
Phytochemical Composition and Anti-Inflammatory Activity of Extracts from the Whole-Meal Flour of Italian Durum Wheat Cultivars


 , , ,
, , ,
Abstract
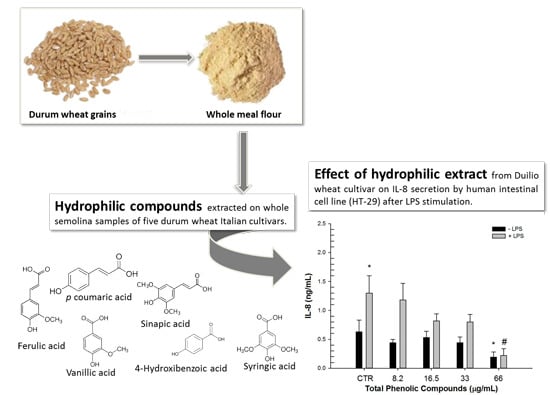
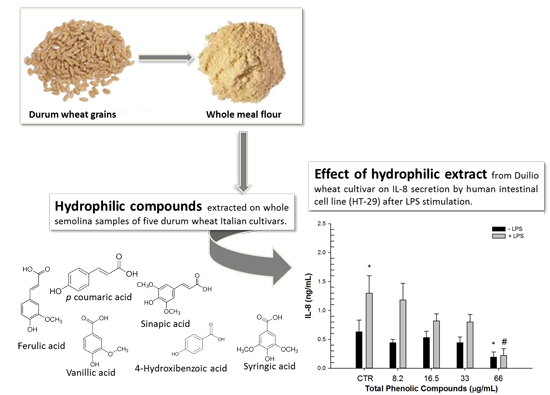
:
1. Introduction
2. Results and Discussion
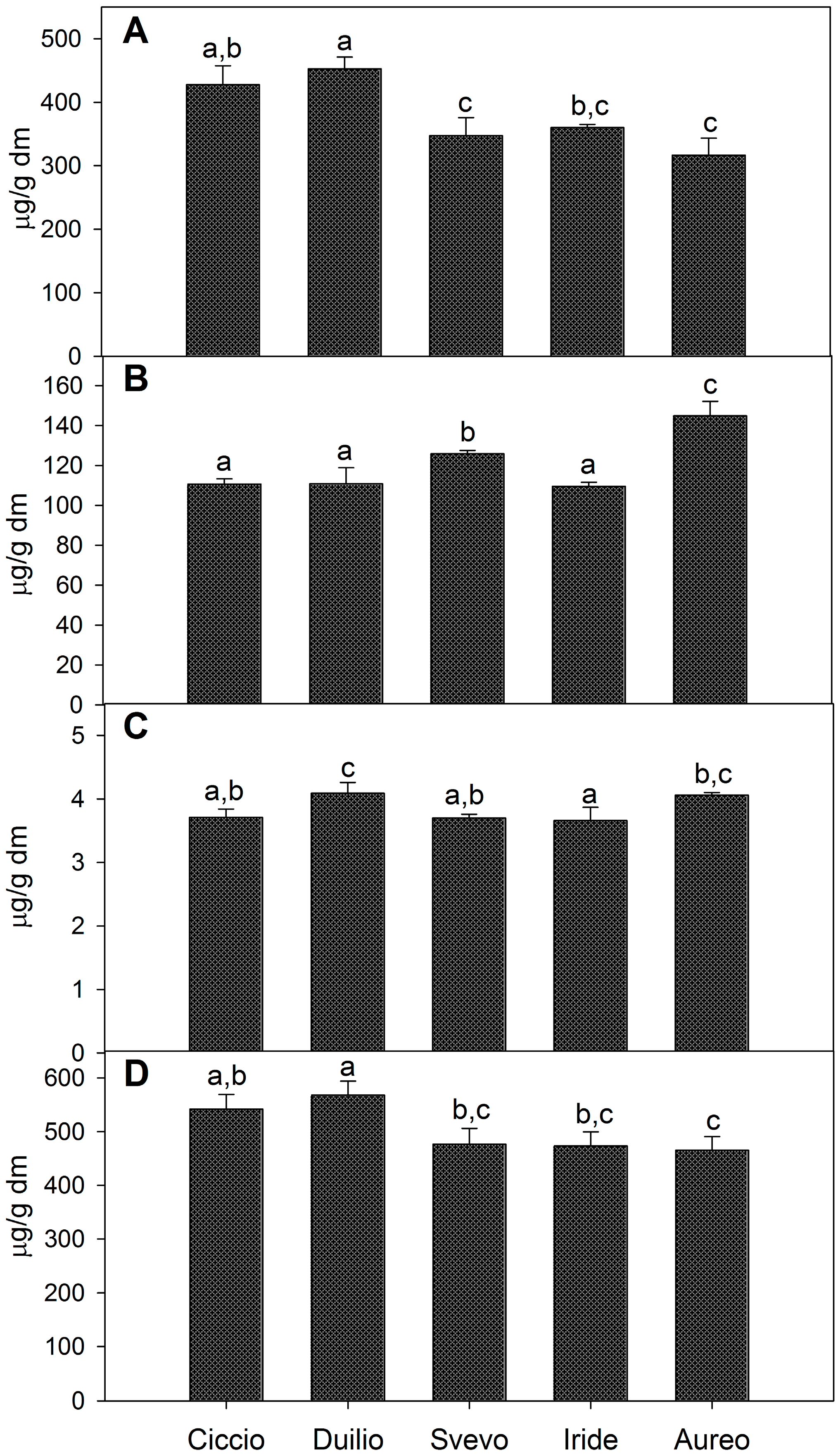
2.1. Composition of Phenolic Extracts

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Acids | Durum Wheat Cultivars | |||||
|---|---|---|---|---|---|---|
| Ciccio Mean ± SD μg/g dm | Duilio Mean ± SD μg/g dm | Svevo Mean ± SD μg/g dm | Iride Mean ± SD μg/g dm | Aureo Mean ± SD μg/g dm | ||
| Ferulic acid | ||||||
| Bound | 408.54 a,b ± 29.6 | 431.67 a ± 17.7 | 333.60 c ± 27.8 | 345.61 b,c ± 23.2 | 302.09 c ± 25.8 | |
| Conjugated | 23.21 a,b ± 0.86 | 26.32 a,b,c ± 1.65 | 28.92 c,d ± 1.31 | 22.71 a ± 1.12 | 27.55 b,d ± 2.07 | |
| Free | 1.41 a ± 0.03 | 1.62 a ± 0.09 | 1.57 a ± 0.07 | 1.60 a ± 0.26 | 1.69 a ± 0.03 | |
| p-Coumaric acid | ||||||
| Bound | 8.72 a ± 0.42 | 11.25 b ± 0.79 | 5.88 c ± 0.69 | 7.92 a,c ± 1.14 | 8.47 a ± 0.83 | |
| Conjugated | 0.85 a,b ± 0.09 | 0.62 b,c ± 0.09 | 0.44 c ± 0.10 | 0.51 c ± 0.10 | 1.05 a ± 0.19 | |
| Free | 0.87 a ± 0.08 | 0.85 a ± 0.03 | 0.76 a,b ± 0.04 | 0.70 b ± 0.02 | 0.80 a,b ± 0.03 | |
| Vanillic acid | ||||||
| Bound | 5.02 a,b ± 0.13 | 5.21 a ± 0.53 | 3.85 b,c ± 0.46 | 3.24 c ± 0.56 | 4.55 a,b ± 0.56 | |
| Conjugated | 4.44 a ± 0.09 | 5.79 b ± 0.33 | 5.08 c ± 0.10 | 4.66 a,c ± 0.09 | 5.80 b ± 0.08 | |
| Free | 1.43 a,b ± 0.03 | 1.62 c ± 0.09 | 1.36 a ± 0.05 | 1.36 a ± 0.04 | 1.58 c,b ± 0.06 | |
| Syringic acid | ||||||
| Bound | 4.71 a ± 0.58 | 3.87 a,b ± 0.13 | 3.04 b,c ± 0.12 | 2.71 c ± 0.38 | 1.19 d ± 0.08 | |
| Conjugated | 1.78 a,b,c ± 0.18 | 2.23 a,b ± 0.51 | 2.45 a ± 0.27 | 1.64 b,d ± 0.19 | 1.11 c,d ± 0.17 | |
| Free | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| 4-Hydroxibenzoic acid | ||||||
| Bound | 0.61 a ± 0.03 | 0.87 b ± 0.02 | 0.87 b ± 0.04 | 0.71 c ± 0.02 | 0.37 d ± 0.02 | |
