DADS Suppresses Human Esophageal Xenograft Tumors through RAF/MEK/ERK and Mitochondria-Dependent Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
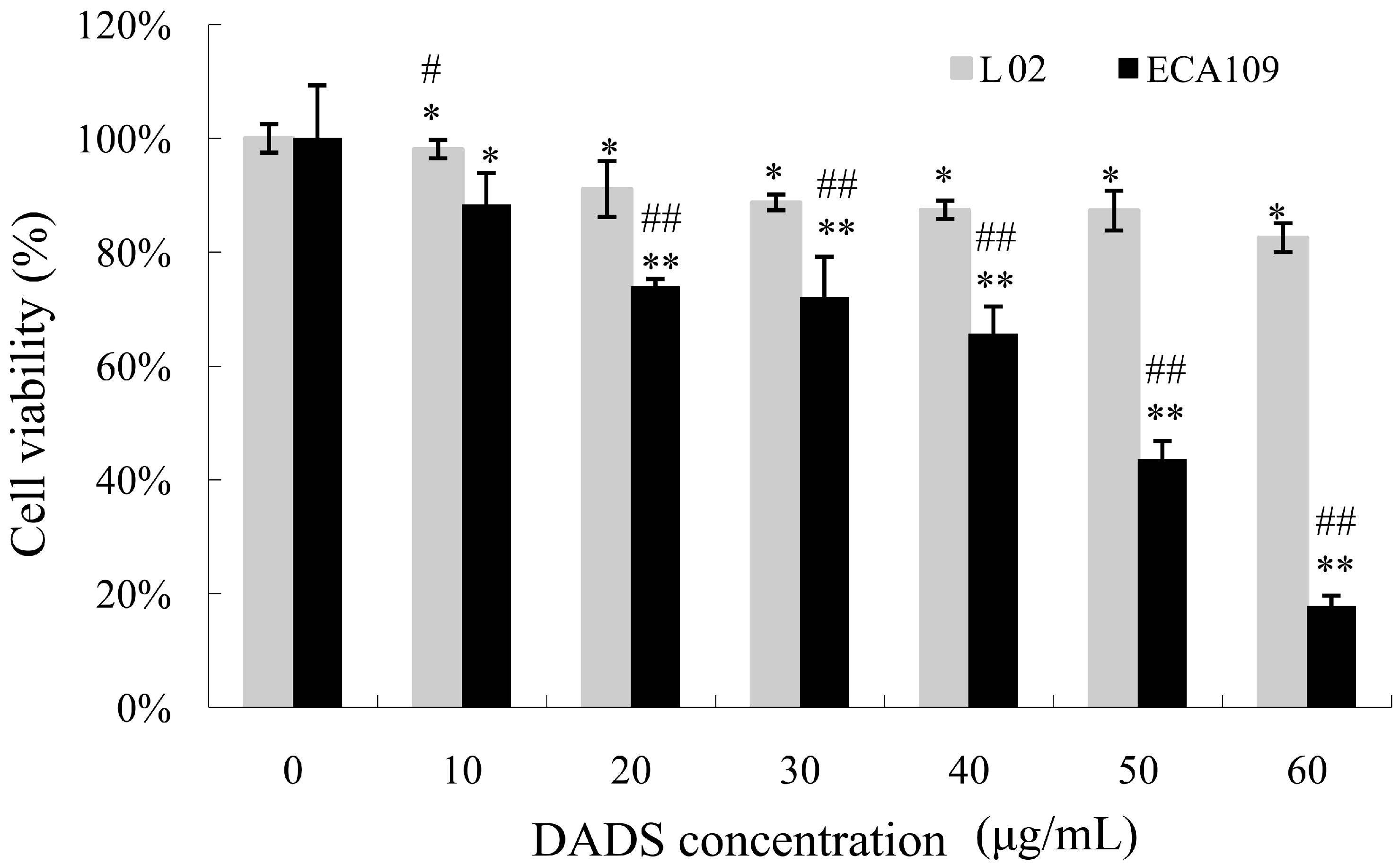
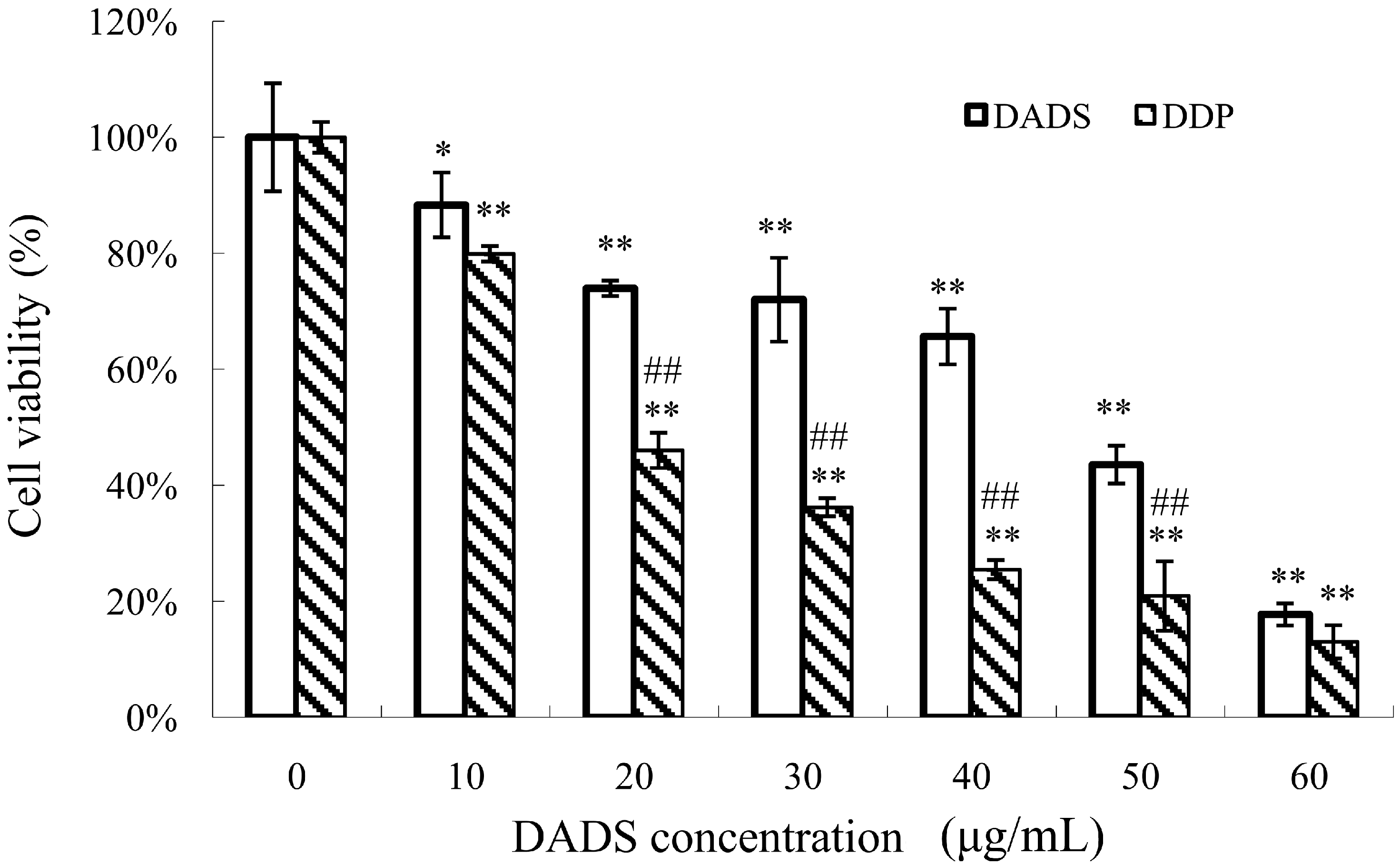
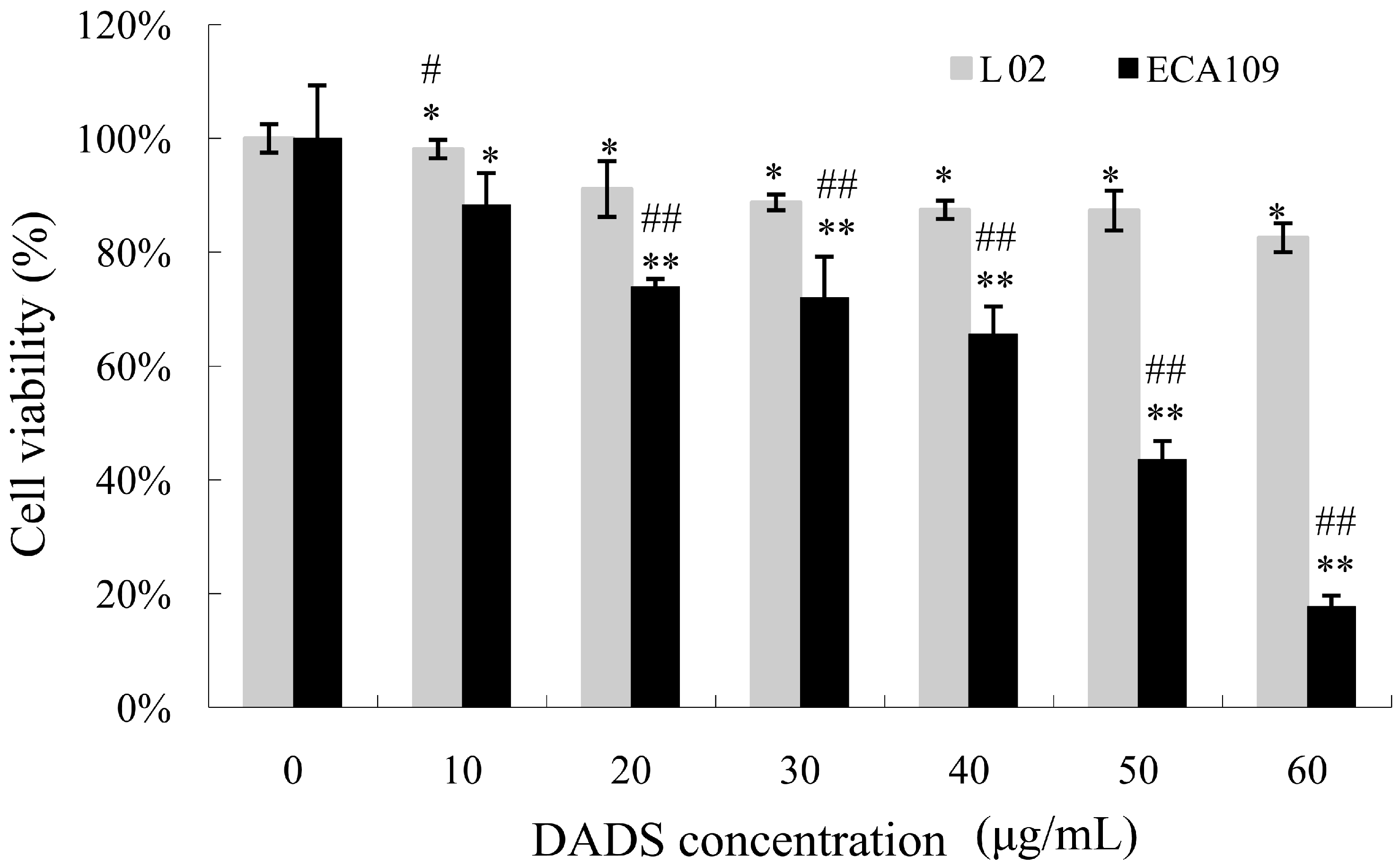
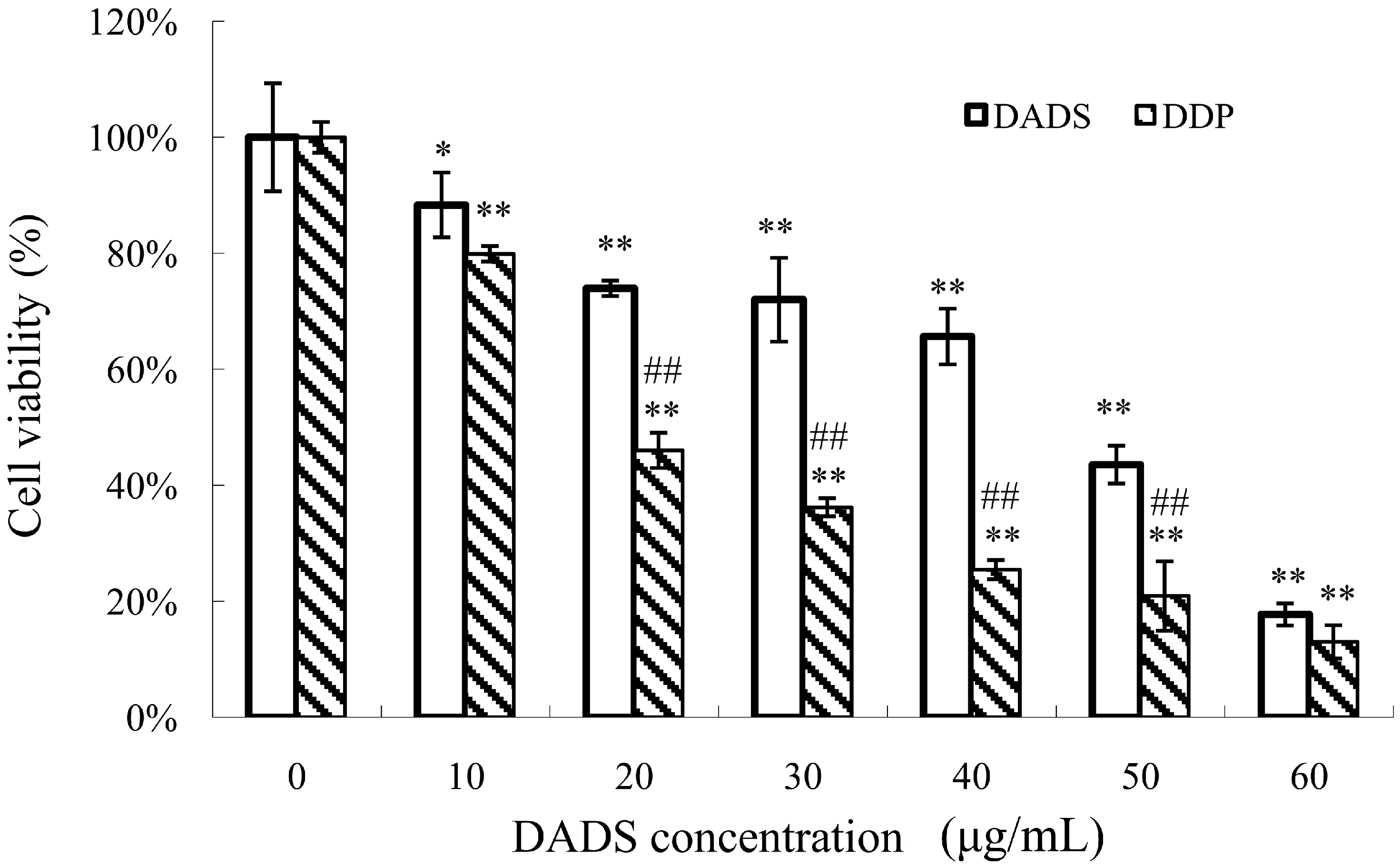
2.1. Diallyl Disulfide (DADS) Inhibits Cell Viability
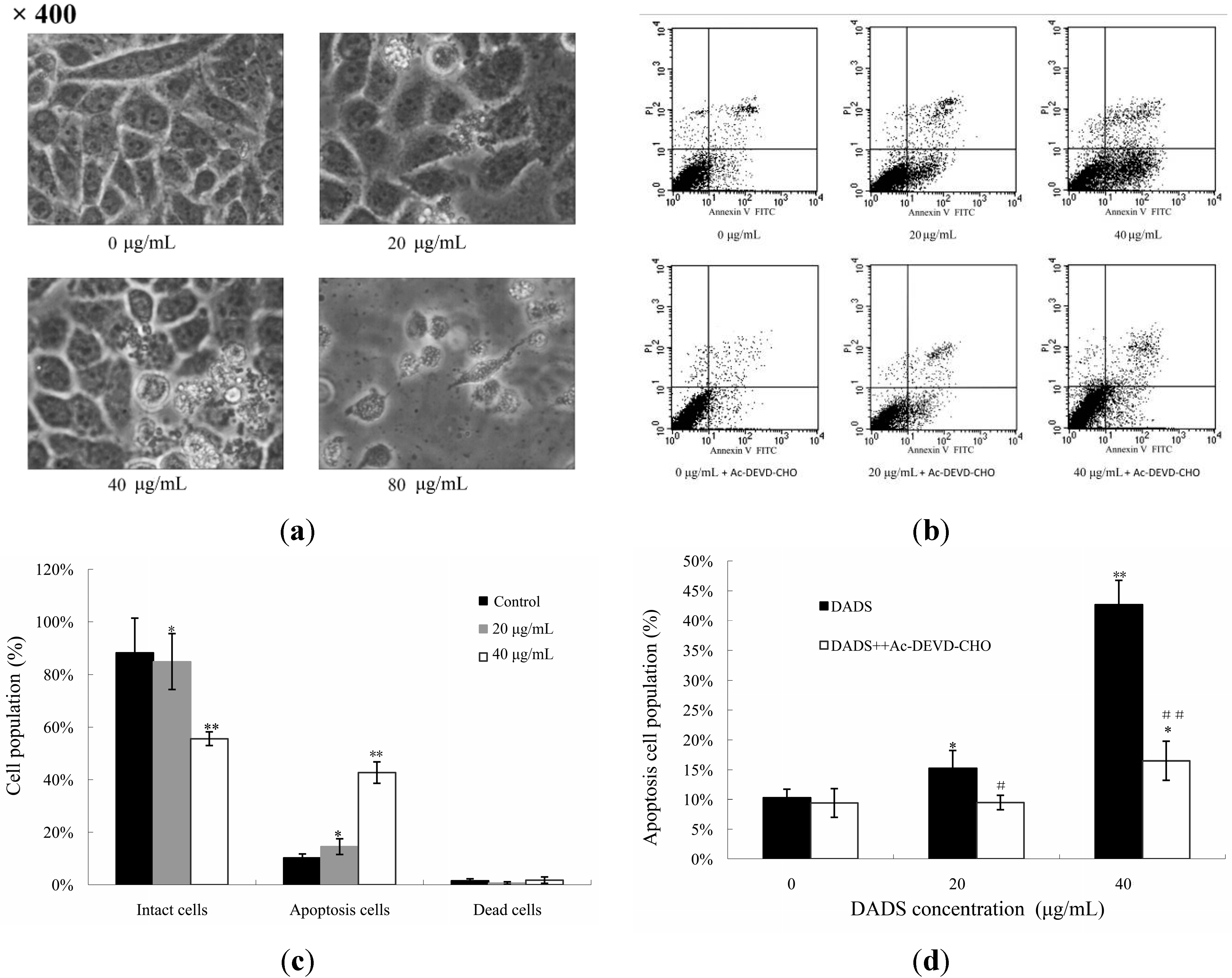
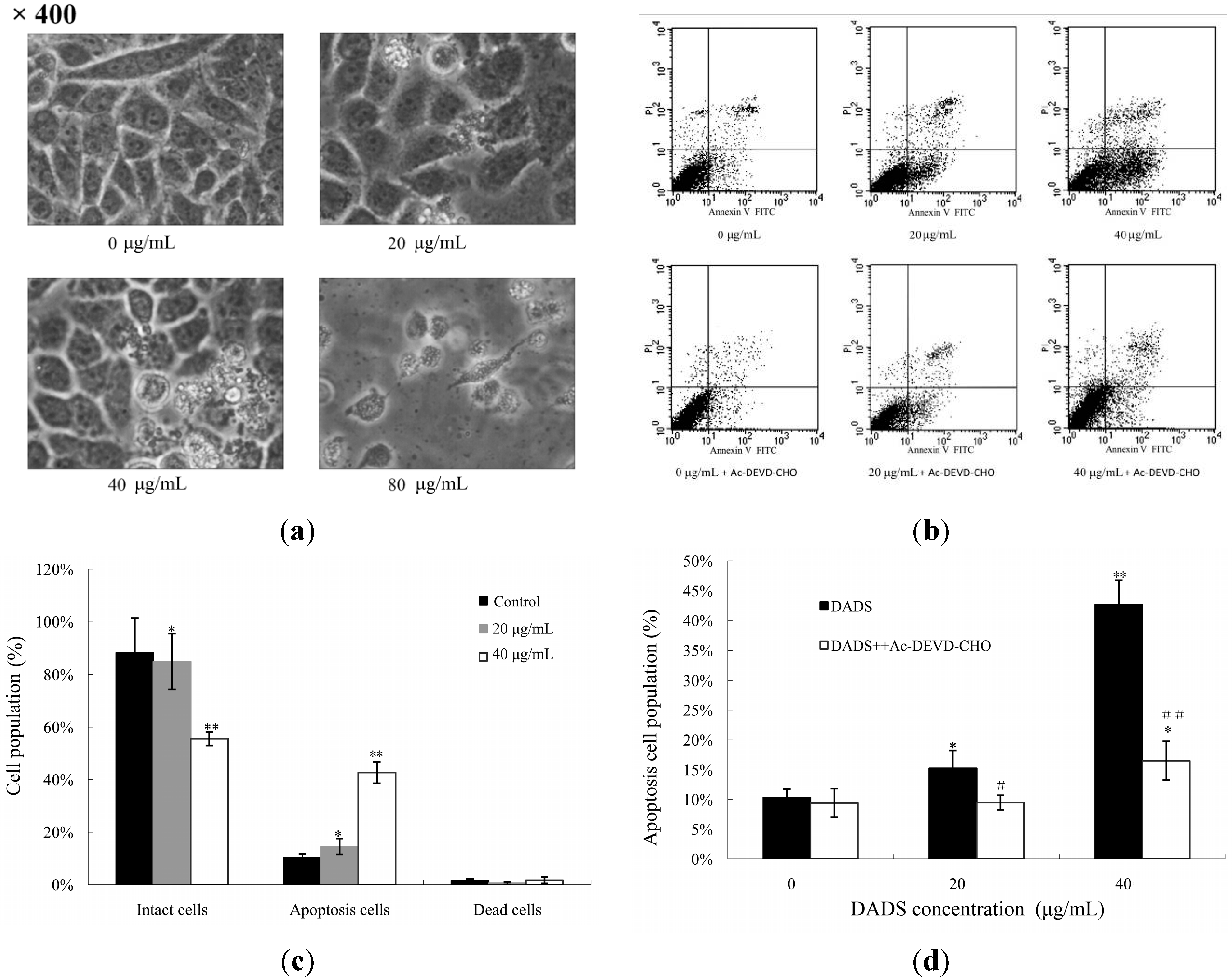
2.2. DADS-Induced Apoptosis
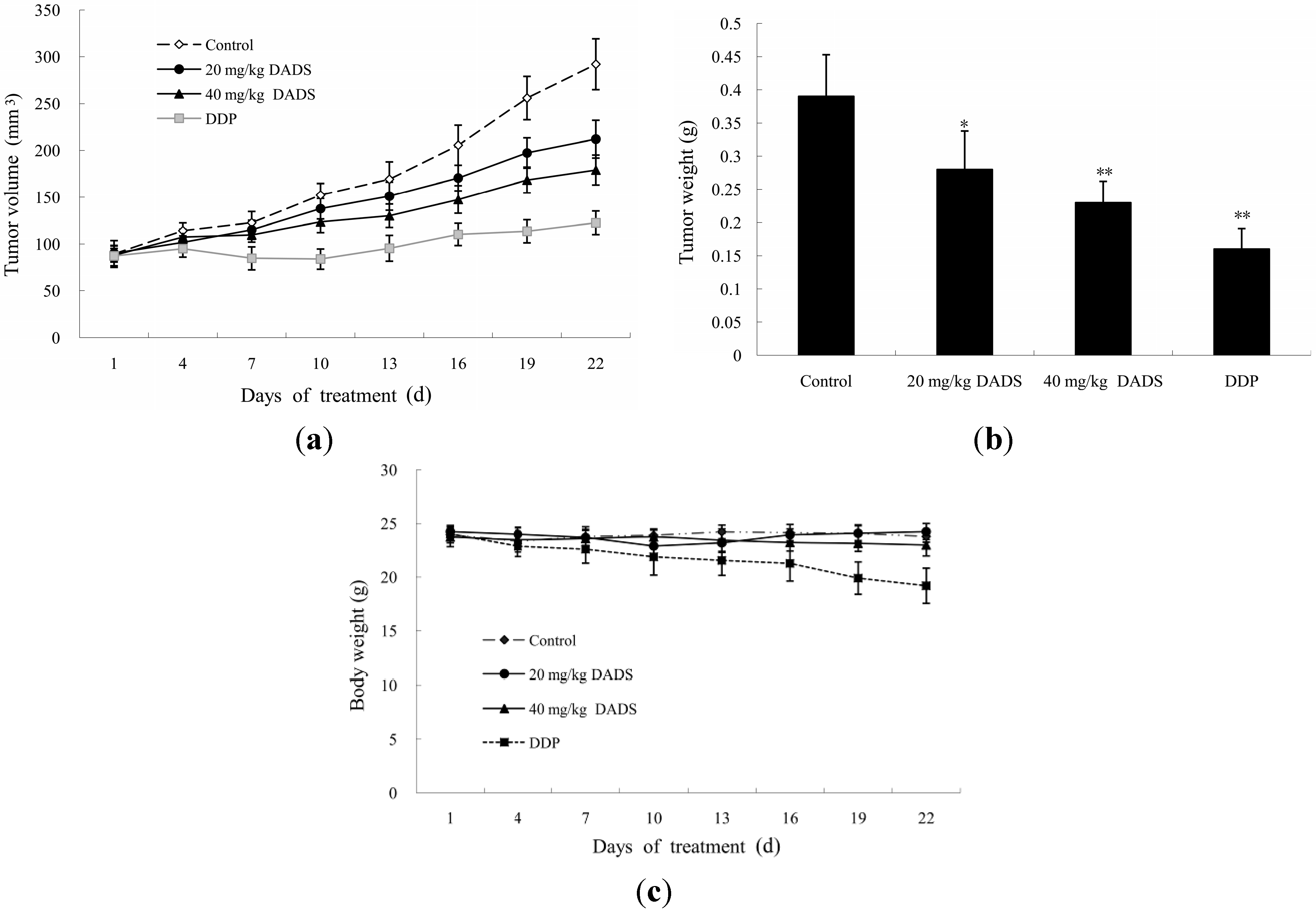
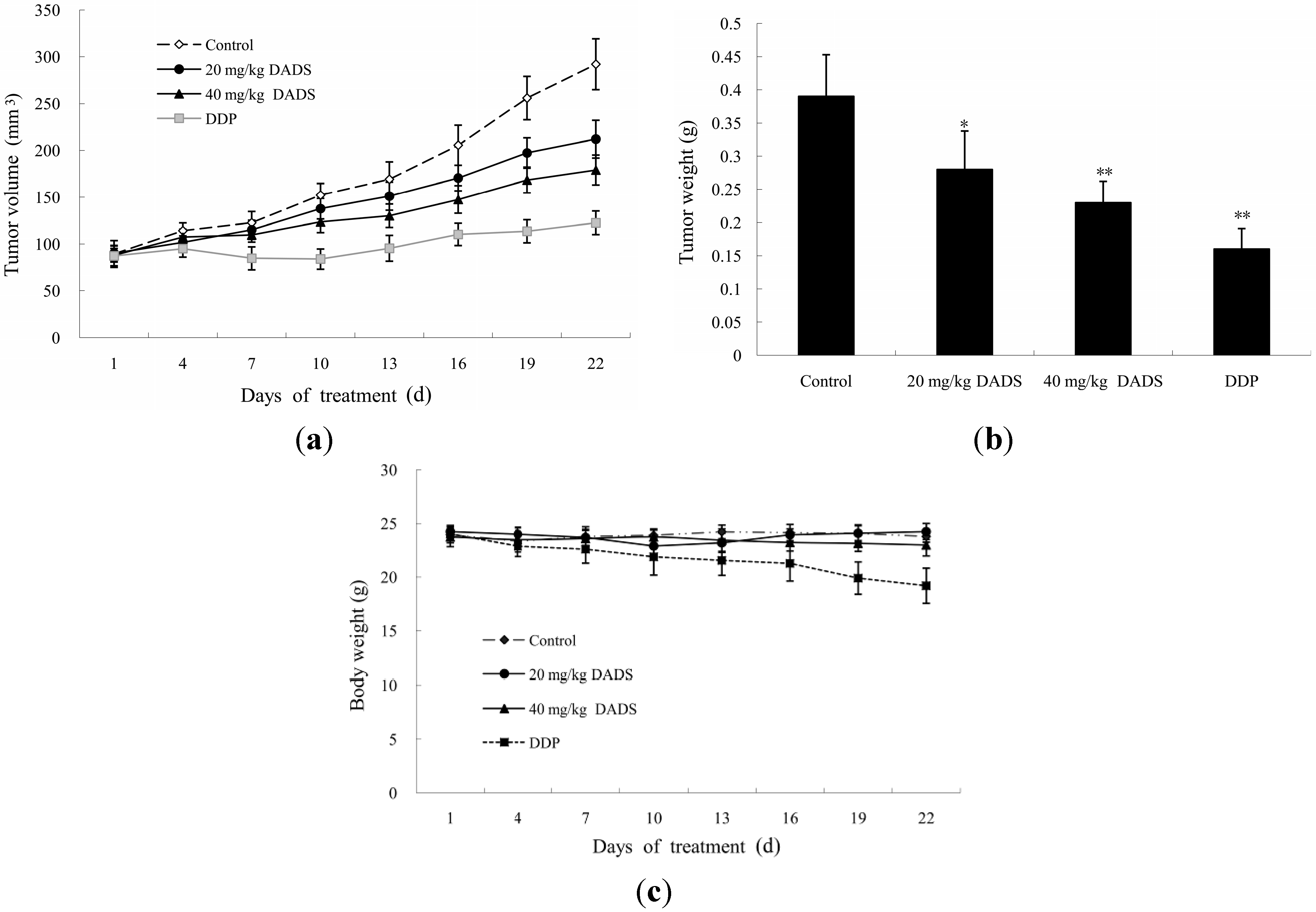
2.3. DADS Blocked the Growth of Xenograft Tumor
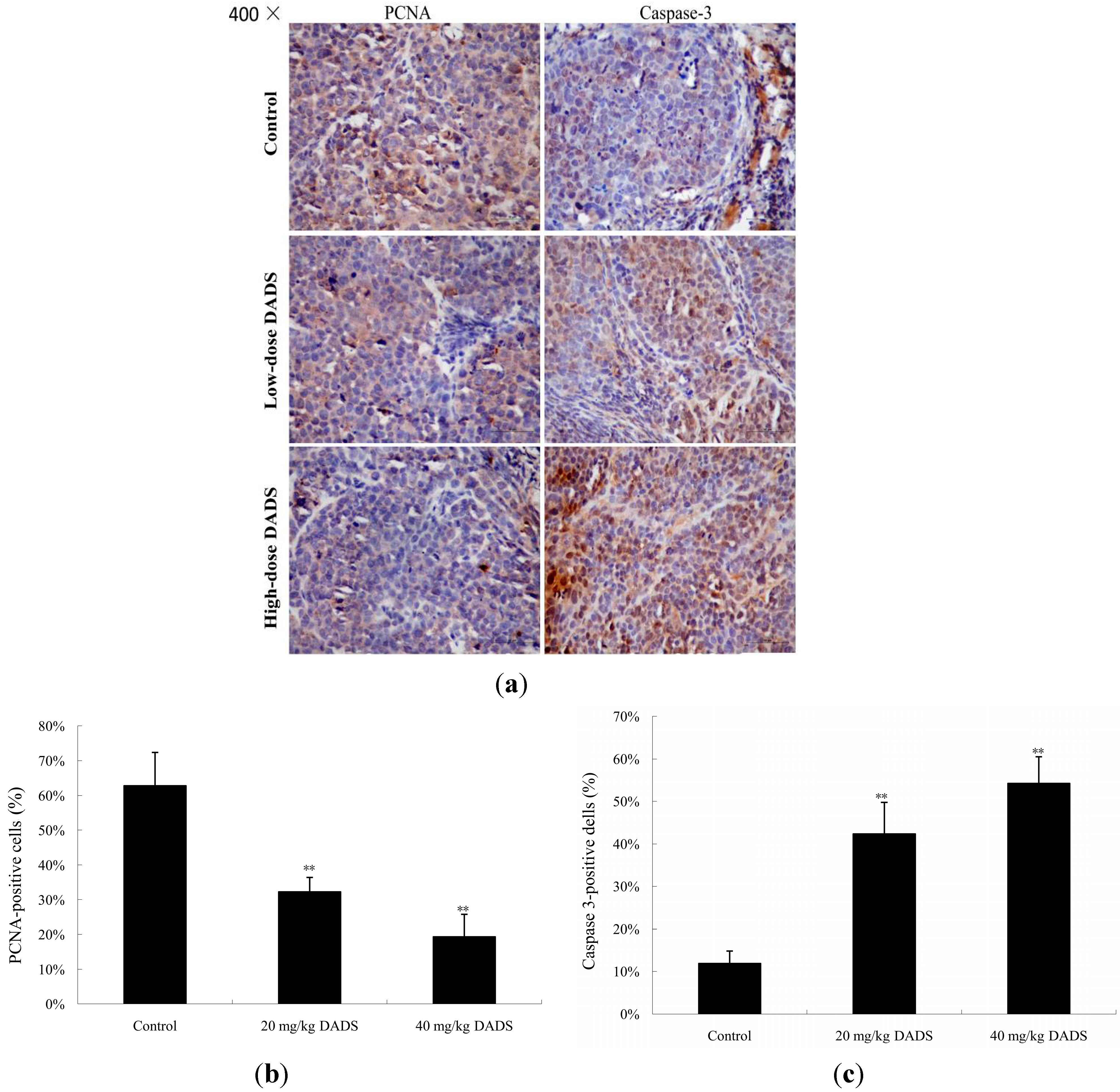
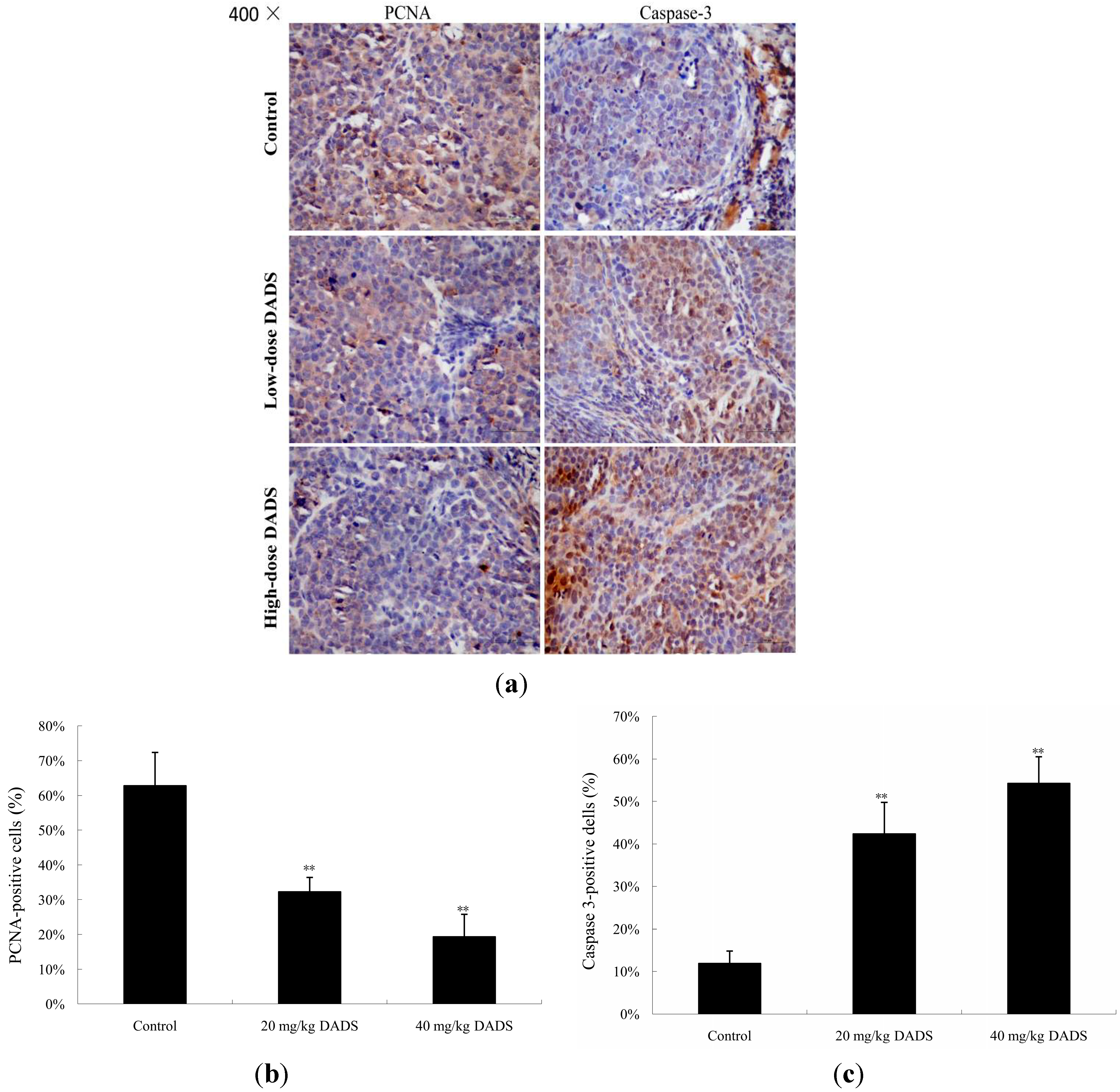
2.4. DADS Inhibited Cell Proliferation and Induced Apoptosis in Xenograft Tumor
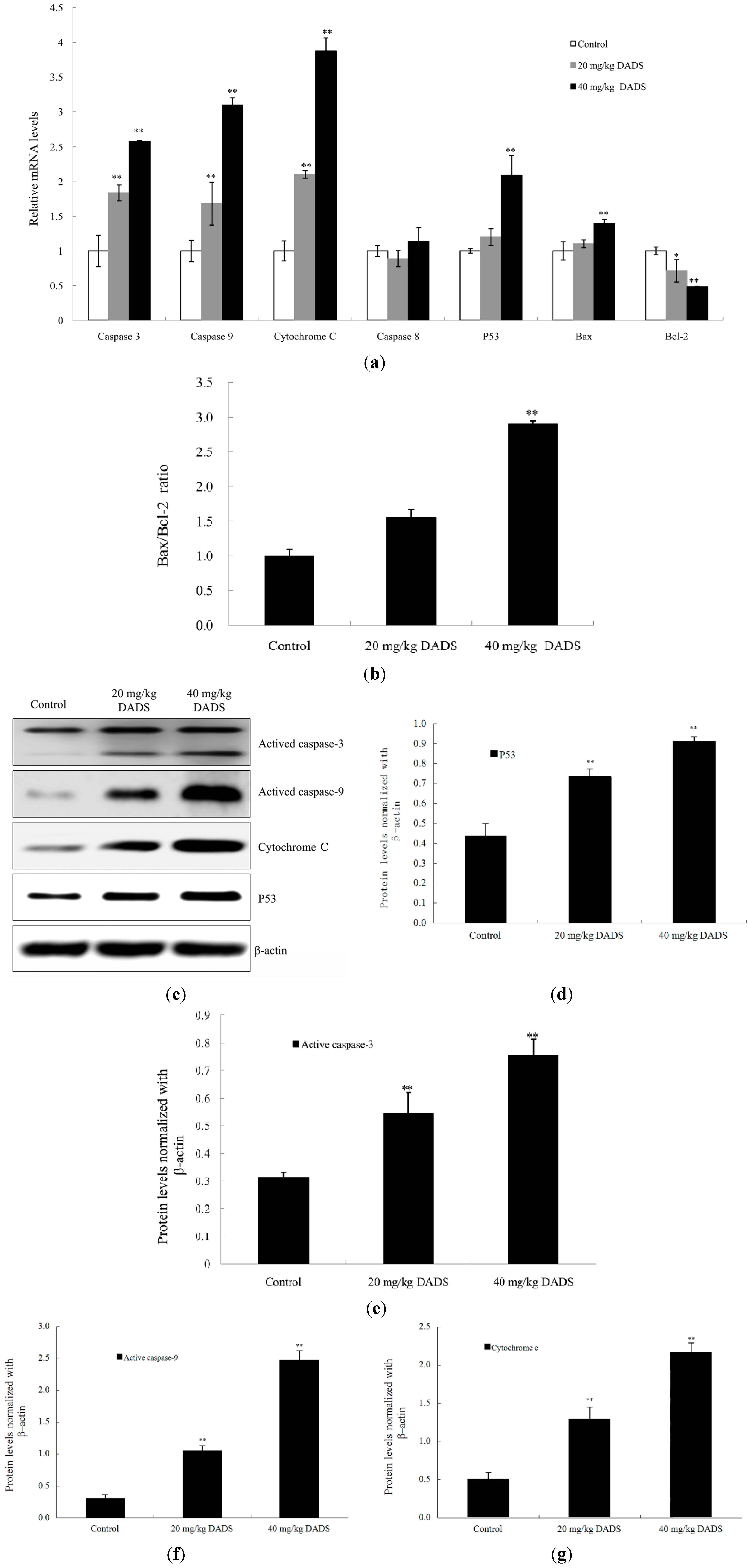
2.5. DADS Activated Mitochondria-Dependent Pathway and Up-Regulated Bax/Bcl-2 Ratio in Xenograft Tumors
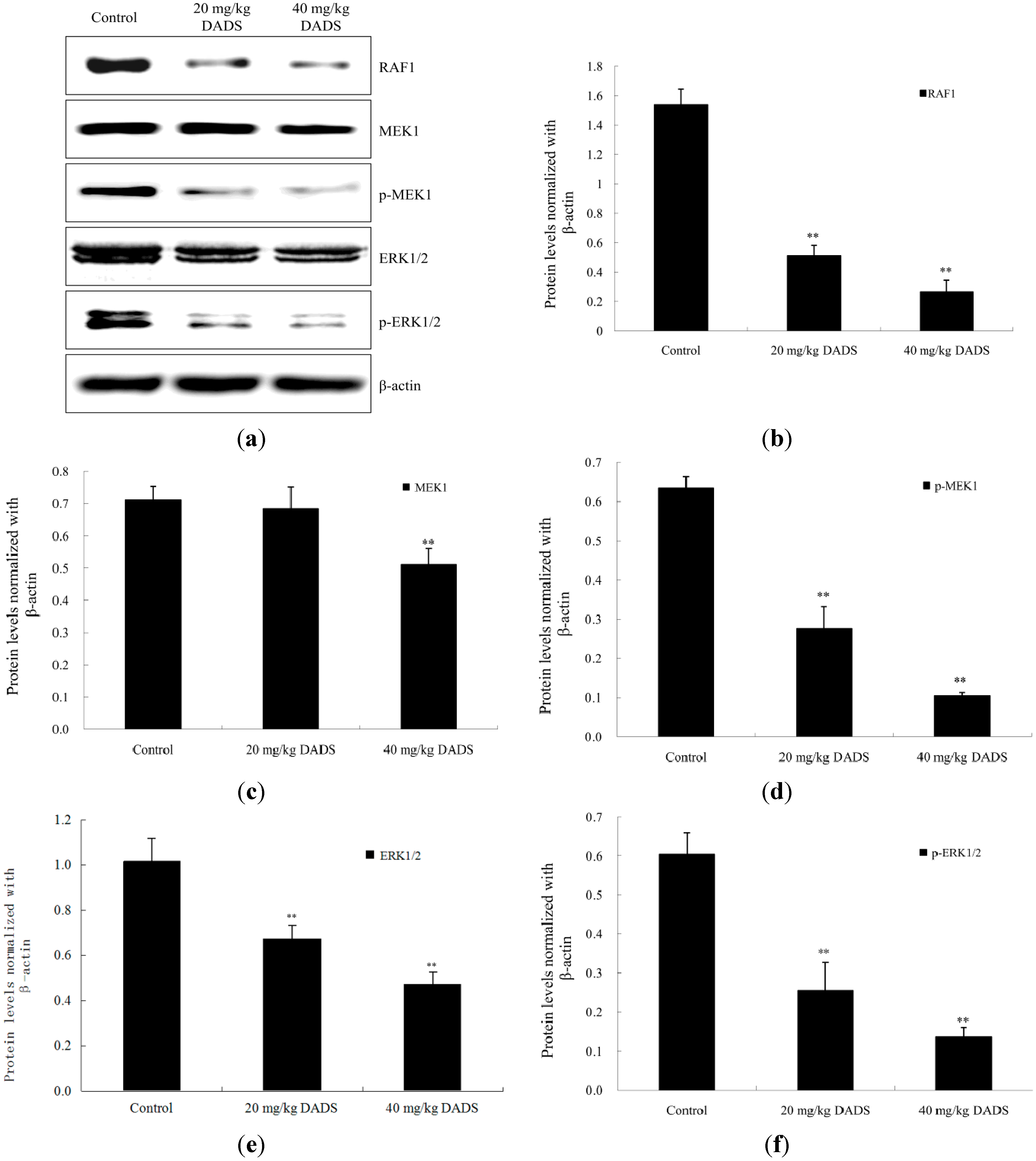
2.6. DADS Alters the RAF/MEK/ERK Pathway in Xenograft Tumors
