
Anti-Inflammatory and Analgesic Effects of Pyeongwisan on LPS-Stimulated Murine Macrophages and Mouse Models of Acetic Acid-Induced Writhing Response and Xylene-Induced Ear Edema

Abstract
:
1. Introduction
2. Results and Discussion
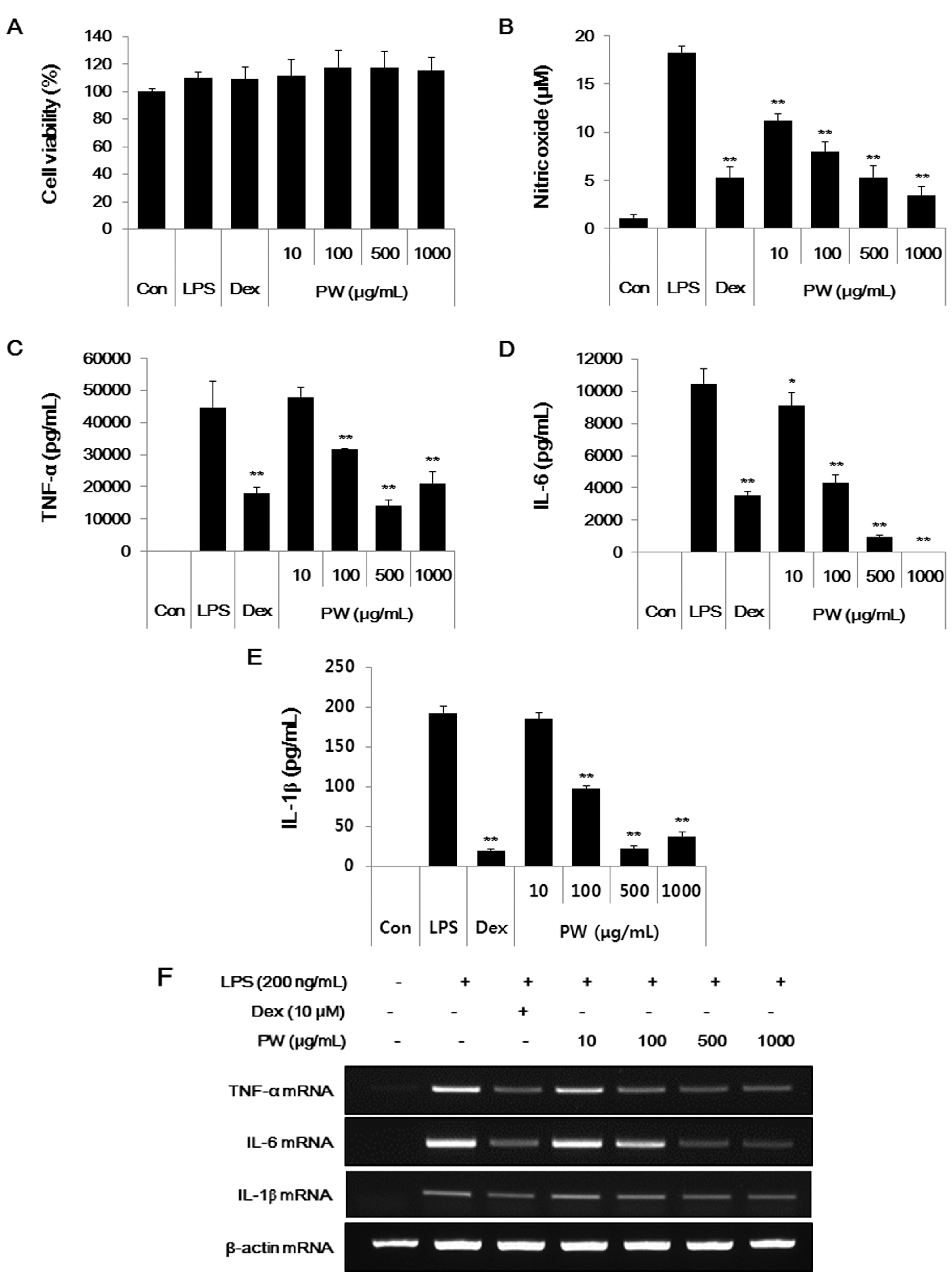
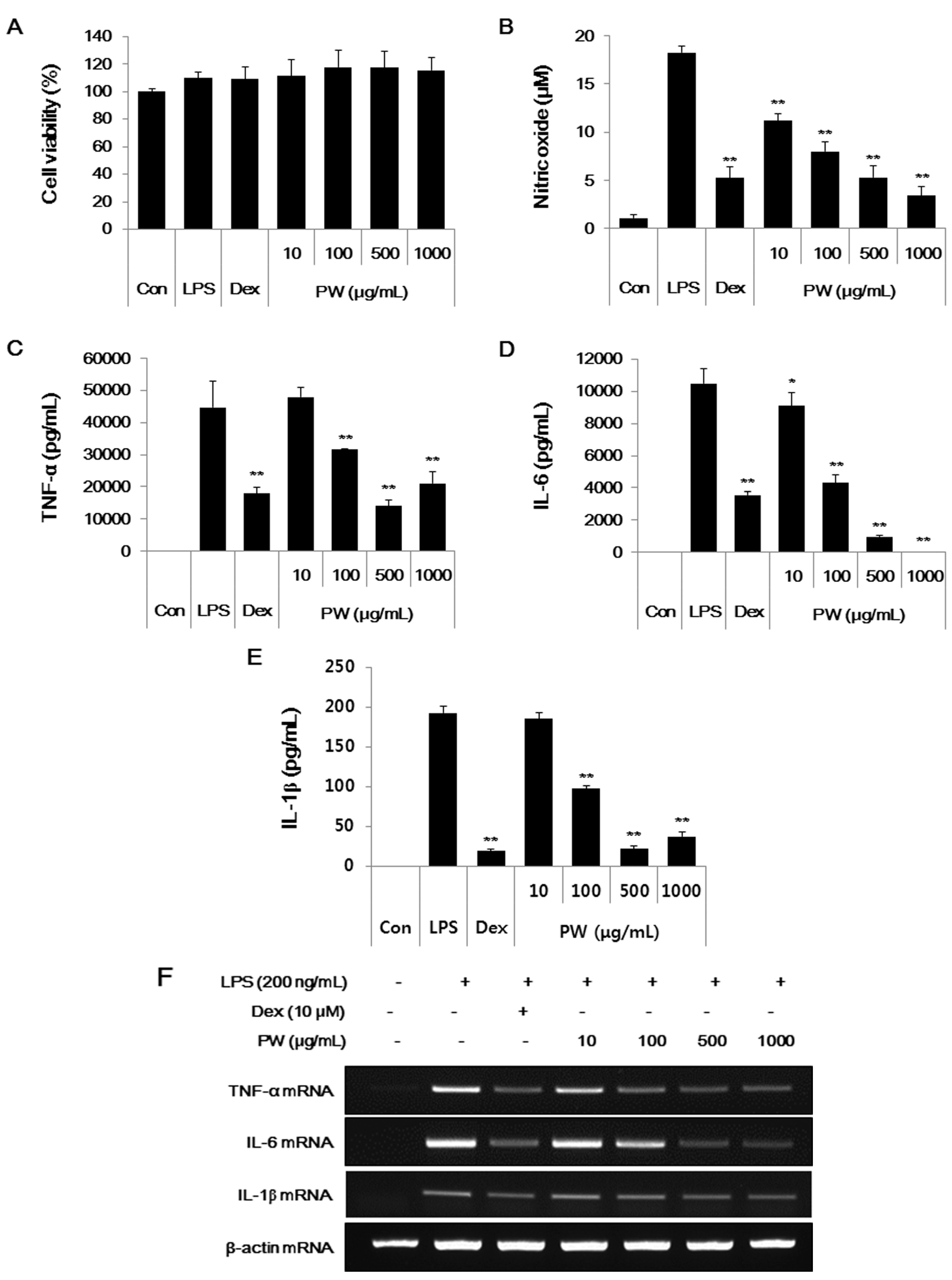
2.1. PW Is not Cytotoxic and Shows Inhibitory Effects on NO and Inflammatory Cytokine Production in RAW 264.7 Cells
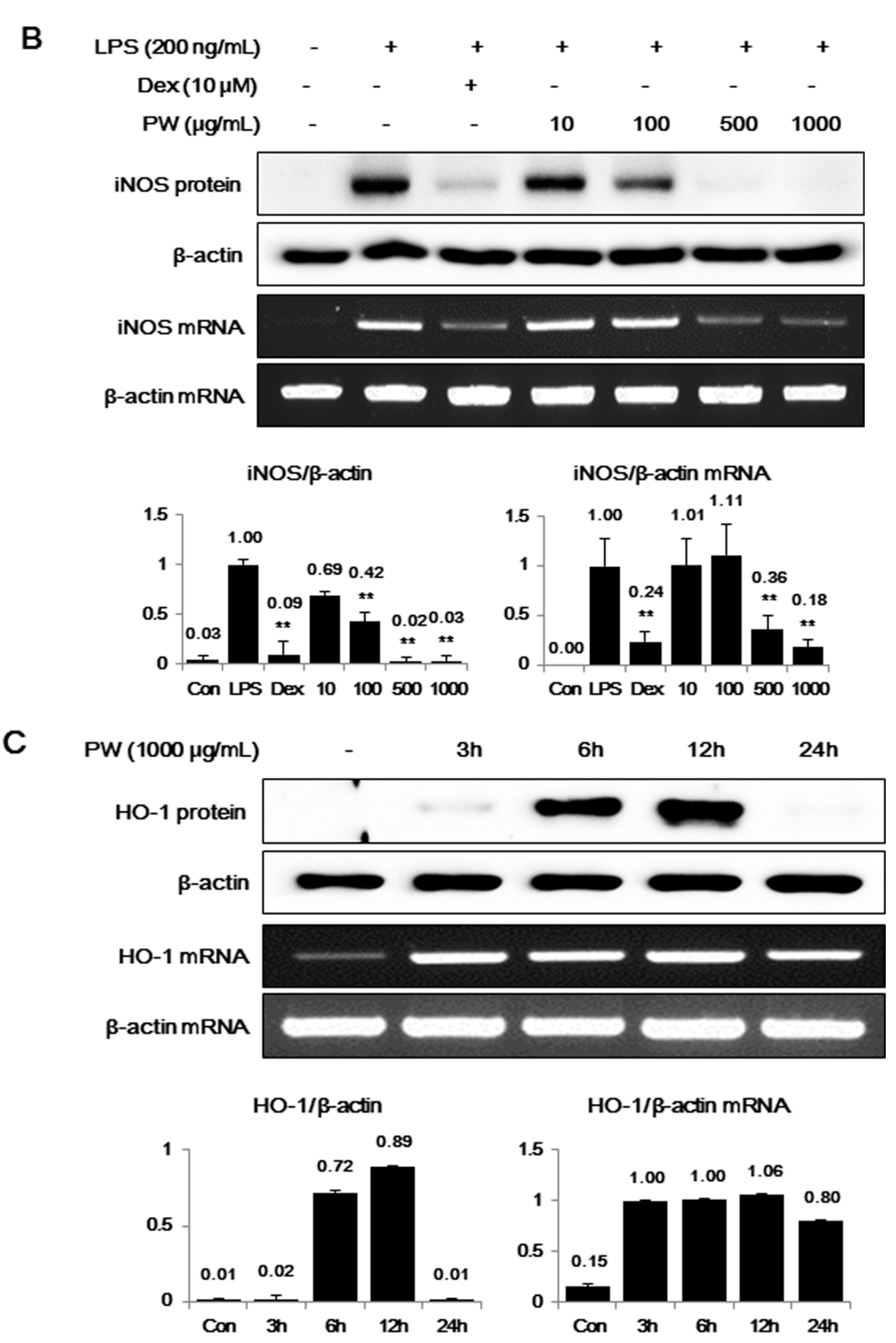
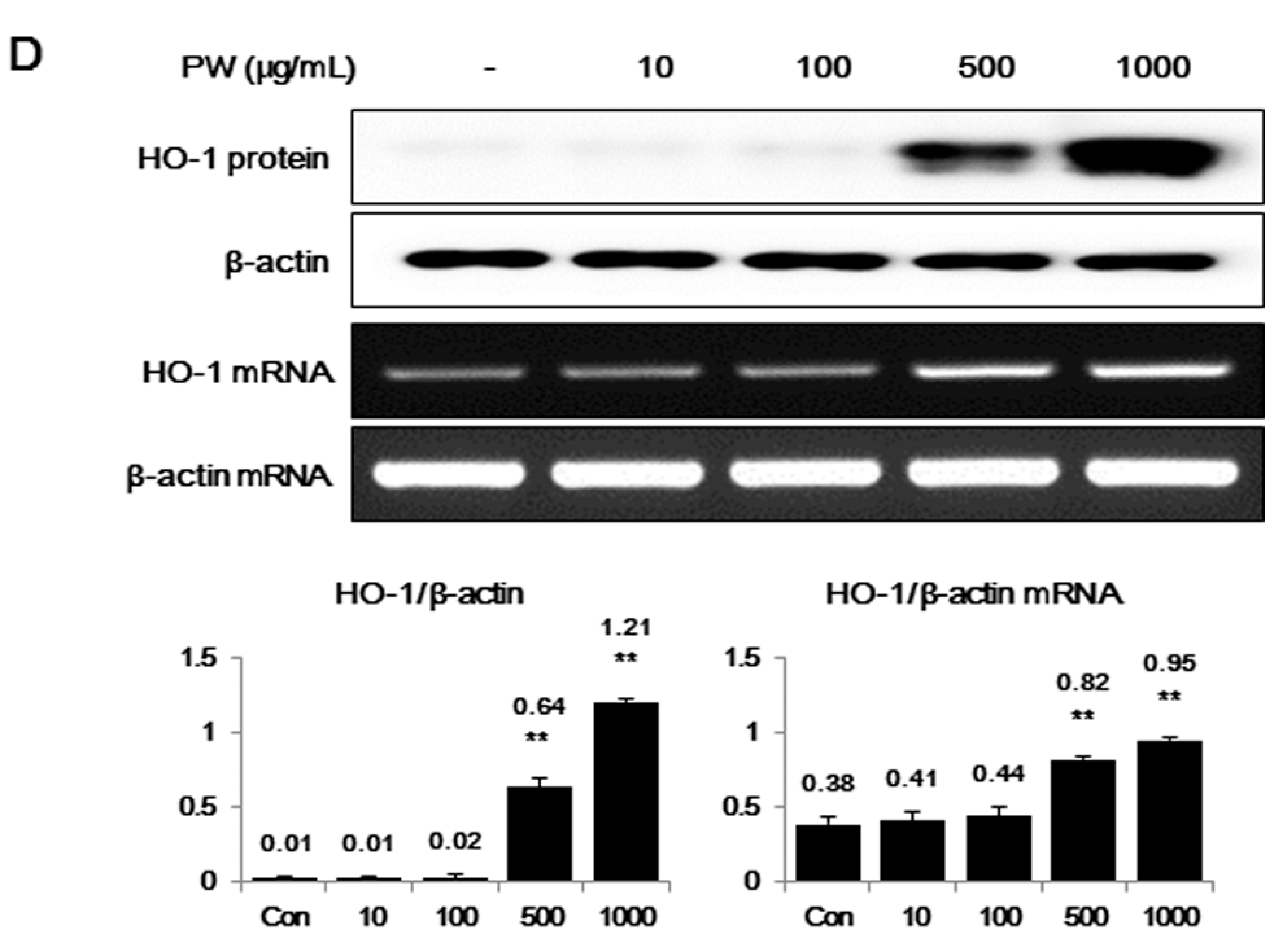
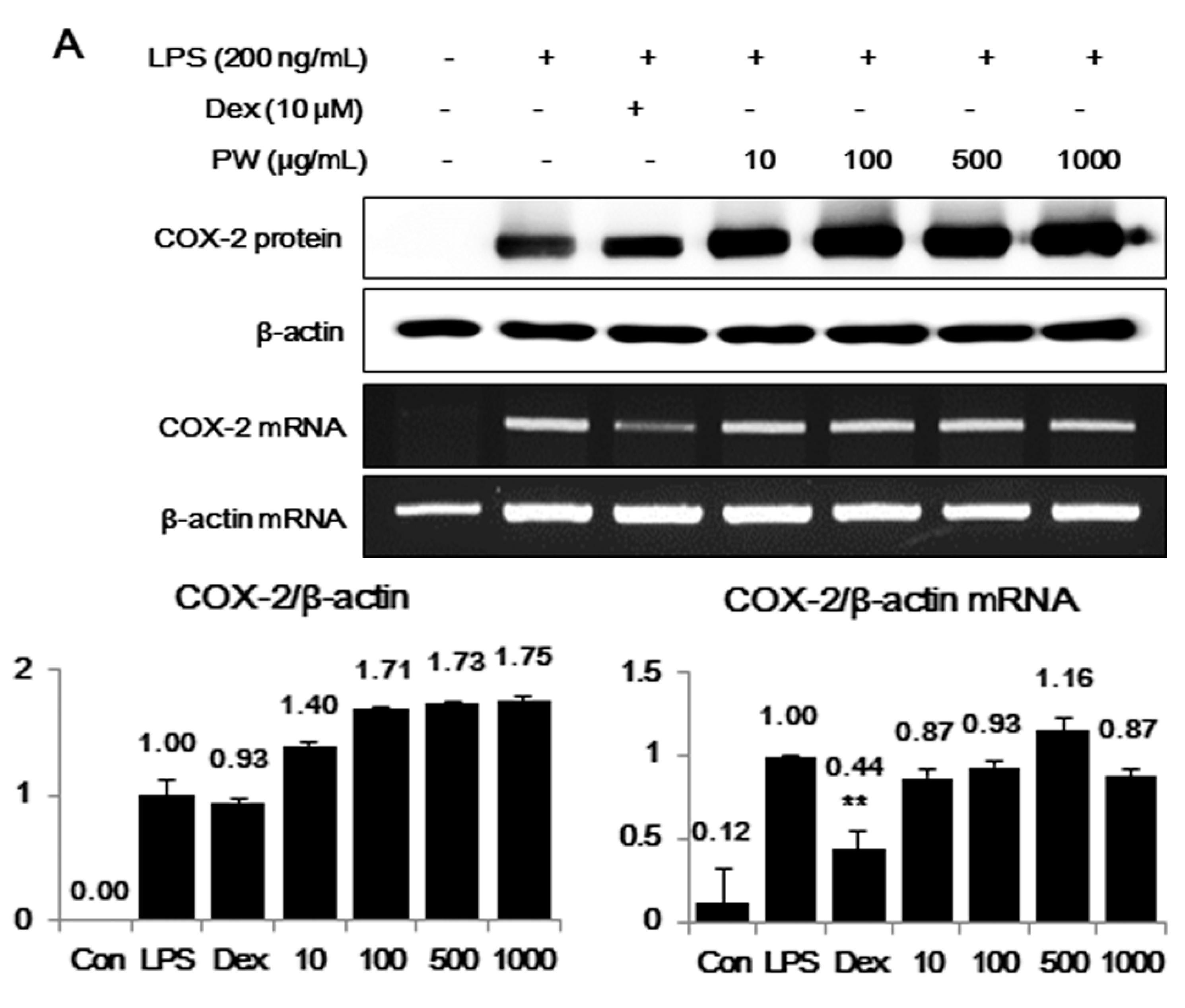
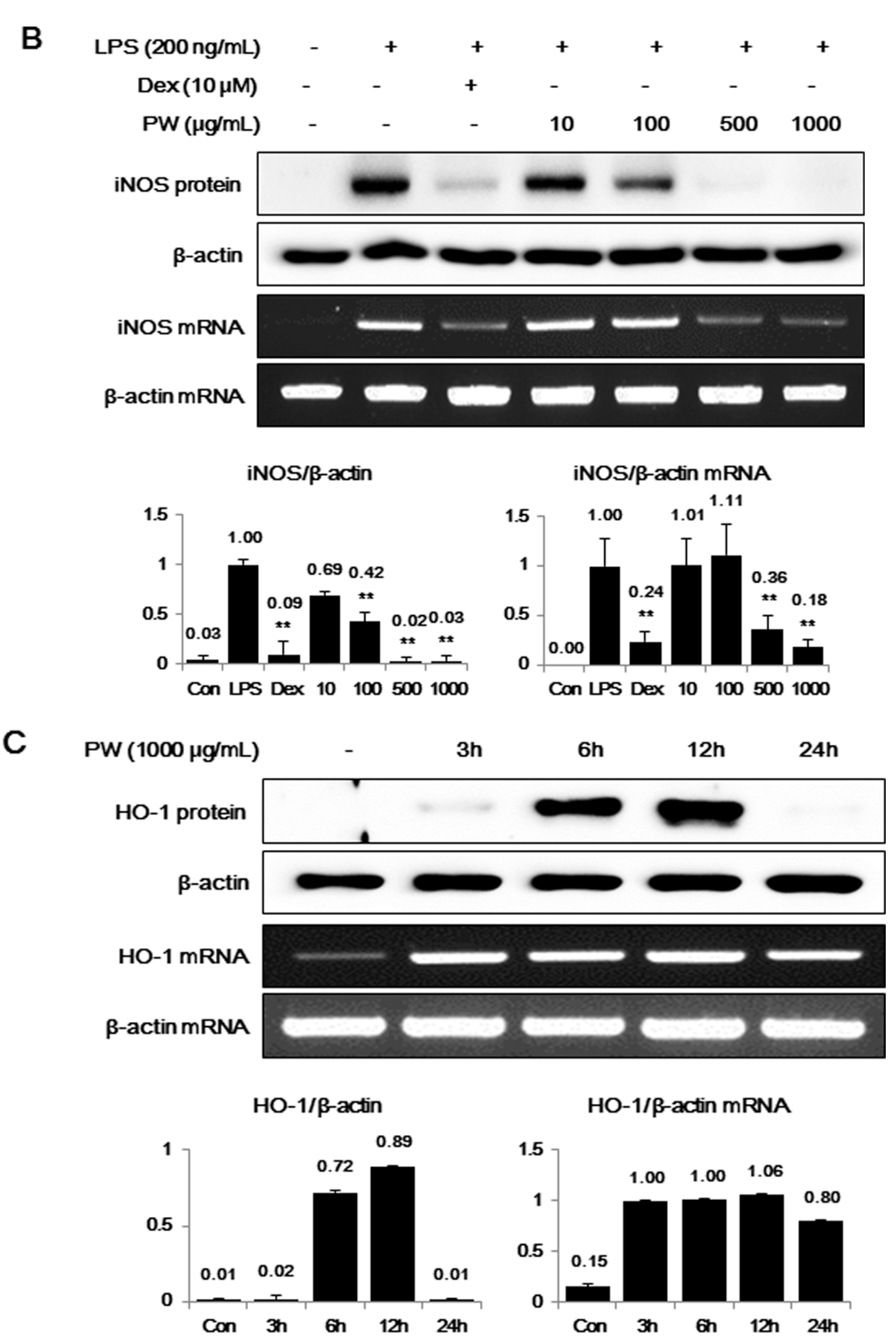
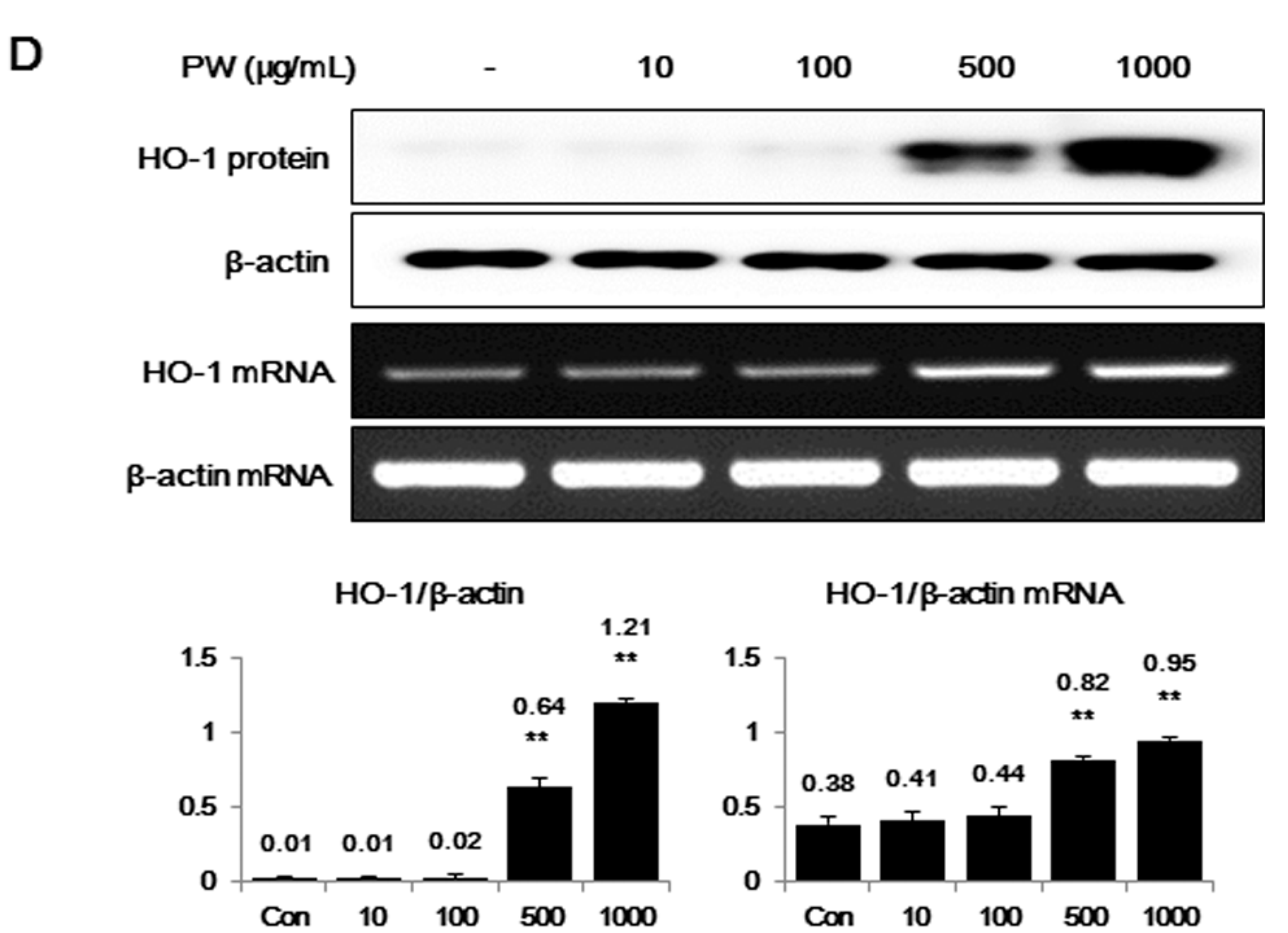
2.2. PW Strongly Inhibits LPS-Induced iNOS, but not COX-2, Expression and Induces HO-1 Induction


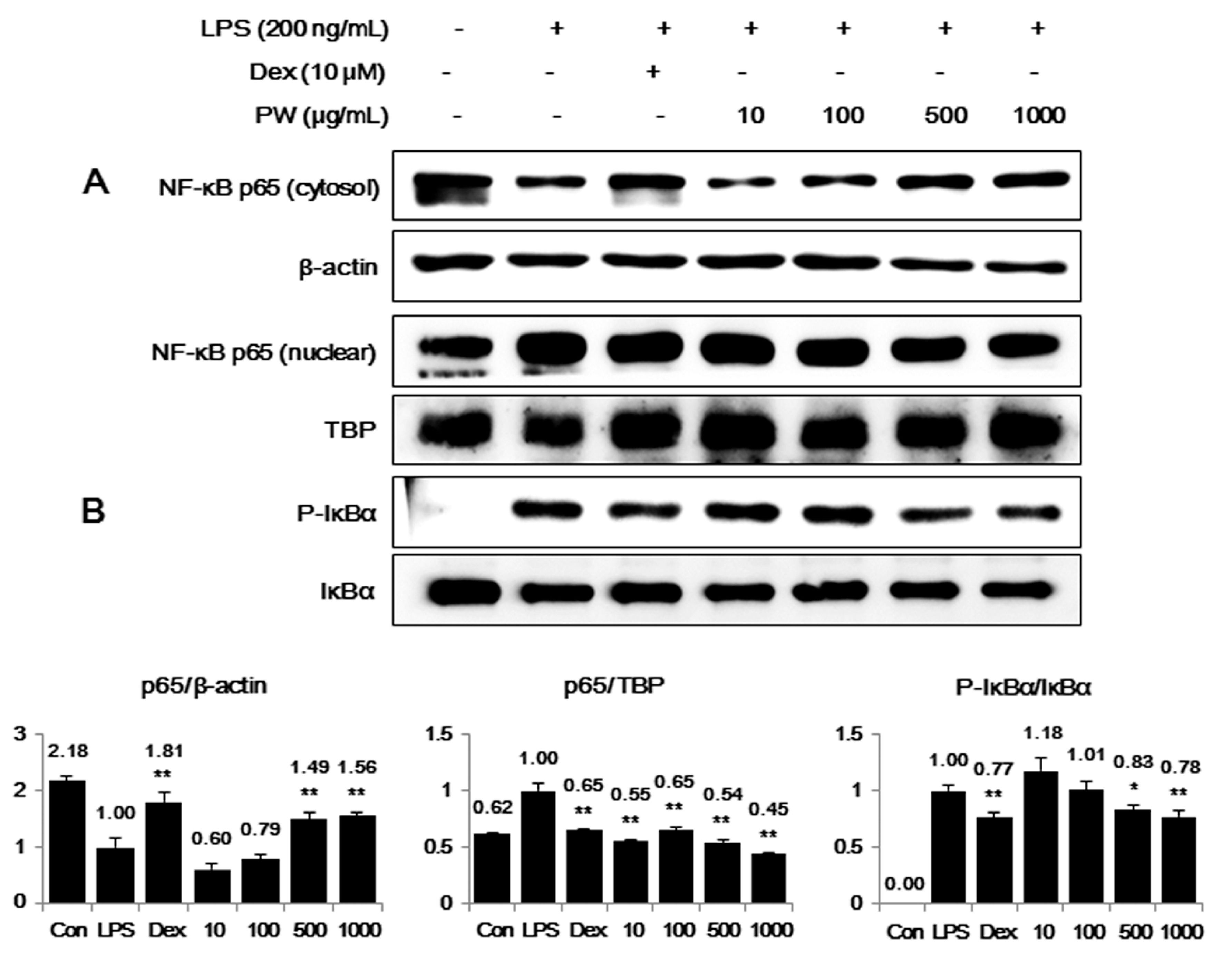
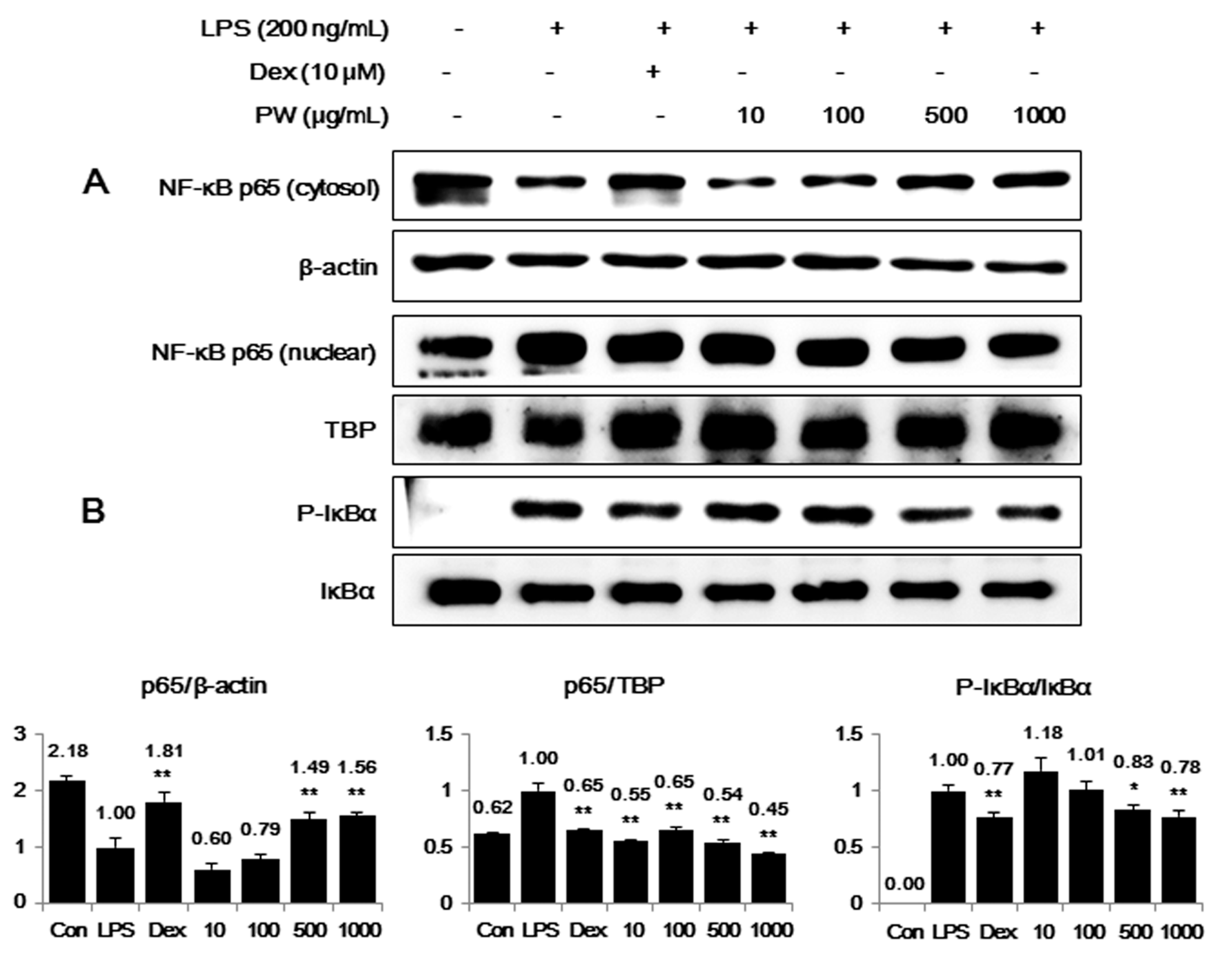
2.3. PW Inhibits NF-κB Nuclear Translocation through Suppression of IκBα Degradation and Phosphorylation in LPS-Stimulated Macrophages
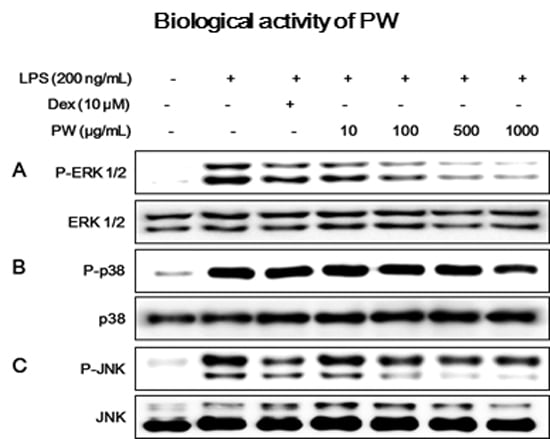
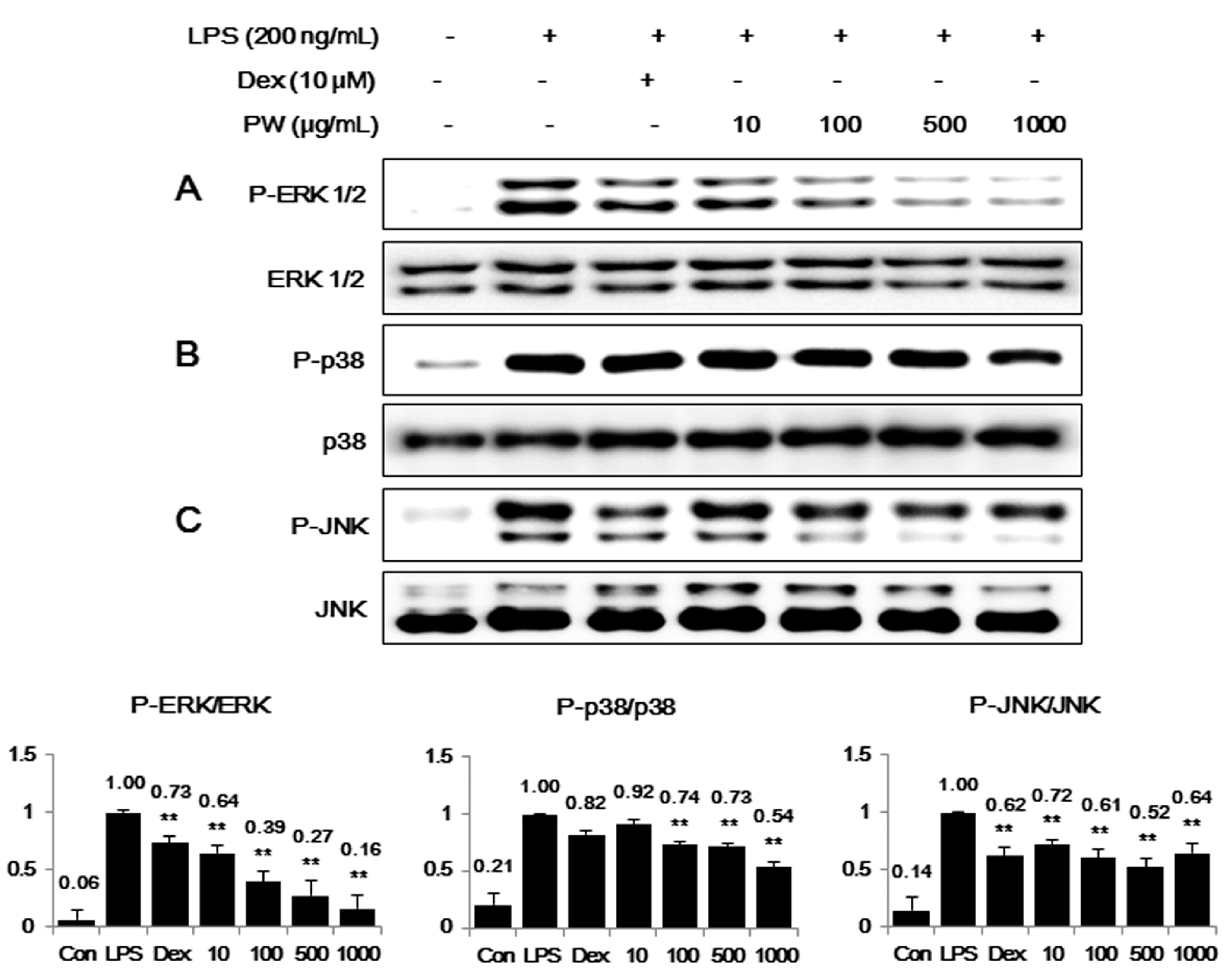
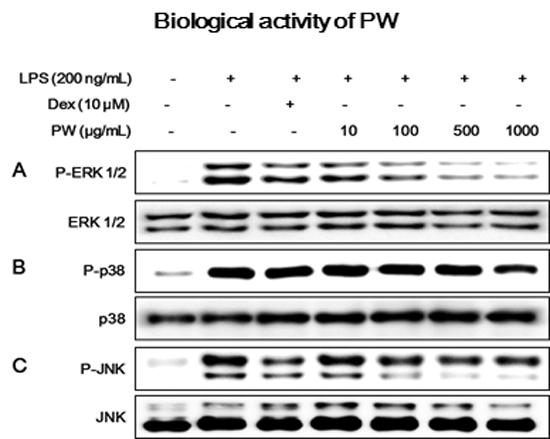
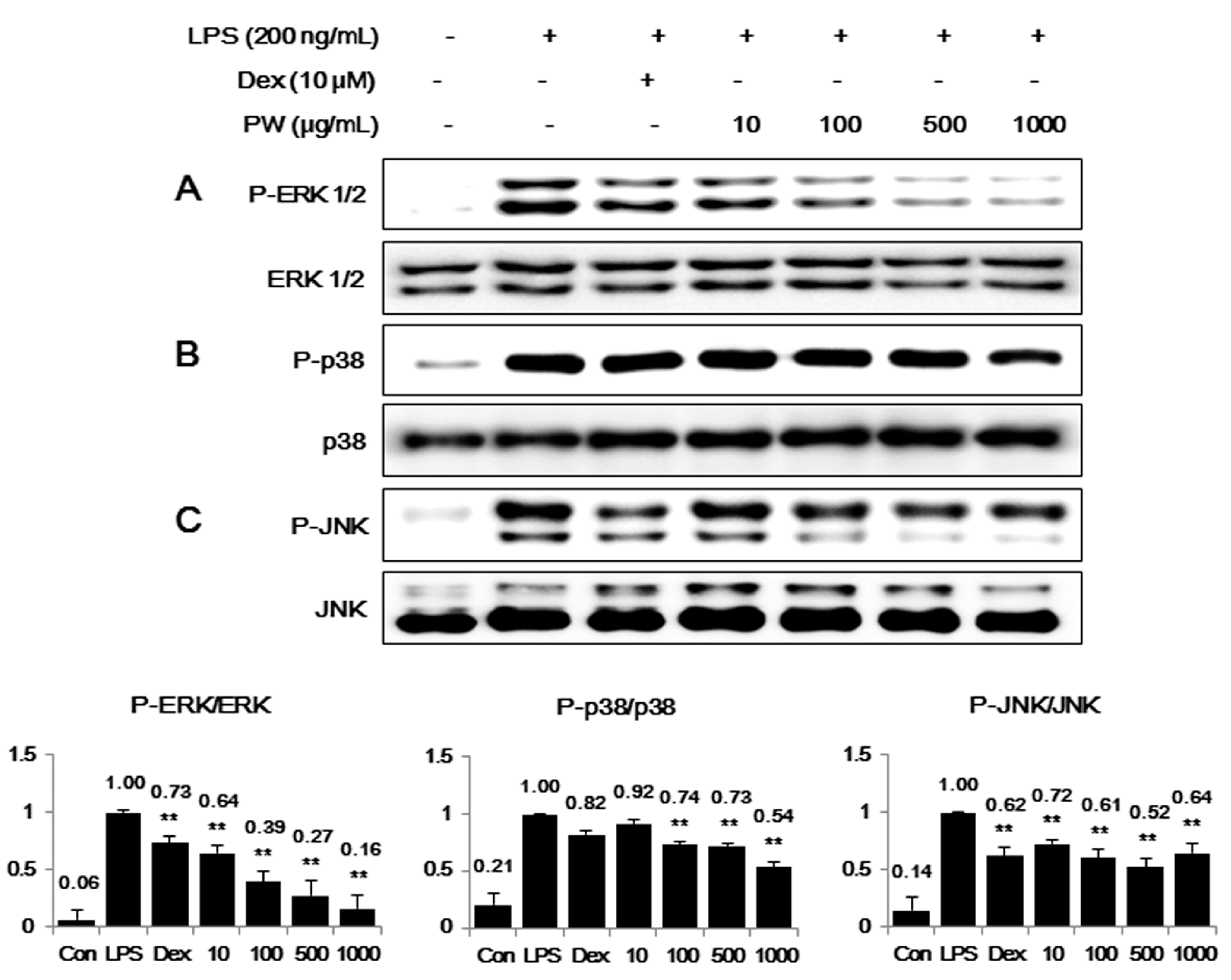
2.4. PW Suppresses Activation of MAPKs via Phosphorylation after LPS Stimulation in Macrophages
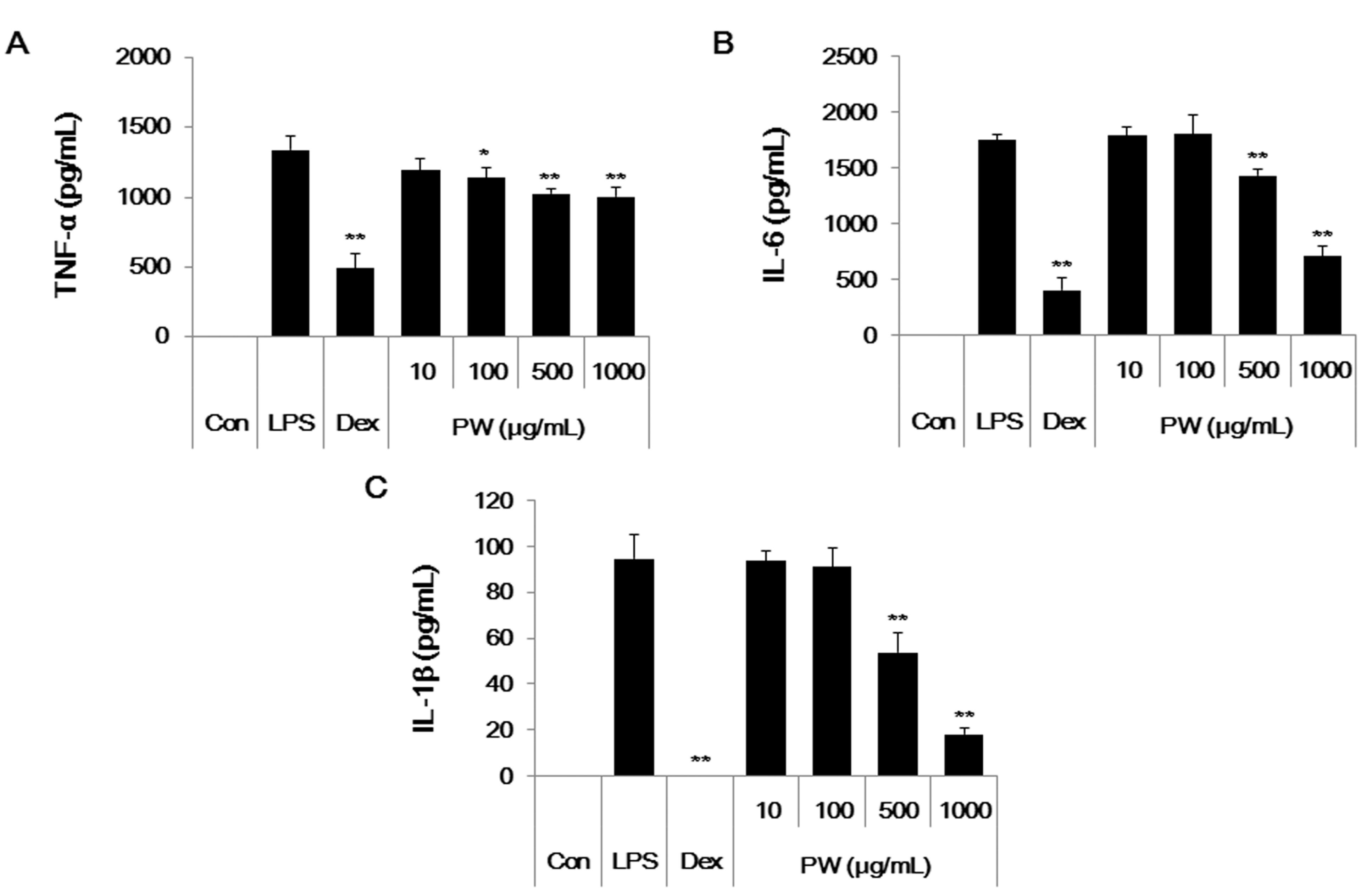
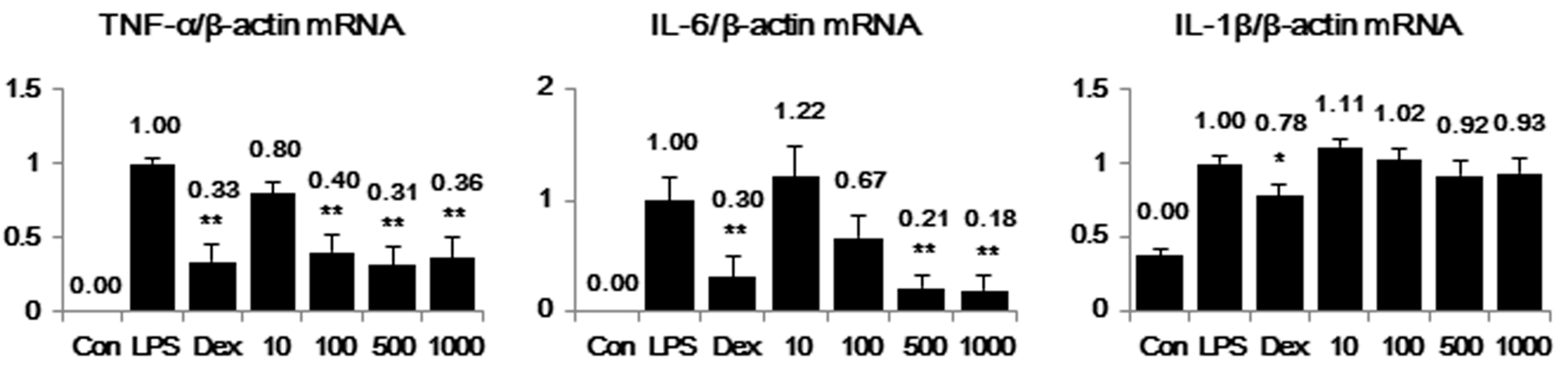
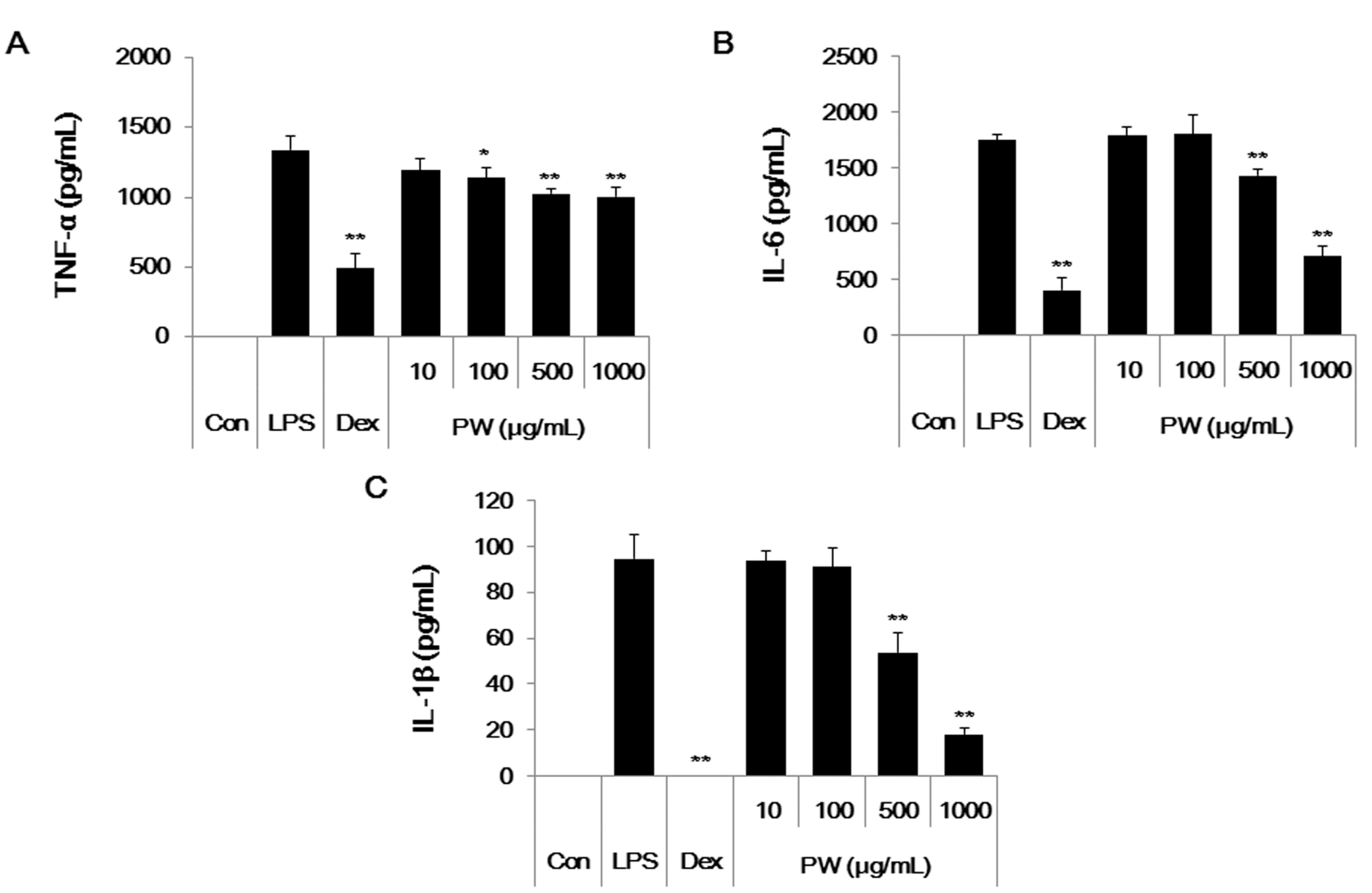
2.5. PW Represses Inflammatory Cytokine Production in LPS-Stimulated Mouse Peritoneal Macrophage Cells
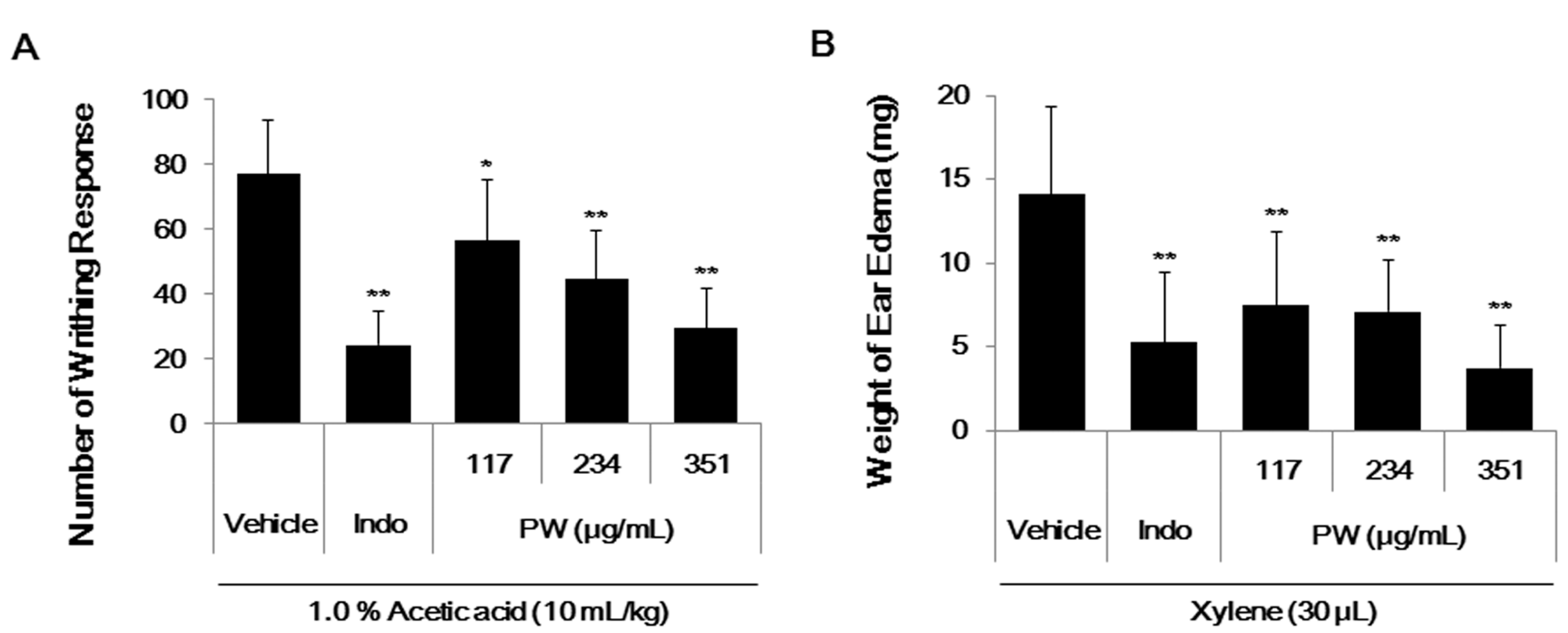
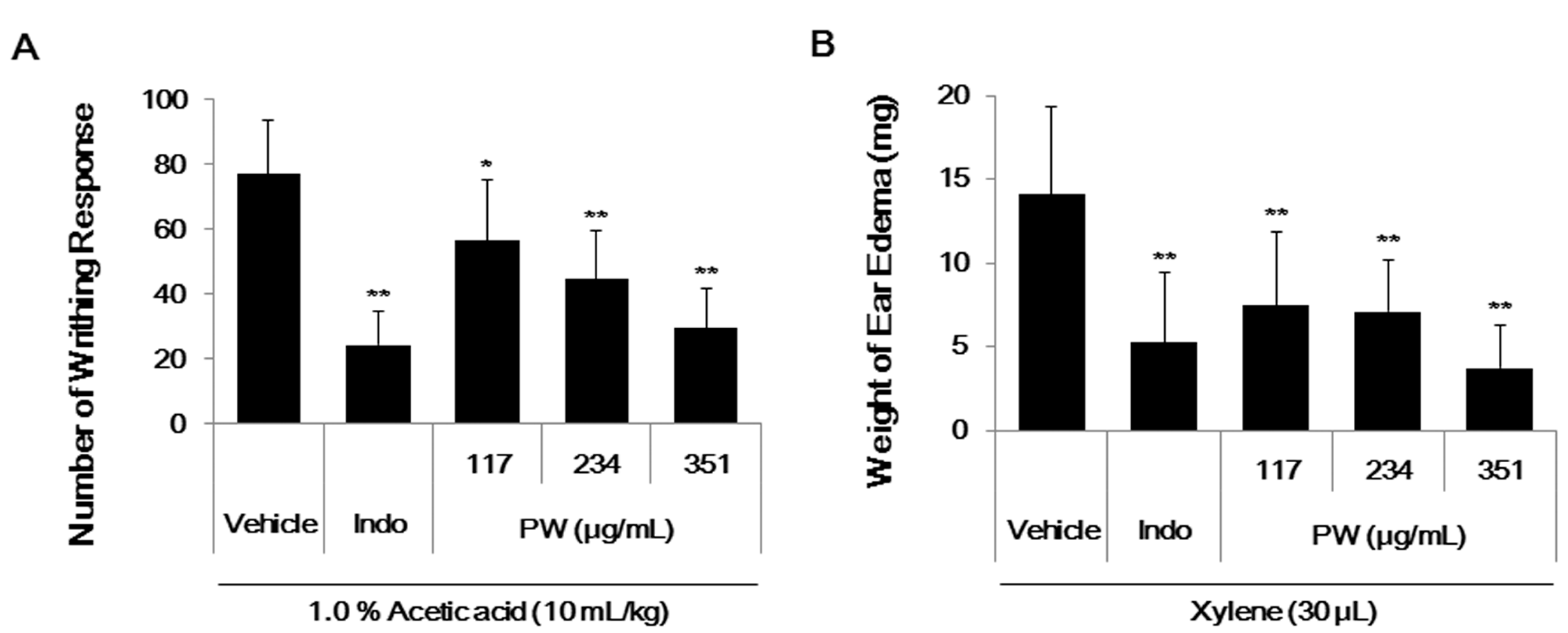
2.6. PW Showes Analgesic Activity on Acetic Acid-Induced Abdominal Writhing Response and an Inhibitory Effect on Xylene-Induced Mice Ear Edema
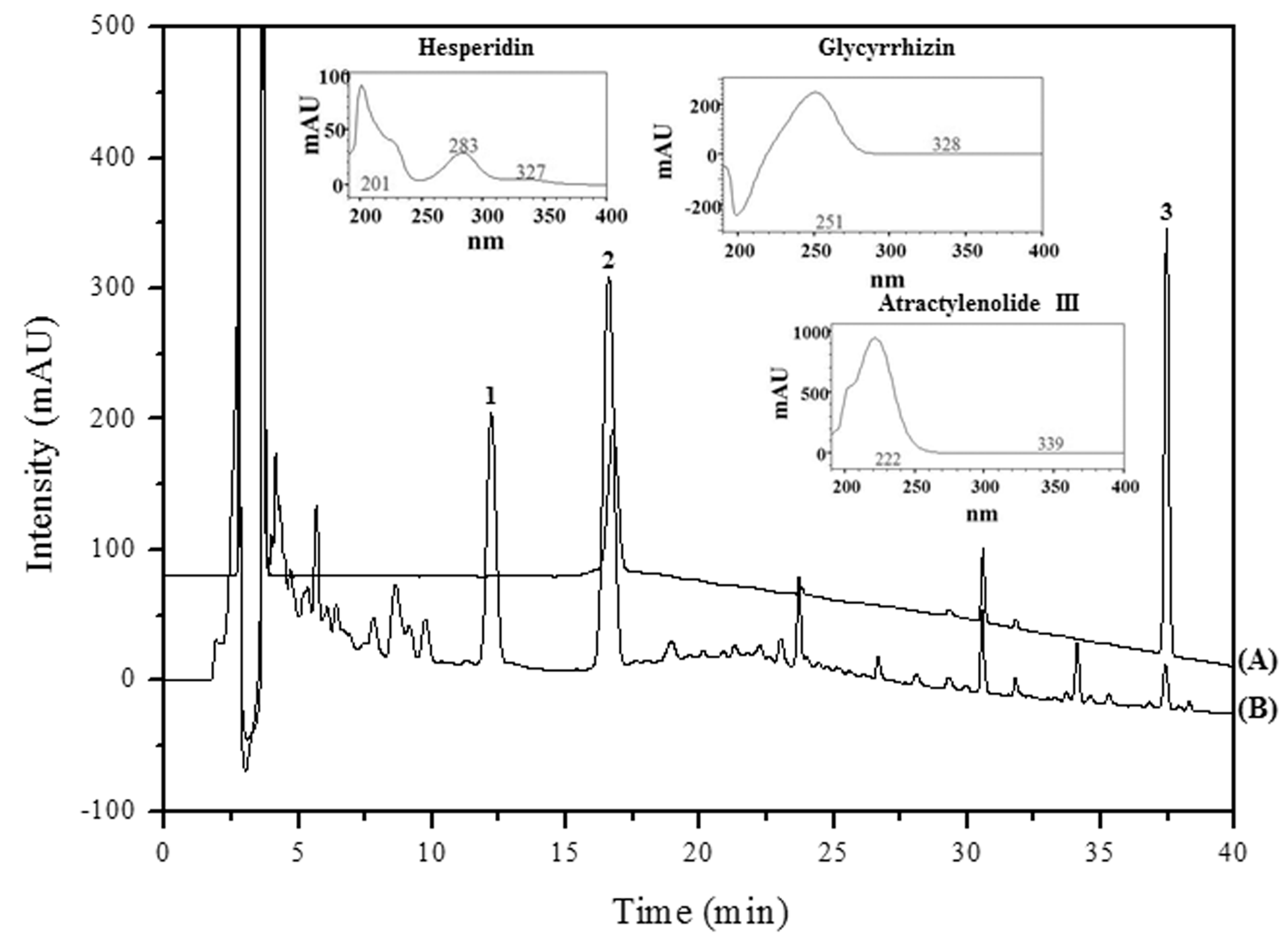
2.7. High Performance Liquid Chromatography (HPLC) Analysis and Previous Studies on the Main PW Constituents

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Condition | ||
|---|---|---|---|
