
A Ribosomal Protein AgRPS3aE from Halophilic Aspergillus glaucus Confers Salt Tolerance in Heterologous Organisms

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. AgRPS3aE Gene and Protein Sequences from A. glaucus
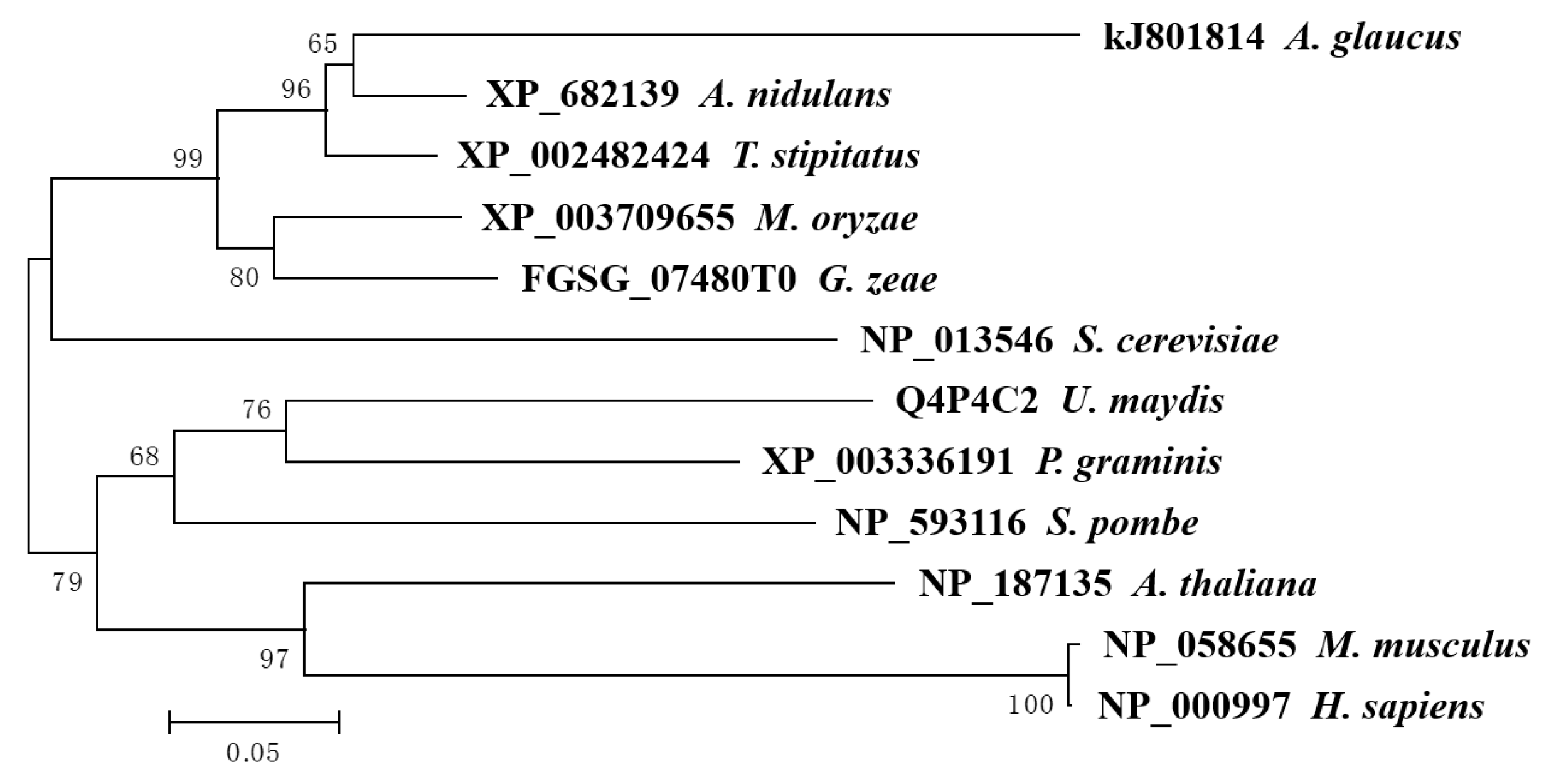
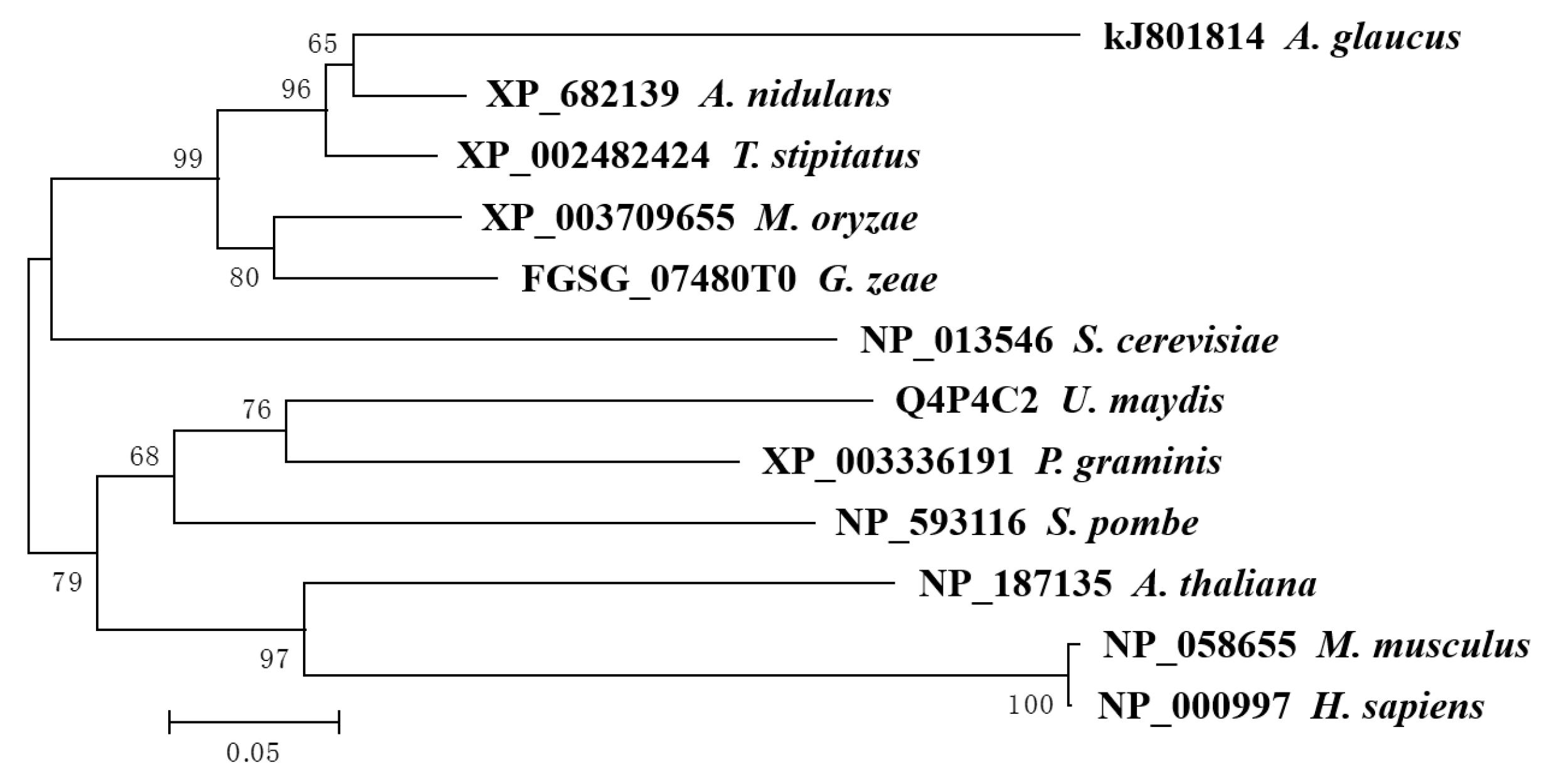
2.2. Evolution and Primary Structure of the Ribosomal Protein AgRPS3aE

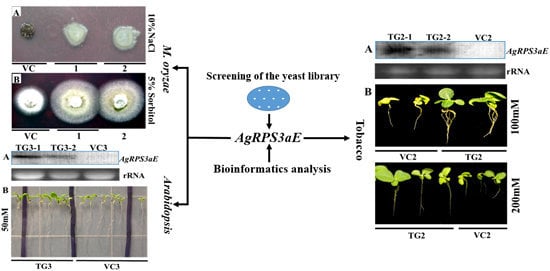
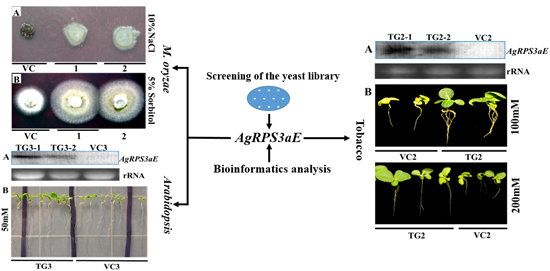
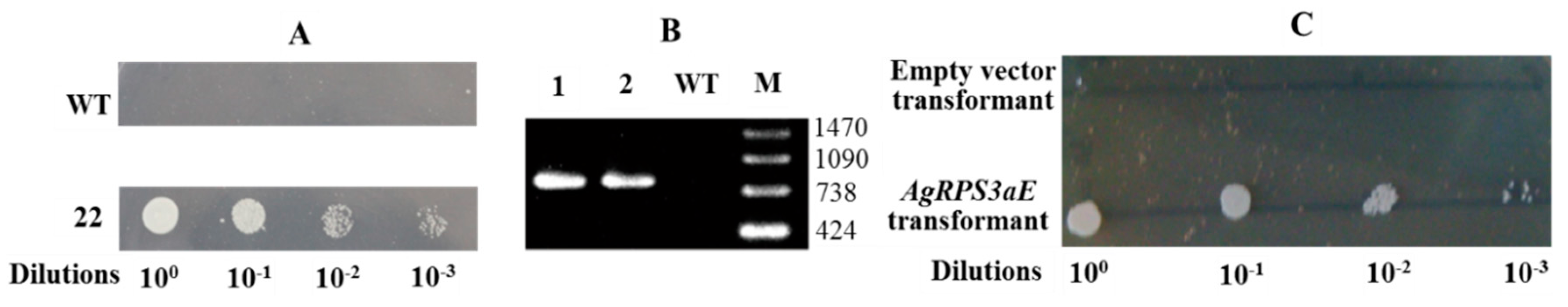
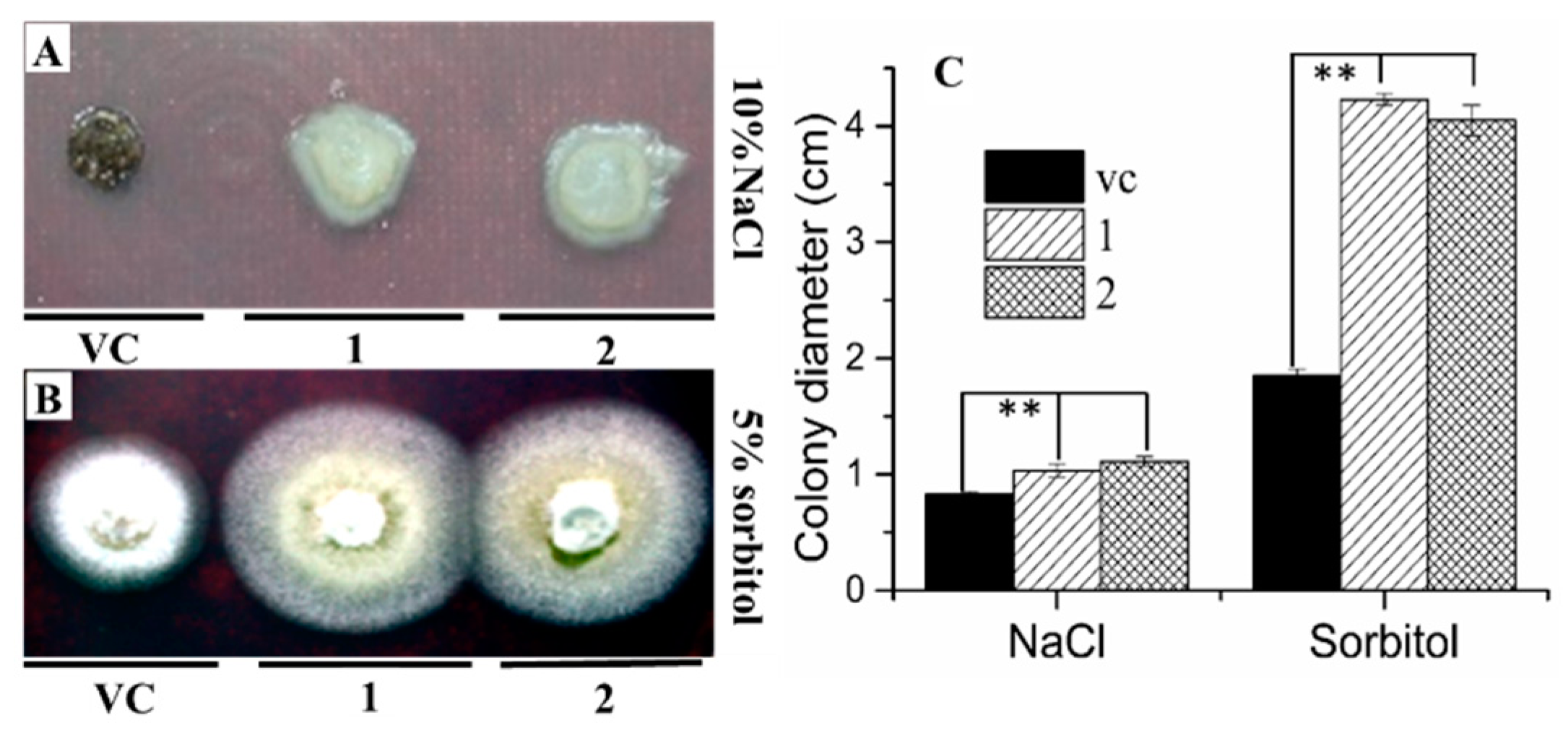
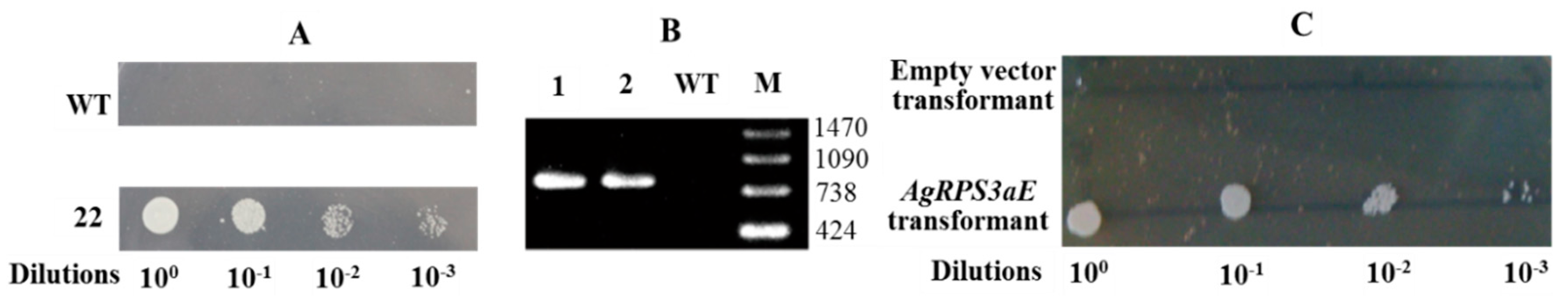
2.3. AgRPS3aE Confers Tolerance to Osmotic Stress in M. oryzae
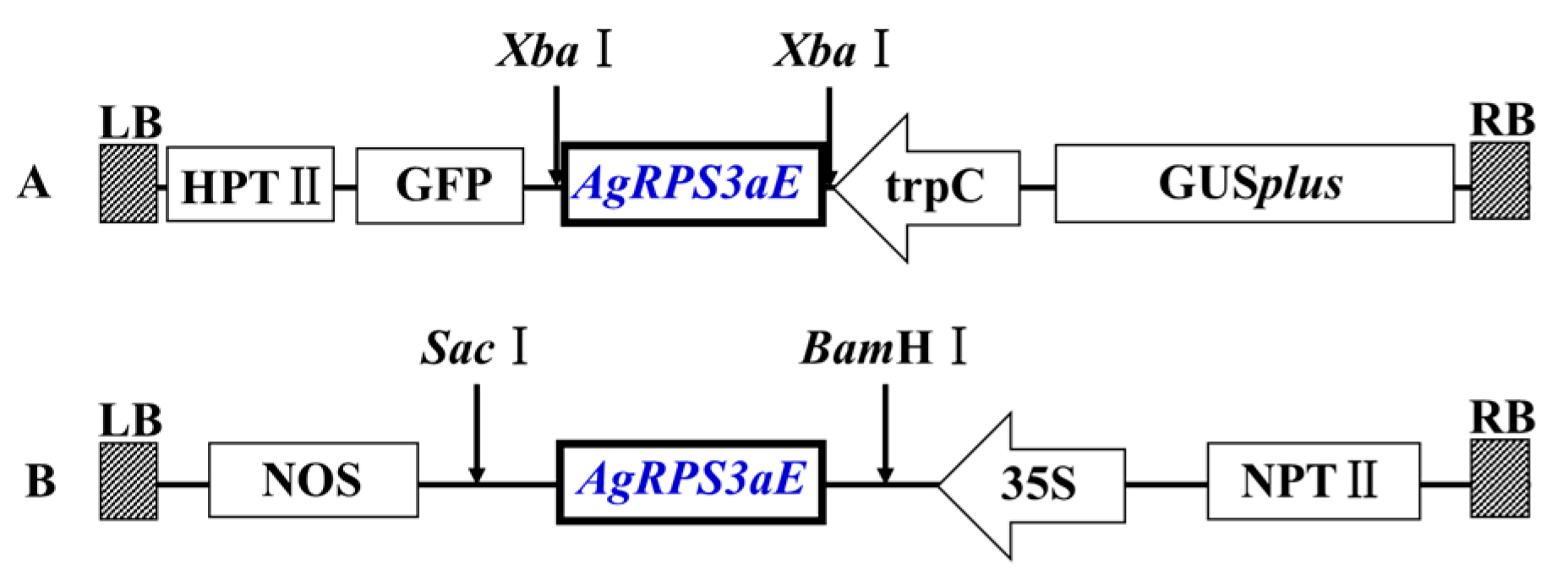
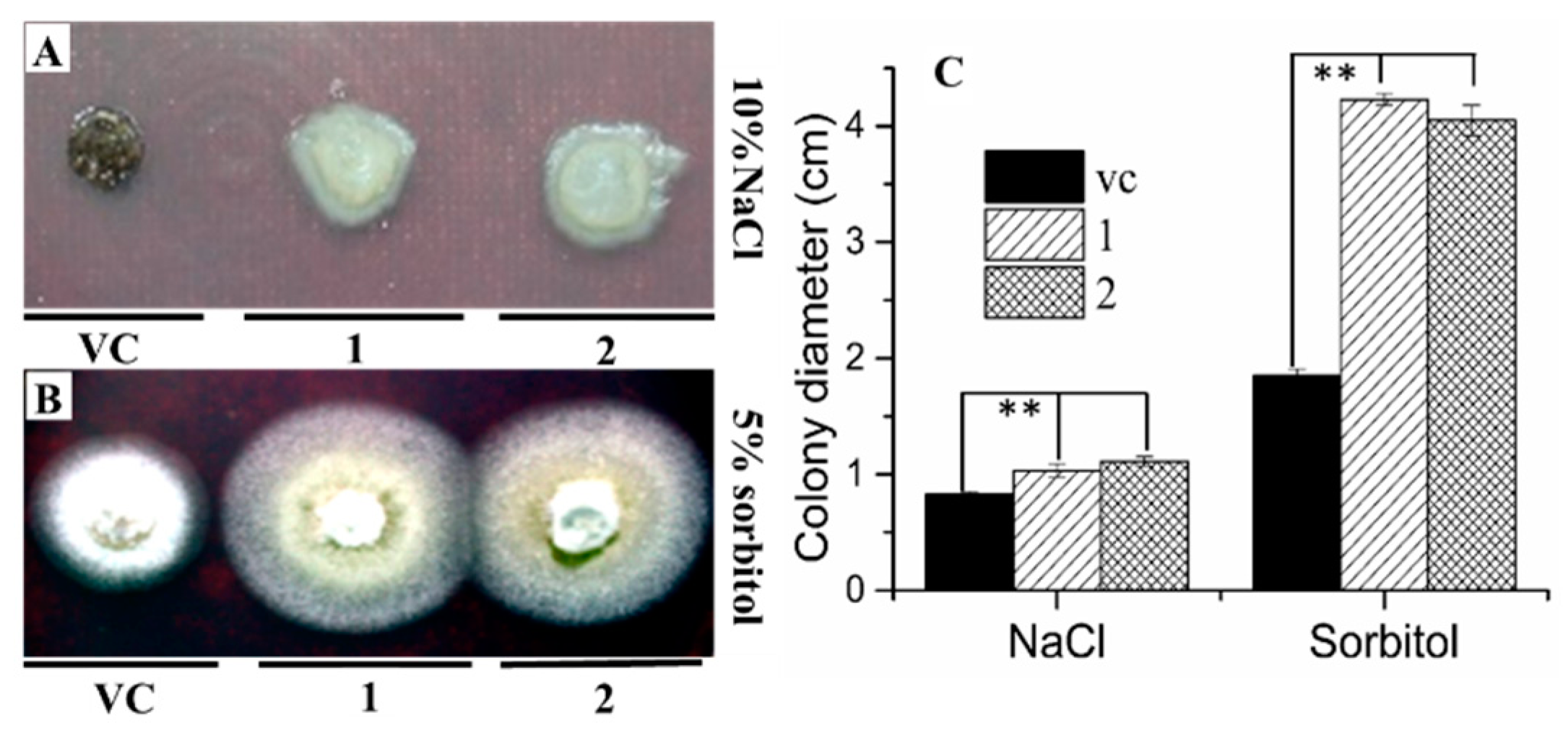
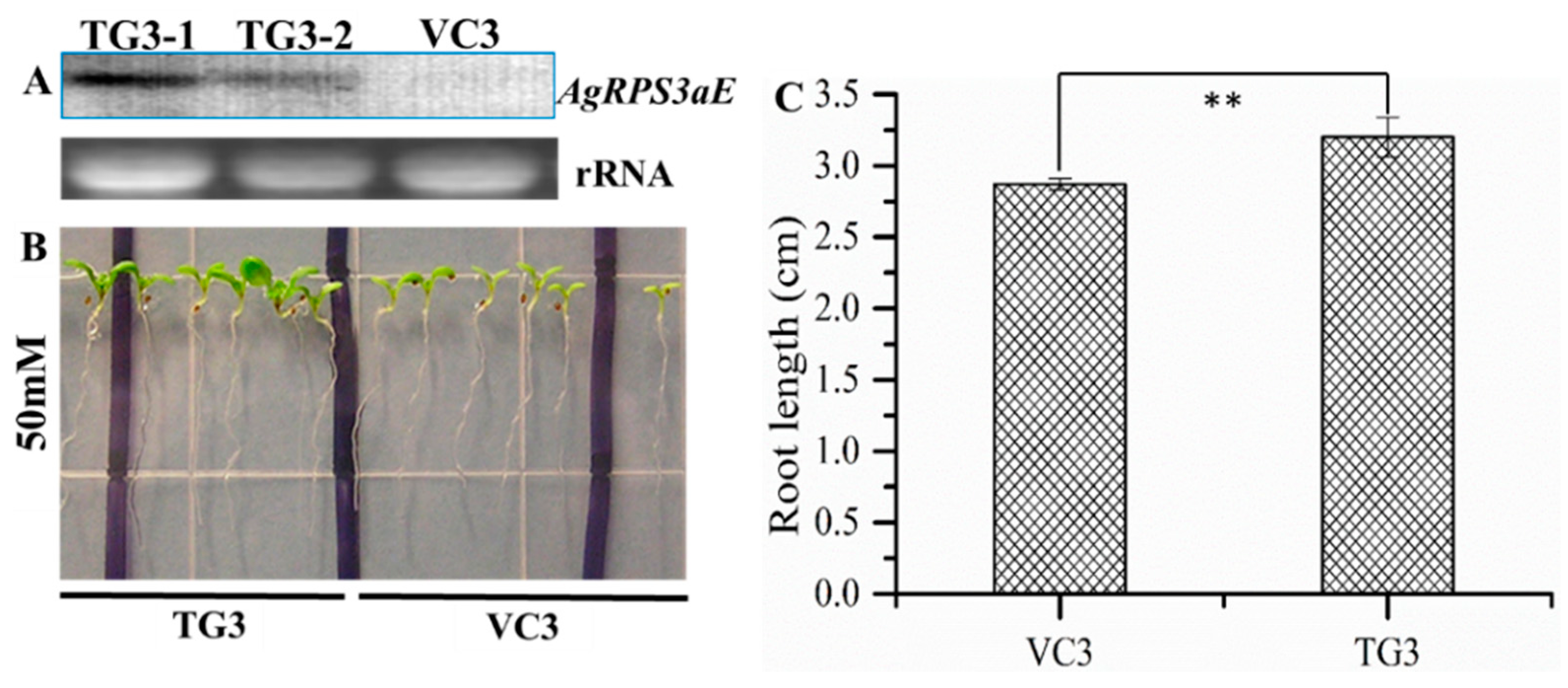
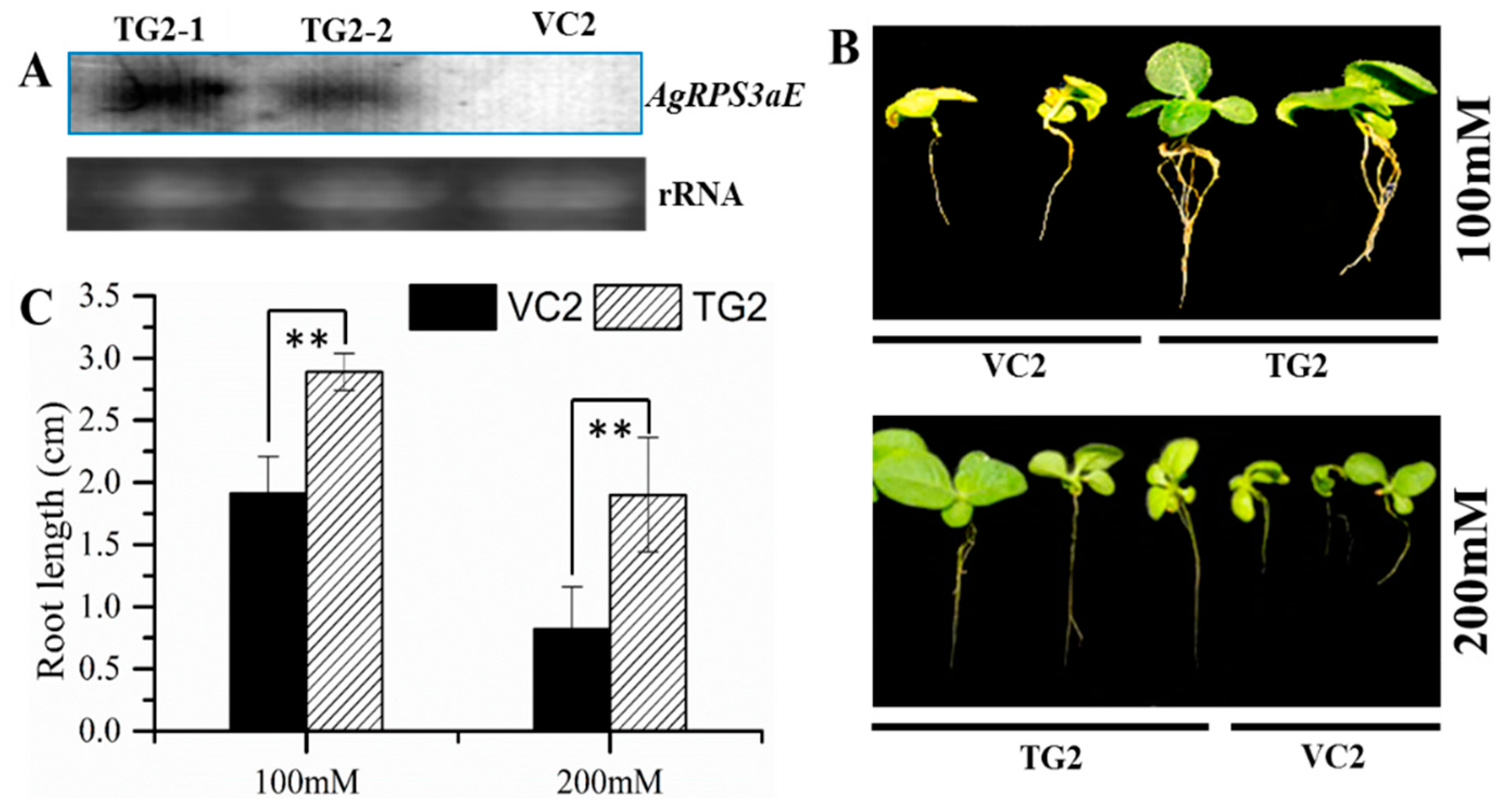
2.4. Overexpression of AgRPS3aE Enhances Salt Tolerance of Plants

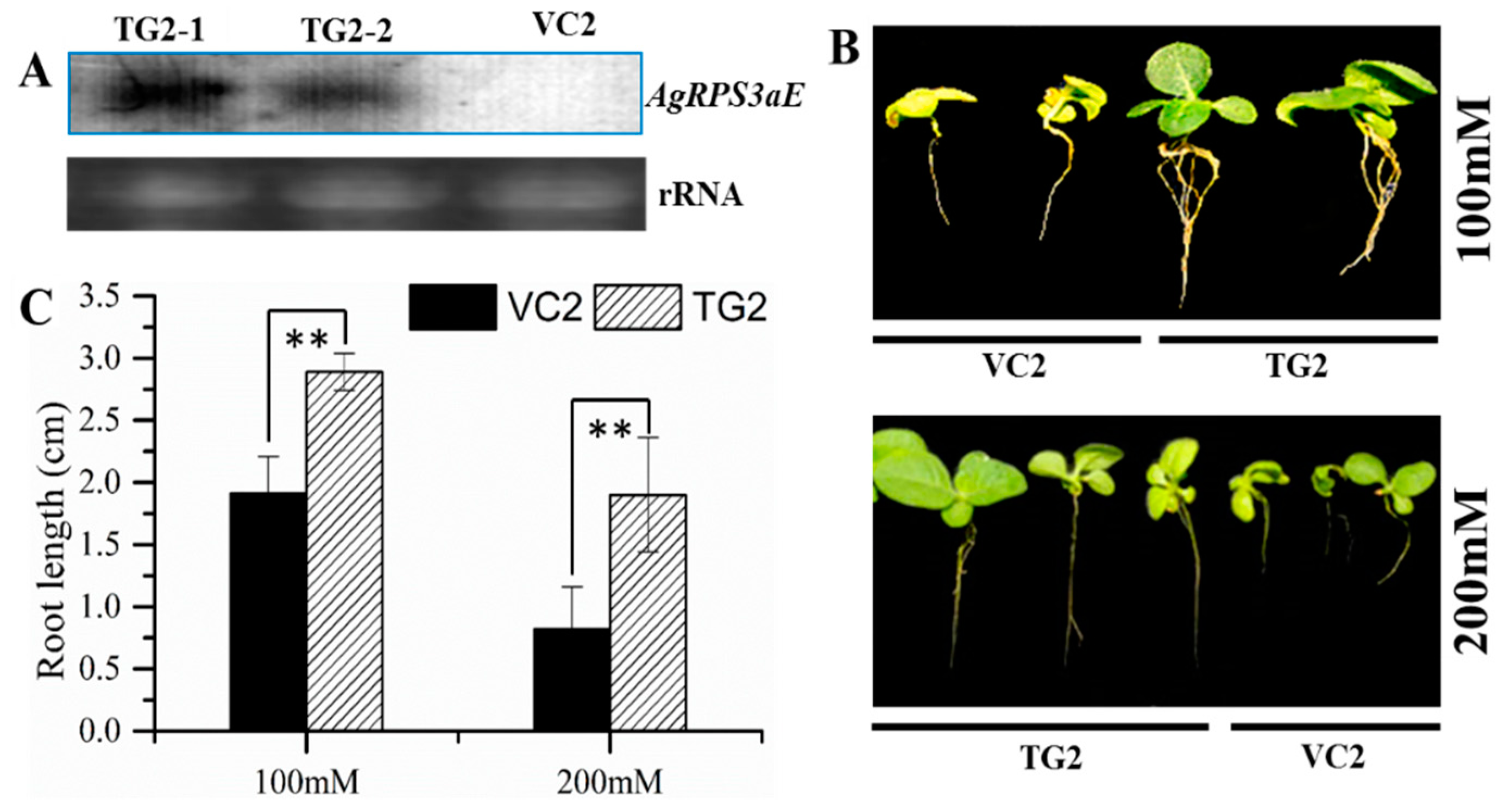
3. Discussion
4. Experimental Section
4.1. Organisms and Growth Conditions
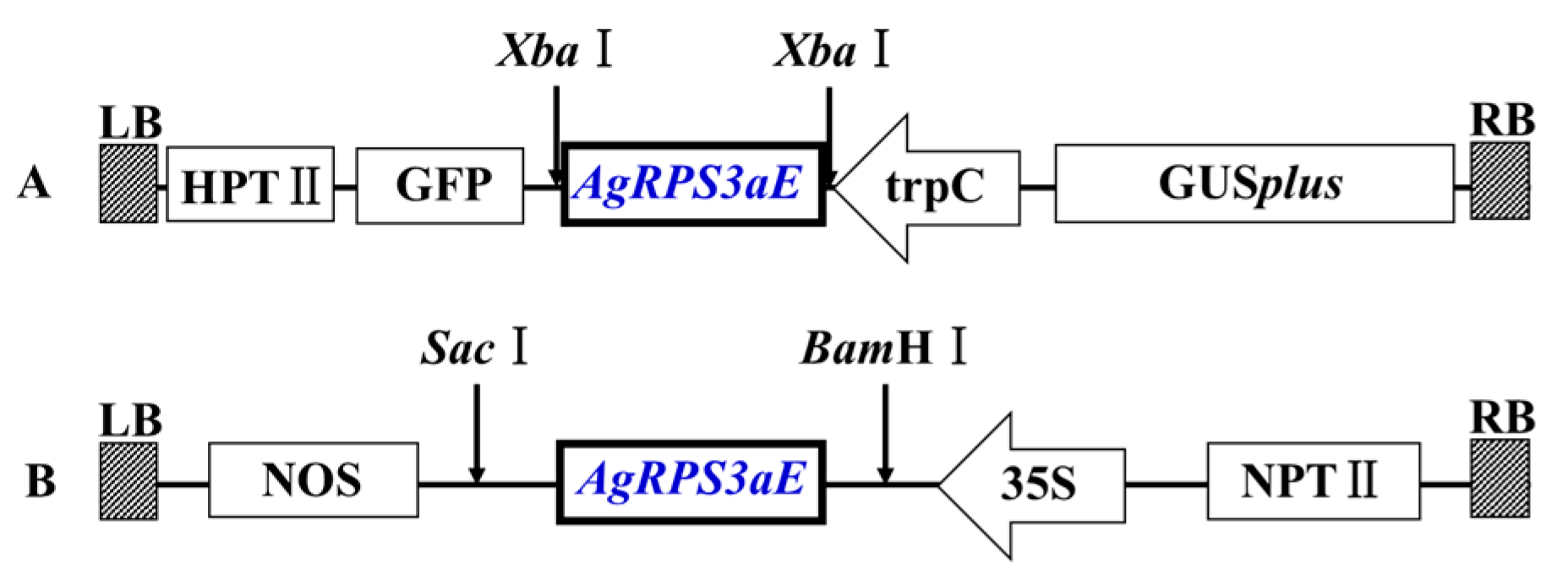
4.2. Cloning and Sequence Analysis of the AgRPS3aE Gene
4.3. Analysis of Osmotic Stress-Tolerance in M. oryzae Containing AgRPS3aE
4.4. Northern Blotting Analysis
4.5. Plant Transformation and Salt Tolerance Analysis
5. Conclusions