| Conjugated | 1.23 a ± 0.02 | 1.75 b ± 0.09 | 2.67 c ± 0.04 | 3.21 d ± 0.02 | 1.98 e ± 0.02 | |
| Free | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| Sinapic acid | ||||||
| Bound | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| Conjugated | 79.12 a,b ± 2.22 | 74.09 a ± 5.94 | 86.38 b ± 1.63 | 76.83 a,b ± 3.30 | 107.39 c ± 4.74 | |
| Free | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
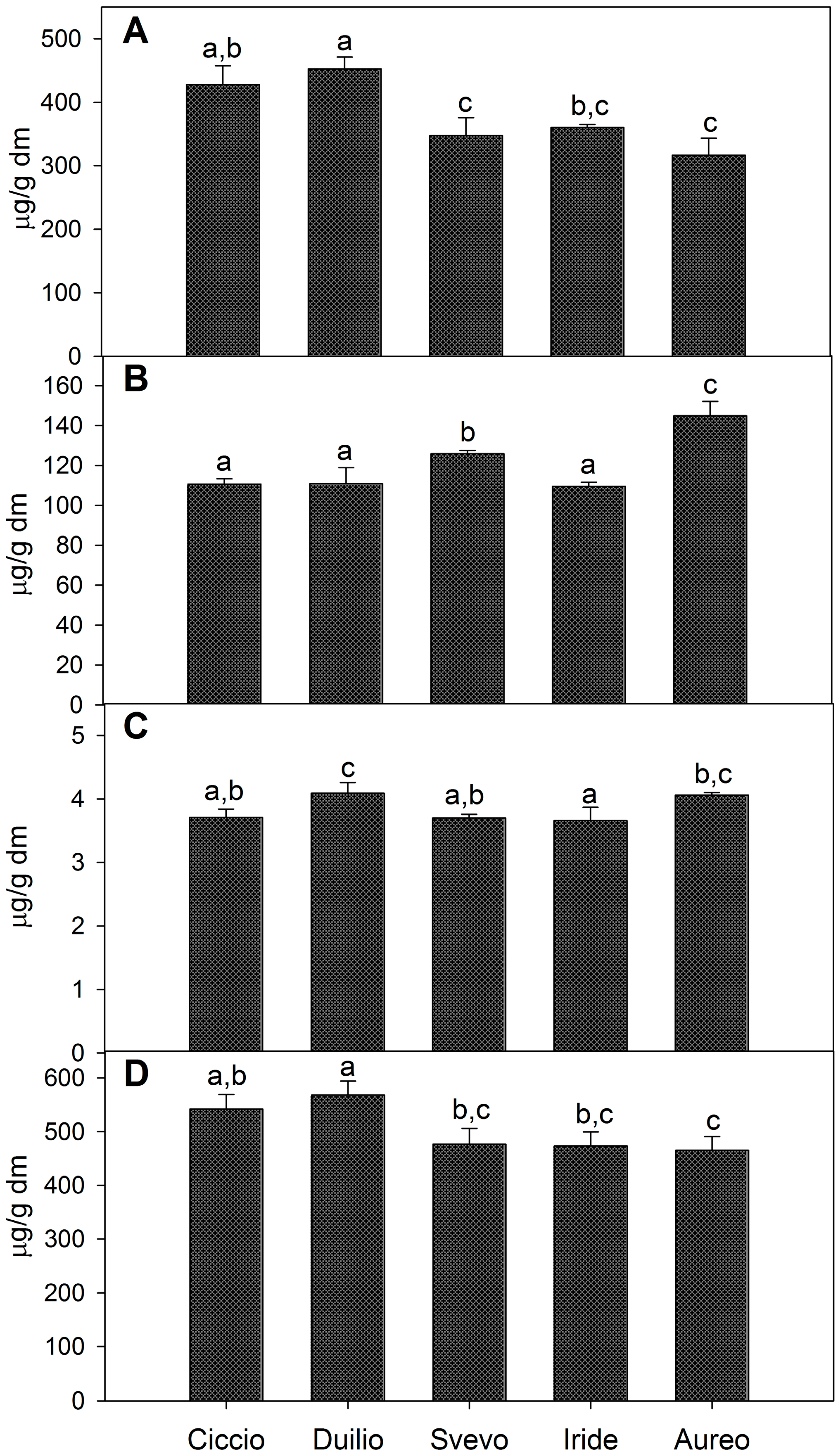
2.2. Composition of Isoprenoid Extracts


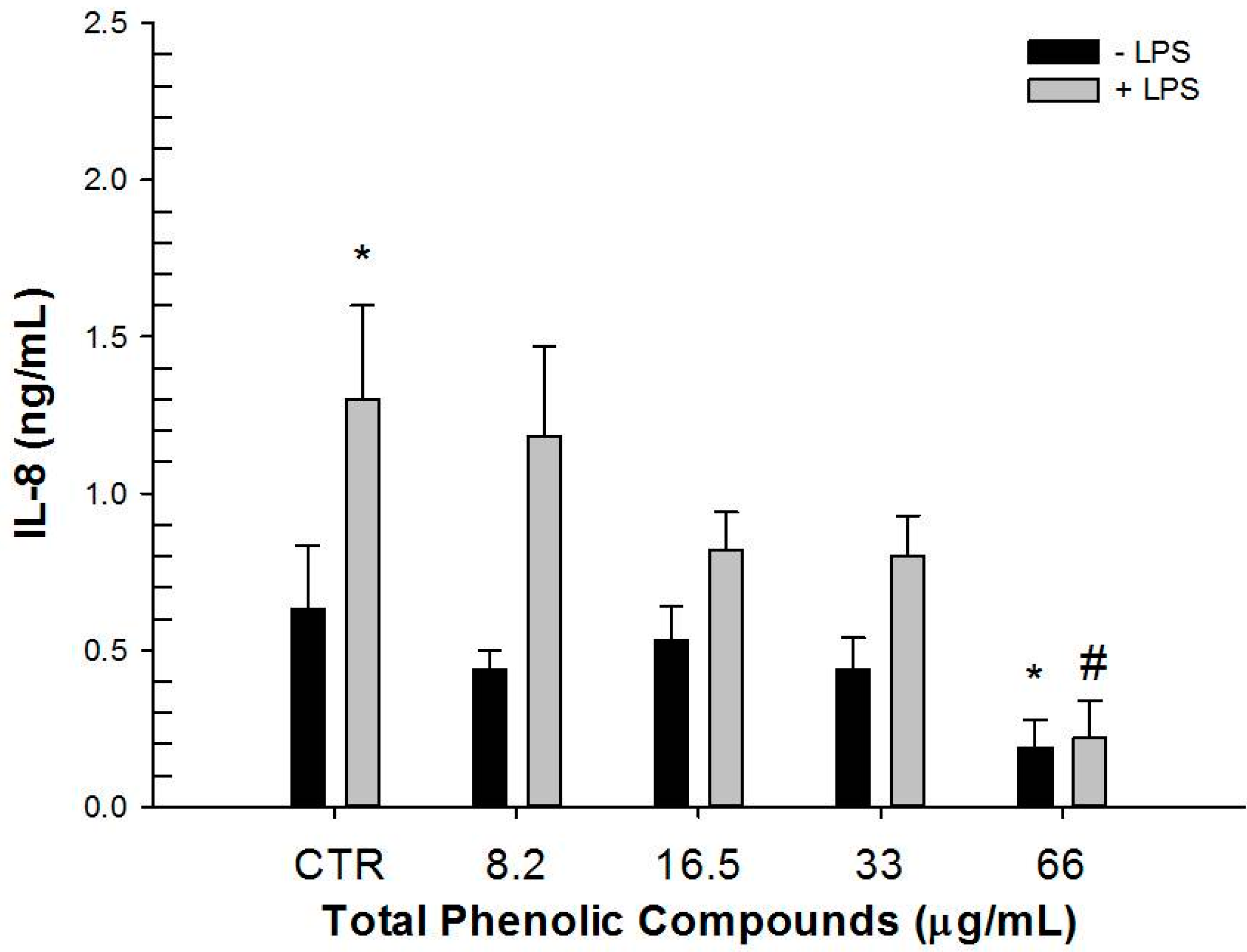
2.3. Effect of Hydrophilic and Lipophilic Extracts on IL-8 and TGF-β1 Production by HT-29 Cells
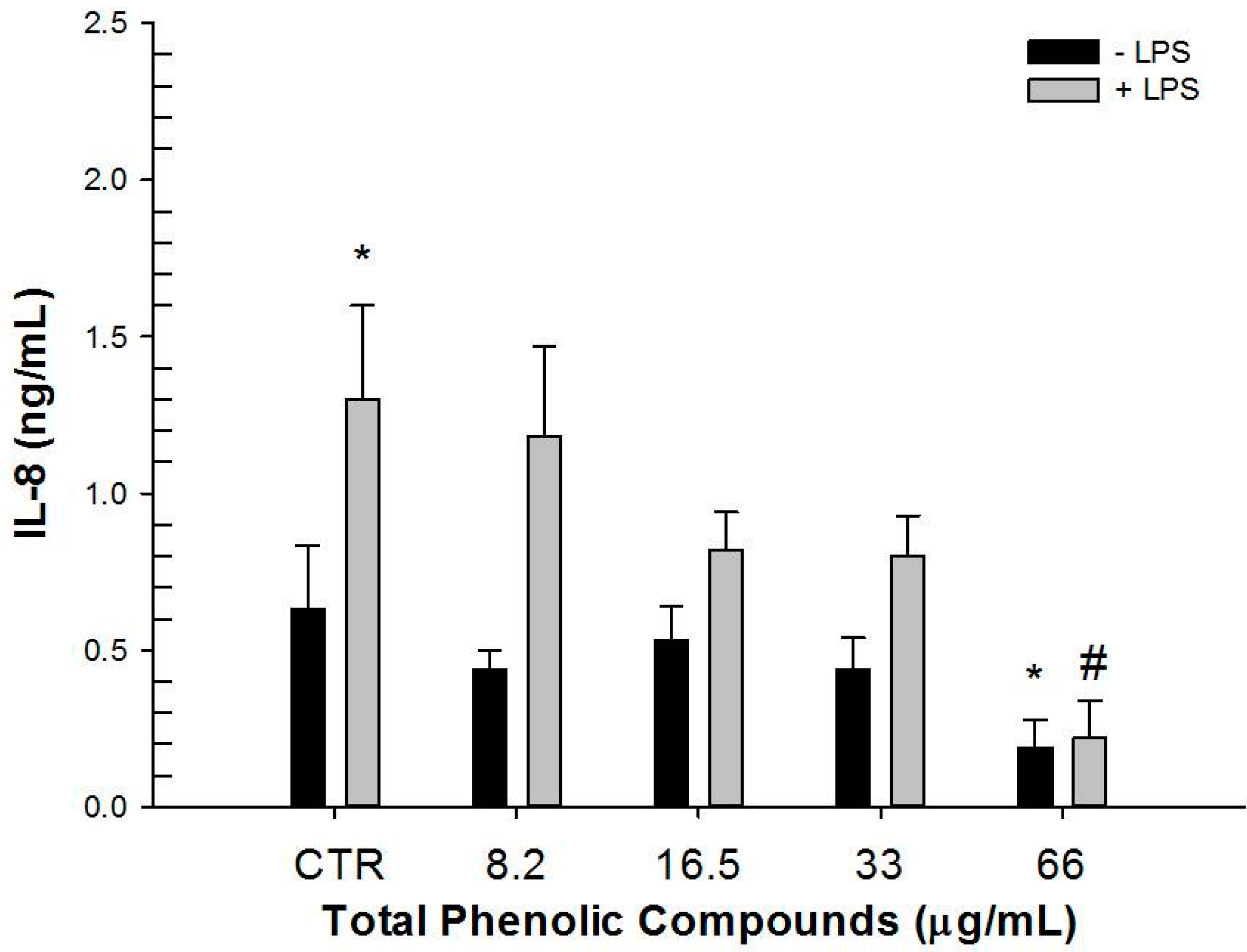
| IL-8 (ng/mL) | ||
|---|---|---|
| SVEVO Total Isoprenoids (μg/mL) | −LPS | +LPS |
| CTR | 0.33 ± 0.02 a | 1.77 ± 0.17 b |
| 0.01 | 0.37 ± 0.09 | 1.42 ± 0.27 |
| 0.02 | 0.36 ± 0.06 | 1.80 ± 0.40 |
| 0.1 | 0.31 ± 0.07 | 1.88 ± 0.23 |
| 0.2 | 0.34 ± 0.04 | 1.34 ± 0.33 * |
3. Experimental Section
3.1. Wheat Samples
3.2. Extraction of Phenolic Acids from Whole-Meal Flour and HPLC Analysis
3.3. Extraction of Isoprenoids from Whole-Meal Flour and HPLC Analysis
3.4. Determination of the Trolox Equivalent Antioxidant Capacity (TEAC)
3.5. Quantification of IL-8 and TGF-β Produced by HT-29 Cell Line with and without LPS Stimulation
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Notes
Conflicts of Interest
References
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Lenucci, M.S.; Rescio, L.; Mita, G.; Caretto, S. Durum wheat by-products as natural sources of valuable nutrients. Phytochem. Rev. 2012, 11, 255–262. [Google Scholar] [CrossRef]
- Žilić, S.; Serpen, A.; Akıllıoğlu, G.; Janković, M.; Gökmen, V. Distributions of phenolic compounds, yellow pigments and oxidative enzymes in wheat grains and their relation to antioxidant capacity of bran and debranned flour. J. Cereal Sci. 2012, 56, 652–658. [Google Scholar] [CrossRef]
