
Identification of Retinopathy of Prematurity Related miRNAs in Hyperoxia-Induced Neonatal Rats by Deep Sequencing

Abstract
:1. Introduction
2. Results and Discussion
2.1. Analysis of Sequenced Small RNAs
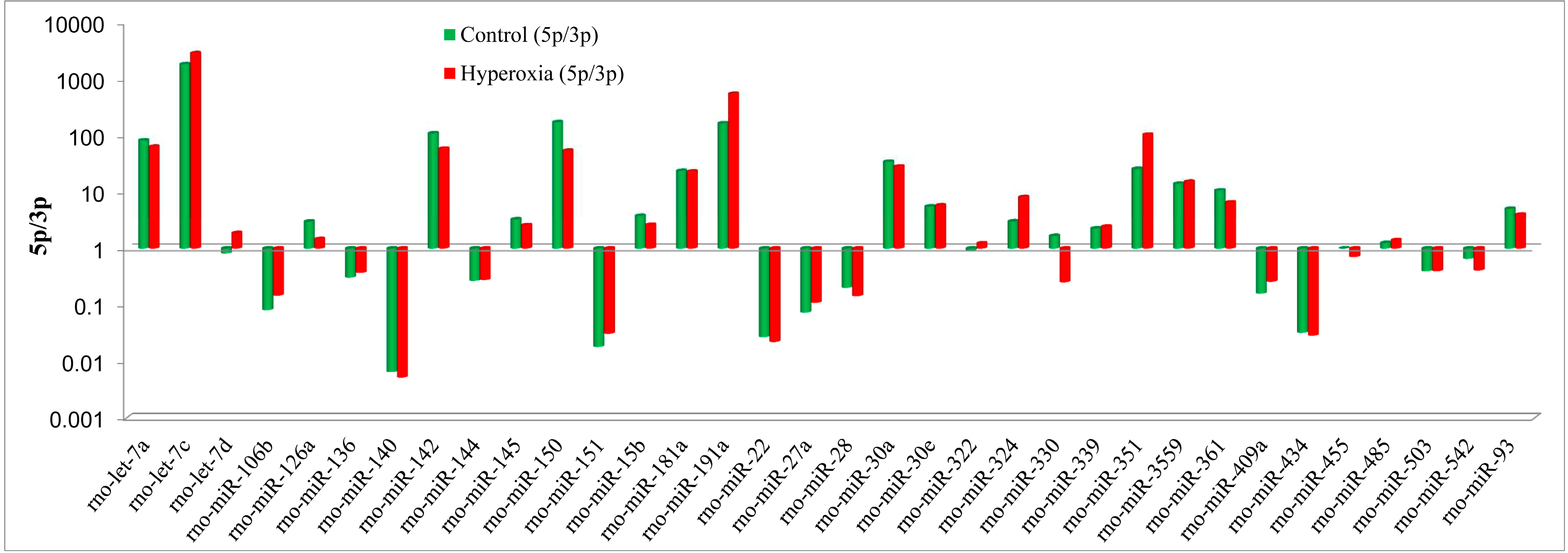
2.2. Differential Expression Analysis of the miRNAs from the Hyperoxia Rats and Controls

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Down-Regulated miRNAs | Log 2 (Fold-Change) |
|---|---|
| rno-miR-330-5p | −3.84522 |
| rno-miR-223-3p | −3.35767 |
| rno-miR-191a-3p | −3.33064 |
| rno-miR-377-3p | −3.10825 |
| rno-miR-128-3p | −2.535 |
| rno-miR-181c-3p | −2.52329 |
| rno-miR-324-3p | −2.52329 |
| rno-miR-340-3p | −2.52329 |
| rno-miR-378a-5p | −2.52329 |
| rno-miR-326-3p | −2.26025 |
| rno-miR-425-3p | −1.97075 |
| rno-miR-191a-5p | −1.58496 |
| rno-miR-122-5p | −1.52329 |
| rno-miR-1306-5p | −1.52329 |
| rno-miR-350 | −1.52329 |
| rno-miR-383-5p | −1.52329 |
| rno-miR-483-3p | −1.52329 |
| rno-miR-667-3p | −1.52329 |
| rno-miR-708-5p | −1.52329 |
| rno-miR-328a-3p | −1.50559 |
| rno-miR-674-3p | −1.39776 |
| rno-miR-423-3p | −1.3147 |
| rno-miR-132-3p | −1.3009 |
| rno-miR-342-3p | −1.28949 |
| rno-miR-451-5p | −1.24595 |
| rno-miR-345-3p | −1.23378 |
| rno-miR-92a-3p | −1.17894 |
| rno-miR-181d-5p | −1.14478 |
| rno-miR-133a-3p | −1.1243 |
| rno-miR-29a-3p | −1.10989 |
| rno-miR-1843-5p | −1.10825 |
| rno-miR-18a-5p | −1.10825 |
| rno-miR-25-5p | −1.10825 |
| rno-miR-324-5p | −1.10825 |
| rno-miR-330-3p | −1.10825 |
| rno-miR-3473 | −1.10825 |
| rno-miR-493-3p | −1.10825 |
| rno-miR-872-3p | −1.10825 |
| rno-miR-652-3p | −1.09265 |
| rno-miR-505-3p | −1.0538 |
| rno-miR-19b-3p | −1.04983 |
| rno-miR-351-3p | −1.03786 |
| rno-miR-126a-5p | −1.03563 |
| rno-miR-17-5p | −1.02817 |
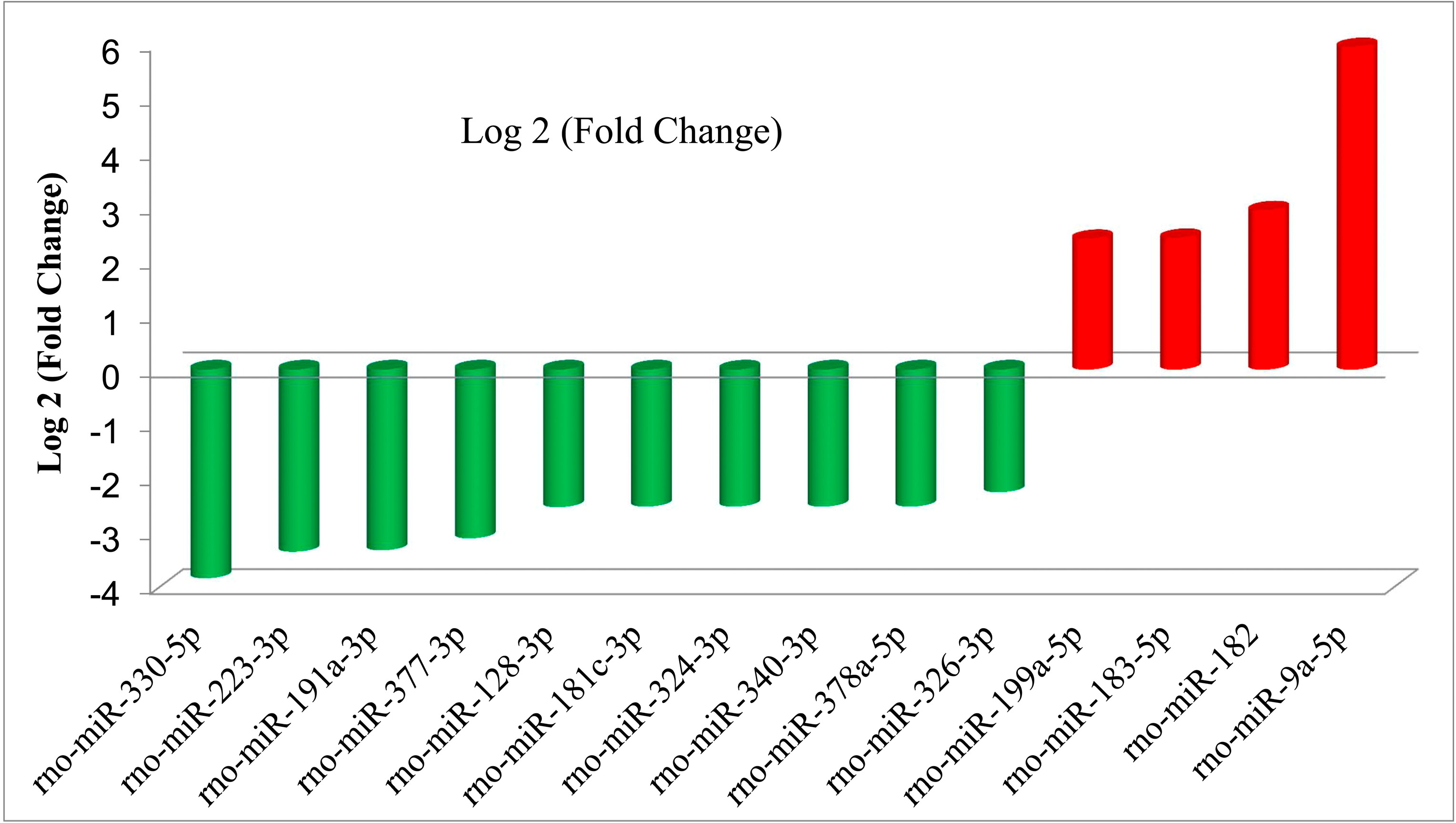
| Up-Regulated miRNAs | Log 2 (Fold-Change) |
|---|---|