Acknowledgments
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Irfan, M.; Ahmad, A.; Hayat, S. Causes of salinity and plant manifestations to salt stress: A review. J. Environ. Biol. 2011, 32, 667–685. [Google Scholar] [PubMed]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Apse, M.P.; Blumwald, E. Na+ transport in plants. FEBS Lett. 2007, 581, 2247–2254. [Google Scholar] [CrossRef] [PubMed]
- Sobhanian, H.; Aghaei, K.; Komatsu, S. Changes in the plant proteome resulting from salt stress: Toward the creation of salt-tolerant crops? J. Proteomics 2011, 74, 1323–1337. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M. Breeding for salinity tolerance in plants. Crit. Rev. Plant Sci. 1994, 13, 17–42. [Google Scholar] [CrossRef]
- Rai, M.K.; Kalia, R.K.; Singh, R.; Gangola, M.P.; Dhawan, A. Developing stress tolerant plants through in vitro selection—An overview of the recent progress. Environ. Exp. Bot. 2011, 71, 89–98. [Google Scholar] [CrossRef]
- Owens, S. Salt of the earth. EMBO Rep. 2001, 2, 877–879. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Leslie, P.; Zhang, Y. Ribosomal proteins as unrevealed caretakers for cellular stress and genomic instability. Oncotarget 2014, 5, 860–871. [Google Scholar] [PubMed]