| Mobile Phase | Time (min) | Water (Containing 0.1% Trifluoroacetic acid (TFA)) | Acetonitrile |
| 0 | 80 | 20 | |
| 10 | 80 | 20 | |
| 40 | 20 | 80 | |
| Flow Rate | 1.0 mL/min | ||
| Inject Volume | 20 µL | ||
| Column | OptimaPak C18 (4.6 × 250 mm, 5 µm, RS Tech Co., Daejeon, Korea) | ||
| Column Temperature | 35 °C | ||
| UV Wavelength | 230 nm | ||

3. Experimental Section
3.1. Materials and Reagents
3.2. Preparation of the PW Herbal Decoction
| Herbs | Amount of Herbs (g) |
|---|---|
| Atractylodes Rhizome | 400 |
| Citrus Unshiu Peel | 280 |
| Magnolia Bark | 200 |
| Glycyrrhizae Radix et Rhizoma | 120 |
| Zingiberis Rhizoma Recens | 74.5 |
| Zizyphi Fructus | 100 |
3.3. Cell Culture and Drug Treatment
3.4. Peritoneal Macrophage Isolation and Cell Culture
3.5. Cell Viability Assay
3.6. Measurement of NO Production
3.7. Determination of Cytokine Production
3.8. Preparation of Cytosolic and Nuclear Extracts for NF-κB Detection
3.9. Western Blot Analysis
3.10. RNA Extraction and Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
| Antibody | Corporation and Product Number | Dilution Rate |
|---|---|---|
| COX-2 | Cell signaling technology #4842 (Beverly, MA, USA) | 1:5000 |
| iNOS | Cell signaling technology #2977 | 1:1000 |
| HO-1 | Santa Cruz biotechnology #SC-10789 (Santa Cruz, CA, USA) | 1:500 |
| NF-κB p65 | Cell signaling technology #3034 | 1:1000 |
| P-IκBα | Cell signaling technology #2859 | 1:1000 |
| IκBα | Cell signaling technology #4814 | 1:1000 |
| β-actin | Santa Cruz biotechnology #SC-47778 | 1:5000 |
| TBP | Santa Cruz biotechnology #SC-33736 | 1:1000 |
| P-ERK | Cell signaling technology #4377 | 1:1000 |
| ERK | Cell signaling technology #9102 | 1:1000 |
| P-p38 | Cell signaling technology #9211 | 1:1000 |
| p38 | Cell signaling technology #9212 | 1:1000 |
| P-JNK | Cell signaling technology #9251 | 1:1000 |
| JNK | Cell signaling technology #9252 | 1:1000 |
