
CREB Negatively Regulates IGF2R Gene Expression and Downstream Pathways to Inhibit Hypoxia-Induced H9c2 Cardiomyoblast Cell Death


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
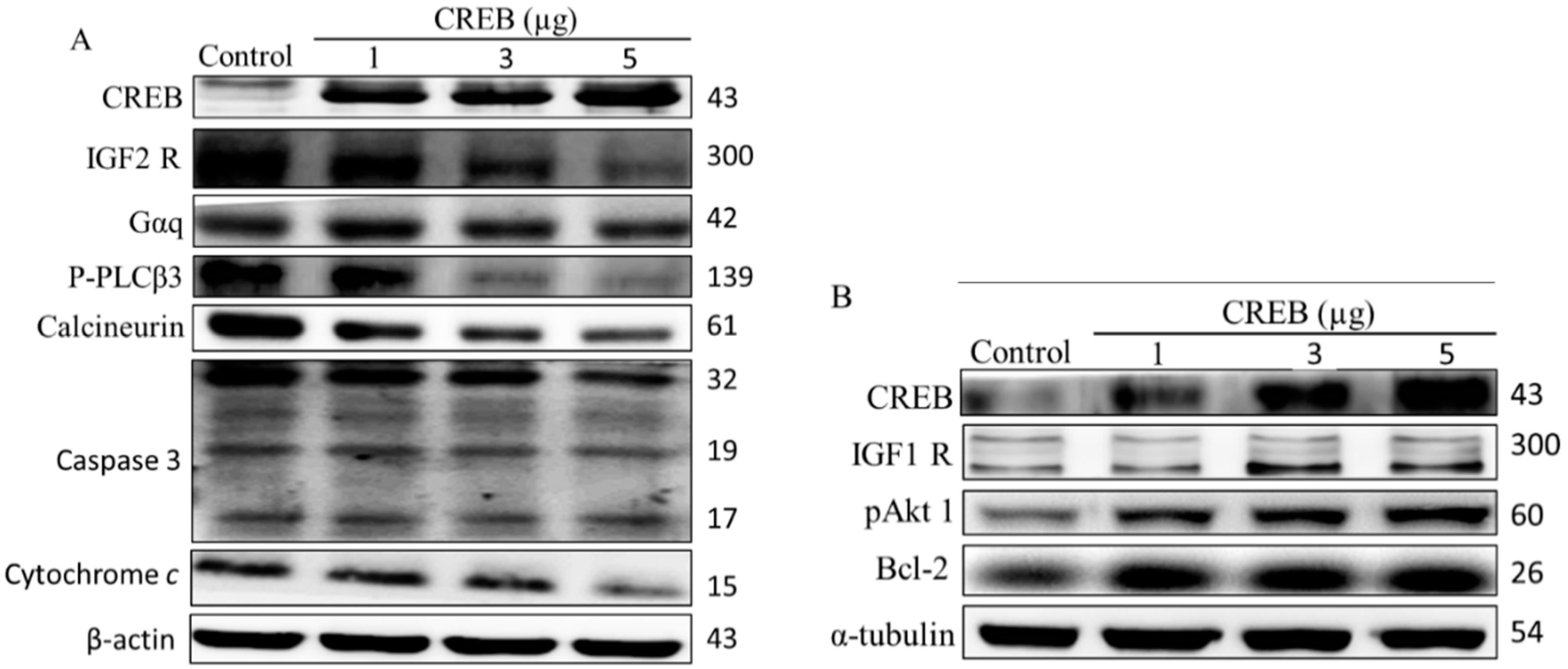
2.1. cAMP Responsive Element-Binding Protein (CREB) Was Involved in Cell Survival in H9c2 Cardiomyoblast Cells
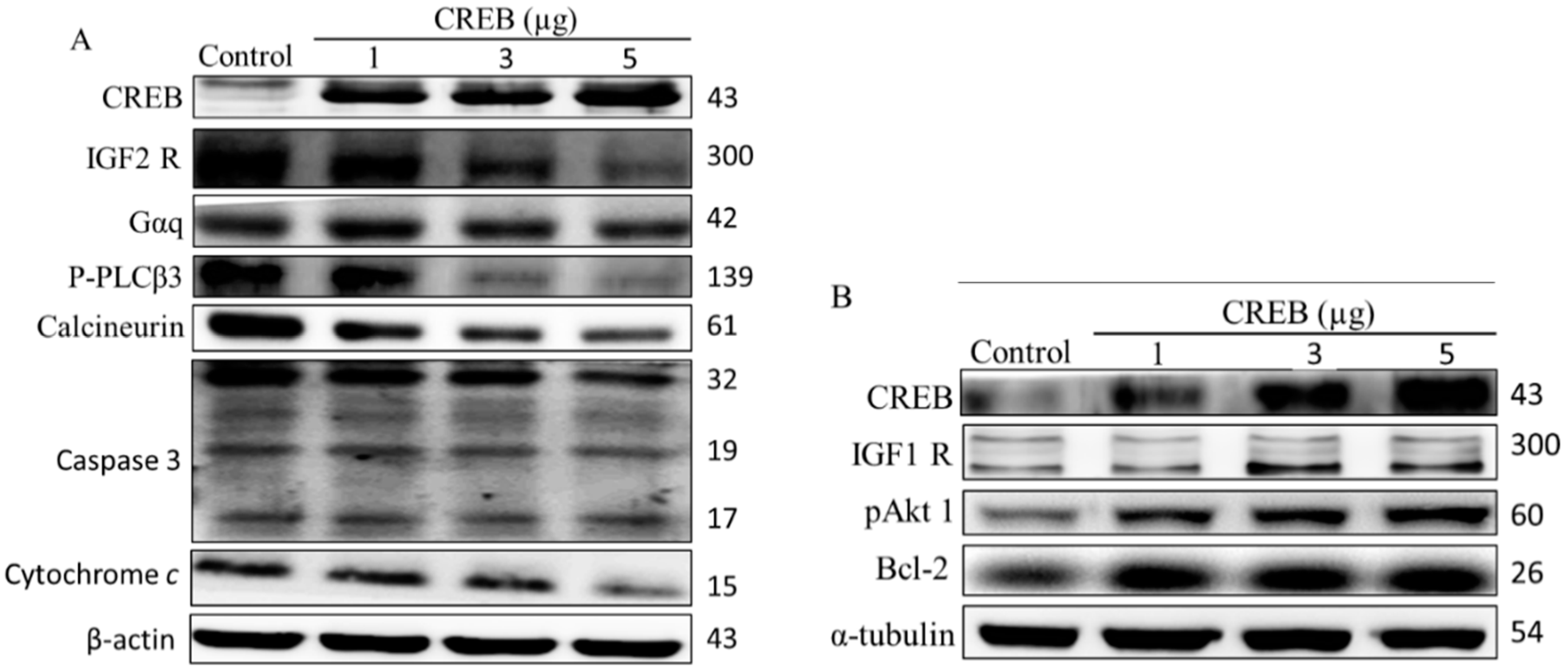
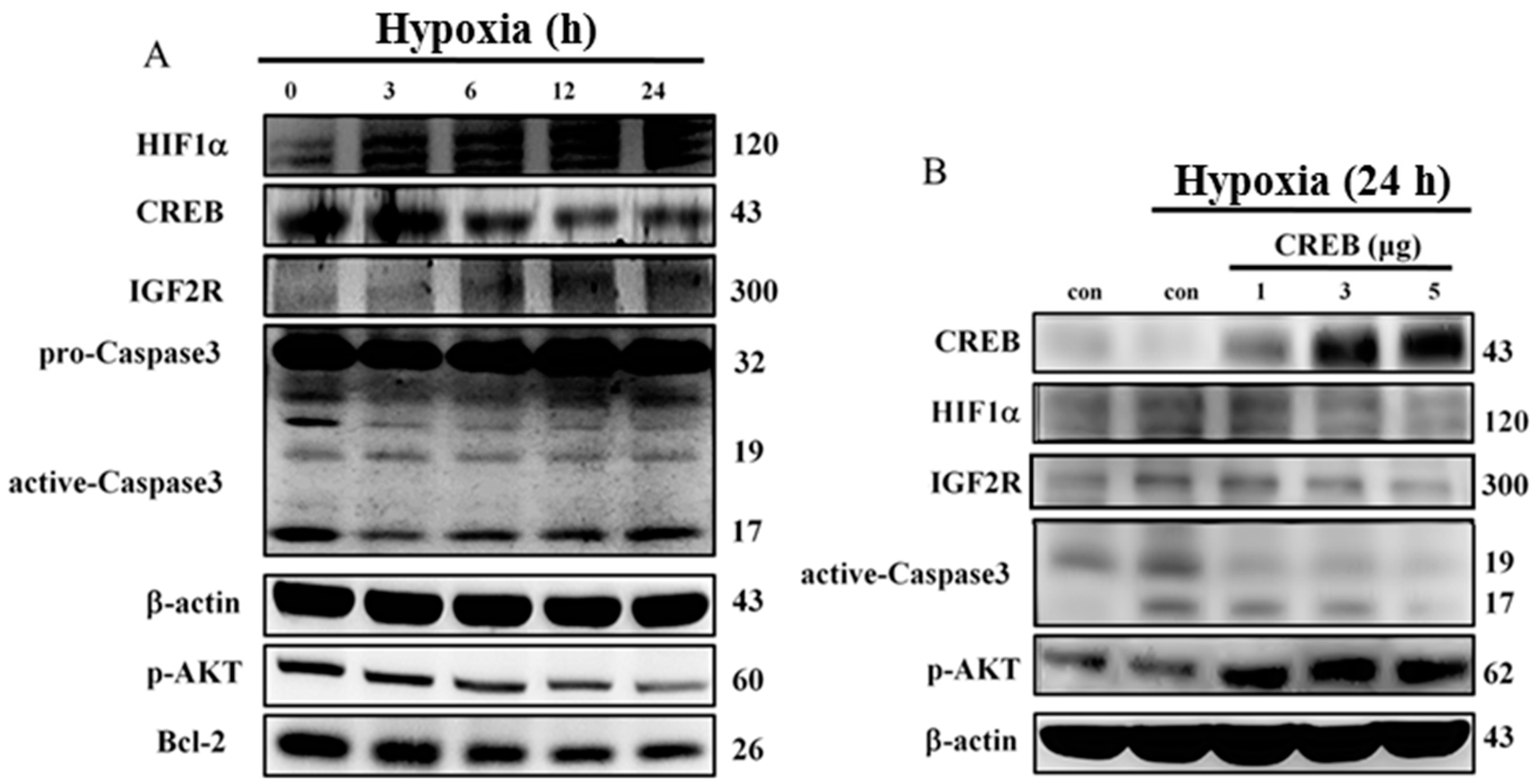
2.2. Hypoxia-Induced Apoptosis Was Suppressed by CREB Transfection in H9c2 Cardiomyoblasts
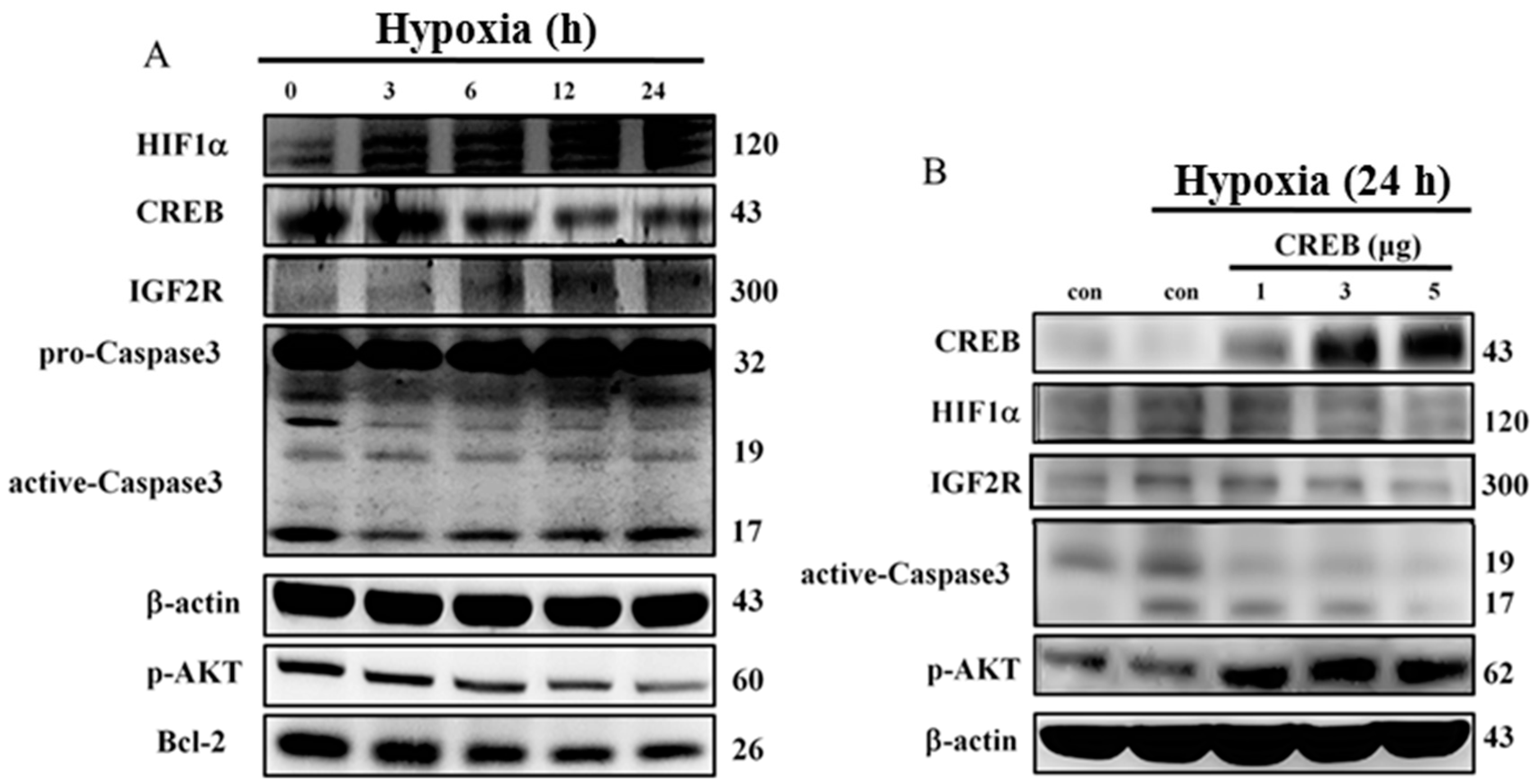
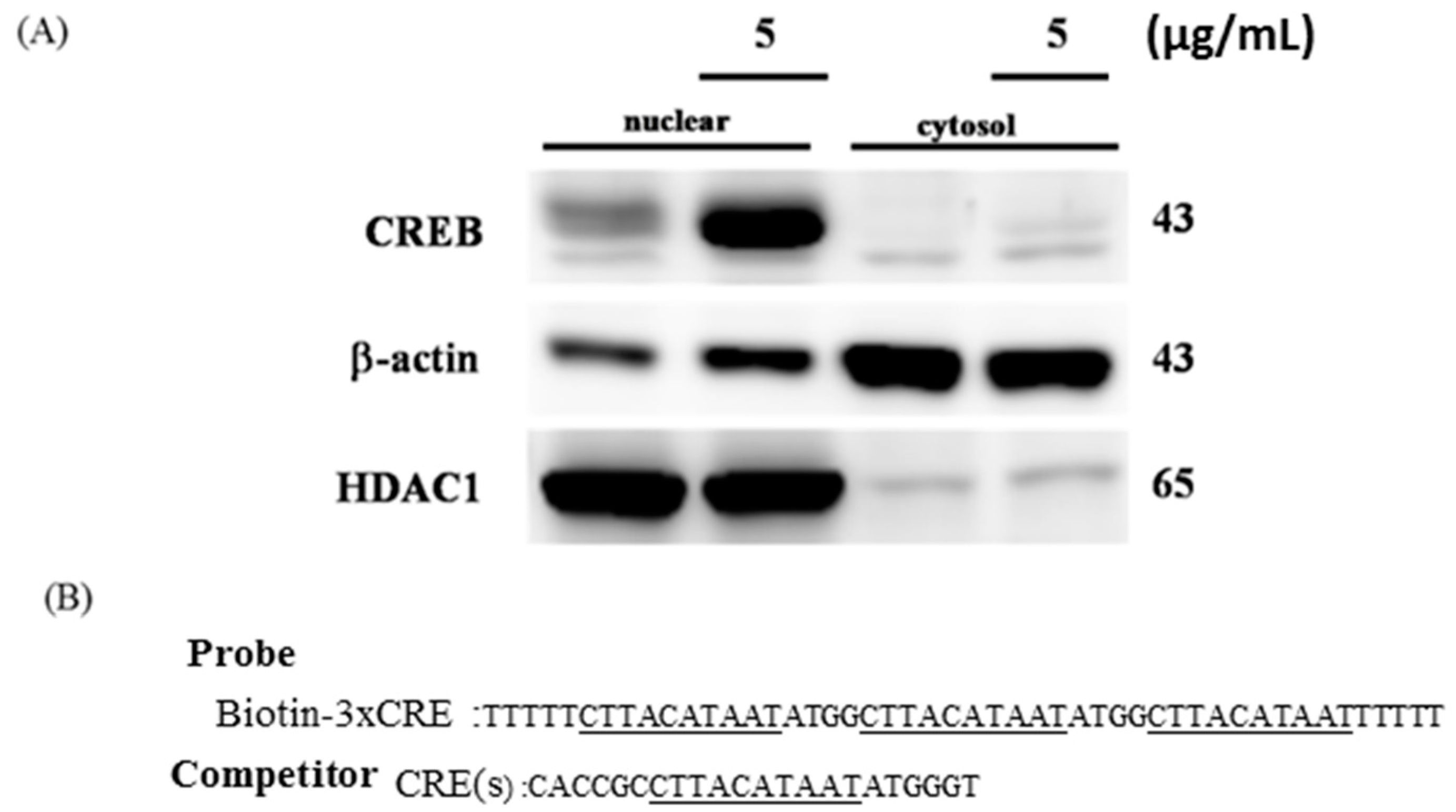
2.3. CREB Translocated to the Nuclei of H9c2 Cells to Directly Target the Promoter Region of the IGF2R
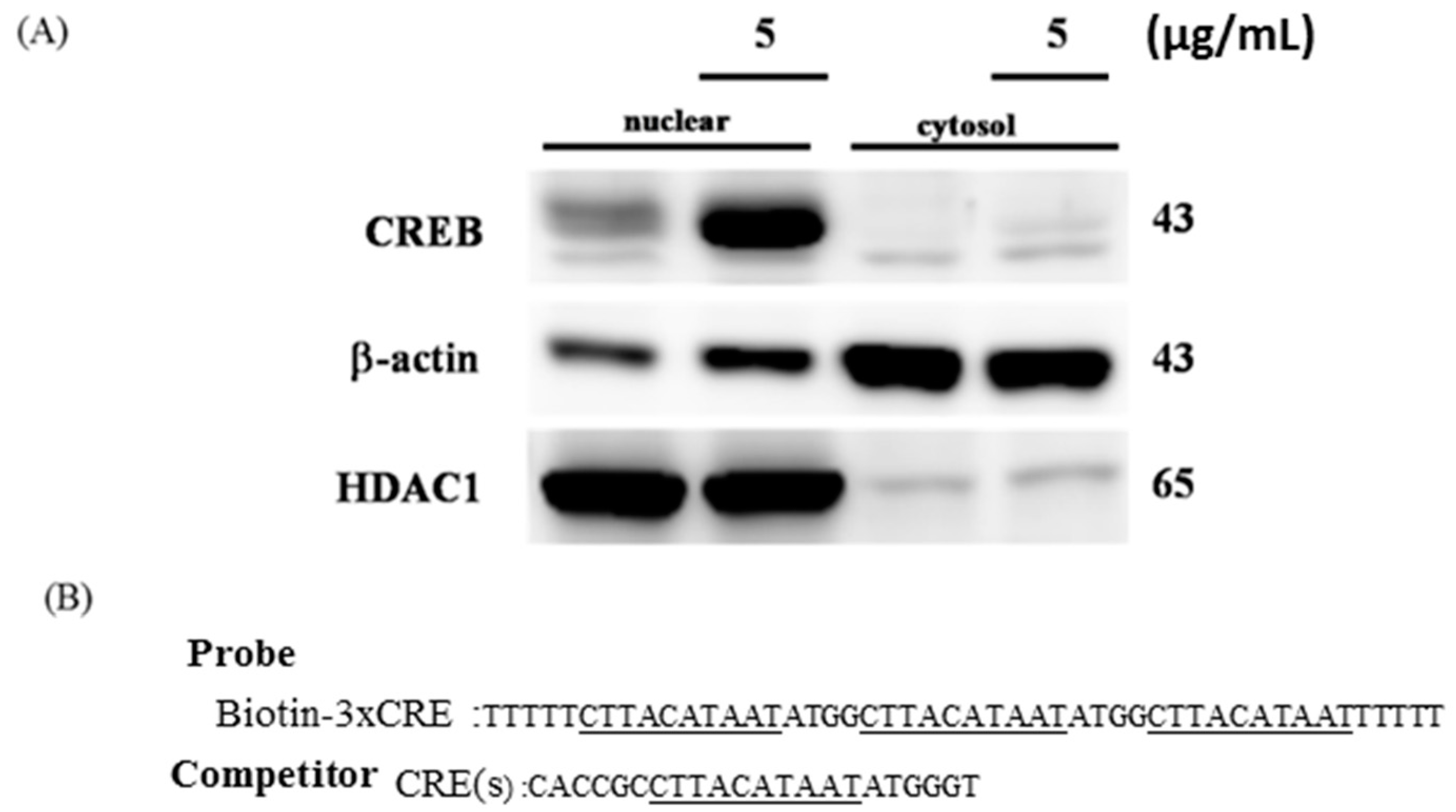
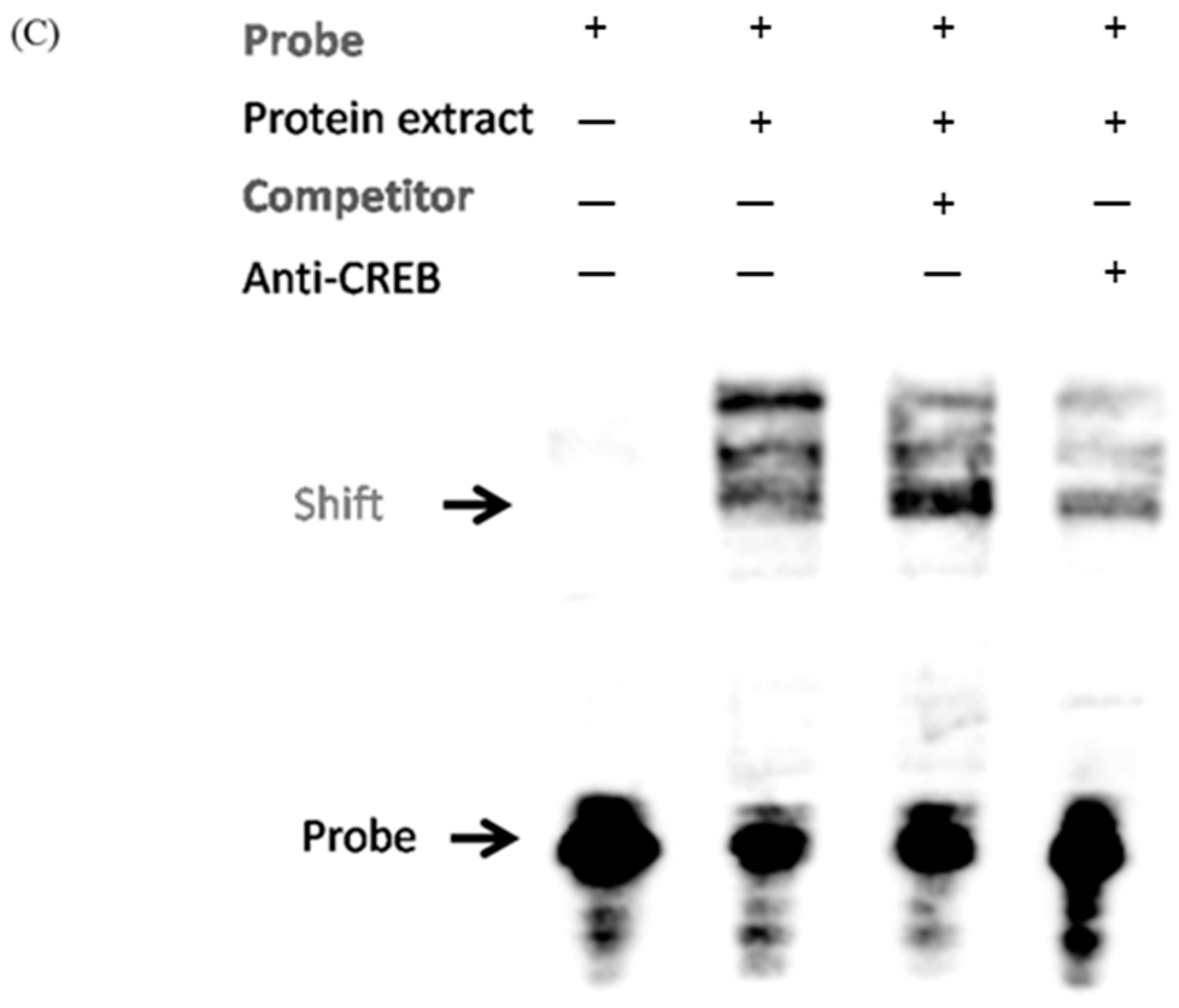
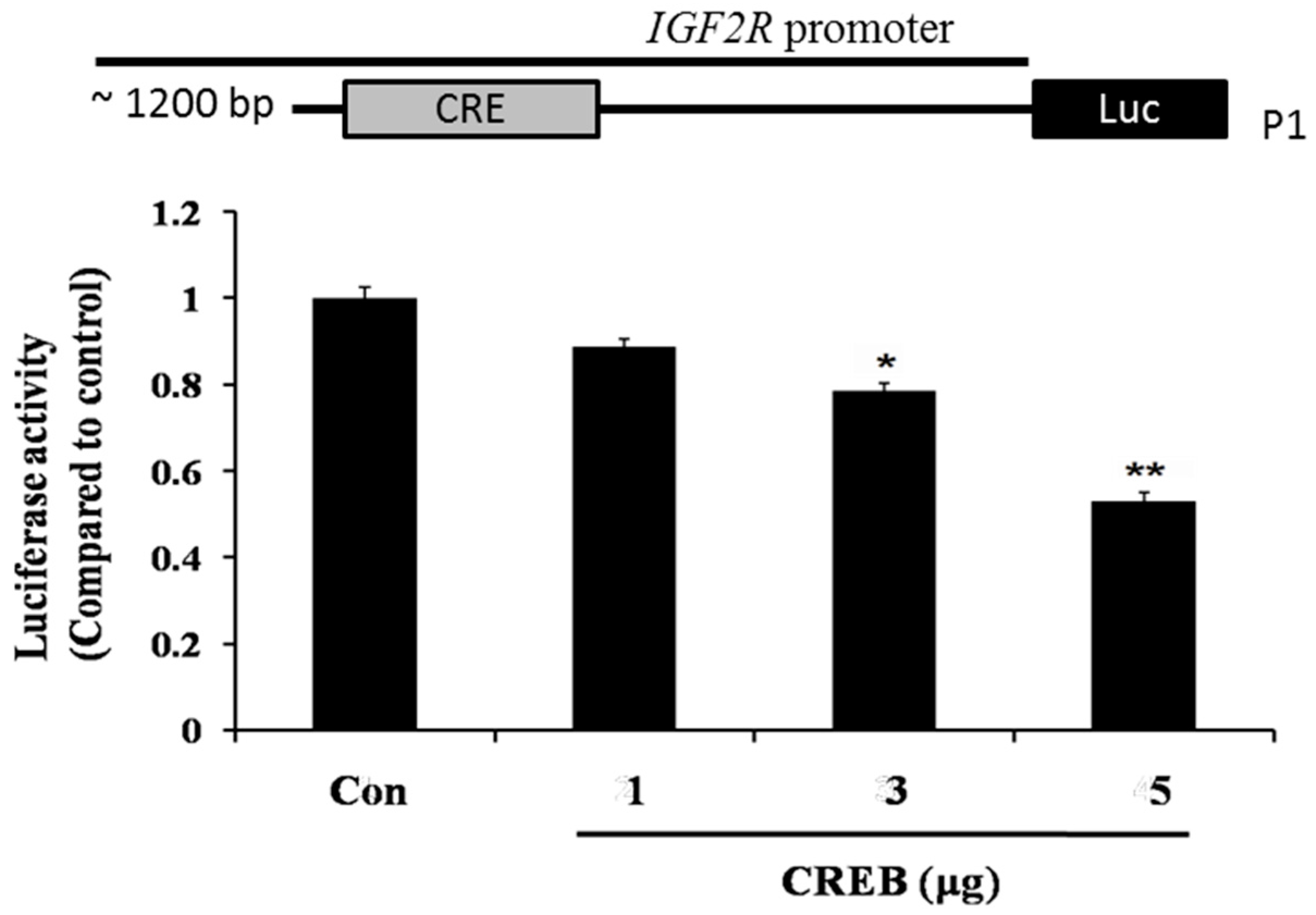
2.4. CREB Suppresses IGF2R Expression by Negatively Regulating the IGF2R Promoter Activity
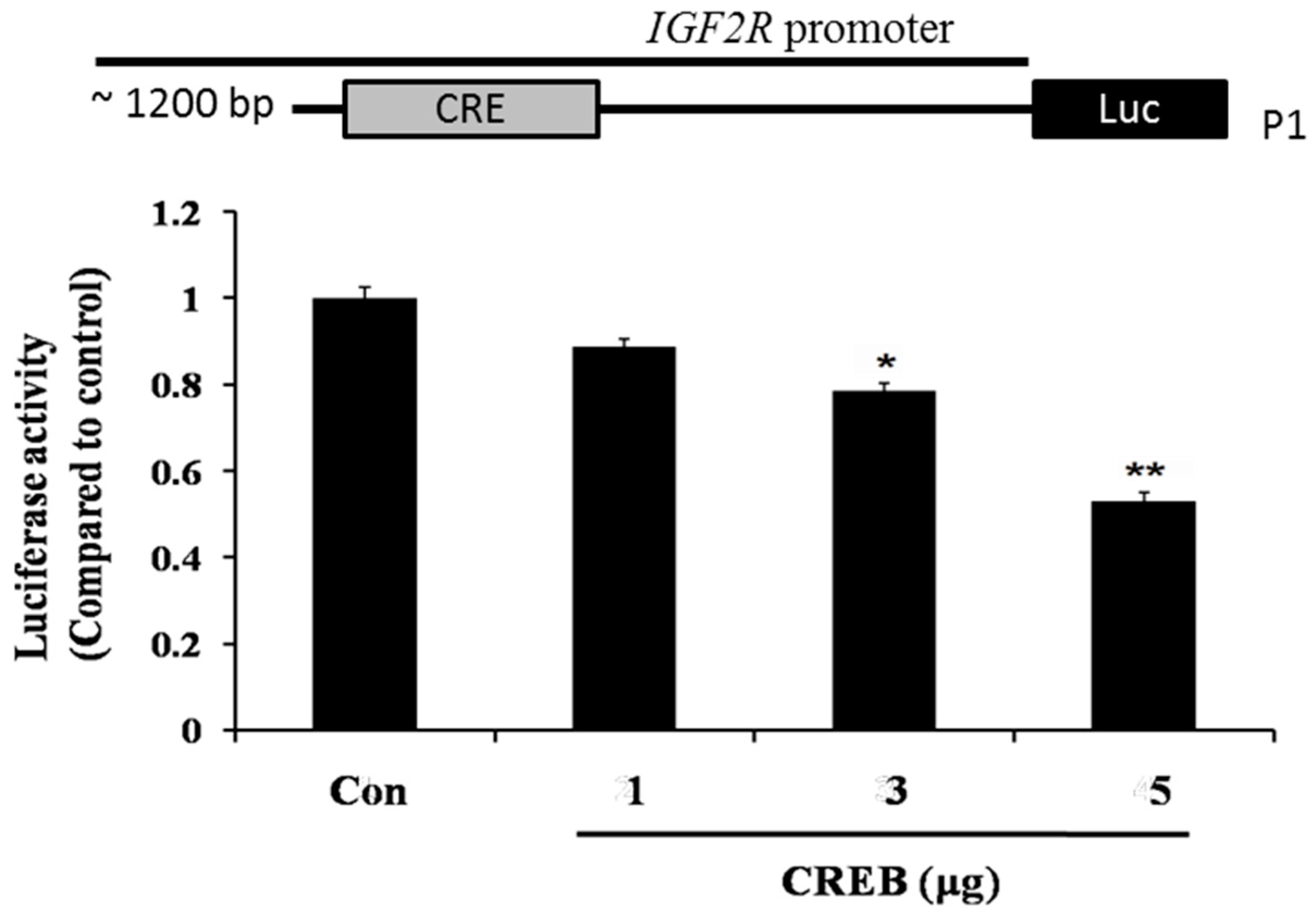
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Transfection
4.2. Western Blot
4.3. Nuclear Extracts and the Electrophoretic Mobility Shift Assay (EMSA)
4.4. Luciferase Assay
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hoeffler, J.P.; Meyer, T.E.; Yun, Y.; Jameson, J.L.; Habener, J.F. Cyclic AMP-responsive DNA-binding protein: Structure based on a cloned placental cDNA. Science 1988, 242, 1430–1433. [Google Scholar] [CrossRef] [PubMed]
- Schindler, M.; Fischer, S.; Thieme, R.; Fischer, B.; Santos, A.N. cAMP-responsive element binding protein: A vital link in embryonic hormonal adaptation. Endocrinology 2013, 154, 2208–2221. [Google Scholar] [CrossRef] [PubMed]