- Warner, J.R.; McIntosh, K.B. How common are extraribosomal functions of ribosomal proteins? Mol. Cell 2009, 34, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Navakanitworakul, R.; Deachamag, P.; Wonglapsuwan, M.; Chotigeat, W. The roles of ribosomal protein S3a in ovarian development of Fenneropenaeus merguiensis (De Man). Aquaculture 2012, 338, 208–215. [Google Scholar] [CrossRef]
- Yusupova, G.; Yusupov, M. High-resolution structure of the eukaryotic 80S ribosome. Annu. Rev. Biochem. 2014, 83, 467–486. [Google Scholar] [CrossRef] [PubMed]
- Kho, C.J.; Wang, Y.; Zarbl, H. Effect of decreased fte-1 gene expression on protein synthesis, cell growth, and transformation. Cell Growth Differ. 1996, 7, 1157–1166. [Google Scholar] [PubMed]
- Naora, H.; Naora, H. Involvement of ribosomal proteins in regulating cell growth and apoptosis: Translational modulation or recruitment for extraribosomal activity? Immunol. Cell Biol. 1999, 77, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Naora, H.; Takai, I.; Adachi, M.; Naora, H. Altered cellular responses by varying expression of a ribosomal protein gene: Sequential coordination of enhancement and suppression of ribosomal protein S3a gene expression induces apoptosis. J. Cell Biol. 1998, 141, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Ruggero, D.; Pandolfi, P.P. Does the ribosome translate cancer? Nat. Rev. Cancer 2003, 3, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Russell, L.; Naora, H.; Naora, H. Down-regulated RPS3a/nbl expression during retinoid-induced differentiation of HL-60 Cells. A close association with diminished susceptibility to Actinomycin D-stimulated apoptosis. Cell Struct. Funct. 2000, 25, 103–113. [Google Scholar]
- Zapata, M.; Tanguy, A.; David, E.; Moraga, D.; Riquelme, C. Transcriptomic response of Argopecten purpuratus post-larvae to copper exposure under experimental conditions. Gene 2009, 442, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.; Vuong, T.; VanToai, T.; Lee, J.; Wu, X.; Mian, M.; Dorrance, A.; Shannon, J.; Nguyen, H. Mapping of quantitative trait loci associated with resistance to and flooding tolerance in soybean. Crop Sci. 2012, 52, 2481–2493. [Google Scholar] [CrossRef]
- Liu, X.D.; Xie, L.; Wei, Y.; Zhou, X.; Jia, B.; Liu, J.; Zhang, S. Abiotic stress resistance, a novel moonlighting function of ribosomal protein RPL44 in halophilic Aspergillus glaucus. Appl. Environ. Microb. 2014, 80, 4294–4300. [Google Scholar] [CrossRef]
