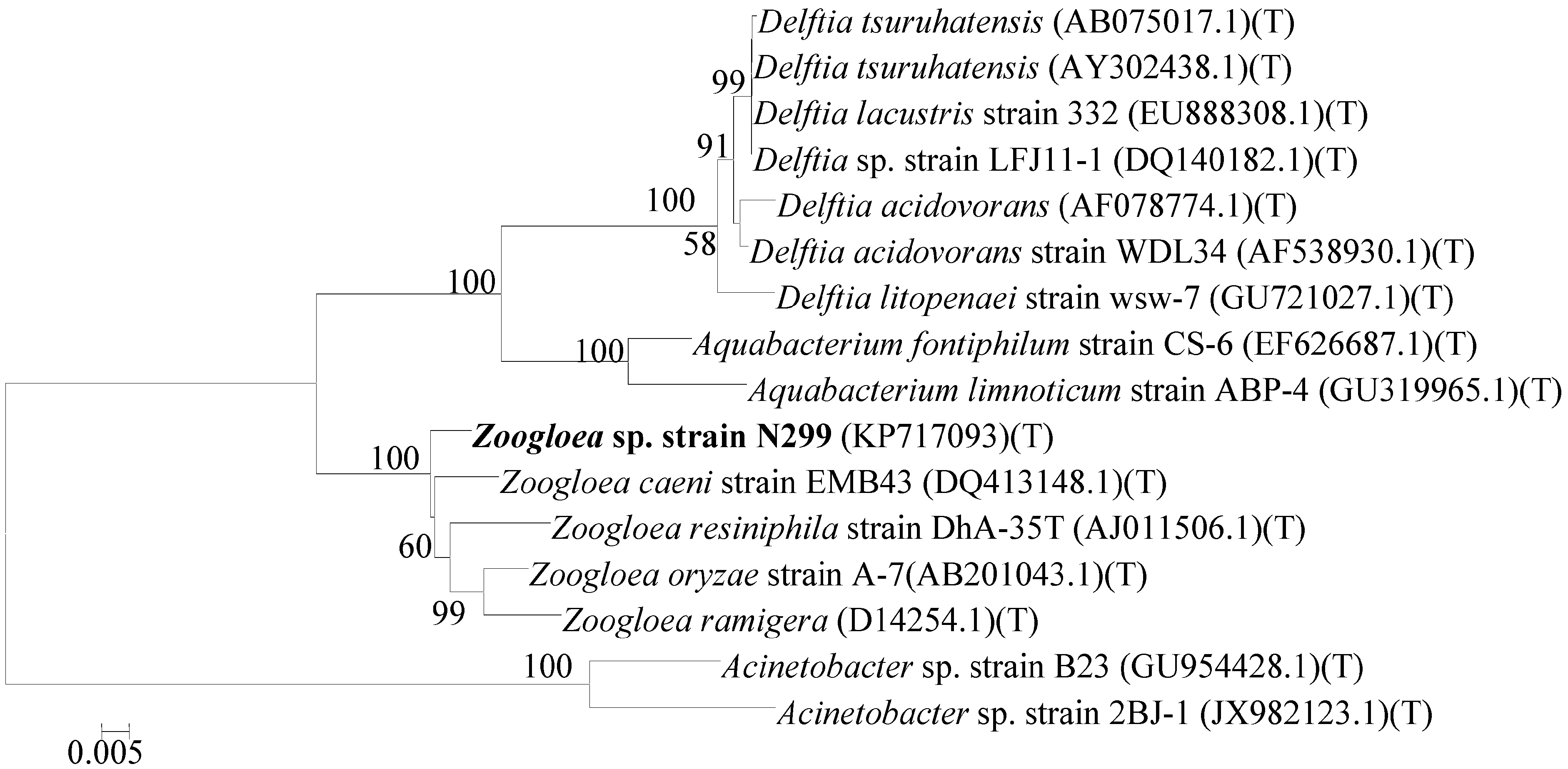
2.1. Identification and Phylogenetic Analysis of Aerobic Denitrifier N299


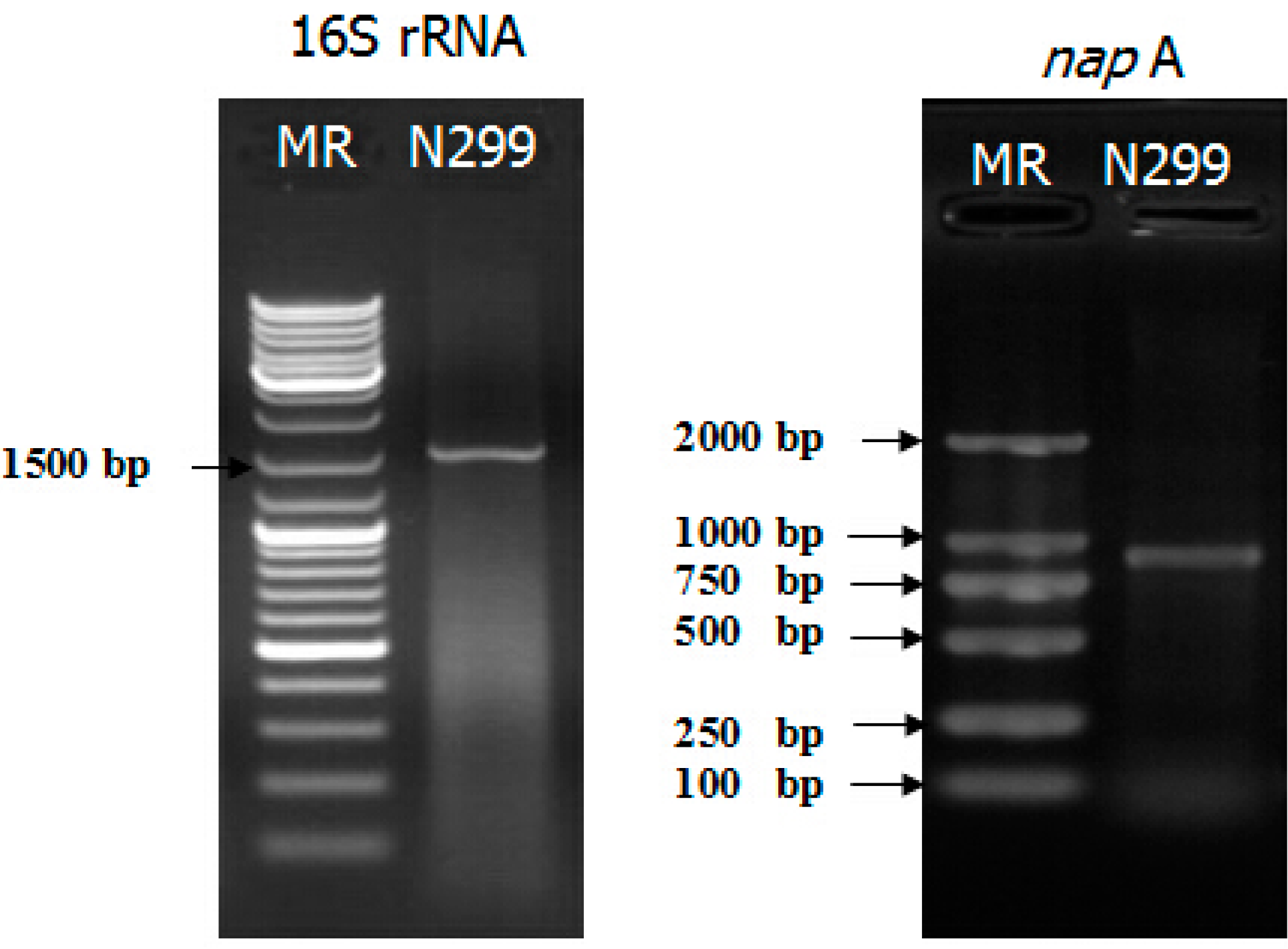
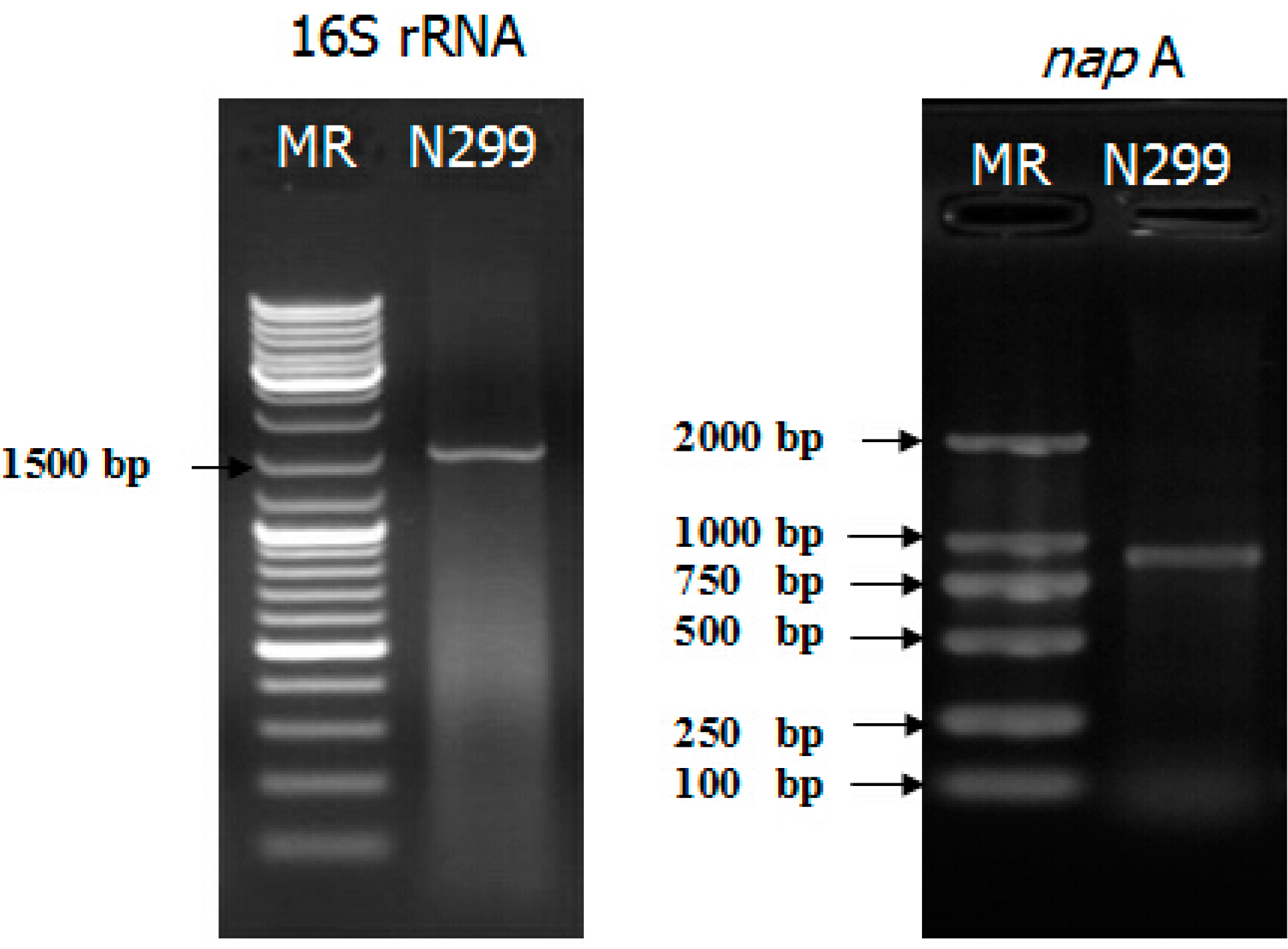
2.2. napA Examination
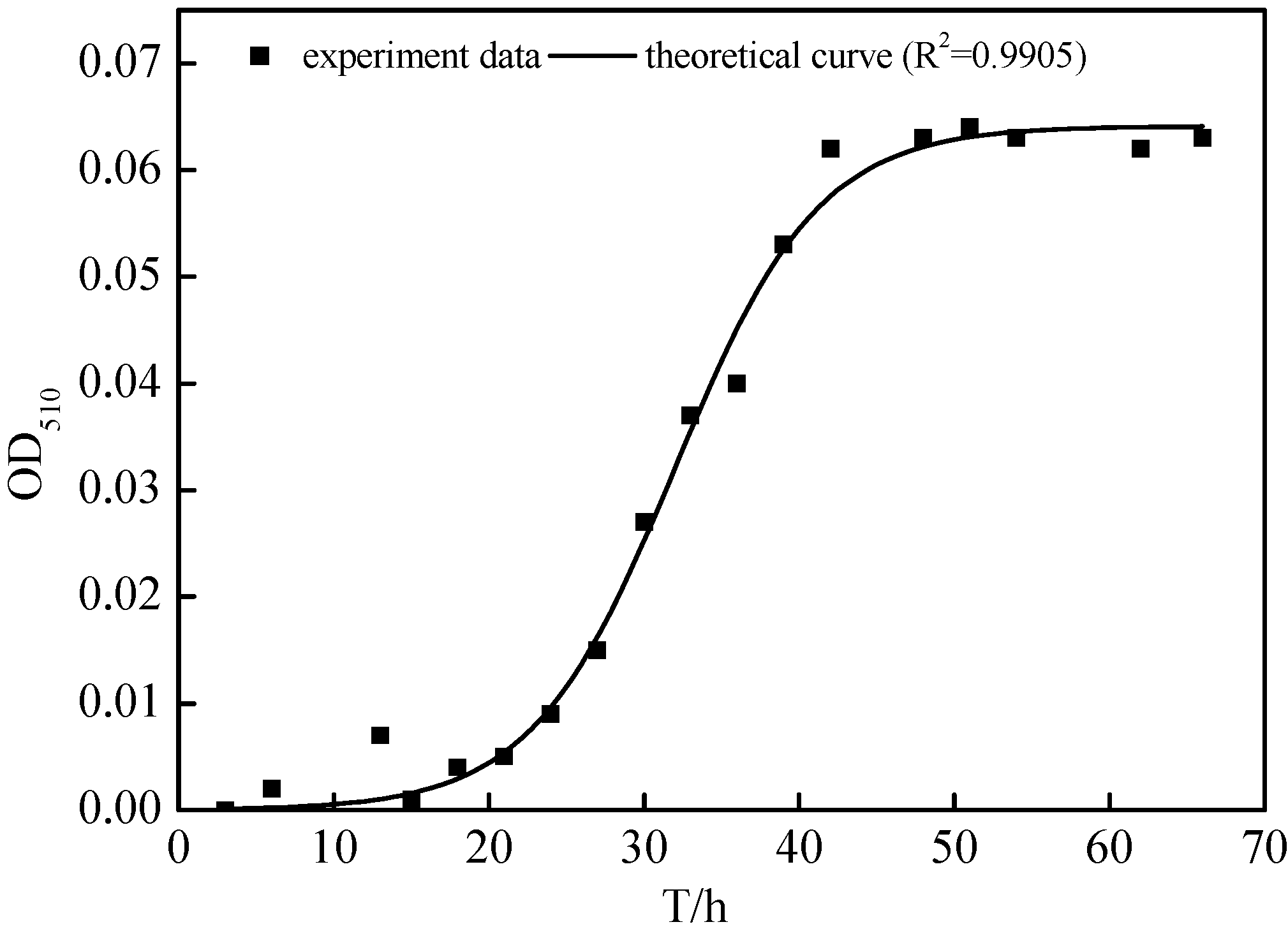
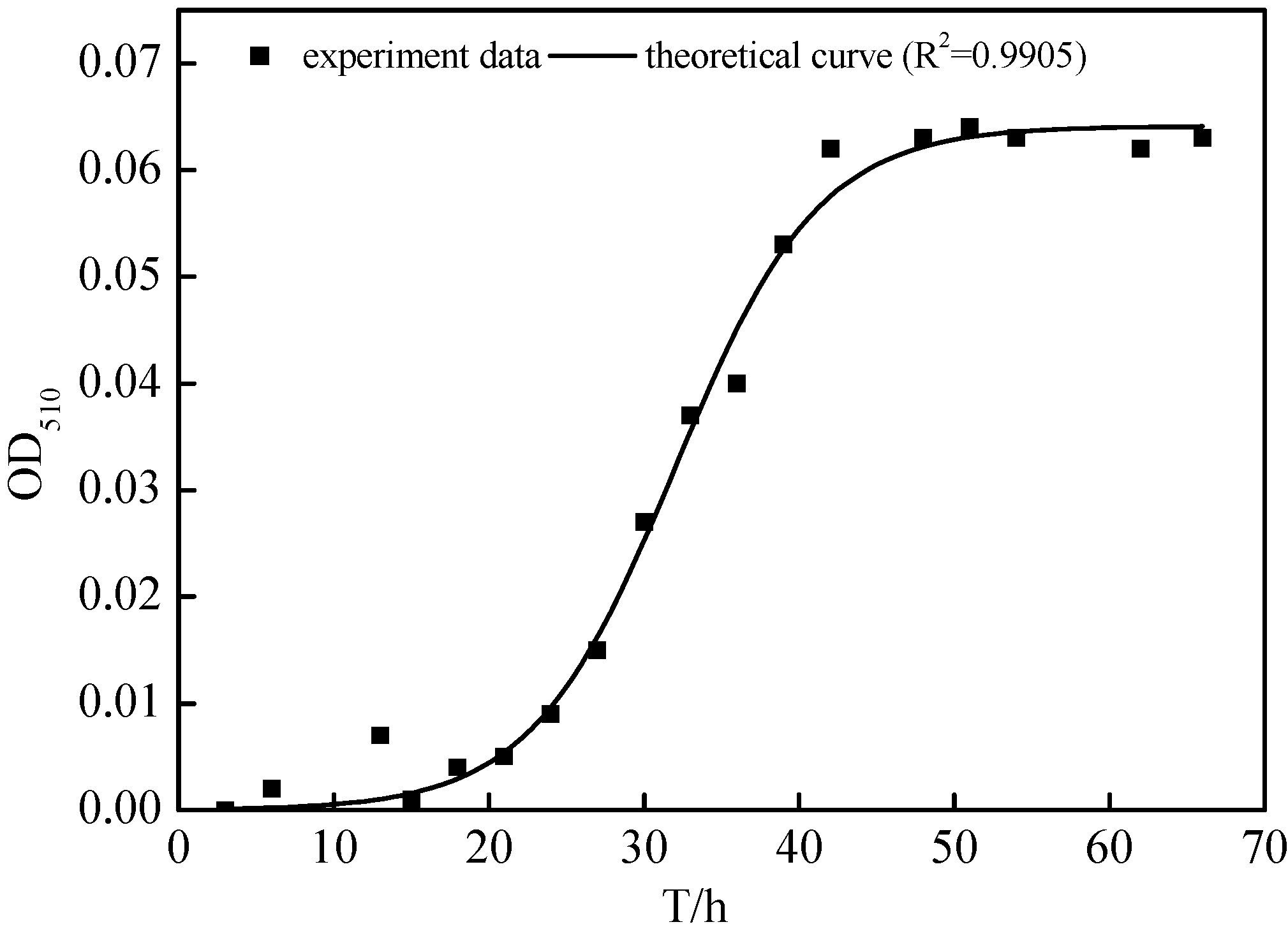
2.3. Growth Characteristics of N299
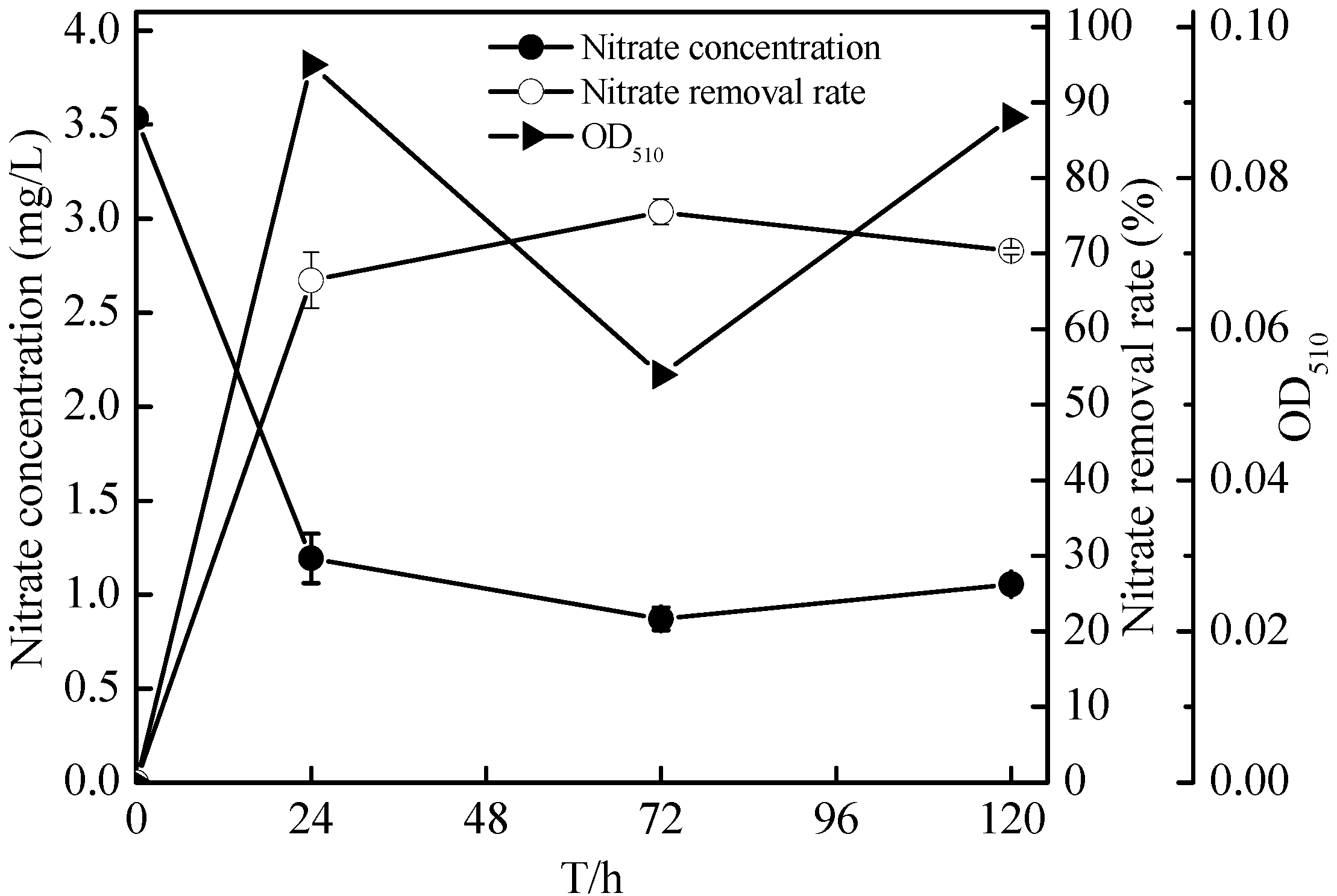
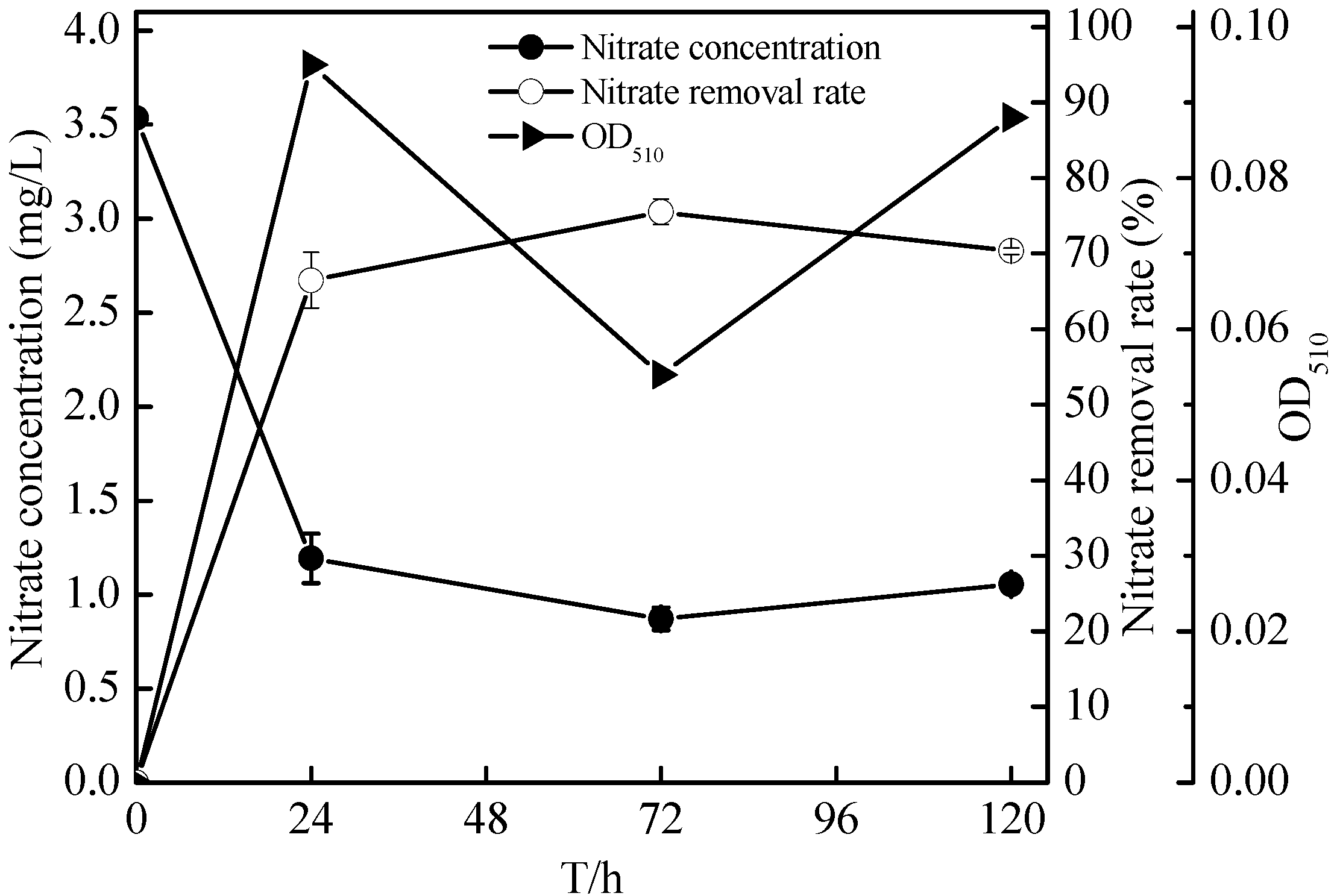
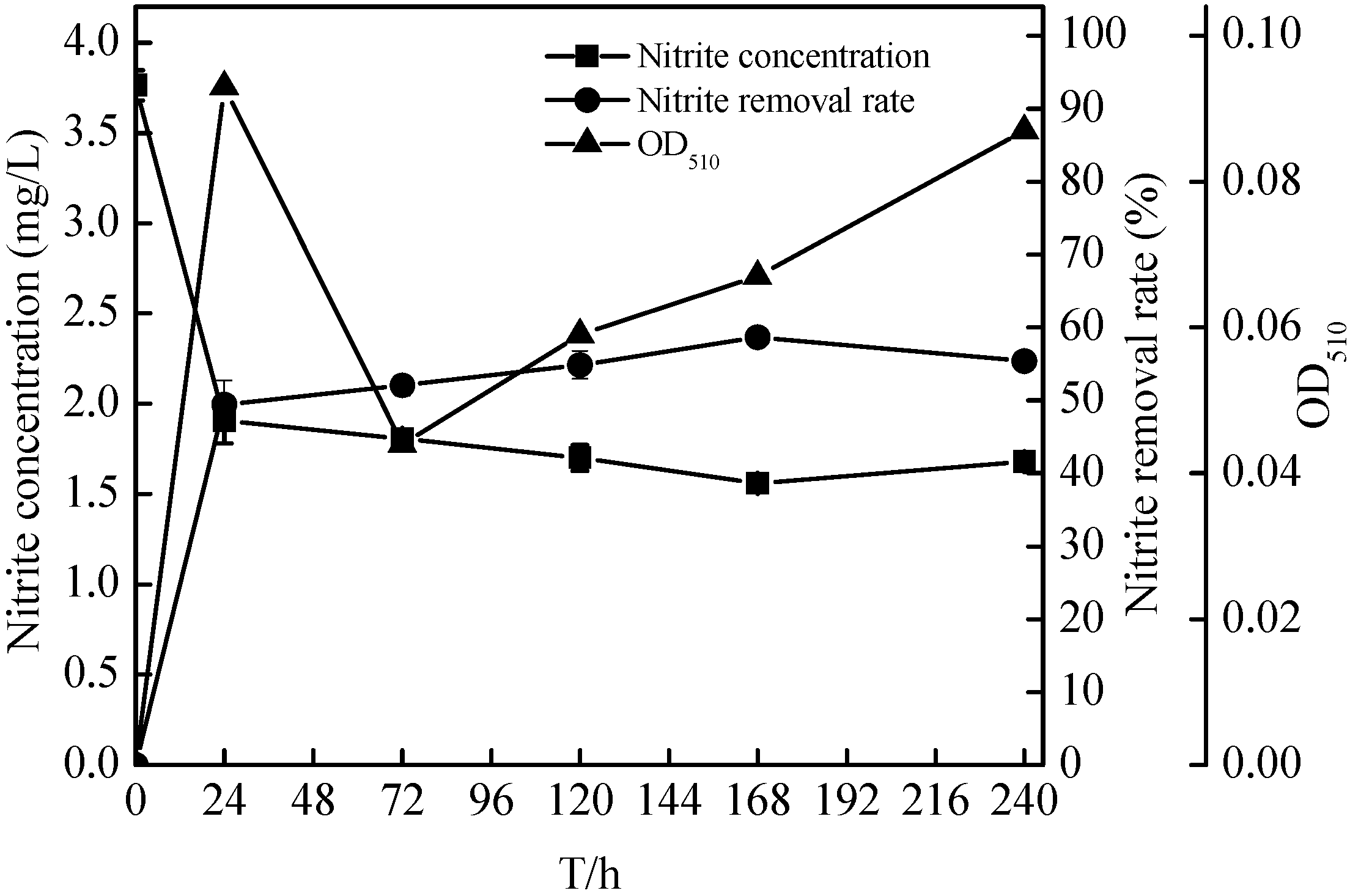
2.4. Denitrification of Zoogloea sp. N299 Using Nitrate and Nitrite as Sole Nitrogen Source
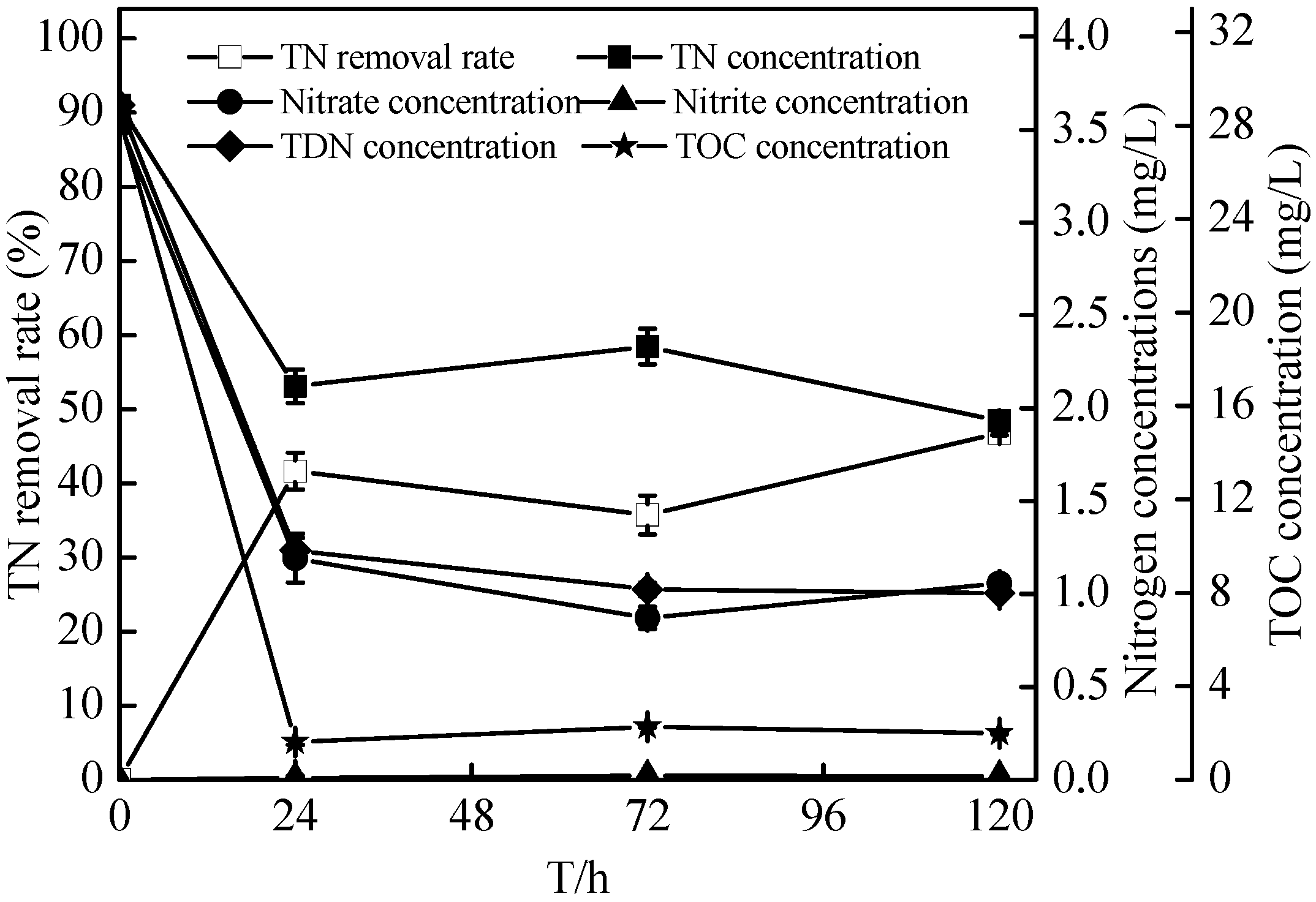
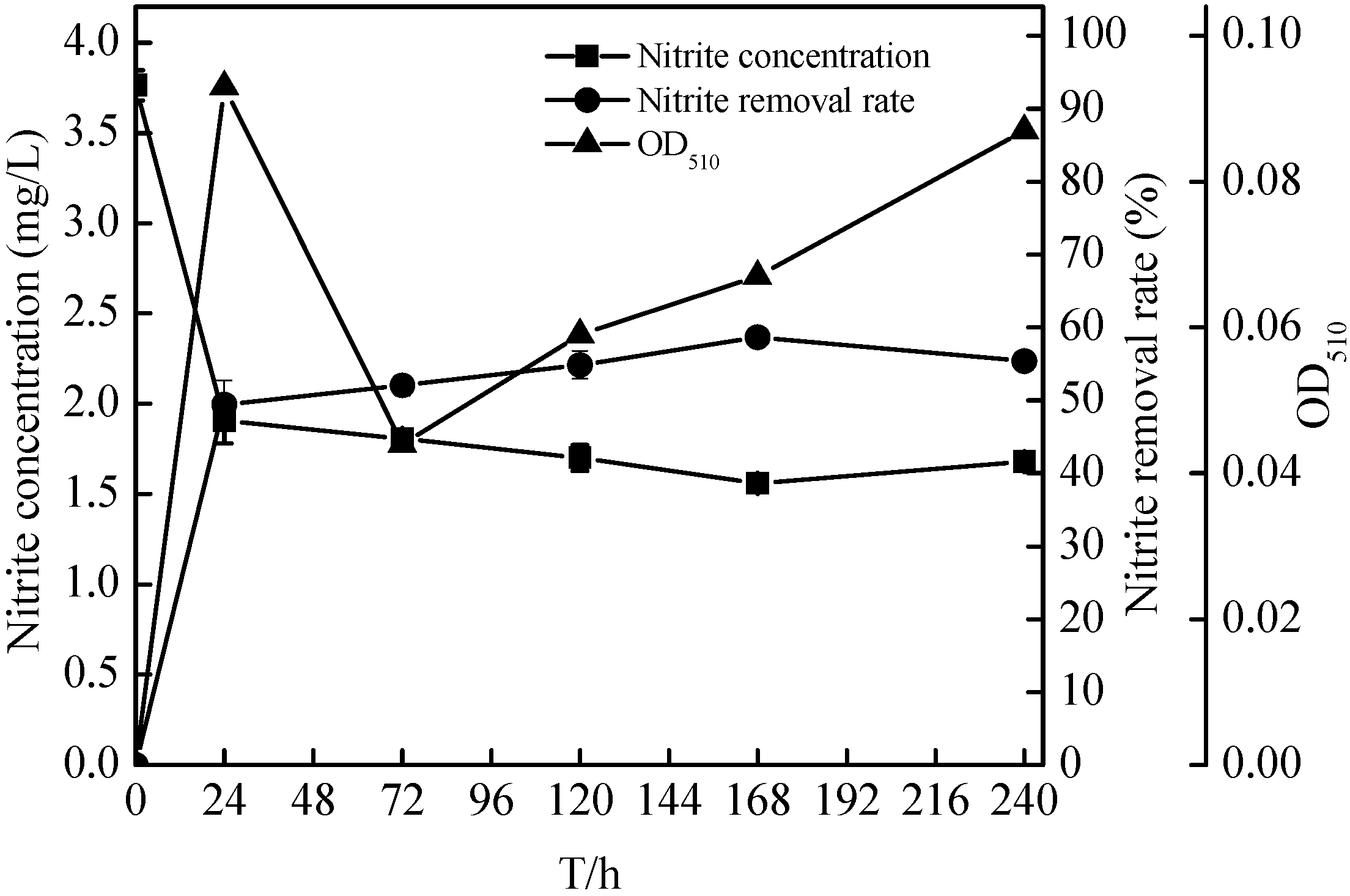
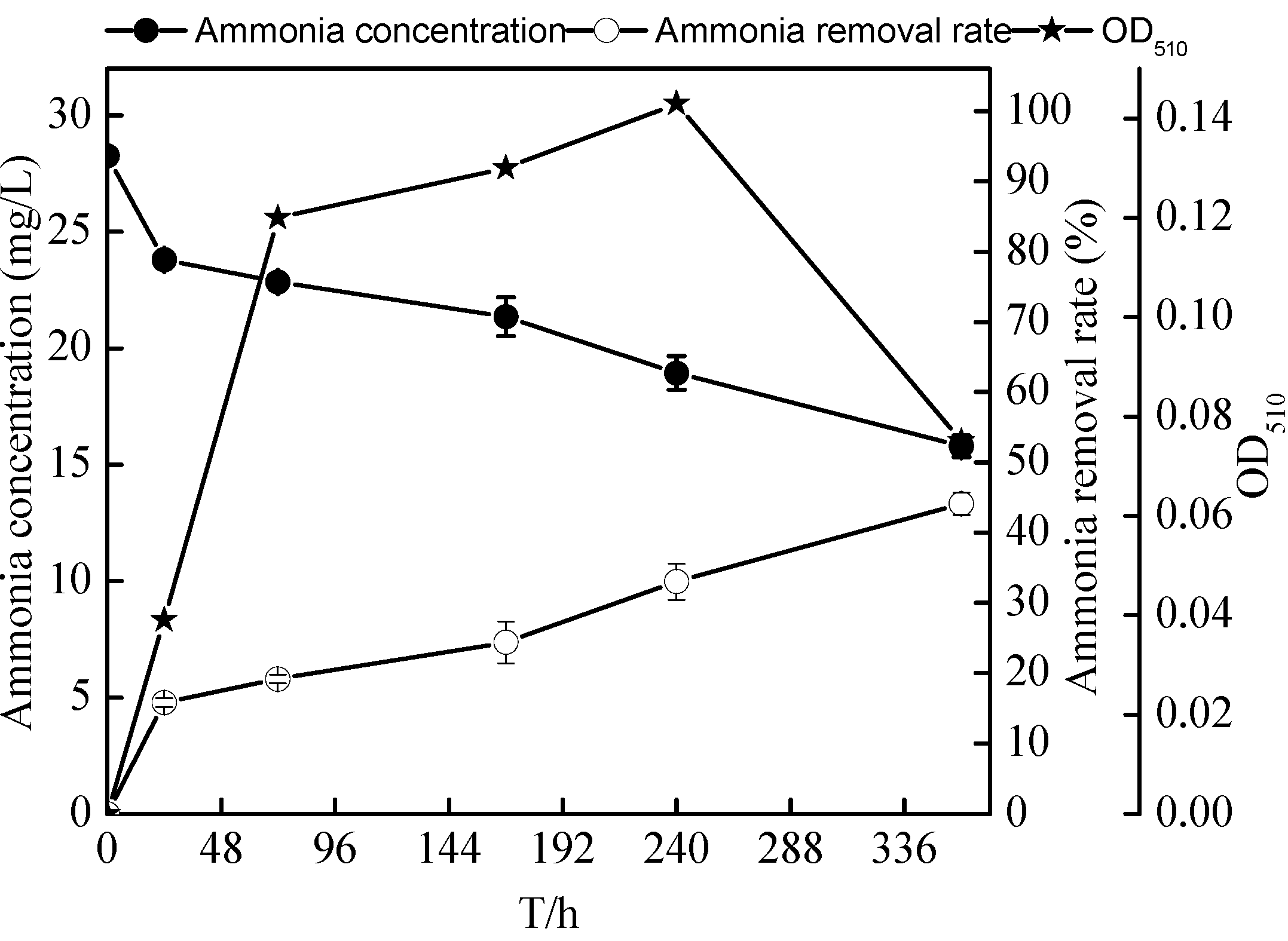
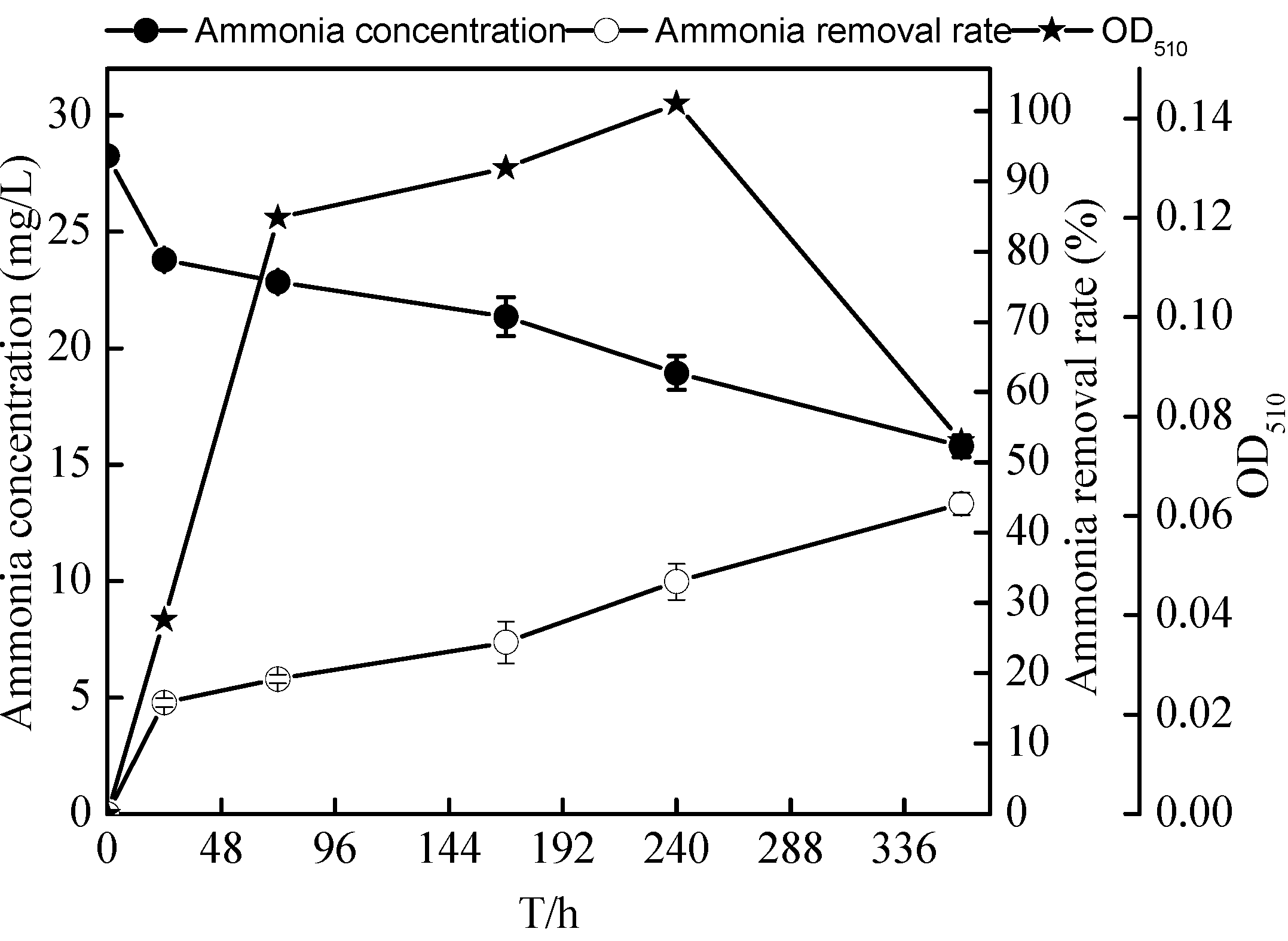
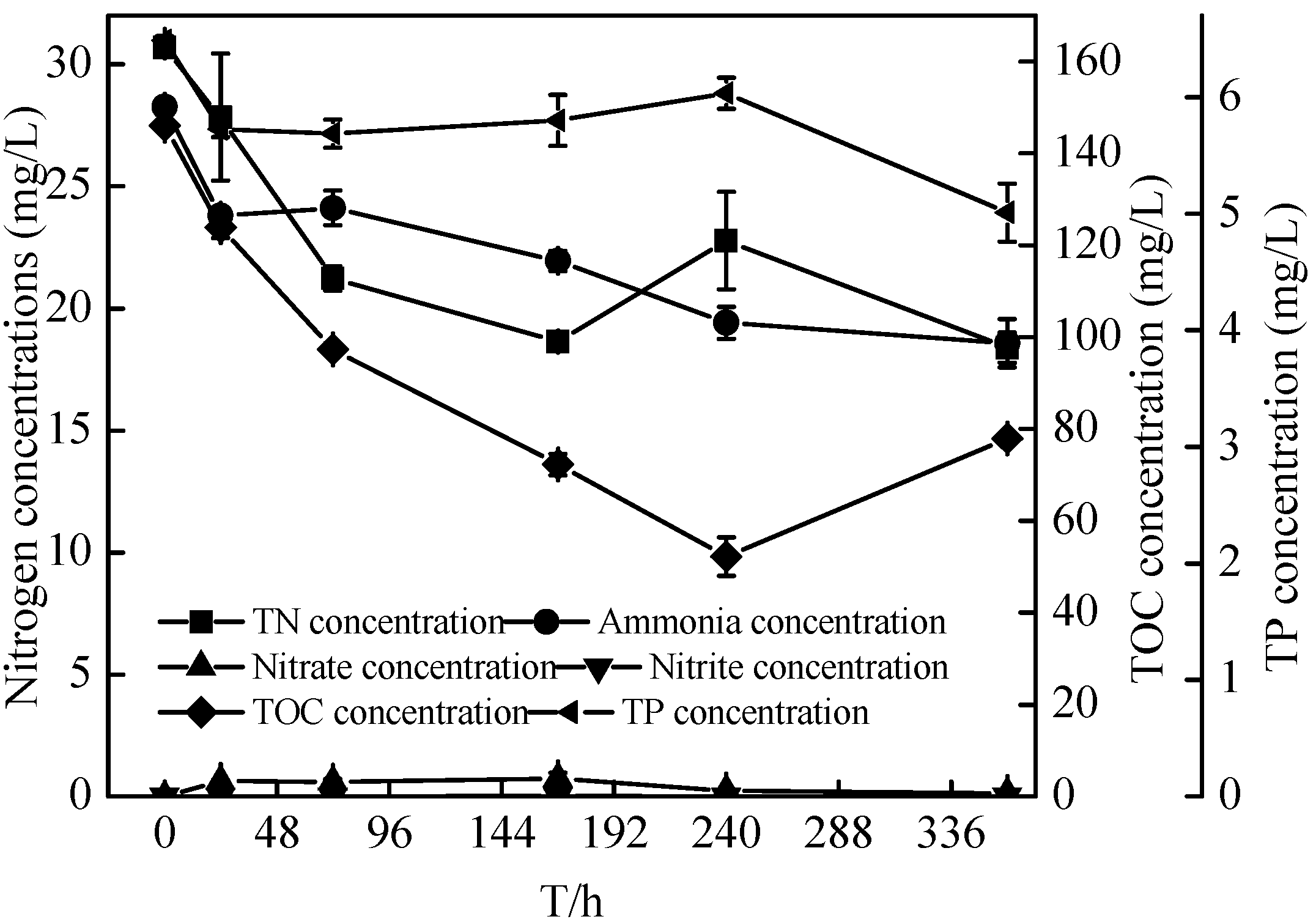
2.5. Nitrification Characteristics of the Zoogloea sp. N299
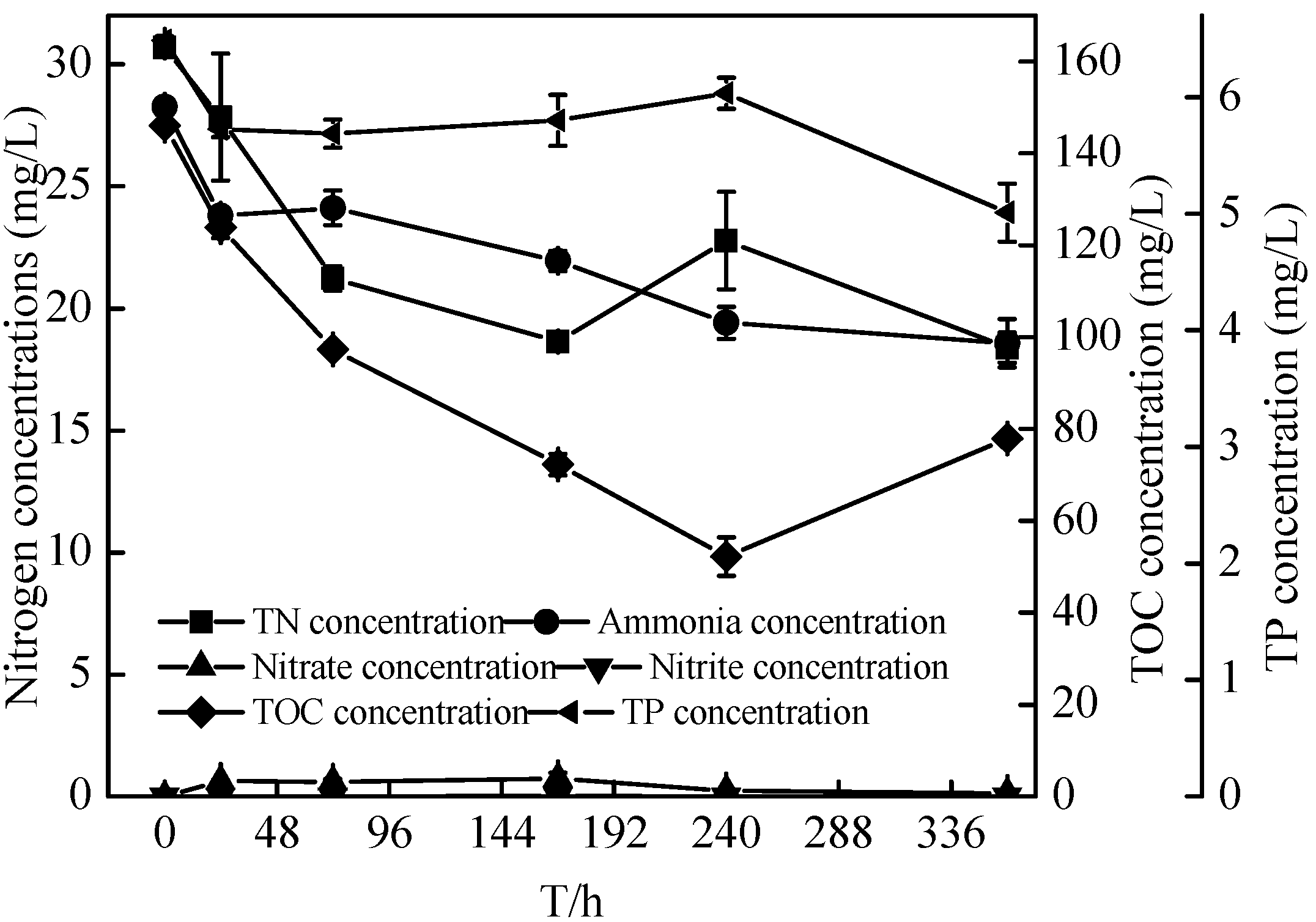
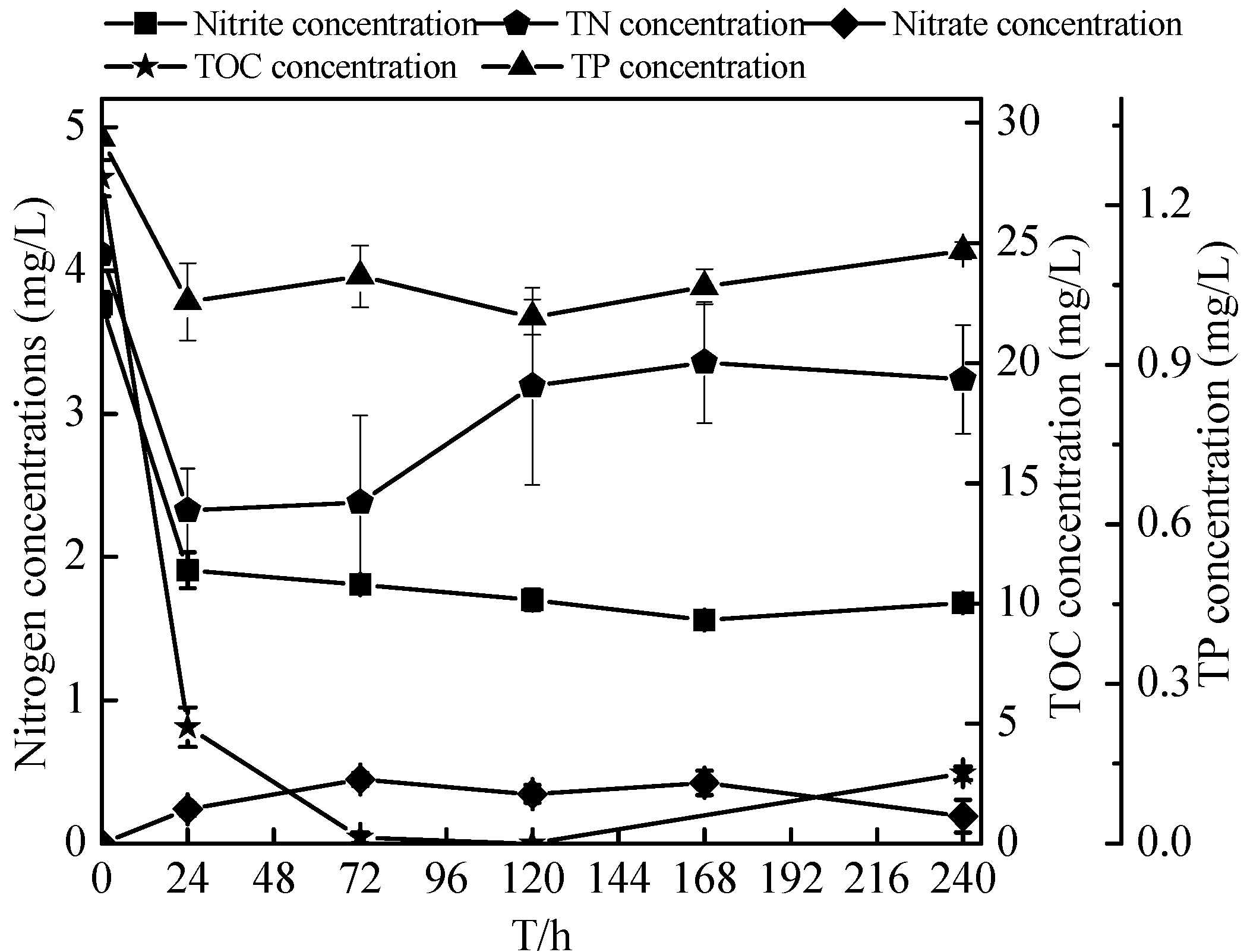
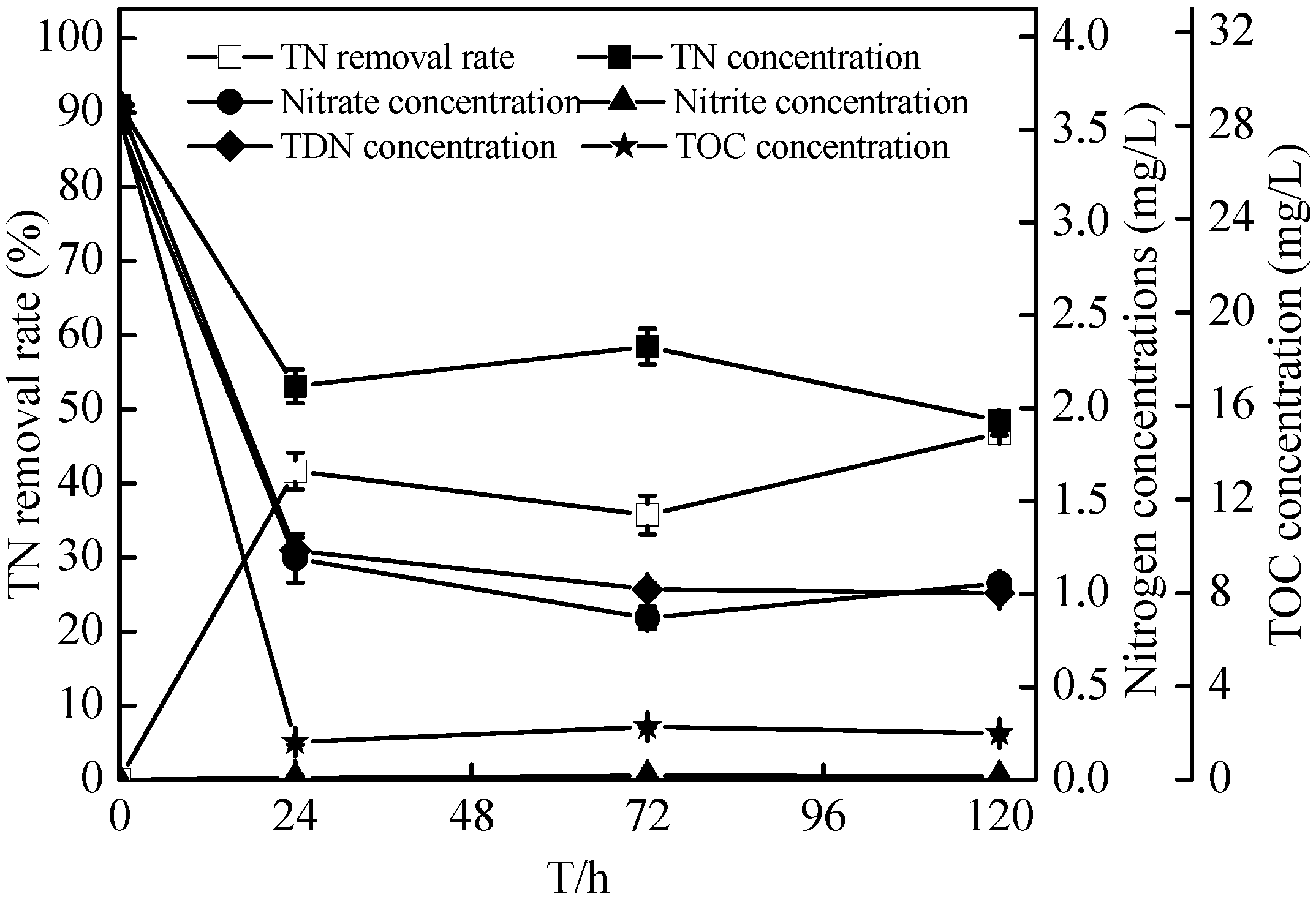
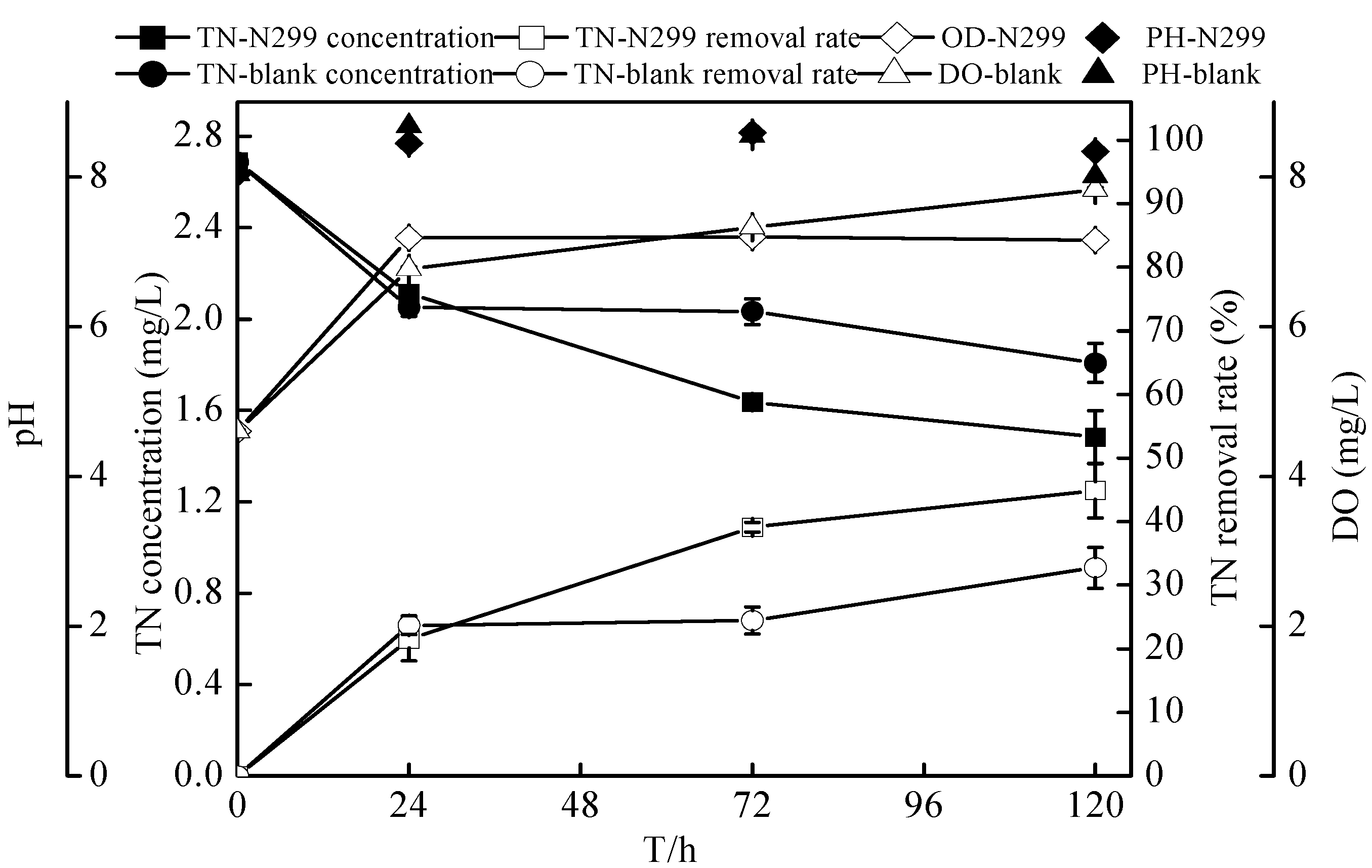
2.6. Sterilized Reservoir Source Water Experiment of the Zoogloea sp. N299

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
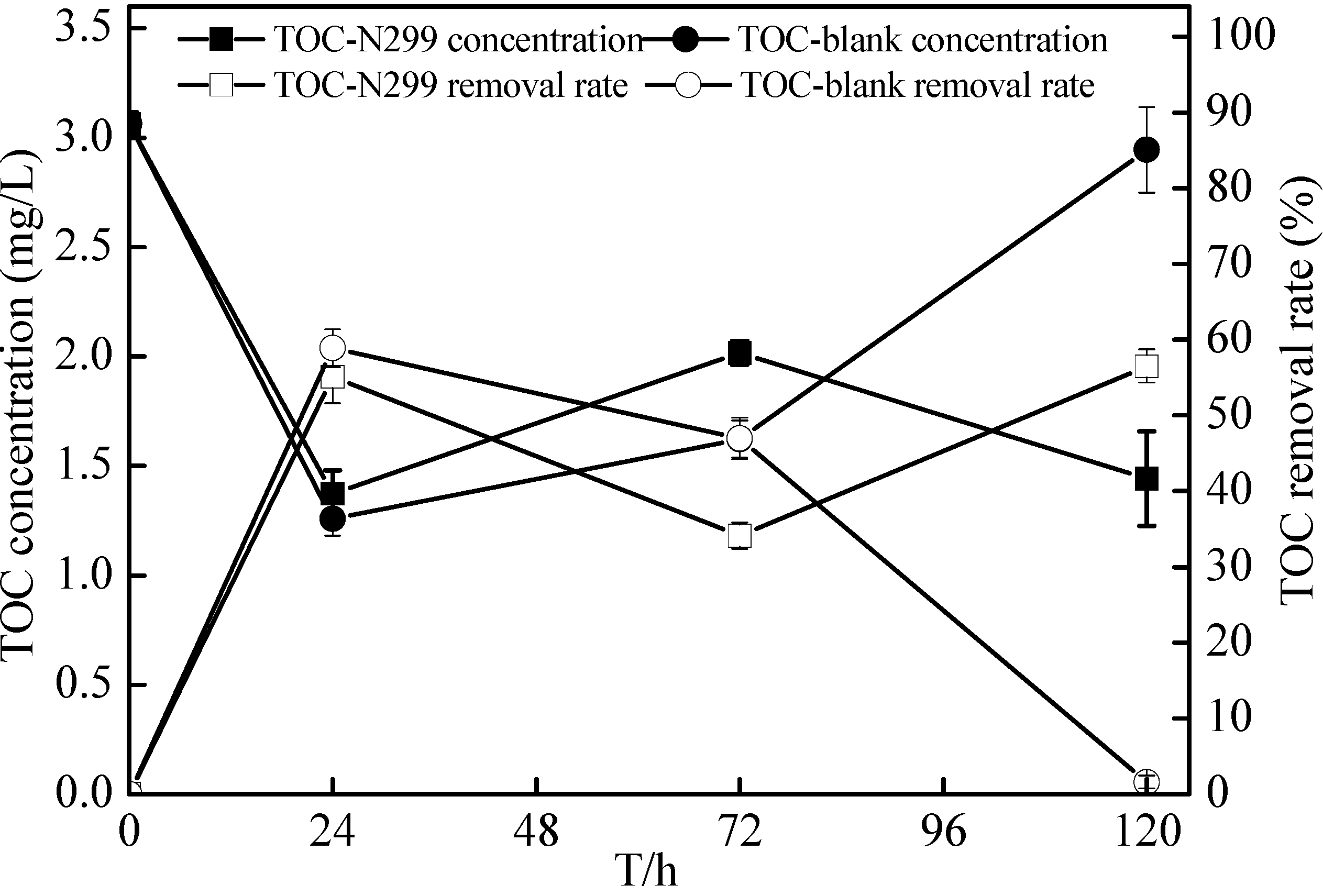
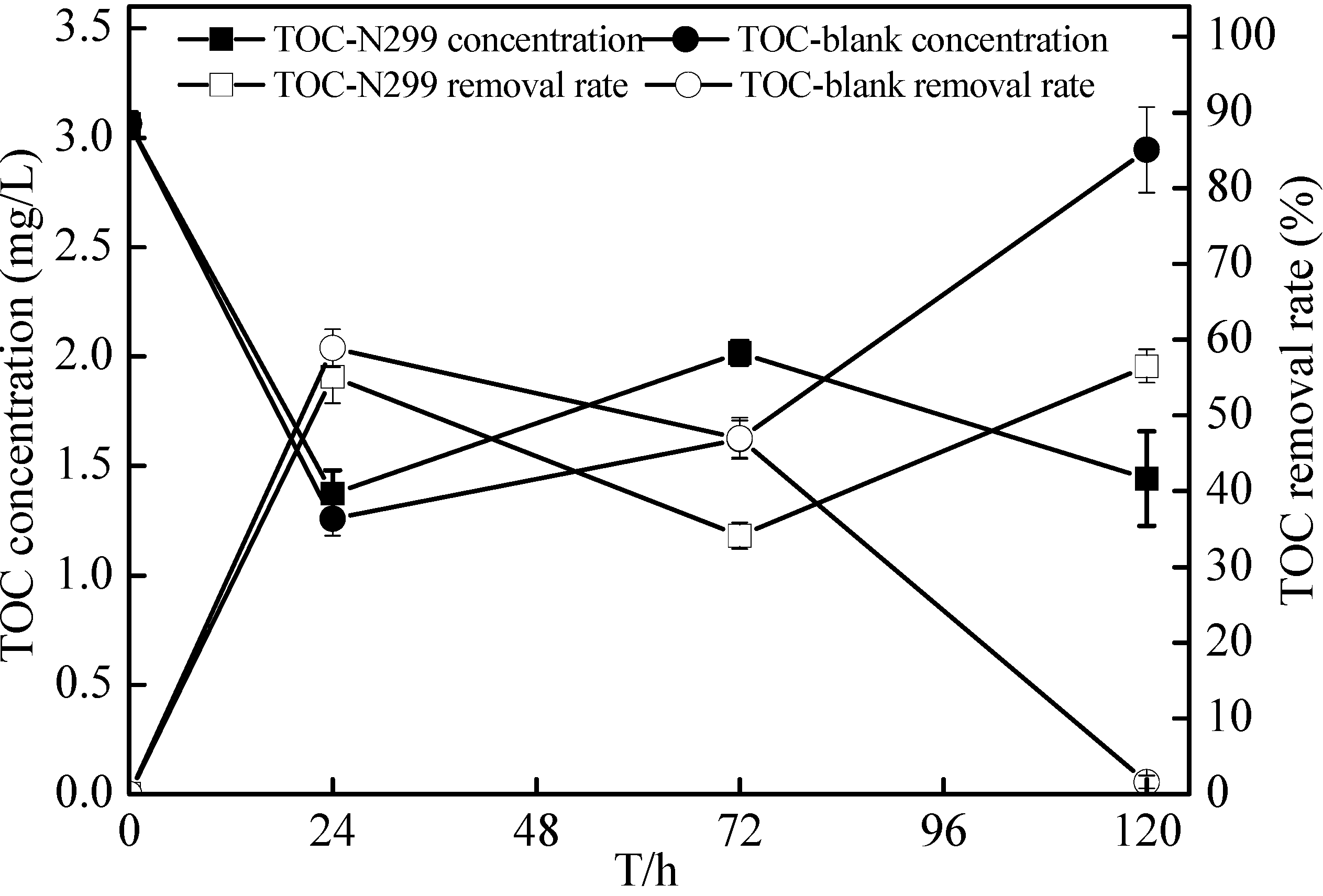{kind=link}
| Reservoir Water | Nitrate (mg/L) | Nitrite (mg/L) | TN (mg/L) | TOC (mg/L) | pH | DO (mg/L) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| sterilized reservoir source water | 1.36 | 0.02 | 0.02 | 0.00 | 2.69 | 0.03 | 3.06 | 0.05 | 8.37 | 0.01 | 8.53 | 0.03 |
| non-sterilized reservoir source water | 1.48 | 0.02 | 0.02 | 0.00 | 2.69 | 0.03 | 3.06 | 0.05 | 8.04 | 0.00 | 4.6 | 0.02 |
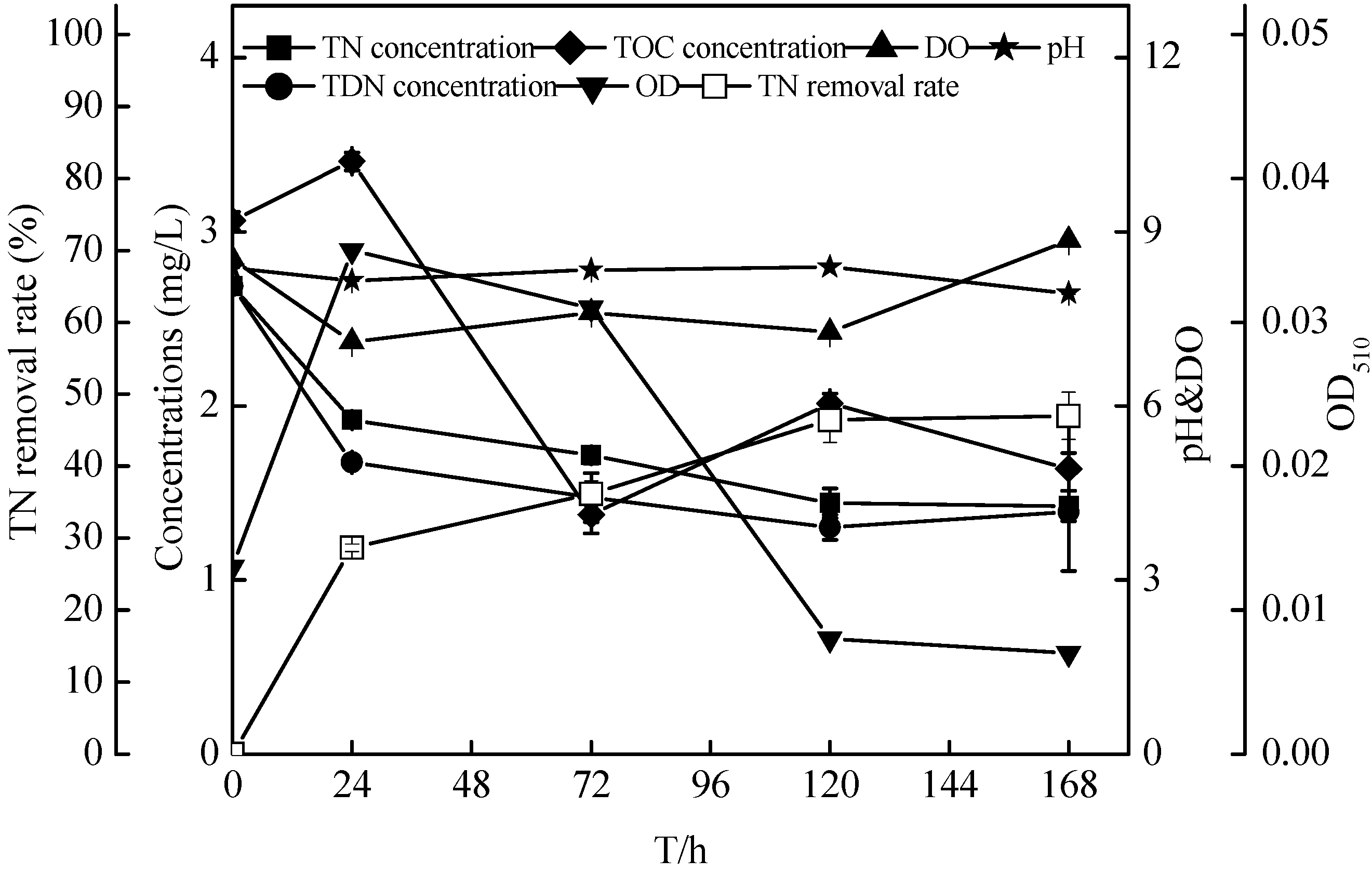
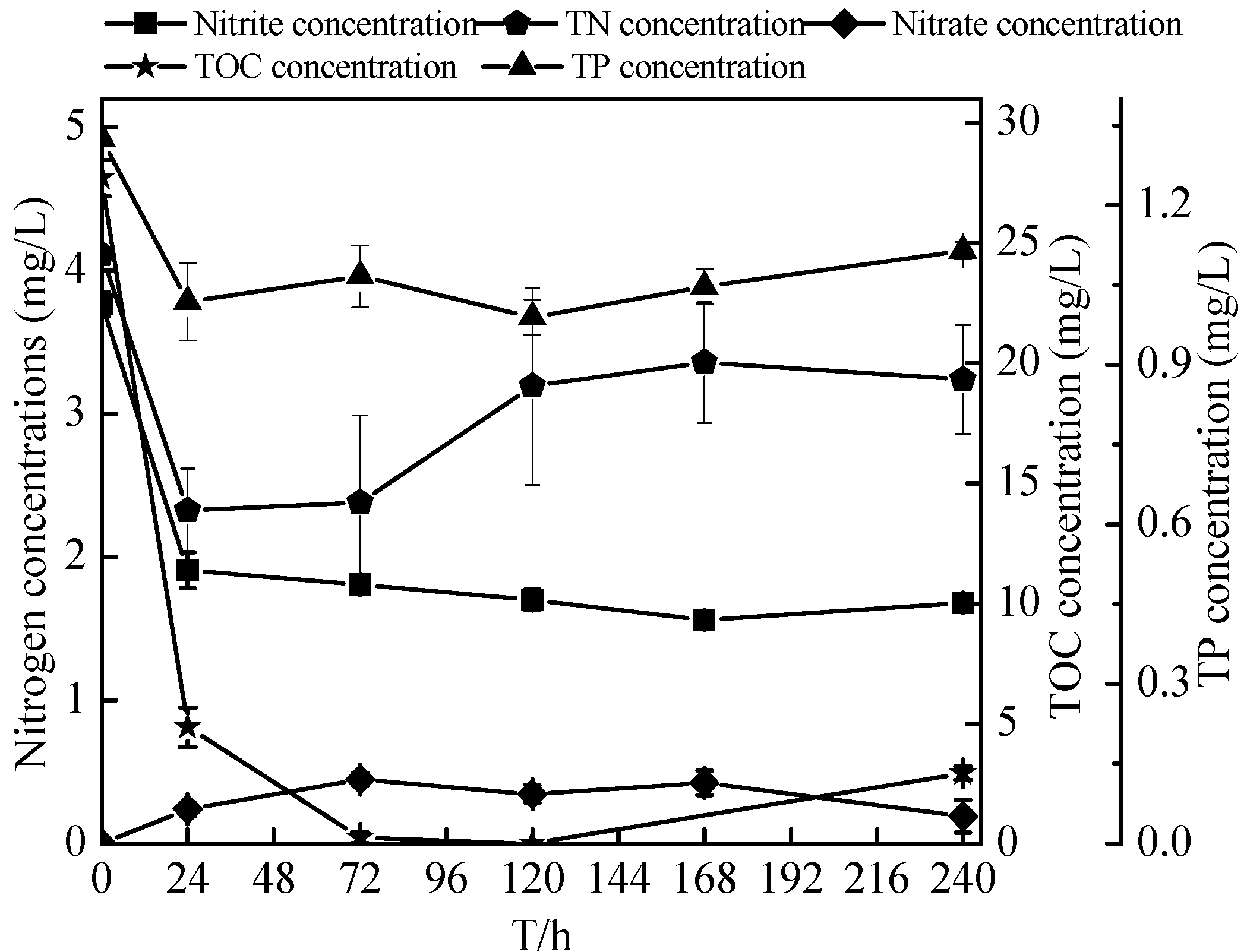
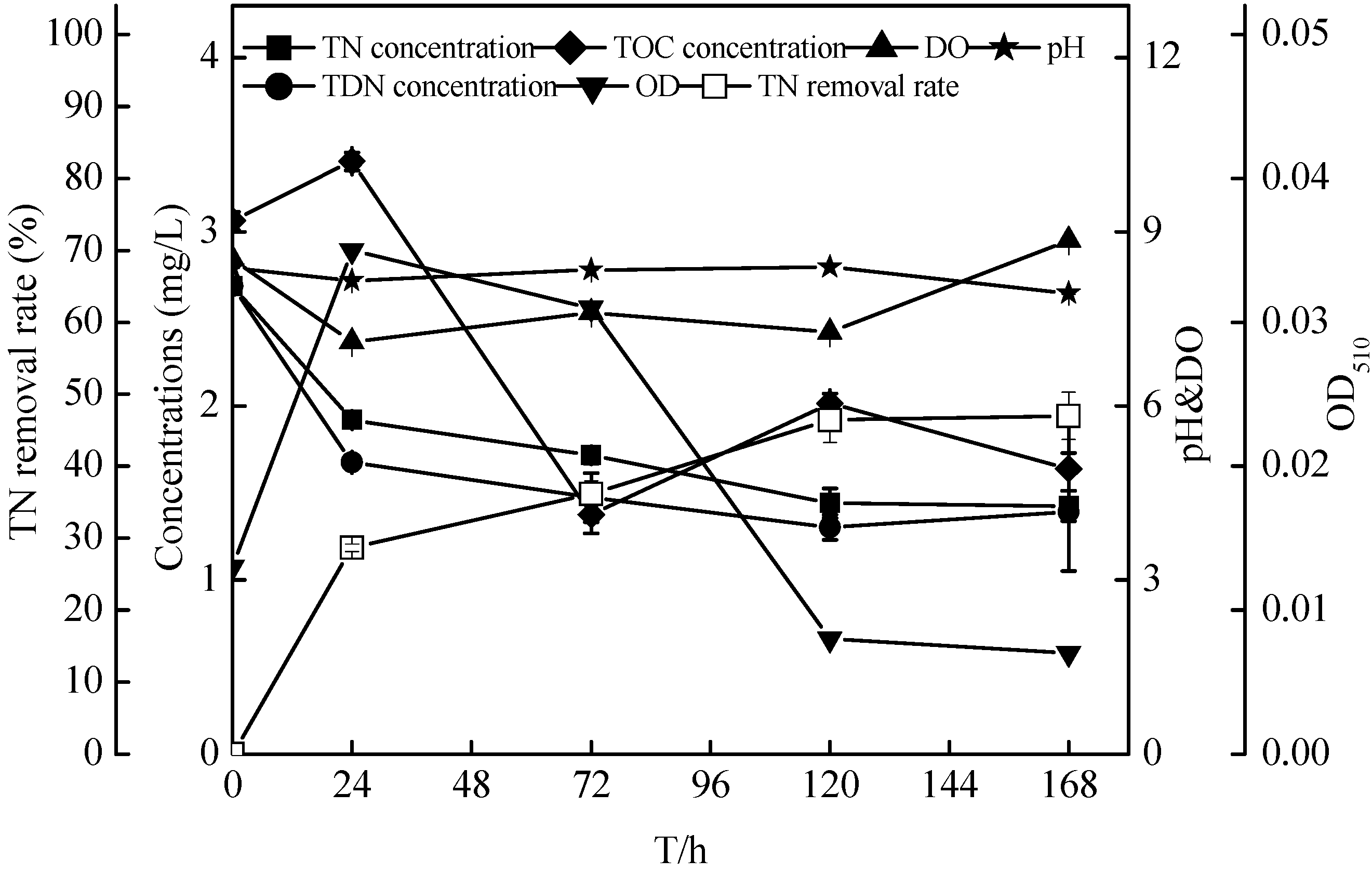
2.7. Non-Sterilized Reservoir Source Water Experiment of the Zoogloea spp. N299

| Systems/Time (h) | TN (mg/L) | TOC (mg/L) | C/N(TOC/TN) (mg/L) | pH | DO (mg/L) | OD510 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||
| Bacteria system | |||||||||||
| 0 | 2.69 | 0.03 | 3.06 | 0.05 | 1.14 | 0.02 | 8.04 | 0.01 | 4.60 | 0.02 | 0.02 |
| 24 | 2.07 | 0.04 | 1.35 | 0.14 | 0.65 | 0.04 | 8.45 | 0.01 | 7.19 | 0.01 | 0.02 |
| 72 | 1.64 | 0.16 | 1.21 | 0.04 | 1.23 | 0.05 | 8.59 | 0.01 | 7.20 | 0.01 | 0.02 |
| 120 | 1.58 | 0.13 | 1.72 | 0.25 | 0.90 | 0.12 | 8.34 | 0.02 | 7.16 | 0.05 | 0.00 |
| Blank system | |||||||||||
| 0 | 2.69 | 0.03 | 3.06 | 0.05 | 1.14 | 0.02 | 8.04 | 0.01 | 4.60 | 0.02 | 0.02 |
| 24 | 2.05 | 0.04 | 1.26 | 0.08 | 0.63 | 0.05 | 8.68 | 0.00 | 6.77 | 0.03 | 0.04 |