- Ciccoritti, R.; Carbone, K.; Bellato, S.; Pogna, N.; Sgrulletta, D. Content and relative composition of some phytochemicals in diploid, tetraploid and hexaploid Triticum species with potential nutraceutical properties. J. Cereal Sci. 2013, 57, 200–206. [Google Scholar] [CrossRef]
- Lampi, A.M.; Nurmi, T.; Ollilainen, V.; Piironen, V. Tocopherols and tocotrienols in wheat genotypes in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2008, 56, 9716–9721. [Google Scholar] [CrossRef]
- Digesù, A.M.; Platani, C.; Cattivelli, L.; Mangini, G.; Blanco, A. Genetic variability in yellow pigment components in cultivated and wild tetraploid wheats. J. Cereal Sci. 2009, 50, 210–218. [Google Scholar] [CrossRef]
- Shewry, P.R.; Piironen, V.; Lampi, A.M.; Edelmann, M.; Kariluoto, S.; Nurmi, T.; Fernandez-Orozco, R.; Ravel, C.; Charmet, G.; Andersson, A.A.M.; et al. The HEALTHGRAIN wheat diversity screen: Effects of genotype and environment on phytochemicals and dietary fiber components. J. Agric. Food Chem. 2010, 58, 9291–9298. [Google Scholar] [CrossRef] [PubMed]
- Brenchley, R.; Spannagl, M.; Pfeifer, M.; Barker, G.L.A.; D’Amore, R.; Allen, A.M.; McKenzie, N.; Kramer, M.; Kerhornou, A.; Bolser, D.; et al. Analysis of the bread wheat genome using whole-genome shotgun sequencing. Nature 2012, 491, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Rawat, N.; Laddomada, B.; Gill, B.S. Genomics of cereal-based functional foods. In Cereal Genomics II; Varshney, R.K., Gupta, P.K., Eds.; Springer Sciences Business Media: Dordrecht, The Netherlands, 2013; pp. 247–274. [Google Scholar]
- Rampino, P.; Mita, G.; Fasano, P.; Borrelli, G.M.; Aprile, A.; Dalessandro, G.; de Bellis, L.; Perrotta, C. Novel durum wheat genes up-regulated in response to a combination of heat and drought stress. Plant Physiol. Biochem. 2012, 56, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Brandolini, A.; Castoldi, P.; Plizzari, L.; Hidalgo, A. Phenolic acids composition, total polyphenols content and antioxidant activity of Triticum monococcum, Triticum turgidum and Triticum aestivum: A two-years evaluation. J. Cereal Sci. 2013, 58, 123–131. [Google Scholar] [CrossRef]