| Secondary anti-mouse | Thermo scientific #31437 (Hudson, NH, USA) | 1:5000 |
| Secondary anti-rabbit | Thermo scientific #31463 | 1:5000 |
| Target Gene | Primer Sequence | Annealing Temperature | |||
|---|---|---|---|---|---|
| TNF-α | F: 5'-AGCACAGAAAGCATGATCCG-3' | 55 °C | |||
| R: 5'-GTTTGCTACGACGTGGGCTA-3' | |||||
| IL-6 | F: 5'-CATGTTCTCTGGGAAATCGTGG-3' | 58 °C | |||
| R: 5'-AACGCACTAGGTTTGCCGAGTA-3' | |||||
| IL-1β | F: 5'-TGCAGAGTTCCCCAACTGGTACATC-3' | 64 °C | |||
| R: 5'-GTGCTGCCTAATGTCCCCTTGAATC-3' | |||||
| COX-2 | F: 5'-CACTCAGTTTGTTGAGTCATTC-3' | 45 °C | |||
| R: 5'-GATTAGTACTGTAGGGTTAATG-3' | |||||
| iNOS | F: 5'-AGCCCAACAATACAAATGACCCTA-3' | 56 °C | |||
| R: 5'-TTCCTGTTGTTTCTATTTCCTTTGT-3' | |||||
| HO-1 | F: 5'-TGAAGGAGGCCACCAAGGAGG-3' | 62 °C | |||
| R: 5'-AGAGGTCACCCAGGTAGCGGG-3' | |||||
| β-actin | F: 5'-ATGAAGATCCTGACCGAGCGT-3' | 58 °C | |||
| R: 5'-AACGCAGCTCAGTAACAGTCCG-3' |
3.11. Experimental Animals
3.12. Acetic Acid-Induced Abdominal Writhing Response
3.13. Xylene-Induced Mice Ear Edema
3.14. Chromatographic Separation
3.15. Preparation of Standard Solutions and Samples
3.16. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ferrero-Miliani, L.; Nielsen, O.H.; Andersen, P.S.; Girardin, S.E. Chronic inflammation: Importance of NOD2 and NALP3 in interleukin-1β generation. Clin. Exp. Immunol. 2007, 147, 227–235. [Google Scholar]
- Pierce, G.F. Macrophages: Important physiologic and pathologic sources of polypeptide growth factors. Am. J. Respir. Cell Mol. Biol. 1990, 2, 233–234. [Google Scholar]
- Wadleigh, D.J.; Reddy, S.T.; Kopp, E.; Ghosh, S.; Herschman, H.R. Transcriptional activation of the cyclooxygenase-2 gene in endotoxin-treated RAW 264.7 macrophages. J. Biol. Chem. 2000, 275, 6259–6266. [Google Scholar]