- Hansen, R.T., 3rd; Zhang, H.T. Senescent-induced dysregulation of cAMP/CREB signaling and correlations with cognitive decline. Brain Res. 2013, 1516, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.; Vanhoutte, P.; Pages, C.; Caboche, J.; Laroche, S. The MAPK/ERK cascade targets both Elk-1 and cAMP response element-binding protein to control long-term potentiation-dependent gene expression in the dentate gyrus in vivo. J. Neurosci. 2000, 20, 4563–4572. [Google Scholar] [PubMed]
- Lamph, W.W.; Dwarki, V.J.; Ofir, R.; Montminy, M.; Verma, I.M. Negative and positive regulation by transcription factor cAMP response element-binding protein is modulated by phosphorylation. Proc. Natl. Acad. Sci. USA 1990, 87, 4320–4324. [Google Scholar] [CrossRef] [PubMed]
- Ofir, R.; Dwarki, V.J.; Rashid, D.; Verma, I.M. CREB represses transcription of fos promoter: Role of phosphorylation. Gene Expr. 1991, 1, 55–60. [Google Scholar] [PubMed]
- Mayr, B.; Montminy, M. Transcriptional regulation by the phosphorylation-dependent factor CREB. Nat. Rev. Mol. Cell Biol. 2001, 2, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Meyer, T.E.; Habener, J.F. Cyclic adenosine 3′,5′-monophosphate response element binding protein (CREB) and related transcription-activating deoxyribonucleic acid-binding proteins. Endocr. Rev. 1993, 14, 269–290. [Google Scholar] [PubMed]
- Lee, K.A.; Masson, N. Transcriptional regulation by CREB and its relatives. Biochim. Biophys. Acta 1993, 1174, 221–233. [Google Scholar] [CrossRef]
- Lonze, B.E.; Ginty, D.D. Function and regulation of CREB family transcription factors in the nervous system. Neuron 2002, 35, 605–623. [Google Scholar] [CrossRef]
- Liu, W.; Chin-Chance, C.; Lee, E.J.; Lowe, W.L., Jr. Activation of phosphatidylinositol 3-kinase contributes to insulin-like growth factor I-mediated inhibition of pancreatic β-cell death. Endocrinology 2002, 143, 3802–3812. [Google Scholar] [CrossRef] [PubMed]