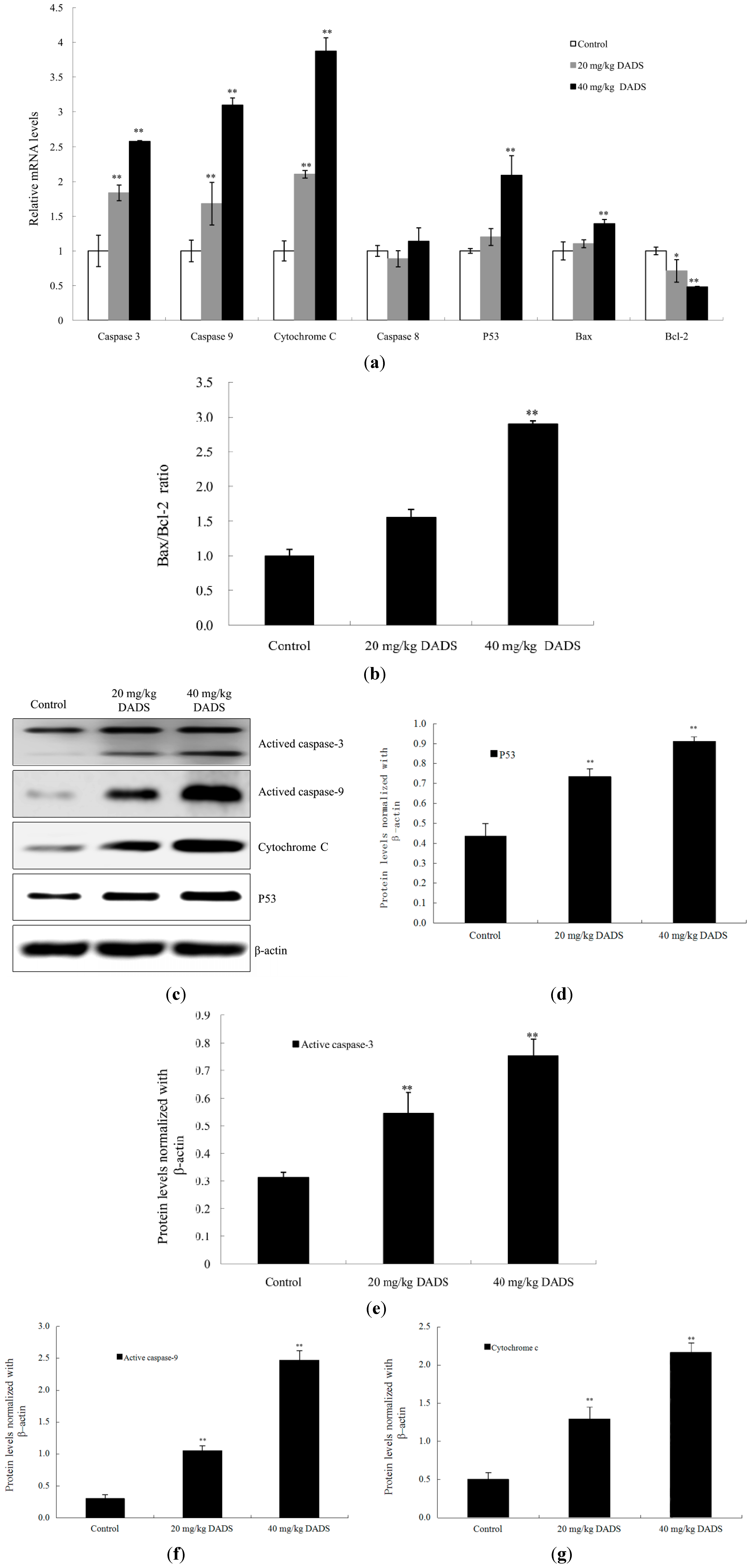

2.7. Discussion
3. Experimental Section
3.1. Reagents and Antibodies
3.2. Cell Culture
3.3. Cell Viability Assay
3.4. Apoptosis Assay
3.5. Xenograft Tumor Assay in Vivo
3.6. Immunohistochemical Staining
3.7. Quantitative Real-Time PCR
3.8. Western Blot Assay
3.9. Quantification and Statistic Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Liu, R.; Peng, Y.; Li, X.; Wang, Y.; Pan, E.; Guo, W.; Pu, Y.; Yin, L. Identification of plasma metabolomic profiling for diagnosis of esophageal squamous-cell carcinoma using an UPLC/TOF/MS platform. Int. J. Mol. Sci. 2013, 14, 8899–8911. [Google Scholar]
- Song, Y.; Li, L.; Ou, Y.; Gao, Z.; Li, E.; Li, X.; Zhang, W.; Wang, J.; Xu, L.; Zhou, Y.; et al. Identification of genomic alterations in oesophageal squamous cell cancer. Nature 2014, 509, 91–95. [Google Scholar] [CrossRef]