| rno-miR-351-5p | 1.036533 |
| rno-miR-200c-3p | 1.049601 |
| rno-miR-219a-2-3p | 1.061673 |
| rno-miR-92b-3p | 1.061673 |
| rno-miR-200a-3p | 1.077104 |
| rno-miR-127-3p | 1.107432 |
| rno-miR-181a-2-3p | 1.139676 |
| rno-let-7e-5p | 1.177151 |
| rno-miR-433-3p | 1.177151 |
| rno-miR-337-5p | 1.213677 |
| rno-miR-494-3p | 1.35118 |
| rno-miR-200b-3p | 1.424244 |
| rno-miR-204-5p | 1.512335 |
| rno-miR-195-5p | 1.535605 |
| rno-miR-125b-5p | 1.552595 |
| rno-miR-3068-3p | 1.564174 |
| rno-miR-375-3p | 1.747109 |
| rno-miR-429 | 1.837458 |
| rno-miR-199a-5p | 2.406502 |
| rno-miR-183-5p | 2.424824 |
| rno-miR-182 | 2.933923 |
| rno-miR-9a-5p | 5.936143 |
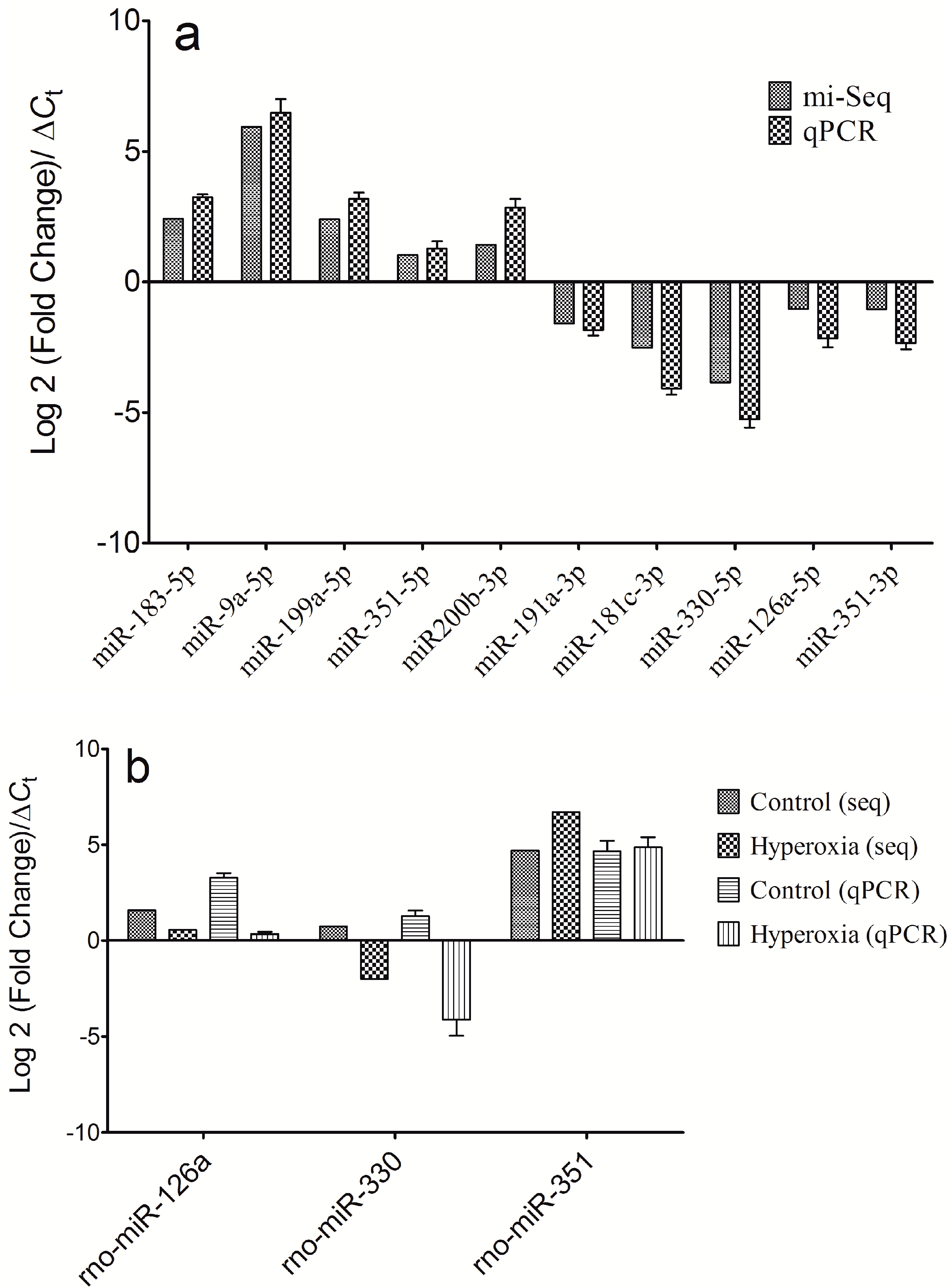
2.3. qPCR Validation of the Differential Expressions of the miRNAs


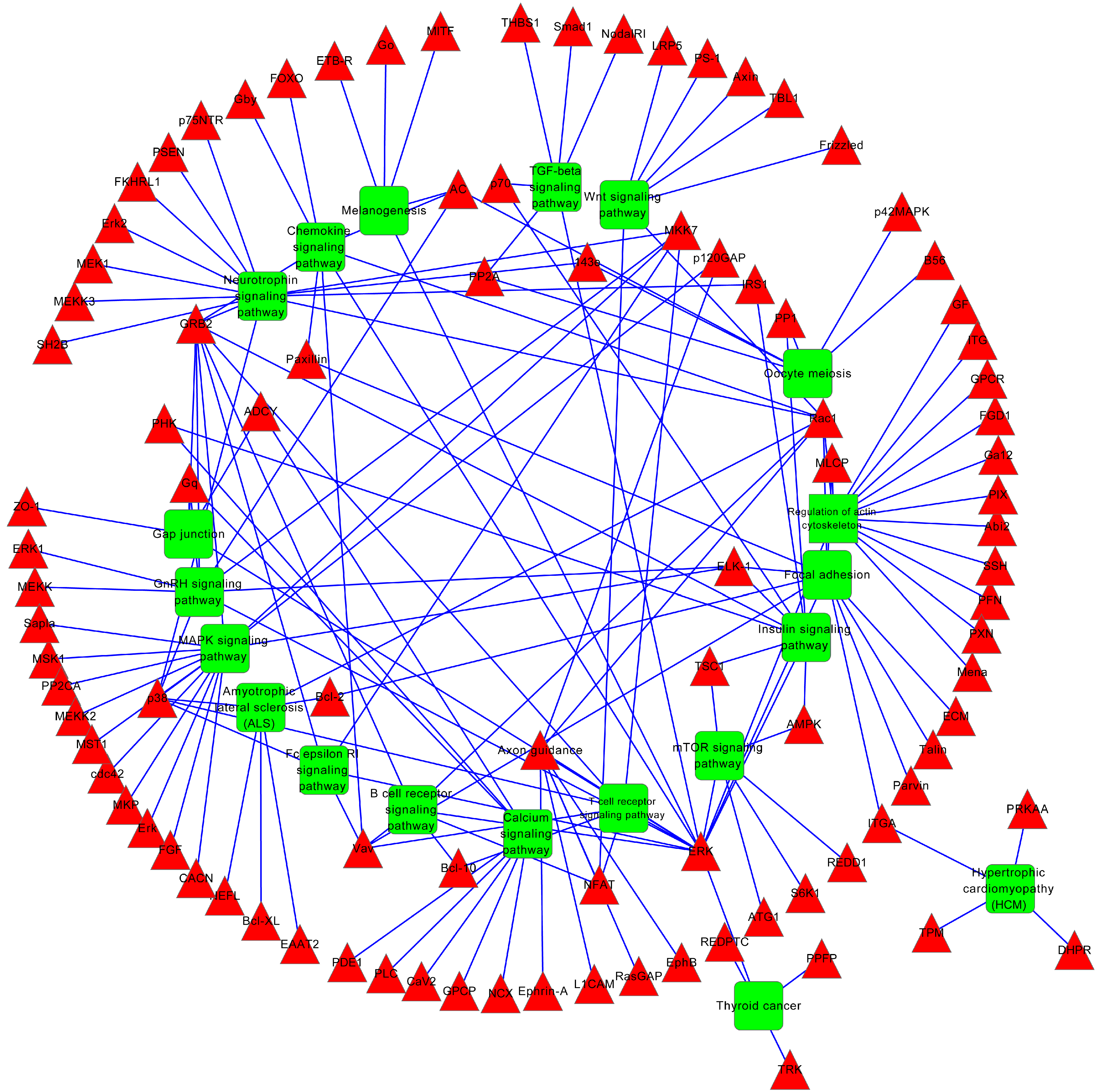
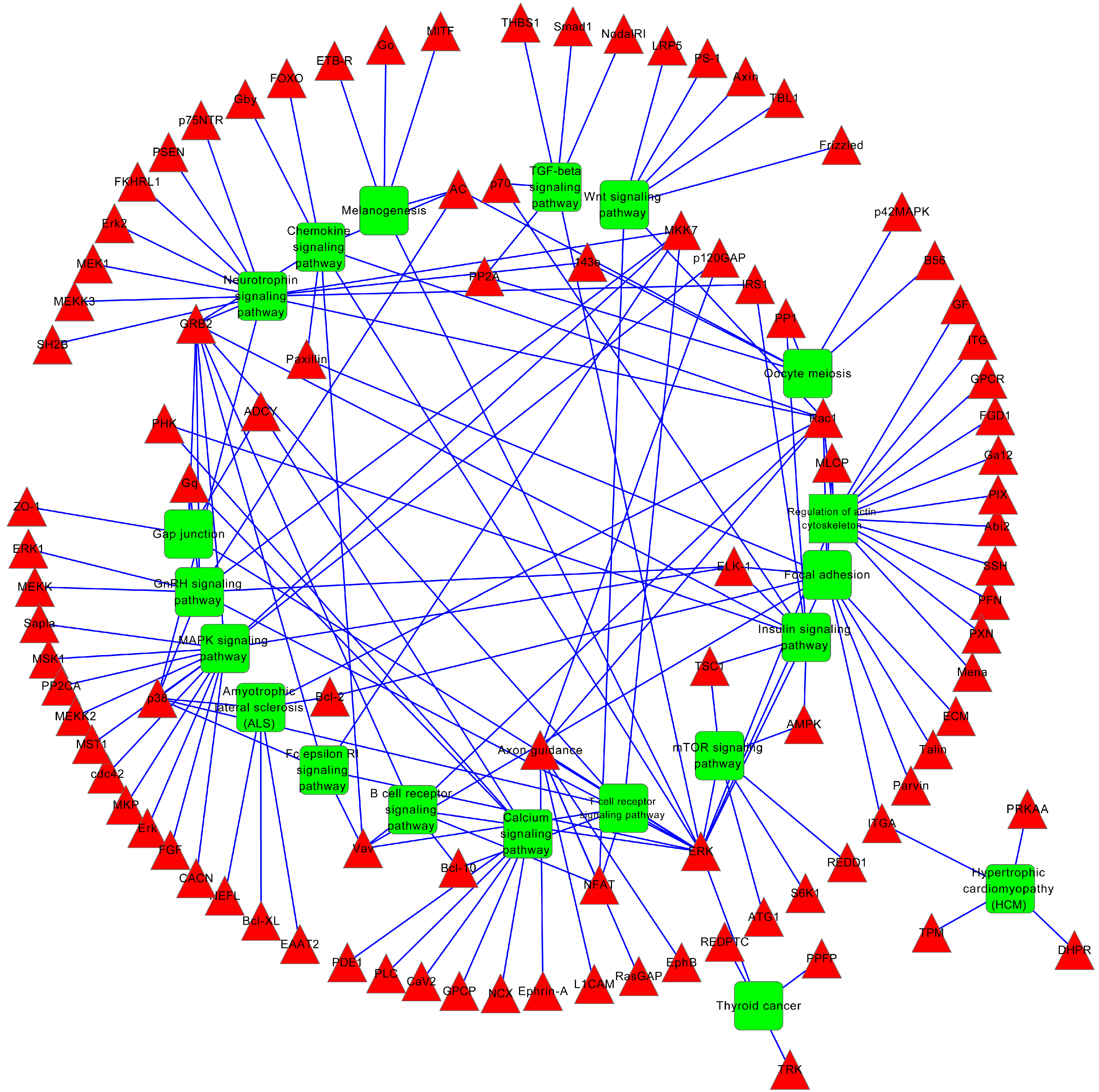
2.4. miRNA Target Prediction and Functional Annotation
| GO Term | Gene Count | p-Value | FDR (Benjamini Hochberg) * |
|---|---|---|---|
| Embryonic development ending in birth or egg hatching | 37 | 7.10 × 10−10 | 8.80 × 10−7 |
| Chordate embryonic development | 37 | 4.90 × 10−10 | 1.20 × 10−6 |
| Intracellular signaling cascade | 60 | 1.40 × 10−8 | 1.20 × 10−5 |
| Cell migration | 27 | 5.40 × 10−8 | 3.40 × 10−5 |
| Cell motion | 34 | 2.60 × 10−7 | 1.30 × 10−4 |
| In utero embryonic development | 24 | 4.40 × 10−7 | 1.80 × 10−4 |
| Embryonic morphogenesis | 28 | 1.20 × 10−6 | 4.10 × 10−4 |
| Cell morphogenesis | 28 | 3.30 × 10−6 | 1.00 × 10−3 |
| Cell motility | 27 | 4.80 × 10−6 | 1.30 × 10−3 |
| Localization of cell | 27 | 4.80 × 10−6 | 1.30 × 10−3 |
| Positive regulation of gene expression | 37 | 6.10 × 10−6 | 1.50 × 10−3 |
| Embryonic organ development | 21 | 6.70 × 10−6 | 1.50 × 10−3 |
| Positive regulation of nucleobase, nucleoside, nucleotide and nucleic acid metabolic process | 39 | 1.10 × 10−5 | 1.90 × 10−3 |