- Sigrist, C.J.A.; Cerutti, L.; de Castro, E.; Langendijk-Genevaux, P.S.; Bulliard, V.; Bairoch, A.; Hulo, N. PROSITE, a protein domain database for functional characterization and annotation. Nucleic Acids Res. 2010, 38, D161–D166. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.F.; Zhao, L.N.; Hu, B.; Chen, X.W.; Zhang, F.; Zhu, Z.M.; Chen, D.F. Identifying novel salt-tolerant genes from Dunaliella salina using a Haematococcus pluvialis expression system. Plant Cell Tissue Organ 2014, 117, 113–124. [Google Scholar] [CrossRef]
- Parker, M.S.; Sah, R.; Balasubramaniam, A.; Sallee, F.R.; Park, E.A.; Parker, S.L. On the expansion of ribosomal proteins and RNAs in eukaryotes. Amino Acids 2014, 46, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Vásquez, J.; Gallois, P.; Delseny, M. Accumulation and nuclear targeting of BnC24, a Brassica napus ribosomal protein corresponding to a mRNA accumulating in response to cold treatment. Plant Sci. 2000, 156, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Park, S.W.; Chung, Y.S.; Chung, C.H.; Kim, J.I.; Lee, J.H. Molecular cloning of low-temperature-inducible ribosomal proteins from soybean. J. Exp. Bot. 2004, 55, 1153–1155. [Google Scholar] [CrossRef] [PubMed]