| 72 | 2.03 | 0.06 | 1.62 | 0.08 | 0.82 | 0.08 | 8.55 | 0.01 | 7.33 | 0.01 | 0.02 |
| 120 | 2.25 | 0.03 | 2.94 | 0.20 | 1.35 | 0.03 | 8.01 | 0.00 | 7.83 | 0.03 | 0.00 |
2.8. Effect of Different Factors on Nitrate Removal
2.8.1. Carbon Sources
| Systems | Nitrate | TN | OD510 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Concentration | Removal Rate | Concentration | Removal Rate | ||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||
| Carbon source | |||||||||
| Glucose | 0.99 | 0.01 | 70.79 | 0.24 | 1.01 | 0.02 | 73.58 | 0.40 | 0.046 |
| Sodium succinate | 1.57 | 0.01 | 53.55 | 0.23 | 1.83 | 0.02 | 52.10 | 0.40 | 0.036 |
| Sodium citrate | 0.74 | 0.01 | 78.32 | 0.18 | 1.67 | 0.03 | 56.43 | 0.65 | 0.049 |
| Sodium acetate | 0.72 | 0.01 | 78.86 | 0.29 | 1.08 | 0.02 | 71.76 | 0.54 | 0.063 |
| Temperature | |||||||||
| 10 | 1.86 | 0.01 | 45.25 | 0.35 | 2.03 | 0.03 | 47.07 | 0.79 | 0.033 |
| 20 | 1.09 | 0.01 | 67.75 | 0.36 | 1.19 | 0.03 | 68.99 | 0.65 | 0.046 |
| 30 | 0.72 | 0.01 | 78.86 | 0.29 | 1.08 | 0.02 | 71.76 | 0.54 | 0.063 |
| C/N ratio | |||||||||
| 1 | 2.49 | 0.03 | 17.25 | 0.93 | 3.02 | 0.03 | 21.17 | 0.65 | 0.012 |
| 3 | 2.15 | 0.01 | 38.43 | 0.29 | 2.97 | 0.03 | 22.56 | 0.78 | 0.02 |
| 5 | 1.93 | 0.04 | 43.94 | 1.04 | 2.32 | 0.08 | 39.45 | 2.06 | 0.037 |
| 8 | 0.72 | 0.01 | 78.86 | 0.29 | 1.08 | 0.02 | 71.76 | 0.54 | 0.063 |
| 10 | 0.44 | 0.02 | 85.18 | 0.80 | 0.95 | 0.03 | 75.31 | 0.78 | 0.068 |
| Inoculum dosage | |||||||||
| 2% | 0.72 | 0.01 | 78.86 | 0.29 | 1.08 | 0.02 | 71.76 | 0.54 | 0.063 |
| 3% | 0.77 | 0.01 | 77.23 | 0.41 | 1.16 | 0.02 | 69.68 | 0.54 | 0.055 |
| 5% | 0.77 | 0.01 | 77.34 | 0.36 | 1.25 | 0.03 | 67.34 | 0.79 | 0.048 |
| 10% | 0.72 | 0.01 | 78.79 | 0.41 | 1.10 | 0.03 | 71.24 | 0.65 | 0.051 |
| pH | |||||||||
| 6 | 1.83 | 0.01 | 45.89 | 0.25 | 2.37 | 0.02 | 38.06 | 0.40 | 0.044 |
| 7 | 0.72 | 0.01 | 78.86 | 0.29 | 1.08 | 0.02 | 71.76 | 0.54 | 0.063 |
| 8 | 0.86 | 0.01 | 74.53 | 0.29 | 1.42 | 0.02 | 62.93 | 0.54 | 0.042 |
| 9 | 1.97 | 0.01 | 41.82 | 0.29 | 1.88 | 0.03 | 50.97 | 0.65 | 0.043 |
| 10 | 2.02 | 0.02 | 40.41 | 0.49 | 2.30 | 0.03 | 39.97 | 0.78 | 0.059 |
2.8.2. Temperature
2.8.3. C/N Ratio
2.8.4. Inoculum Dosage
2.8.5. pH
3. Discussion
4. Experimental Section
4.1. Samples
4.2. Enrichment Cultures and Isolation of Aerobic Denitrifiers
4.3. Analysis of 16S rRNA Gene Sequence
4.4. Amplification of the napA Gene
4.5. Growth Characteristics of the Zoogloea sp. N299
4.6. Nitrogen Removal Performance in Pure Culture Medium System