- Cui, X.; Zhao, Z.; Liu, D.; Guo, T.; Li, S.; Hu, J.; Liu, C.; Yang, L.; Cao, Y.; Jiang, J.; et al. Inactivation of miR-34a by aberrant CpG methylation in Kazakh patients with esophageal carcinoma. J. Exp. Clin. Cancer Res. 2014, 33, 20. [Google Scholar] [CrossRef]
- Shen, Z.T.; Wu, X.H.; Li, B.; Shen, J.S.; Wang, Z.; Li, J.; Zhu, X.X. Nedaplatin concurrent with three-dimensional conformal radiotherapy for treatment of locally advanced esophageal carcinoma. World J. Gastroenterol. 2013, 19, 9447–9452. [Google Scholar] [CrossRef]
- Yeruva, L.; Elegbede, J.A.; Carper, S.W. Methyl jasmonate decreases membrane fluidity and induces apoptosis through tumor necrosis factor receptor 1 in breast cancer cells. Anti-Cancer Drugs 2008, 19, 766–776. [Google Scholar] [CrossRef]
- Eswar, K.; Venkateshbabu, N.; Rajeswari, K.; Kandaswamy, D. Dentinal tubule disinfection with 2% chlorhexidine, garlic extract, and calcium hydroxide against Enterococcus faecalis by using real-time polymerase chain reaction: In vitro study. J. Conserv. Dent. 2013, 16, 194–198. [Google Scholar]