- Ferreyra, M.L.F.; Pezza, A.; Biarc, J.; Burlingame, A.L.; Casati, P. Plant L10 ribosomal proteins have different roles during development and translation under ultraviolet-B stress. Plant Physiol. 2010, 153, 1878–1894. [Google Scholar] [CrossRef] [PubMed]
- Mok, Y.G.; Lee, B.D.; Kim, Y.J.; Lee, C.E.; Kim, D.G.; Lee, J.; Shim, J.; Meng, Y.; Rosen, B.P.; Choi, J.S. The tobacco gene Ntcyc07 confers arsenite tolerance in Saccharomyces cerevisiae by reducing the steady state levels of intracellular arsenic. FEBS Lett. 2008, 582, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Thorrez, L.; van Deun, K.; Tranchevent, L.C.; van Lommel, L.; Engelen, K.; Marchal, K.; Moreau, Y.; van Mechelen, I.; Schuit, F. Using ribosomal protein genes as reference: A tale of caution. PLoS One 2008, 3, e1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzdin, A.; Gogvadze, E.; Lebrun, M.H. Chimeric retrogenes suggest a role for the nucleolus in LINE amplification. FEBS Lett. 2007, 581, 2877–2882. [Google Scholar] [CrossRef] [PubMed]
- Wee, C.; Lee, S.; Robin, C.; Heckel, D. Identification of candidate genes for fenvalerate resistance in Helicoverpa armigera using cDNA-AFLP. Insect Mol. Biol. 2008, 17, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, Y.; Hu, X.; Yang, Q.; Kang, J.; Zhang, T. Molecular cloning and characterization of a novel gene encoding DREB protein from Buchloe dactyloides (Nutt.) Engelm. J. Agric. Sci. 2011, 4, 12–22. [Google Scholar]