- Herzig, S.; Hedrick, S.; Morantte, I.; Koo, S.H.; Galimi, F.; Montminy, M. CREB controls hepatic lipid metabolism through nuclear hormone receptor PPAR-γ. Nature 2003, 426, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Herzig, S.; Long, F.; Jhala, U.S.; Hedrick, S.; Quinn, R.; Bauer, A.; Rudolph, D.; Schutz, G.; Yoon, C.; Puigserver, P.; et al. CREB regulates hepatic gluconeogenesis through the coactivator PGC-1. Nature 2001, 413, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Fentzke, R.C.; Korcarz, C.E.; Lang, R.M.; Lin, H.; Leiden, J.M. Dilated cardiomyopathy in transgenic mice expressing a dominant-negative CREB transcription factor in the heart. J. Clin. Investig. 1998, 101, 2415–2426. [Google Scholar] [CrossRef] [PubMed]
- Mehrhof, F.B.; Muller, F.U.; Bergmann, M.W.; Li, P.; Wang, Y.; Schmitz, W.; Dietz, R.; von Harsdorf, R. In cardiomyocyte hypoxia, insulin-like growth factor-I-induced antiapoptotic signaling requires phosphatidylinositol-3-OH-kinase-dependent and mitogen-activated protein kinase-dependent activation of the transcription factor cAMP response element-binding protein. Circulation 2001, 104, 2088–2094. [Google Scholar] [PubMed]
- Kang, P.M.; Izumo, S. Apoptosis and heart failure: A critical review of the literature. Circ. Res. 2000, 86, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-Y.; Lee, S.-D. Possible pathophysiology of heart failure in obesity: Cardiac apoptosis. BioMedicine 2012, 2, 36–40. [Google Scholar] [CrossRef]
- Chu, C.H.; Lo, J.F.; Hu, W.S.; Lu, R.B.; Chang, M.H.; Tsai, F.J.; Tsai, C.H.; Weng, Y.S.; Tzang, B.S.; Huang, C.Y. Histone acetylation is essential for ANG-II-induced IGF-IIR gene expression in H9c2 cardiomyoblast cells and pathologically hypertensive rat heart. J. Cell. Physiol. 2012, 227, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.H.; Tzang, B.S.; Chen, L.M.; Kuo, C.H.; Cheng, Y.C.; Chen, L.Y.; Tsai, F.J.; Tsai, C.H.; Kuo, W.W.; Huang, C.Y. IGF-II/mannose-6-phosphate receptor signaling induced cell hypertrophy and atrial natriuretic peptide/BNP expression via Gαq interaction and protein kinase C-α/CaMKII activation in H9c2 cardiomyoblast cells. J. Endocrinol. 