- Li, L.; Shewry, P.R.; Ward, J.L. Phenolic acids in wheat varieties in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2008, 56, 9732–9739. [Google Scholar] [CrossRef] [PubMed]
- Vitaglione, P.; Napolitano, A.; Fogliano, V. Cereal dietary fibre: A natural functional ingredient to deliver phenolic compounds into the gut. Trends Food Sci. Technol. 2008, 19, 451–463. [Google Scholar] [CrossRef]
- Lachman, J.; Hejtmánková, K.; Kotíková, Z. Tocols and carotenoids of einkorn, emmer and spring wheat varieties: Selection for breeding and production. J. Cereal Sci. 2013, 57, 207–214. [Google Scholar] [CrossRef]
- Liu, R.H. Whole grain phytochemicals and health. J. Cereal. Sci. 2007, 46, 207–219. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Gianotti, A.; Danesi, F.; Verardo, V.; Serrazanetti, D.I.; Valli, V.; Russo, A.; Riciputi, Y.; Tossani, N.; Caboni, M.F.; Guerzoni, M.E.; et al. Role of cereal type and processing in whole grain in vivo protection from oxidative stress. Front. Biosci. 2011, 16, 1609–1618. [Google Scholar] [CrossRef]
- Harada, A.; Mukaida, N.; Matsushima, K. Interleukin 8 as a novel target for intervention therapy in acute inflammatory diseases. Mol. Med. Today 1996, 2, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, T.; Matsushima, K.; Tanaka, S.; Robinson, E.A.; Appella, E.; Oppenheim, J.J.; Leonard, E.J. Purification of a human monocyte-derived neutrophil chemotactic factor that has peptide sequence similarity to other host defense cytokines. Proc. Natl. Acad. Sci. USA 1987, 84, 9233–9237. [Google Scholar] [CrossRef] [PubMed]
- Seitz, M.; Dewald, B.; Gerber, N.; Baggiolini, M. Enhanced production of neutrophil-activating peptide-1/interleukin-8 in rheumatoid arthritis. J. Clin. Investig. 1991, 87, 463–469. [Google Scholar] [CrossRef]