- Becker, S.; Mundandhara, S.; Devlin, R.B.; Madden, M. Regulation of cytokine production in human alveolar macrophages and airway epithelial cells in response to ambient air pollution particles: Further mechanistic studies. Toxicol. Appl. Pharmacol. 2005, 207, 269–275. [Google Scholar]
- Kim, J.B.; Han, A.R.; Park, E.Y.; Kim, J.Y.; Cho, W.; Lee, J.; Seo, E.K.; Lee, K.T. Inhibition of LPS-induced iNOS, COX-2 and cytokines expression by poncirin through the NF-κB inactivation in RAW 264.7 macrophage cells. Biol. Pharm. Bull. 2007, 30, 2345–2351. [Google Scholar]
- Terry, C.M.; Clikeman, J.A.; Hoidal, J.R.; Callahan, K.S. Effect of tumor necrosis factor-α and interleukin-1 α on heme oxygenase-1 expression in human endothelial cells. Am. J. Physiol. 1998, 274, 883–891. [Google Scholar]
- Srisook, K.; Cha, Y.N. Super-induction of HO-1 in macrophages stimulated with lipopolysaccharide by prior depletion of glutathione decreases iNOS expression and NO production. Nitric Oxide 2005, 12, 70–79. [Google Scholar]
- Kaur, H.; Hughes, M.N.; Green, C.J.; Naughton, P.; Foresti, R.; Motterlini, R. Interaction of bilirubin and biliverdin with reactive nitrogen species. FEBS Lett. 2003, 543, 113–119. [Google Scholar]
- Wang, W.W.; Smith, D.L.; Zucker, S.D. Bilirubin inhibits iNOS expression and NO production in response to endotoxin in rats. Hepatology 2004, 40, 424–433. [Google Scholar]
- Ashino, T.; Yamanaka, R.; Yamamoto, M.; Shimokawa, H.; Sekikawa, K.; Iwakura, Y.; Shioda, S.; Numazawa, S.; Yoshida, T. Negative feedback regulation of lipopolysaccharide-induced inducible nitric oxide synthase gene expression by heme oxygenase-1 induction in macrophages. Mol. Immunol. 2008, 45, 2106–2115. [Google Scholar]
- Makarov, S.S. NF-κB as a therapeutic target in chronic inflammation recent advances. Mol. Med. Today 2000, 6, 441–448. [Google Scholar]
- De Martin, R.; Vanhove, B.; Cheng, Q.; Hofer, E.; Csizmadia, V.; Winkler, H.; Bach, F.H. Cytokine-inducible expression in endothelial cells of an IκBα-like gene is regulated by NF-κB. EMBO J. 1993, 12, 2773–2779. [Google Scholar]