2008, 197, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Motyka, B.; Korbutt, G.; Pinkoski, M.J.; Heibein, J.A.; Caputo, A.; Hobman, M.; Barry, M.; Shostak, I.; Sawchuk, T.; Holmes, C.F.; et al. Mannose 6-phosphate/insulin-like growth factor II receptor is a death receptor for granzyme B during cytotoxic T cell-induced apoptosis. Cell 2000, 103, 491–500. [Google Scholar] [CrossRef]
- Chu, C.H.; Tzang, B.S.; Chen, L.M.; Liu, C.J.; Tsai, F.J.; Tsai, C.H.; Lin, J.A.; Kuo, W.W.; Bau, D.T.; Yao, C.H.; et al. Activation of insulin-like growth factor II receptor induces mitochondrial-dependent apoptosis through Gαq and downstream calcineurin signaling in myocardial cells. Endocrinology 2009, 150, 2723–2731. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.H.; Huang, C.Y.; Lu, M.C.; Lin, J.A.; Tsai, F.J.; Tsai, C.H.; Chu, C.Y.; Kuo, W.H.; Chen, L.M.; Chen, L.Y. Enhancement of AG1024-induced H9c2 cardiomyoblast cell apoptosis via the interaction of IGF2R with Gα proteins and its downstream PKA and PLC-β modulators by IGF-II. Chin. J. Physiol. 2009, 52, 31–37. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, S.M.; Leonard, M.O.; Kieran, N.; Comerford, K.M.; Cummins, E.; Pouliot, M.; Lee, S.B.; Taylor, C.T. Hypoxia induces epithelial amphiregulin gene expression in a CREB-dependent manner. Am. J. Physiol. Cell Physiol. 2006, 290, C592–C600. [Google Scholar] [CrossRef] [PubMed]
- Crawford, R.M.; Jovanovic, S.; Budas, G.R.; Davies, A.M.; Lad, H.; Wenger, R.H.; Robertson, K.A.; Roy, D.J.; Ranki, H.J.; Jovanovic, A. Chronic mild hypoxia protects heart-derived H9c2 cells against acute hypoxia/reoxygenation by regulating expression of the SUR2A subunit of the ATP-sensitive K+ channel. J. Biol. Chem. 2003, 278, 31444–31455. [Google Scholar] [CrossRef] [PubMed]