4.7. Nitrogen Removal Performance in Oligotrophic Reservoir Source Water System
4.8. Effect of Different Factors on Nitrate Removal
4.9. Analytical Methods
4.10. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Duce, R.A.; LaRoche, J.; Altieri, K.; Arrigo, K.R.; Baker, A.R.; Capone, D.G.; Cornell, S.; Dentener, F.; Galloway, J.; Ganeshram, R.S. Impacts of atmospheric anthropogenic nitrogen on the open ocean. Science 2008, 320, 893–897. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Camargo, J.A.; Alonso, Á. Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: A global assessment. Environ. Int. 2006, 32, 831–849. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Takenaka, S.; Murakami, S.; Seesuriyachan, P.; Kuntiya, A.; Aoki, K. Screening and characterization of bacteria that can utilize ammonium and nitrate ions simultaneously under controlled cultural conditions. J. Biosci. Bioeng. 2007, 103, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.Q.; Zhu, G.W.; Gao, G.; Zhang, Y.L.; Li, W.; Paerl, H.W.; Carmichael, W.W. A drinking water crisis in lake Taihu, China: Linkage to climatic variability and lake management. Environ. Manag. 2010, 45, 105–112. [Google Scholar] [CrossRef]
- Jiang, C.L.; Zhu, L.Q.; Hu, X.Q.; Cheng, J.Y.; Xie, M.H. Reasons and control of eutrophication in new reservoirs. In Eutrophication: Causes,Consequences and Control; Springer: New York, NY, USA, 2011; pp. 325–340. [Google Scholar]
- Cai, Q.; Hu, Z. Studies on eutrophication problem and control strategy in the Three Gorges Reservoir. Acta Hydrobiol. Sin. 2006, 30, 11. [Google Scholar]
- Li, L.; Wang, H.W.; Lu, J.H. Nitrogen removal using air stripping tower in urban wastewater treatment plant. China Water Wastewater 2006, 22, 92. [Google Scholar]
- Huang, H.M.; Song, Q.W.; Wang, W.J.; Wu, S.W.; Dai, J.K. Treatment of anaerobic digester effluents of nylon wastewater through chemical precipitation and a sequencing batch reactor process. J. Environ. Manag. 2012, 101, 68–74. [Google Scholar] [CrossRef]
- Zhu, G.B.; Peng, Y.Z.; Li, B.K.; Guo, J.H.; Yang, Q.; Wang, S.Y. Biological removal of nitrogen from wastewater. In Reviews of Environmental Contamination and Toxicology; Whitacre, D., Ed.; Springer: New York, NY, USA, 2008; Volume 192, pp. 159–195. [Google Scholar]
- Joo, H.S.; Hirai, M.; Shoda, M. Characteristics of ammonium removal by heterotrophic nitrification-aerobic denitrification by Alcaligenes faecalis No. 4. J. Biosci. Bioeng. 2005, 100, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Van Rijn, J.; Tal, Y.; Schreier, H.J. Denitrification in recirculating systems: Theory and applications. Aquac. Eng. 2006, 34, 364–376. [Google Scholar] [CrossRef]