| Cell projection organization | 28 | 9.90 × 10−6 | 2.00 × 10−3 |
| Forebrain development | 18 | 1.00 × 10−5 | 2.00 × 10−3 |
| Positive regulation of cellular biosynthetic process | 41 | 1.50 × 10−5 | 2.40 × 10−3 |
| Positive regulation of transcription | 35 | 2.20 × 10−5 | 2.90 × 10−3 |
| Appendage morphogenesis | 13 | 2.30 × 10−5 | 2.90 × 10−3 |
| Limb morphogenesis | 13 | 2.30 × 10−5 | 2.90 × 10−3 |
| Positive regulation of biosynthetic process | 41 | 2.20 × 10−5 | 3.00 × 10−3 |
| KEGG Term | Gene Count | p-Value | FDR (Benjamini Hochberg) * |
|---|---|---|---|
| Neurotrophin signaling pathway | 17 | 1.00 × 10−6 | 1.20 × 10−4 |
| Regulation of actin cytoskeleton | 16 | 1.60 × 10−3 | 6.10 × 10−2 |
| MAPK signaling pathway | 19 | 1.10 × 10−3 | 6.20 × 10−2 |
| Focal adhesion | 14 | 6.10 × 10−3 | 1.60 × 10−1 |
| GnRH signaling pathway | 9 | 7.50 × 10−3 | 1.60 × 10−1 |
| mTOR signaling pathway | 6 | 2.20 × 10−2 | 2.30 × 10−1 |
| Melanogenesis | 8 | 2.10 × 10−2 | 2.40 × 10−1 |
| Chemokine signaling pathway | 12 | 1.50 × 10−2 | 2.50 × 10−1 |
| Oocyte meiosis | 9 | 1.90 × 10−2 | 2.50 × 10−1 |
| Insulin signaling pathway | 10 | 1.90 × 10−2 | 2.70 × 10−1 |
| Amyotrophic lateral sclerosis (ALS) | 6 | 3.30 × 10−2 | 2.80 × 10−1 |
| Wnt signaling pathway | 10 | 3.20 × 10−2 | 2.90 × 10−1 |
| ErbB signaling pathway | 7 | 4.40 × 10−2 | 3.10 × 10−1 |
| TGF-beta signaling pathway | 7 | 4.60 × 10−2 | 3.10 × 10−1 |
| Maturity onset diabetes of the young | 4 | 4.30 × 10−2 | 3.30 × 10−1 |
| Vascular smooth muscle contraction | 8 | 5.60 × 10−2 | 3.30 × 10−1 |
| Dilated cardiomyopathy | 7 | 5.50 × 10−2 | 3.40 × 10−1 |
| Thyroid cancer | 4 | 6.20 × 10−2 | 3.40 × 10−1 |
| Natural killer cell mediated cytotoxicity | 7 | 8.30 × 10−2 | 3.80 × 10−1 |
| B cell receptor signaling pathway | 6 | 7.70 × 10−2 | 3.90 × 10−1 |
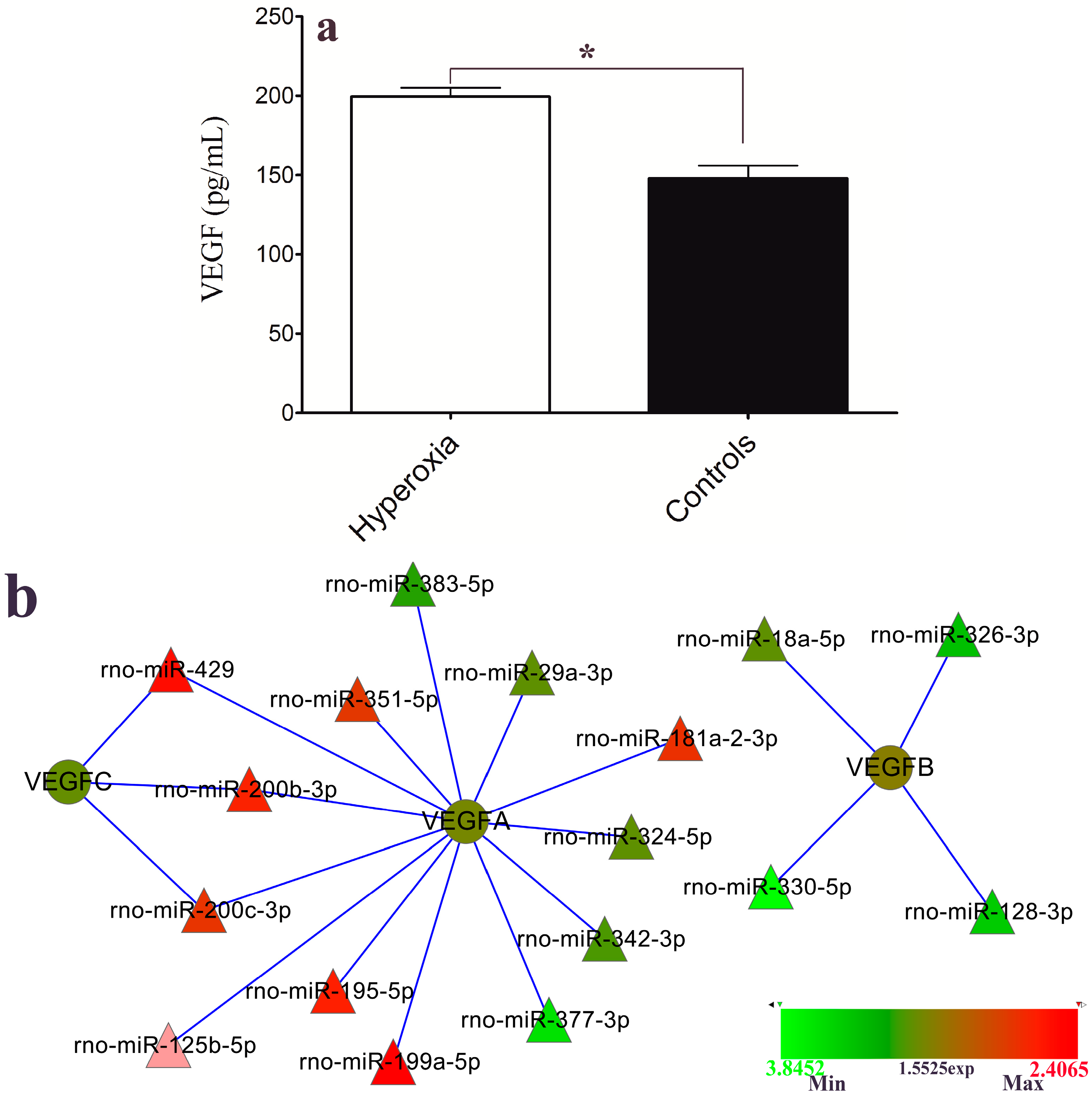