- Abdul, K.S.M.; Joyanovic, S.; Sukhodub, A.; Du, Q.Y.; Jovanovic, A. Upregulation of cardioprotective SUR2A by sub-hypoxic drop in oxygen. BBA Mol. Cell Res. 2014, 1843, 2424–2431. [Google Scholar] [Green Version]
- Shameem, K.; Abdul, M.; Jovanovic, S.; Du, Q.Y.; Sukhodub, A.; Jovanovic, A. A link between ATP and SUR2A: A novel mechanism explaining cardioprotection at high altitude. Int. J. Cardiol. 2015, 189, 73–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul, K.S.M.; Jovanovic, S.; Du, Q.; Sukhodub, A.; Jovanovic, A. Mild hypoxia in vivo regulates cardioprotective SUR2A: A role for Akt and LDH. BBA Mol. Basis Dis. 2015, 1852, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.D.; Kuo, W.W.; Wu, C.H.; Lin, Y.M.; Lin, J.A.; Lu, M.C.; Yang, A.L.; Liu, J.Y.; Wang, S.G.P.; Liu, C.J.; et al. Effects of short- and long-term hypobaric hypoxia on Bcl2 family in rat heart. Int. J. Cardiol. 2006, 108, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.L.; Lin, J.W.; Hsieh, D.J.; Yeh, Y.L.; Shen, C.Y.; Day, C.H.; Ho, T.J.; Viswanadha, V.P.; Kuo, W.W.; Huang, C.Y. Long-term hypoxia exposure enhanced IGFBP-3 protein synthesis and secretion resulting in cell apoptosis in H9c2 myocardial cells. Growth Factors 2015, 33, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.M.; Kuo, W.W.; Yang, J.J.; Wang, S.G.P.; Yeh, Y.L.; Tsai, F.J.; Ho, Y.J.; Chang, M.H.; Huang, C.Y.; Lee, S.D. Eccentric cardiac hypertrophy was induced by long-term intermittent hypoxia in rats. Exp. Physiol. 2007, 92, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Kotla, S.; Singh, N.K.; Heckle, M.R.; Tigyi, G.J.; Rao, G.N. The transcription factor CREB enhances interleukin-17A production and inflammation in a mouse model of atherosclerosis. Sci. Signal. 2013, 6, ra83. [Google Scholar] [CrossRef] [PubMed]