- Robertson, L.A.; Kuenen, J.G. Thiosphaera pantotropha gen. nov. sp. nov., a facultatively anaerobic, facultatively autotrophic sulphur bacterium. J. Gen. Microbiol. 1983, 129, 2847–2855. [Google Scholar]
- Robertson, L.A.; Kuenen, J.G. Aerobic denitrification: A controversy revived. Arch. Microbiol. 1984, 139, 351–354. [Google Scholar] [CrossRef]
- Robertson, L.A.; Kuenen, J.G.; Kleijntjens, R. Aerobic denitrification and heterotrophic nitrification by Thiosphaera pantotropha. Antonie Van Leeuwenhoek 1985, 51, 445–445. [Google Scholar] [CrossRef]
- Gao, H.; Schreiber, F.; Collins, G.; Jensen, M.M.; Kostka, J.E.; Lavik, G.; de Beer, D.; Zhou, H.; Kuypers, M.M.M. Aerobic denitrification in permeable wadden sea sediments. ISME J. 2009, 4, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Jechalke, S.; Vogt, C.; Reiche, N.; Franchini, A.G.; Borsdorf, H.; Neu, T.R.; Richnow, H.H. Aerated treatment pond technology with biofilm promoting mats for the bioremediation of benzene, MTBE and ammonium contaminated groundwater. Water Res. 2010, 44, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Perelo, L.W. Review: In situ and bioremediation of organic pollutants in aquatic sediments. J. Hazard. Mater. 2010, 177, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.S.; Hirai, M.; Shoda, M. Piggery wastewater treatment using Alcaligenes faecalis strain No. 4 with heterotrophic nitrification and aerobic denitrification. Water Res. 2006, 40, 3029–3036. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.P.; Hsaio, Y.; Spiro, S.; Richardson, D.J. Soil and sediment bacteria capable of aerobic nitrate respiration. Appl. Environ. Microbiol. 1995, 61, 2852–2858. [Google Scholar] [PubMed]
- Zhang, D.Y.; Li, W.G.; Huang, X.F.; Qin, W.; Liu, M. Removal of ammonium in surface water at low temperature by a newly isolated Microbacterium sp. Strain SFA13. Bioresour. Technol. 2013, 137, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Ding, W.; Feng, L.J.; Dai, X.; Xu, X.Y. Characteristics of an aerobic denitrifier that utilizes ammonium and nitrate simultaneously under the oligotrophic niche. Environ. Sci. Pollut. Res. 2012, 19, 3185–3191. [Google Scholar] [CrossRef]