- Grimm, M.C.; Elsbury, S.K.; Pavli, P.; Doe, W.F. Interleukin 8: Cells of origin in inflammatory bowel disease. Gut 1996, 38, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Schroder, J.M.; Gregory, H.; Young, J.; Christophers, E. Neutrophil activating proteins in psoriasis. J. Investig. Dermatol. 1992, 98, 241–247. [Google Scholar] [CrossRef]
- Mahida, Y.R.; Ceska, M.; Effenberger, F.; Kurlak, L.; Lindley, I.; Hawkey, C.J. Enhanced synthesis of neutrophil-activating peptide-1/interleukin-8 in active ulcerative colitis. Clin. Sci. 1992, 82, 273–275. [Google Scholar] [PubMed]
- Gibson, P.; Rosella, O. Interleukin 8 secretion by colonic crypt cells in vitro: Response to injury suppressed by butyrate and enhanced in inflammatory bowel disease. Gut 1995, 37, 536–543. [Google Scholar] [CrossRef]
- Yomogida, S.; Hua, J.; Tsutsumi-Ishii, Y.; Sakamoto, K.; Nagaoka, I. Effect of glucosamine on interleukin-8 production from human colonic epithelial cell line. Inflamm. Regen. 2006, 26, 513–518. [Google Scholar] [CrossRef]
- Rieder, F.; Biancani, P.; Harnett, K.; Yerian, L.; Falk, J.W. Inflammatory mediators in gastroesophageal reflux disease: Impact on esophageal motility, fibrosis, and carcinogenesis. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, 571–581. [Google Scholar] [CrossRef]
- Drygiannakis, I.; Valatas, V.; Sfakianaki, O.; Bourikas, L.; Manousou, P.; Kambas, K.; Ritis, K.; Kolios, G.; Kouroumalis, E. Proinflammatory cytokines induce crosstalk between colonic epithelial cells and subepithelial myofibroblasts: Implication in intestinal fibrosis. J. Crohns Colitis 2013, 7, 286–300. [Google Scholar] [CrossRef]
- Bahrami, B.; Macfarlane, S.; Macfarlane, G.T. Induction of cytokine formation by human intestinal bacteria in gut epithelial cell lines. J. Appl. Microbiol. 2010, 110, 353–363. [Google Scholar]
- Hoffmann, P.; Sturm, A.; Stein, J.; Dignass, A.U. Interferon-γ modulates intestinal epithelial cell function in vitro through a TGF-β-dependent mechanism. Regul. Pept. 2011, 168, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, A.J.; Geuking, M.B.; Slack, E.; Hapfelmeier, S.; McCoy, K.D. The habitat, double life, citizenship, and forgetfulness of IgA. Immunol. Rev. 2011, 245, 132–146. [Google Scholar]