- Truong, D.; Hindmarsh, W.; O’Brien, P.J. The molecular mechanisms of diallyl disulfide and diallyl sulfide induced hepatocyte cytotoxicity. Chem. Biol. Interact. 2009, 180, 79–88. [Google Scholar] [CrossRef]
- Lee, I.C.; Kim, S.H.; Baek, H.S.; Moon, C.; Kim, S.H.; Kim, Y.B.; Yun, W.K.; Kim, H.C.; Kim, J.C. Protective effects of diallyl disulfide on carbon tetrachloride-induced hepatotoxicity through activation of Nrf2. Environ. Toxicol. 2013. Available online: http://onlinelibrary.wiley.com/doi/10.1002/tox.21930/abstract;jsessionid=1C5EEE912870EFFD9CEB32D9B9CFC1D2.f02t04?deniedAccessCustomisedMessage=&userIsAuthenticated=false (accessed on 1 December 2013). [CrossRef]
- Alam, M.; Zubair, S.; Farazuddin, M.; Ahmad, E.; Khan, A.; Zia, Q.; Malik, A.; Mohammad, O. Development, characterization and efficacy of niosomal diallyl disulfide in treatment of disseminated murine candidiasis. Nanomed. Nanotechnol. Biol. Med. 2013, 9, 247–256. [Google Scholar] [CrossRef]