2.5. VEGF Level and Hyperoxia-Related miRNAs
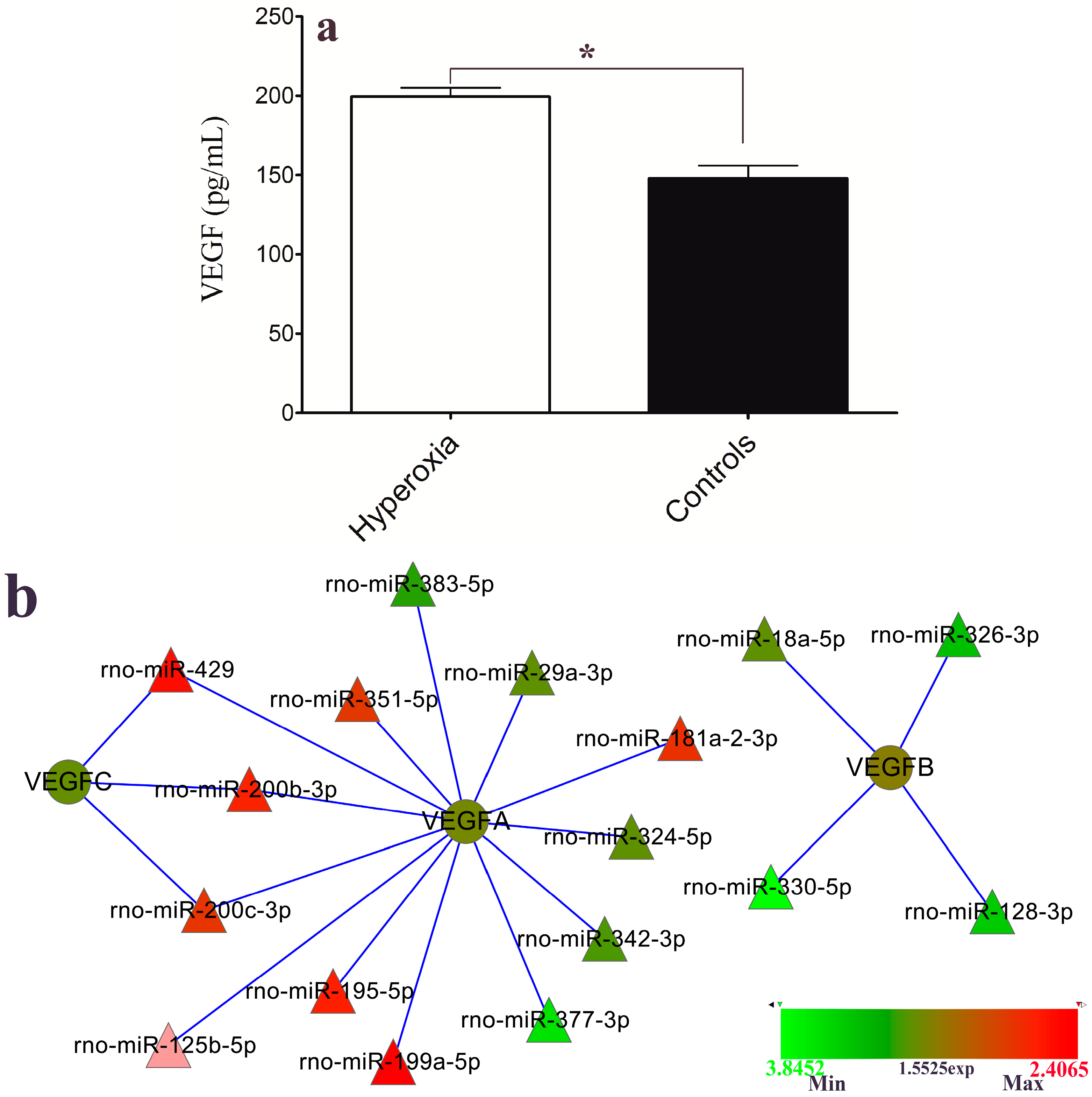
3. Experimental Section
3.1. Experimental Animals and Treatment
3.1.1. Experimental Animals
3.1.2. Exposure of Neonatal Rats to Cyclic Hyperoxia
3.2. Plasma Separation and RNA Extraction
3.3. Deep Sequencing
3.4. qPCR Confirmation
3.5. Prediction and Enrichment Analyses of the Target Genes
3.6. ELISA
3.7. Statistics
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gyllensten, L.J.; Hellstrom, B.E. Experimental approach to the pathogenesis of retrolental fibroplasia. Am. J. Ophthalmol. 1955, 39, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Gole, G.A.; Browning, J.; Elts, S.M. The mouse model of oxygen-induced retinopathy: A suitable animal model for angiogenesis research. Doc. Ophthalmol. 1990, 74, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.; Whyte, R.K.; Asztalos, E.V.; Moddemann, D.; Poets, C.; Rabi, Y.; Solimano, A.; Roberts, R.S. Effects of targeting higher vs lower arterial oxygen saturations on death or disability in extremely preterm infants: A randomized clinical trial. JAMA 2013, 309, 2111–2120. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Smith, L.E. Retinopathy of prematurity. Angiogenesis 2007, 10, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.J.; Wang, K.; Koroglu, O.; di Fiore, J.; Kc, P. Intermittent hypoxic episodes in preterm infants: Do they matter? Neonatology 2011, 100, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Barnett, J.M.; Yanni, S.E.; Penn, J.S. The development of the rat model of retinopathy of prematurity. Doc. Ophthalmol. 2010, 120, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Sanchez, M.A.; Liu, J.; Hannon, G.J.; Parker, R. Control of translation and mRNA degradation by mirnas and sirnas. Genes Dev. 2006, 20, 515–524. [Google Scholar] [CrossRef]