- Whent, M.; Huang, H.; Xie, Z.; Lutterodt, H.; Yu, L.; Fuerst, E.P.; Craig, F.; Morris, C.F.; Yu, L.; Luthria, D. Phytochemical composition, anti-inflammatory, and antiproliferative activity of whole wheat flour. J. Agric. Food Chem. 2012, 60, 2129–2135. [Google Scholar] [CrossRef]
- Fernandez-Orozco, R.; Li, L.; Harflett, C.; Shewry, P.R.; Ward, J.L. Effects of environment and genotype on phenolic acids in wheat in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2010, 58, 9341–9352. [Google Scholar] [CrossRef]
- Shewry, P.R.; Charmet, G.; Branlard, G.; Lafiandra, D.; Gergely, S.; Salgó, A.; Saulnier, L.; Bedȍ, Z.; Clare Mills, E.N.; Ward, J.L. Developing new types of wheat with enhanced health benefits. Trends Food Sci. Technol. 2012, 25, 70–77. [Google Scholar] [CrossRef]
- Dinelli, G.; Segura Carretero, A.; di Silvestro, R.; Marotti, I.; Fu, S.; Benedettelli, S.; Ghiselli, L.; Gutierrez, A.F. Determination of phenolic compounds in modern and old varieties of durum wheat using liquid chromatography coupled with time-of-flight mass spectrometry. J. Chromatogr. A 2009, 1216, 7229–7240. [Google Scholar]
- Menga, V.; Fares, C.; Troccoli, A.; Cattivelli, A.; Baiano, A. Effects of genotype, location and baking on the phenolic content and some antioxidant properties of cereal species. Int. J. Food Sci. Technol. 2010, 45, 7–16. [Google Scholar]
- Abdel-Aal, E.M.; Rabalski, I. Bioactive compounds and their antioxidant capacity in selected primitive and modern wheat species. Open Agric. J. 2008, 2, 7–14. [Google Scholar] [CrossRef]
- Chowers, Y.; Cahalon, L.; Lahav, M.; Schor, H.; Tal, R.; Bar-Meir, S.; Levite, M. Somatostatin through its specific receptor inhibits spontaneous and TNFα and bacteria-induced IL-8 and IL-1β secretion from intestinal epithelial cells. J. Immunol. 2000, 165, 2955–2961. [Google Scholar] [CrossRef]