- Pahl, H.L. Activators and target genes of Rel/NF-κB transcription factors. Oncogene 1999, 18, 6853–6866. [Google Scholar]
- Baeuerle, P.A.; Baltimore, D. NF-κB: Ten years after. Cell 1996, 87, 13–20. [Google Scholar]
- Robinson, M.J.; Cobb, M.H. Mitogen-activated protein kinase pathways. Curr. Opin. Cell Biol. 1997, 9, 180–186. [Google Scholar]
- Yang, S.H.; Yu, C.L. Antiinflammatory effects of Bu-zhong-yi-qi-tang in patients with perennial allergic rhinitis. J. Ethnopharmacol. 2008, 115, 104–109. [Google Scholar]
- Ji, H.C.; Baek, T.H. A comparative study of Pyeongwi-san, Ijin-tang and Pyeongjintang extracts on indomethacin-induced gastric mucosal lesions in mice. J. Korean Orient. Med. 2011, 32, 102–117. [Google Scholar]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar]
- Southan, G.J.; Szabo, C. Selective pharmacological inhibition of distinct nitric oxide synthase isoforms. Biochem. Pharmacol. 1996, 51, 383–394. [Google Scholar]
- Brasier, A.R. The NF-κB regulatory network. Cardiovasc. Toxicol. 2006, 6, 111–130. [Google Scholar]
- Gilmore, T.D. Introduction to NF-κB: Players, pathways, perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar]
- Tian, B.; Brasier, A.R. Identification of a nuclear factor kappa B-dependent gene network. Recent Prog. Horm. Res. 2003, 58, 95–130. [Google Scholar]
- Chen, F.; Kuhn, D.C.; Sun, S.C.; Gaydos, L.J.; Demers, L.M. Dependence and reversal of nitric oxide production on NF-κB in silica and lipopolysaccharide-induced macrophages. Biochem. Biophys. Res. Commun. 1995, 214, 839–846. [Google Scholar]
- Roshak, A.K.; Jackson, J.R.; McGough, K.; Chabot-Fletcher, M.; Mochan, E.; Marshall, L.A. Manipulation of distinct NF-κB proteins alters interleukin-1β-induced human rheumatoid synovial fibroblast prostaglandin E2 formation. J. Biol. Chem. 1996, 271, 31496–31501. [Google Scholar]