- Shin, D.Y.; Kim, G.Y.; Lee, J.H.; Choi, B.T.; Yoo, Y.H.; Choi, Y.H. Apoptosis induction of human prostate carcinoma DU145 cells by diallyl disulfide via modulation of JNK and PI3K/AKT signaling pathways. Int. J. Mol. Sci. 2012, 13, 14158–14171. [Google Scholar]
- Tang, H.; Kong, Y.; Guo, J.; Tang, Y.; Xie, X.; Yang, L.; Su, Q.; Xie, X. Diallyl disulfide suppresses proliferation and induces apoptosis in human gastric cancer through Wnt-1 signaling pathway by up-regulation of miR-200b and miR-22. Cancer Lett. 2013, 340, 72–81. [Google Scholar] [CrossRef]
- Myzak, M.C.; Dashwood, R.H. Histone deacetylases as targets for dietary cancer preventive agents: Lessons learned with butyrate, diallyl disulfide, and sulforaphane. Curr. Drug Targets 2006, 7, 443–452. [Google Scholar] [CrossRef]
- Jun, Z.; Suzuki, M.; Xiao, J.; Wen, J.; Talbot, S.G.; Li, G.C.; Xu, M. Comparative effects of natural and synthetic diallyl disulfide on apoptosis of human breast-cancer MCF-7 cells. Biotechnol. Appl. Biochem. 2009, 52, 113–119. [Google Scholar] [CrossRef]