- Schauer, I.E.; Knaub, L.A.; Lloyd, M.; Watson, P.A.; Gliwa, C.; Lewis, K.E.; Chait, A.; Klemm, D.J.; Gunter, J.M.; Bouchard, R.; et al. CREB downregulation in vascular disease: A common response to cardiovascular risk. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Ozgen, N.; Guo, J.; Gertsberg, Z.; Danilo, P., Jr.; Rosen, M.R.; Steinberg, S.F. Reactive oxygen species decrease cAMP response element binding protein expression in cardiomyocytes via a protein kinase D1-dependent mechanism that does not require Ser133 phosphorylation. Mol. Pharmacol. 2009, 76, 896–902. [Google Scholar] [CrossRef] [PubMed]
- Fiol, C.J.; Williams, J.S.; Chou, C.H.; Wang, Q.M.; Roach, P.J.; Andrisani, O.M. A secondary phosphorylation of CREB341 at Ser129 is required for the cAMP-mediated control of gene expression. A role for glycogen synthase kinase-3 in the control of gene expression. J. Biol. Chem. 1994, 269, 32187–32193. [Google Scholar] [PubMed]
- Gonzalez, G.A.; Montminy, M.R. Cyclic AMP stimulates somatostatin gene transcription by phosphorylation of CREB at serine 133. Cell 1989, 59, 675–680. [Google Scholar] [CrossRef]
- Balogh, A.; Nemeth, M.; Koloszar, I.; Marko, L.; Przybyl, L.; Jinno, K.; Szigeti, C.; Heffer, M.; Gebhardt, M.; Szeberenyi, J.; et al. Overexpression of CREB protein protects from tunicamycin-induced apoptosis in various rat cell types. Apoptosis Int. J. Program. Cell Death 2014, 19, 1080–1098. [Google Scholar] [CrossRef] [PubMed]
- Bonni, A.; Brunet, A.; West, A.E.; Datta, S.R.; Takasu, M.A.; Greenberg, M.E. Cell survival promoted by the Ras-MAPK signaling pathway by transcription-dependent and -independent mechanisms. Science 1999, 286, 1358–1362. [Google Scholar] [CrossRef] [PubMed]