- Xie, W.; Merrill, J.R.; Bradshaw, W.S.; Simmons, D.L. Structural determination and promoter analysis of the chicken mitogen-inducible prostaglandin G/H synthase gene and genetic mapping of the murine homolog. Arch. Biochem. Biophys. 1993, 300, 247–252. [Google Scholar]
- Ahn, K.S.; Noh, E.J.; Zhao, H.L.; Jung, S.H.; Kang, S.S.; Kim, Y.S. Inhibition of inducible nitric oxide synthase and cyclooxygenase II by Platycodon grandiflorum saponins via suppression of nuclear factor-κB activation in RAW 264.7 cells. Life Sci. 2005, 76, 2315–2328. [Google Scholar]
- Barnes, P.J.; Karin, M. Nuclear factor-κB. A pivotal transcription factor in chronic inflammatory diseases. N. Engl. J. Med. 1997, 336, 1066–1071. [Google Scholar]
- Kim, Y.M.; Lee, B.S.; Yi, K.Y.; Paik, S.G. Upstream NF-κB site is required for the maximal expression of mouse inducible nitric oxide synthase gene in interferon-γ plus lipopolysaccharide induced RAW 264.7 macrophages. Biochem. Pharmacol. 1997, 236, 655–660. [Google Scholar]
- Caivano, M. Role of MAP kinase cascades in inducing arginine transporters and nitric oxide synthetase in RAW 264.7 macrophages. FEBS Lett. 1999, 429, 249–253. [Google Scholar]
- Saiprasad, G.; Chitra, P.; Manikandan, R.; Sudhandiran, G. Hesperidin alleviates oxidative stress and downregulates the expressions of proliferative and inflammatory markers in azoxymethane-induced experimental colon carcinogenesis in mice. Inflamm. Res. 2013, 62, 425–440. [Google Scholar]
- Kato, T.; Horie, N.; Hashimoto, K.; Satoh, K.; Shimoyama, T.; Kaneko, T.; Kusama, K.; Sakagami, H. Bimodal effect of glycyrrhizin on macrophage nitric oxide and prostaglandin E2 production. In Vivo 2008, 22, 583–586. [Google Scholar]
- Li, C.Q.; He, L.C.; Jin, J.Q. Atractylenolide I and atractylenolide III inhibit Lipopolysaccharide-induced TNF-α and NO production in macrophages. Phytother. Res. 2007, 21, 347–353. [Google Scholar]