- Lai, K.C.; Kuo, C.L.; Ho, H.C.; Yang, J.S.; Ma, C.Y.; Lu, H.F.; Huang, H.Y.; Chueh, F.S.; Yu, C.C.; Chung, J.G. Diallyl sulfide, diallyl disulfide and diallyl trisulfide affect drug resistant gene expression in colo 205 human colon cancer cells in vitro and in vivo. Phytomed. Int. J. Phytother. Phytopharmacol. 2012, 19, 625–630. [Google Scholar] [CrossRef]
- Hui, C.; Jun, W.; Ya, L.N.; Ming, X. Effect of Allium sativum (garlic) diallyl disulfide (DADS) on human non-small cell lung carcinoma H1299 cells. Trop. Biomed. 2008, 25, 37–45. [Google Scholar]
- Tan, H.; Ling, H.; He, J.; Yi, L.; Zhou, J.; Lin, M.; Su, Q. Inhibition of ERK and activation of p38 are involved in diallyl disulfide induced apoptosis of leukemia HL-60 cells. Arch. Pharm. Res. 2008, 31, 786–793. [Google Scholar] [CrossRef]
- Liao, Q.J.; Su, J.; Zhou, X.T.; Tang, H.L.; Song, Y.; Su, Q. Inhibitory effect of diallyl disulfide on proliferation of human colon cancer cell line SW480 in nude mice. Chin. J. Cancer 2007, 26, 828–832. (In Chinese) [Google Scholar]