- Dwinell, M.B.; Eckmann, L.; Leopard, J.D.; Varki, N.M.; Kagnoff, M.F. Chemokine receptor expression by human intestinal epithelial cells. Gastroenterology 1999, 177, 359–387. [Google Scholar] [CrossRef]
- DeForge, L.E.; Fantone, J.C.; Kenney, J.S.; Remick, D.G. Oxygen radical scavengers selectively inhibit interleukin 8 production in human whole blood. J. Clin. Investig. 1992, 90, 2123–2129. [Google Scholar] [CrossRef]
- Massion, P.P.; Lindén, A.; Inoue, H.; Mathy, M.; Grattan, K.M.; Nadel, J.A. Dimethyl sulfoxide decreases interleukin-8-mediated neutrophil recruitment in the airways. Am. J. Physiol. 1996, 271, 838–843. [Google Scholar]
- Hollebeeck, S.; Raas, T.; Piront, N.; Schneider, Y.J.; Toussaint, O.; Larondelle, Y.; During, A. Dimethyl sulfoxide (DMSO) attenuates the inflammatory response in the in vitro intestinal Caco-2 cell model. Toxicol. Lett. 2011, 206, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Mateo Anson, N.; Aura, A.; Selinheimo, E.; Mattila, I.; Poutanen, K.; van den Berg, R.; Havenaar, R.; Bast, A.; Haenen, G.R.M.M. Bioprocessing of wheat bran in whole wheat bread increases the bioavailability of phenolic acids in men and exerts antiinflammatory effects ex vivo. J. Nutr. Immunol. 2011, 141, 137–143. [Google Scholar] [CrossRef]
- Lotito, S.B.; Zhang, W.J.; Yang, C.S.; Crozier, A.; Frei, B. Metabolic conversion of dietary flavonoids alters their anti-inflammatory and antioxidant properties. Free Radic. Biol. Med. 2011, 51, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Kroon, P.A.; Clifford, M.N.; Crozier, A.; Day, A.J.; Donovan, J.L.; Manach, C.; Williamson, G. How should we assess the effects of exposure to dietary polyphenols in vitro? Am. J. Clin. Nutr. 2004, 80, 15–21. [Google Scholar] [PubMed]
- Manak, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [PubMed]
- Yam, M.L.; Hafid, S.R.A.; Cheng, H.M.; Nesaretnam, K. Tocotrienols suppress proinflammatory markers and cyclooxygenase-2 expression in RAW264.7 macrophages. Lipids 2009, 44, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Mstatc-C Software, Version 2.6. Drinkwater N, Crop and Soil Science Department, Michigan State University: East Lansing, MI, USA, 1998.