- Ambros, V. MicroRNA pathways in flies and worms: Growth, death, fat, stress, and timing. Cell 2003, 113, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Griffiths-Jones, S. Mirbase: Integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011, 39, D152–D157. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.; Mitra, R.; Maulik, U.; Zhang, M.Q. Development of the human cancer microRNA network. Silence 2010, 1, 6. [Google Scholar] [CrossRef] [PubMed]
- Malleter, M.; Jacquot, C.; Rousseau, B.; Tomasoni, C.; Juge, M.; Pineau, A.; Sakanian, V.; Roussakis, C. Mirnas, a potential target in the treatment of non-small-cell lung carcinomas. Gene 2012, 506, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Karali, M.; Peluso, I.; Marigo, V.; Banfi, S. Identification and characterization of microRNAs expressed in the mouse eye. Investig. Ophthalmol. Vis. Sci. 2007, 48, 509–515. [Google Scholar] [CrossRef]
- Ryan, D.G.; Oliveira-Fernandes, M.; Lavker, R.M. MicroRNAs of the mammalian eye display distinct and overlapping tissue specificity. Mol. Vis. 2006, 12, 1175–1184. [Google Scholar] [PubMed]
- Shen, J.; Yang, X.; Xie, B.; Chen, Y.; Swaim, M.; Hackett, S.F.; Campochiaro, P.A. MicroRNAs regulate ocular neovascularization. Mol. Ther. 2008, 16, 1208–1216. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Bai, X.; Wang, Z.; Zhang, X.; Ruan, C.; Miao, J. MicroRNA-126 inhibits ischemia-induced retinal neovascularization via regulating angiogenic growth factors. Exp. Mol. Pathol. 2011, 91, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Tea, M.; Michael, M.Z.; Brereton, H.M.; Williams, K.A. Stability of small non-coding RNA reference gene expression in the rat retina during exposure to cyclic hyperoxia. Mol. Vis. 2013, 19, 501–508. [Google Scholar] [PubMed]