- Kim, M.; Jeong, S.Y.; Yoon, S.J.; Cho, S.J.; Kim, Y.H.; Kim, M.J.; Ryu, E.Y.; Lee, S.J. Aerobic denitrification of Pseudomonas putida AD-21 at different C/N ratios. J. Biosci. Bioeng. 2008, 106, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Su, J.J.; Liu, B.Y.; Liu, C.Y. Comparison of aerobic denitrification under high oxygen atmosphere by Thiosphaera pantotropha ATCC 35512 and Pseudomonas stutzeri SU2 newly isolated from the activated sludge of a piggery wastewater treatment system. J. Appl. Microbiol. 2001, 90, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Ozeki, S.; Baba, I.; Takaya, N.; Shoun, H. A novel C1-using denitrifier Alcaligenes sp. STC1 and its genes for copper-containing nitrite reductase and azurin. Biosci. Biotechnol. Biochem. 2001, 65, 1206–1210. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.K.; Tseng, S.K. Nitrate reduction by Citrobacter diversus under aerobic environment. Appl. Microbiol. Biotechnol. 2001, 55, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.B.; Wu, P.X.; Hao, B.; Yu, Z.N. Heterotrophic nitrification and aerobic denitrification by the bacterium Pseudomonas stutzeri YZN-001. Bioresour. Technol. 2011, 102, 9866–9869. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.Z.; Li, J.; Li, Q.X.; Wang, Y.C.; Li, S.P.; Ren, T.Z.; Wang, L.G. Simultaneous heterotrophic nitrification and aerobic denitrification by bacterium Rhodococcus sp. CPZ24. Bioresour. Technol. 2012, 116, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Ding, W.; Feng, L.J.; Kong, Y.; Xu, J.; Xu, X.Y. Isolation of aerobic denitrifiers and characterization for their potential application in the bioremediation of oligotrophic ecosystem. Bioresour. Technol. 2012, 108, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.Y.; Chen, Q.K.; Fang, F.; Hu, Z.X.; Wu, J.; Miao, A.J.; Xiao, L.; Chen, X.F.; Yang, L.Y. Application potential of a newly isolated indigenous aerobic denitrifier for nitrate and ammonium removal of eutrophic lake water. Bioresour. Technol. 2013, 142, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Khardenavis, A.A.; Kapley, A.; Purohit, H.J. Simultaneous nitrification and denitrification by diverse Diaphorobacter sp. Appl. Microbiol. Biotechnol. 2007, 77, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Obaja, D.; Macé, S.; Mata-Alvarez, J. Biological nutrient removal by a sequencing batch reactor (SBR) using an internal organic carbon source in digested piggery wastewater. Bioresour. Technol. 2005, 96, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Heaton, T.; Talma, A.; Vogel, J. Origin and history of nitrate in confined groundwater in the western Kalahari. J. Hydrol. 1983, 62, 243–262. [Google Scholar] [CrossRef]