- Gao, J.S.; Hu, L.; Xie, P.; Meng, Y.; Cai, Y.P.; Lin, Y. Isolation and molecular characterization of an ethylene response. J. Biosci. 2014, 39, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Hu, J.J.; Yan, C.F.; Su, H.H.; Ding, J.C.; Guo, Y.Y.; Ye, N.; Huang, S.Q.; Zhang, X.Z.; Zhou, S.F. Characterization of human cytomegalovirus UL145 and UL136 genes in low-passage clinical isolates from infected Chinese infants. Med. Sci. Monit. 2011, 17, CR423–CR431. [Google Scholar] [PubMed]
- Frederich, M.; Roger, B.; David, D.; Seidman, J.; John, A.; Kevin, S. Short Protocols in Molecular Biology; John Wiley & Sons: New York, NY, USA, 1999; pp. 2–12. [Google Scholar]
- Rho, H.S.; Kang, S.; Lee, Y.H. Agrobacterium tumefaciens-mediated transformation of the plant pathogenic fungus, Magnaporthe grisea. Mol. Cells 2001, 12, 407–411. [Google Scholar] [PubMed]
- Liu, J.; Zeng, H.; Li, X.; Xu, L.; Wang, Y.; Tang, W.; Han, L. Isolation and characterization of the betaine aldehyde dehydrogenase gene in Ophiopogon japonicus. Open Biotechnol. J. 2010, 4, 18–25. [Google Scholar] [CrossRef]