- Kosaka, N.; Iguchi, H.; Ochiya, T. Circulating microRNA in body fluid: A new potential biomarker for cancer diagnosis and prognosis. Cancer Sci. 2010, 101, 2087–2092. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.C. Circulating microRNAs as minimally invasive biomarkers for cancer theragnosis and prognosis. Front. Genet. 2011, 2, 7. [Google Scholar] [PubMed]
- Hu, Z.; Chen, X.; Zhao, Y.; Tian, T.; Jin, G.; Shu, Y.; Chen, Y.; Xu, L.; Zen, K.; Zhang, C.; et al. Serum microRNA signatures identified in a genome-wide serum microRNA expression profiling predict survival of non-small-cell lung cancer. J. Clin. Oncol. 2010, 28, 1721–1726. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xu, J.; Wang, J.; Gortner, L.; Zhang, S.; Wei, X.; Song, J.; Zhang, Y.; Li, Q.; Feng, Z. Reduction of microRNA-206 contributes to the development of bronchopulmonary dysplasia through up-regulation of fibronectin 1. PLoS One 2013, 8, e74750. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.T.; Chen, W.J.; Hsieh, W.S.; Tsao, P.N.; Yu, S.L.; Lai, C.Y.; Lee, W.C.; Jeng, S.F. MicroRNA expression aberration associated with bronchopulmonary dysplasia in preterm infants: A preliminary study. Respir. Care 2013, 58, 1527–1535. [Google Scholar] [CrossRef] [PubMed]
- Penn, J.S.; Tolman, B.L.; Henry, M.M. Oxygen-induced retinopathy in the rat: Relationship of retinal nonperfusion to subsequent neovascularization. Investig. Ophthalmol. Vis. Sci. 1994, 35, 3429–3435. [Google Scholar]
- Reynaud, X.; Dorey, C.K. Extraretinal neovascularization induced by hypoxic episodes in the neonatal rat. Investig. Ophthalmol. Vis. Sci. 1994, 35, 3169–3177. [Google Scholar]
- Ventresca, M.R.; Gonder, J.R.; Tanswell, A.K. Oxygen-induced proliferative retinopathy in the newborn rat. Can. J. Ophthalmol. 1990, 25, 186–189. [Google Scholar] [PubMed]
- Penn, J.S.; Henry, M.M.; Wall, P.T.; Tolman, B.L. The range of PaO2 variation determines the severity of oxygen-induced retinopathy in newborn rats. Investig. Ophthalmol. Vis. Sci. 1995, 36, 2063–2070. [Google Scholar]