- Wilson, G.B.; Andrews, J.N.; Bath, A.H. Dissolved gas evidence for denitrification in the Lincolnshire Limestone groundwaters, eastern England. J. Hydrol. 1990, 113, 51–60. [Google Scholar] [CrossRef]
- Huang, T.L.; Wei, W.; Su, J.F.; Zhang, H.H.; Li, N. Denitrification performance and microbial community structure of a combined WLA–OBCO system. PLoS ONE 2012, 7, e48339. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.L.; Wei, W.; Wang, C.Y.; Huang, Z.; Su, J.F.; Zhi, L. Pilot research on micropollutants removal in the raw water by combined process of water-lifting aeration and oligotrophic biofilm. J. Chongqing Univ. 2012, 35, 125–146. [Google Scholar]
- Wei, W.; Huang, T.L.; Su, J.F.; Wang, C.Y.; Huang, Z.; Li, N. Isolation and identification of an oligotrophic and aerobic denitrification and its denitrification characteristics. Ecol. Environ. Sci. 2010, 19, 2166–2171. [Google Scholar]
- Wei, W.; Huang, T.L.; Li, N. Denitrification characteristics of in-situ biological inoculation under conditions of low temperature and poor nutrient. Water Technol. 2012, 6, 8–12. [Google Scholar]
- Huang, T.L.; Li, N.; Zhang, H.H.; Wang, K.; Liu, T.T. Denitrification characters and safety of communities of cold tolerant oligotrophic and aerobic denitrifying bacteria. Chin. J. Environ. Eng. 2013, 7, 2419–2423. [Google Scholar]
- Huang, T.L.; Wei, W.; Su, J.F.; Zhi, L.; Liu, Y. Biological denitrification for micro-polluted source water via in situ oligotrophic bio-contact oxidation system. Technol. Water Treat. 2010, 36, 95–99. [Google Scholar]
- Taylor, S.M.; He, Y.L.; Zhao, B.; Huang, J. Heterotrophic ammonium removal characteristics of an aerobic heterotrophic nitrifying–denitrifying bacterium, Providencia rettgeri YL. J. Environ. Sci. 2009, 21, 1336–1341. [Google Scholar] [CrossRef]
- Padhi, S.K.; Tripathy, S.; Sen, R.; Mahapatra, A.S.; Mohanty, S.; Maiti, N.K. Characterisation of heterotrophic nitrifying and aerobic denitrifying Klebsiella pneumoniae CF-S9 strain for bioremediation of wastewater. Int. Biodeterior. Biodegrad. 2013, 78, 67–73. [Google Scholar] [CrossRef]
- Zhao, B.; He, Y.L.; Huang, J.; Taylor, S.; Hughes, J. Heterotrophic nitrogen removal by Providencia rettgeri strain YL. J. Ind. Microbiol. Biotechnol. 2010, 37, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Ni, J.R. Heterotrophic nitrification–aerobic denitrification by novel isolated bacteria. J. Ind. Microbiol. Biotechnol. 2011, 38, 1305–1310. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; An, Q.; He, Y.L.; Guo, J.S. N2O and N2 production during heterotrophic nitrification by Alcaligenes faecalis strain NR. Bioresour. Technol. 2012, 116, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.K.; Ni, J.R. Ammonium removal by Agrobacterium sp. LAD9 capable of heterotrophic nitrification–aerobic denitrification. J. Biosci. Bioeng. 2012, 113, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.X.; Wang, X.W.; Jin, M.; Shen, Z.Q.; Li, J.W. Development and application of a novel and effective screening method for aerobic denitrifying bacteria. FEMS Microbiol. Lett. 2006, 260, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Gupta, S. Simultaneous carbon and nitrogen removal from high strength domestic wastewater in an aerobic RBC biofilm. Water Res. 2001, 35, 1714–1722. [Google Scholar] [CrossRef] [PubMed]
- Arts, P.A.M.; Robertson, L.A.; Gijs Kuenen, J. Nitrification and denitrification by Thiosphaera pantotropha in aerobic chemostat cultures. FEMS Microbiol. Ecol. 1995, 18, 305–315. [Google Scholar] [CrossRef]
- Wan, C.L.; Yang, X.; Lee, D.J.; Du, M.A.; Wan, F.; Chen, C. Aerobic denitrification by novel isolated strain using as nitrogen source. Bioresour. Technol. 2011, 102, 7244–7248. [Google Scholar] [CrossRef] [PubMed]
- Ji, B.; Yang, K.; Wang, H.Y.; Zhou, J.; Zhang, H.N. Aerobic denitrification by Pseudomonas stutzeri C3 incapable of heterotrophic nitrification. Bioprocess Biosyst. Eng. 2014, 38, 407–409. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Lin, J.G. Co-existence of anammox and denitrification for simultaneous nitrogen and carbon removal—Strategies and issues. J. Hazard. Mater. 2010, 178, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Su, J.J.; Liu, B.Y.; Lin, J.; Yang, C.P. Isolation of an aerobic denitrifying bacterial strain NS2 from the activated sludge of piggery wastewater treatment systems in Taiwan possessing denitrification under 92% oxygen atmosphere. J. Appl. Microbiol. 2001, 91, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.Q.; Xu, X.Y.; Luo, W.; Cao, D.; Yang, Y. Formation and microbial community analysis of chloroanilines-degrading aerobic granules in the sequencing airlift bioreactor. J. Appl. Microbiol. 2008, 104, 152–160. [Google Scholar] [PubMed]
- Zaitsev, G.; Mettänen, T.; Langwaldt, J. Removal of ammonium and nitrate from cold inorganic mine water by fixed-bed biofilm reactors. Miner. Eng. 2008, 21, 10–15. [Google Scholar] [CrossRef]