- Wang, J.W.; Li, Y.; Zhang, Y.X.; Chai, T.Y. Molecular cloning and characterization of a Brassica juncea yellow stripe-like gene, BjYSL7, whose overexpression increases heavy metal tolerance of tobacco. Plant Cell Rep. 2013, 32, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Gallois, P.; Marinho, P. Leaf disk transformation using Agrobacterium tumefaciens-expression of heterologous genes in tobacco. Methods Mol. Biol. 1995, 49, 39–48. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, X.; Liu, Y.; Xie, L.; Liu, X.; Wei, Y.; Zhou, X.; Zhang, S. A Ribosomal Protein AgRPS3aE from Halophilic Aspergillus glaucus Confers Salt Tolerance in Heterologous Organisms. Int. J. Mol. Sci. 2015, 16, 3058-3070. https://doi.org/10.3390/ijms16023058
Liang X, Liu Y, Xie L, Liu X, Wei Y, Zhou X, Zhang S. A Ribosomal Protein AgRPS3aE from Halophilic Aspergillus glaucus Confers Salt Tolerance in Heterologous Organisms. International Journal of Molecular Sciences. 2015; 16(2):3058-3070. https://doi.org/10.3390/ijms16023058
Chicago/Turabian StyleLiang, Xilong, Yiling Liu, Lixia Xie, Xiaodan Liu, Yi Wei, Xiaoyang Zhou, and Shihong Zhang. 2015. "A Ribosomal Protein AgRPS3aE from Halophilic Aspergillus glaucus Confers Salt Tolerance in Heterologous Organisms" International Journal of Molecular Sciences 16, no. 2: 3058-3070. https://doi.org/10.3390/ijms16023058