- Ge, Q.; Bai, Y.; Liu, Z.; Liu, Q.; Yan, L.; Lu, Z. Detection of fetal DNA in maternal plasma by microarray coupled with emulsions PCR. Clin. Chim. Acta 2006, 369, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Oshlack, A.; Robinson, M.D.; Young, M.D. From RNA-seq reads to differential expression results. Genome Biol. 2010, 11, 220. [Google Scholar] [CrossRef] [PubMed]
- Jima, D.D.; Zhang, J.; Jacobs, C.; Richards, K.L.; Dunphy, C.H.; Choi, W.W.; Au, W.Y.; Srivastava, G.; Czader, M.B.; Rizzieri, D.A.; et al. Deep sequencing of the small RNA transcriptome of normal and malignant human B cells identifies hundreds of novel microRNAs. Blood 2010, 116, e118–e127. [Google Scholar] [CrossRef]
- Rohr, C.; Kerick, M.; Fischer, A.; Kuhn, A.; Kashofer, K.; Timmermann, B.; Daskalaki, A.; Meinel, T.; Drichel, D.; Borno, S.T.; et al. High-throughput mirna and mrna sequencing of paired colorectal normal, tumor and metastasis tissues and bioinformatic modeling of miRNA-1 therapeutic applications. PLoS One 2013, 8, e67461. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Evans, J.; Bhagwate, A.; Middha, S.; Bockol, M.; Yan, H.; Kocher, J.P. CAP-miRSeq: A comprehensive analysis pipeline for microRNA sequencing data. BMC Genomics 2014, 15, 423. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed]
- Friedlander, M.R.; Chen, W.; Adamidi, C.; Maaskola, J.; Einspanier, R.; Knespel, S.; Rajewsky, N. Discovering microRNAs from deep sequencing data using miRDeep. Nat. Biotechnol. 2008, 26, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Ridzon, D.A.; Broomer, A.J.; Zhou, Z.; Lee, D.H.; Nguyen, J.T.; Barbisin, M.; Xu, N.L.; Mahuvakar, V.R.; Andersen, M.R.; et al. Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res. 2005. [Google Scholar] [CrossRef]