- Lin, Y.; Kong, H.N.; Wu, D.Y.; Li, C.J.; Wang, R.Y.; Tanaka, S.Z. Physiological and molecular biological characteristics of heterotrophic ammonia oxidation by Bacillus sp. LY. World J. Microbiol. Biotechnol. 2010, 26, 1605–1612. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Liu, Y.; Gao, X.Y.; Ai, G.M.; Miao, L.L.; Liu, Z.P. Characterization of a marine origin aerobic nitrifying–denitrifying bacterium. J. Biosci. Bioeng. 2012, 114, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Wei, W. Properties and Experiments of Enhanced In-Situ Biological Nitrogen Removal by Lifting Water and Aeration for Micro-Polluted Raw Water; Xi’an University of Architecture and Technology: Xi’an, China, 2011; pp. 26–27. [Google Scholar]
- Tan, L.; Zhang, X.Y.; Cao, T.L.; Gai, D.Y.; Tian, X.D. Isolation and identification of a new strain acidophilic heterotrophic bacteria from stone coal drainage. Adv. Mater. Res. 2012, 518, 598–603. [Google Scholar] [CrossRef]
- Kim, J.K.; Park, K.J.; Cho, K.S.; Nam, S.W.; Park, T.J.; Bajpai, R. Aerobic nitrification–denitrification by heterotrophic Bacillus strains. Bioresour. Technol. 2005, 96, 1897–1906. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.F.; Yang, L.Y.; Xiao, L.; Miao, A.J.; Xi, B.D. Nitrogen removal by denitrification during cyanobacterial bloom in lake taihu. J. Freshw. Ecol. 2012, 27, 243–258. [Google Scholar] [CrossRef]
- Chinese, S. Water and Wastewater Monitoring Methods Chinese Environmental Science Publishing House; Chinese Environmental Science Publishing House: Beijing, China, 2002. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, T.-L.; Zhou, S.-L.; Zhang, H.-H.; Bai, S.-Y.; He, X.-X.; Yang, X. Nitrogen Removal Characteristics of a Newly Isolated Indigenous Aerobic Denitrifier from Oligotrophic Drinking Water Reservoir, Zoogloea sp. N299. Int. J. Mol. Sci. 2015, 16, 10038-10060. https://doi.org/10.3390/ijms160510038
Huang T-L, Zhou S-L, Zhang H-H, Bai S-Y, He X-X, Yang X. Nitrogen Removal Characteristics of a Newly Isolated Indigenous Aerobic Denitrifier from Oligotrophic Drinking Water Reservoir, Zoogloea sp. N299. International Journal of Molecular Sciences. 2015; 16(5):10038-10060. https://doi.org/10.3390/ijms160510038
Chicago/Turabian StyleHuang, Ting-Lin, Shi-Lei Zhou, Hai-Han Zhang, Shi-Yuan Bai, Xiu-Xiu He, and Xiao Yang. 2015. "Nitrogen Removal Characteristics of a Newly Isolated Indigenous Aerobic Denitrifier from Oligotrophic Drinking Water Reservoir, Zoogloea sp. N299" International Journal of Molecular Sciences 16, no. 5: 10038-10060. https://doi.org/10.3390/ijms160510038