- Walton, M.; Woodgate, A.M.; Muravlev, A.; Xu, R.; During, M.J.; Dragunow, M. CREB phosphorylation promotes nerve cell survival. J. Neurochem. 1999, 73, 1836–1842. [Google Scholar] [PubMed]
- Ghosh, P.; Dahms, N.M.; Kornfeld, S. Mannose 6-phosphate receptors: New twists in the tale. Nat. Rev. Mol. Cell Biol. 2003, 4, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Bohnsack, R.N.; Patel, M.; Olson, L.J.; Twining, S.S.; Dahms, N.M. Residues essential for plasminogen binding by the cation-independent mannose 6-phosphate receptor. Biochemistry 2010, 49, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Kuo, W.W.; Liu, C.J.; Chen, L.M.; Wu, C.H.; Chu, C.H.; Liu, J.Y.; Lu, M.C.; Lin, J.A.; Lee, S.D.; Huang, C.Y. Cardiomyoblast apoptosis induced by insulin-like growth factor (IGF)-I resistance is IGF-II dependent and synergistically enhanced by angiotensin II. Apoptosis Int. J. Program. Cell Death 2006, 11, 1075–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.D.; Chu, C.H.; Huang, E.J.; Lu, M.C.; Liu, J.Y.; Liu, C.J.; Hsu, H.H.; Lin, J.A.; Kuo, W.W.; Huang, C.Y. Roles of insulin-like growth factor II in cardiomyoblast apoptosis and in hypertensive rat heart with abdominal aorta ligation. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E306–E314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montminy, M. Transcriptional regulation by cyclic AMP. Annu. Rev. Biochem. 1997, 66, 807–822. [Google Scholar] [CrossRef] [PubMed]
- MacGillavry, H.D.; Stam, F.J.; Sassen, M.M.; Kegel, L.; Hendriks, W.T.; Verhaagen, J.; Smit, A.B.; van Kesteren, R.E. NFIL3 and cAMP response element-binding protein form a transcriptional feedforward loop that controls neuronal regeneration-associated gene expression. J. Neurosci. 2009, 29, 15542–15550. [Google Scholar] [CrossRef] [PubMed]