- Hill, D.P. The gene ontology: Enhancements for 2011. Nucleic Acids Res. 2012, 40, D559–D564. [Google Scholar] [CrossRef] [PubMed]
- Kozubek, J.; Ma, Z.; Fleming, E.; Duggan, T.; Wu, R.; Shin, D.G.; Dadras, S.S. In-depth characterization of microRNA transcriptome in melanoma. PLoS One 2013, 8, e72699. [Google Scholar] [CrossRef] [PubMed]
- Lang, M.F.; Yang, S.; Zhao, C.; Sun, G.; Murai, K.; Wu, X.; Wang, J.; Gao, H.; Brown, C.E.; Liu, X.; et al. Genome-wide profiling identified a set of miRNAs that are differentially expressed in glioblastoma stem cells and normal neural stem cells. PLoS One 2012, 7, e36248. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Charbonneau, C.; Wei, L.; Yang, W.; Chen, Q.; Terek, R.M. CXCR4-targeted therapy inhibits VEGF expression and chondrosarcoma angiogenesis and metastasis. Mol. Cancer Ther. 2013, 12, 1163–1170. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wei, L.; Chen, Q.; Terek, R.M. CXCR4/SDF1 mediate hypoxia induced chondrosarcoma cell invasion through ERK signaling and increased MMP1 expression. Mol. Cancer 2010, 9, 17. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, R.; Qian, L.; Jiang, L. Identification of Retinopathy of Prematurity Related miRNAs in Hyperoxia-Induced Neonatal Rats by Deep Sequencing. Int. J. Mol. Sci. 2015, 16, 840-856. https://doi.org/10.3390/ijms16010840
Zhao R, Qian L, Jiang L. Identification of Retinopathy of Prematurity Related miRNAs in Hyperoxia-Induced Neonatal Rats by Deep Sequencing. International Journal of Molecular Sciences. 2015; 16(1):840-856. https://doi.org/10.3390/ijms16010840
Chicago/Turabian StyleZhao, Ruibin, Lijuan Qian, and Li Jiang. 2015. "Identification of Retinopathy of Prematurity Related miRNAs in Hyperoxia-Induced Neonatal Rats by Deep Sequencing" International Journal of Molecular Sciences 16, no. 1: 840-856. https://doi.org/10.3390/ijms16010840