- Panfili, G.; Manzi, P.; Pizzoferrato, L. High performance liquid chromatographic method for the simultaneous determination of tocopherols, carotenes and retinol and its geometric isomers in Italian cheese. Analyst 1994, 119, 1161–1165. [Google Scholar] [CrossRef] [PubMed]
- Fraser, P.D.; Pinto, M.S.E.; Holloway, D.E.; Bramley, P.M. Application of high-performance liquid chromatography with photodiode array detection to the metabolic profiling of plant isoprenoids. Plant J. 2000, 24, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Minervini, F.; Fornelli, F.; Lucivero, G.; Romano, C.; Visconti, A. T-2 toxin immunotoxicity on human B and T lymphoid cell lines. Toxicology 2005, 210, 81–91. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laddomada, B.; Durante, M.; Minervini, F.; Garbetta, A.; Cardinali, A.; D'Antuono, I.; Caretto, S.; Blanco, A.; Mita, G. Phytochemical Composition and Anti-Inflammatory Activity of Extracts from the Whole-Meal Flour of Italian Durum Wheat Cultivars. Int. J. Mol. Sci. 2015, 16, 3512-3527. https://doi.org/10.3390/ijms16023512
Laddomada B, Durante M, Minervini F, Garbetta A, Cardinali A, D'Antuono I, Caretto S, Blanco A, Mita G. Phytochemical Composition and Anti-Inflammatory Activity of Extracts from the Whole-Meal Flour of Italian Durum Wheat Cultivars. International Journal of Molecular Sciences. 2015; 16(2):3512-3527. https://doi.org/10.3390/ijms16023512
Chicago/Turabian StyleLaddomada, Barbara, Miriana Durante, Fiorenza Minervini, Antonella Garbetta, Angela Cardinali, Isabella D'Antuono, Sofia Caretto, Antonio Blanco, and Giovanni Mita. 2015. "Phytochemical Composition and Anti-Inflammatory Activity of Extracts from the Whole-Meal Flour of Italian Durum Wheat Cultivars" International Journal of Molecular Sciences 16, no. 2: 3512-3527. https://doi.org/10.3390/ijms16023512