- Matus, M.; Lewin, G.; Stumpel, F.; Buchwalow, I.B.; Schneider, M.D.; Schutz, G.; Schmitz, W.; Muller, F.U. Cardiomyocyte-specific inactivation of transcription factor CREB in mice. FASEB J. 2007, 21, 1884–1892. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.H.; Kuo, C.H.; Kuo, W.W.; Ho, T.J.; Pai, P.; Chen, W.K.; Pan, L.F.; Wang, C.C.; Padma, V.V.; Huang, C.Y. NFIL3 suppresses hypoxia-induced apoptotic cell death by targeting the insulin-like growth factor 2 receptor. J. Cell. Biochem. 2015, 116, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liu, J.; Jo, M.; Curry, T.E., Jr. A role for nuclear factor interleukin-3 (NFIL3), a critical transcriptional repressor, in down-regulation of periovulatory gene expression. Mol. Endocrinol. 2011, 25, 445–459. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.-K.; Kuo, W.-W.; Hsieh, D.J.-Y.; Chang, H.-N.; Pai, P.-Y.; Lin, K.-H.; Pan, L.-F.; Ho, T.-J.; Viswanadha, V.P.; Huang, C.-Y. CREB Negatively Regulates IGF2R Gene Expression and Downstream Pathways to Inhibit Hypoxia-Induced H9c2 Cardiomyoblast Cell Death. Int. J. Mol. Sci. 2015, 16, 27921-27930. https://doi.org/10.3390/ijms161126067
Chen W-K, Kuo W-W, Hsieh DJ-Y, Chang H-N, Pai P-Y, Lin K-H, Pan L-F, Ho T-J, Viswanadha VP, Huang C-Y. CREB Negatively Regulates IGF2R Gene Expression and Downstream Pathways to Inhibit Hypoxia-Induced H9c2 Cardiomyoblast Cell Death. International Journal of Molecular Sciences. 2015; 16(11):27921-27930. https://doi.org/10.3390/ijms161126067
Chicago/Turabian StyleChen, Wei-Kung, Wei-Wen Kuo, Dennis Jine-Yuan Hsieh, Hsin-Nung Chang, Pei-Ying Pai, Kuan-Ho Lin, Lung-Fa Pan, Tsung-Jung Ho, Vijaya Padma Viswanadha, and Chih-Yang Huang. 2015. "CREB Negatively Regulates IGF2R Gene Expression and Downstream Pathways to Inhibit Hypoxia-Induced H9c2 Cardiomyoblast Cell Death" International Journal of Molecular Sciences 16, no. 11: 27921-27930. https://doi.org/10.3390/ijms161126067