- Fu, Y.; Liu, B.; Liu, J.; Liu, Z.; Liang, D.; Li, F.; Li, D.; Cao, Y.; Zhang, X.; Zhang, N.; et al. Geniposide, from Gardenia jasminoides Ellis, inhibits the inflammatory response in the primary mouse macrophages and mouse models. Int. Immunopharmacol. 2012, 14, 792–798. [Google Scholar]
- Choi, H.J.; Kang, O.H.; Park, P.S.; Chae, H.S.; Oh, Y.C.; Lee, Y.S.; Choi, J.G.; Lee, G.H.; Kweon, O.H.; Kwon, D.Y. Mume Fructus water extract inhibits pro-inflammatory mediators in lipopolysaccharide-stimulated macrophages. J. Med. Food 2007, 10, 460–466. [Google Scholar]
- Jo, H.Y.; Kim, Y.; Nam, S.Y.; Lee, B.J.; Kim, Y.B.; Yun, Y.W.; Ahn, B. The inhibitory effect of quercitrin gallate on iNOS expression induced by lipopolysaccharide in Balb/c mice. J. Vet. Sci. 2008, 9, 267–272. [Google Scholar]
- Kim, H.Y.; Kim, J.K.; Choi, J.H.; Jung, J.Y.; Oh, W.Y.; Kim, D.C.; Lee, H.S.; Kim, Y.S.; Kang, S.S.; Lee, S.H.; et al. Hepatoprotective effect of pinoresinol on carbon tetrachloride-induced hepatic damage in mice. J. Pharmacol. Sci. 2010, 112, 105–112. [Google Scholar]
- Srisook, K.; Palachot, M.; Mongkol, N.; Srisook, E.; Sarapusit, S. Anti-inflammatory effect of ethyl acetate extract from Cissus quadrangularis Linn may be involved with induction of heme oxygenase-1 and suppression of NF-κB activation. J. Ethnopharmacol. 2011, 133, 1008–1014. [Google Scholar]
- Kwon, D.J.; Ju, S.M.; Youn, G.S.; Choi, S.Y.; Park, J. Suppression of iNOS and COX-2 expression by flavokawain A via blockade of NF-κB and AP-1 activation in RAW 264.7 macrophages. Food Chem. Toxicol. 2013, 58, 479–486. [Google Scholar]
- Collier, H.O.; Dinneen, L.C.; Johnson, C.A.; Schneider, C. The abdominal constriction response and its suppression by analgesic drugs in the mouse. Br. J. Pharmacol. Chemother. 1968, 32, 295–310. [Google Scholar]
- Fezai, M.; Senovilla, L.; Jemaà, M.; Ben-Attia, M. Analgesic, anti-inflammatory and anticancer activities of extra virgin olive oil. J. Lipids 2013. [Google Scholar] [CrossRef]
- Liao, J.C.; Chang, W.T.; Lee, M.S.; Chiu, Y.J.; Chao, W.K.; Lin, Y.C.; Lin, M.K.; Peng, W.H. Antinociceptive and anti-inflammatory activities of Cuscuta chinensis seeds in mice. Am. J. Chin. Med. 2014, 42, 223–242. [Google Scholar]
- Xu, Q.; Wang, Y.; Guo, S.; Shen, Z.; Wang, Y.; Yang, L. Anti-inflammatory and analgesic activity of aqueous extract of Flos populi. J. Ethnopharmacol. 2014, 152, 540–545. [Google Scholar]
- Kou, J.; Sun, Y.; Lin, Y.; Cheng, Z.; Zheng, W.; Yu, B.; Xu, Q. Anti-inflammatory activities of aqueous extract from Radix Ophiopogon japonicus and its two constituents. Biol. Pharm. Bull. 2005, 28, 1234–1238. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, Y.-C.; Jeong, Y.H.; Cho, W.-K.; Ha, J.-H.; Gu, M.J.; Ma, J.Y. Anti-Inflammatory and Analgesic Effects of Pyeongwisan on LPS-Stimulated Murine Macrophages and Mouse Models of Acetic Acid-Induced Writhing Response and Xylene-Induced Ear Edema. Int. J. Mol. Sci. 2015, 16, 1232-1251. https://doi.org/10.3390/ijms16011232
Oh Y-C, Jeong YH, Cho W-K, Ha J-H, Gu MJ, Ma JY. Anti-Inflammatory and Analgesic Effects of Pyeongwisan on LPS-Stimulated Murine Macrophages and Mouse Models of Acetic Acid-Induced Writhing Response and Xylene-Induced Ear Edema. International Journal of Molecular Sciences. 2015; 16(1):1232-1251. https://doi.org/10.3390/ijms16011232
Chicago/Turabian StyleOh, You-Chang, Yun Hee Jeong, Won-Kyung Cho, Jeong-Ho Ha, Min Jung Gu, and Jin Yeul Ma. 2015. "Anti-Inflammatory and Analgesic Effects of Pyeongwisan on LPS-Stimulated Murine Macrophages and Mouse Models of Acetic Acid-Induced Writhing Response and Xylene-Induced Ear Edema" International Journal of Molecular Sciences 16, no. 1: 1232-1251. https://doi.org/10.3390/ijms16011232