- Xiang, S.L.; Xiao, X.L.; Ling, H.; Liao, Q.J.; Zhou, X.T.; Dong, L.; Su, Q. Antitumor effect of diallyl disulfide on human gastric cancer MGC803 cells xenograft in nude mice. Chin. J. Cancer 2005, 24, 940–944. (In Chinese) [Google Scholar]
- Zhao, J.; Huang, W.G.; He, J.; Tan, H.; Liao, Q.J.; Su, Q. Diallyl disulfide suppresses growth of HL-60 cell through increasing histone acetylation and p21WAF1 expression in vivo and in vitro. Acta Pharmacol. Sin. 2006, 27, 1459–1466. [Google Scholar] [CrossRef]
- Haneji, T.; Hirashima, K.; Teramachi, J.; Morimoto, H. Okadaic acid activates the PKR pathway and induces apoptosis through PKR stimulation in MG63 osteoblast-like cells. Int. J. Oncol. 2013, 42, 1904–1910. [Google Scholar]
- Ozoren, N.; El-Deiry, W.S. Cell surface death receptor signaling in normal and cancer cells. Semin. Cancer Biol. 2003, 13, 135–147. [Google Scholar] [CrossRef]
- Jin, Z.; El-Deiry, W.S. Overview of cell death signaling pathways. Cancer Biol. Ther. 2005, 4, 139–163. [Google Scholar] [CrossRef]
- Olsson, M.; Zhivotovsky, B. Caspases and cancer. Cell Death Differ. 2011, 18, 1441–1449. [Google Scholar] [CrossRef]
- Thomas, S.A.; Vasudevan, S.; Thamkachy, R.; Lekshmi, S.U.; Santhoshkumar, T.R.; Rajasekharan, K.N.; Sengupta, S. Upregulation of DR5 receptor by the diaminothiazole DAT1 [4-amino-5-benzoyl-2-(4-methoxy phenyl amino) thiazole] triggers an independent extrinsic pathway of apoptosis in colon cancer cells with compromised pro and antiapoptotic proteins. Apoptosis Int. J. Program. Cell Death 2013, 18, 713–726. [Google Scholar] [CrossRef]
- Zheng, T.S.; Hunot, S.; Kuida, K.; Momoi, T.; Srinivasan, A.; Nicholson, D.W.; Lazebnik, Y.; Flavell, R.A. Deficiency in caspase-9 or caspase-3 induces compensatory caspase activation. Nat. Med. 2000, 6, 1241–1247. [Google Scholar] [CrossRef]
- Yang, J.S.; Chen, G.W.; Hsia, T.C.; Ho, H.C.; Ho, C.C.; Lin, M.W.; Lin, S.S.; Yeh, R.D.; Ip, S.W.; Lu, H.F.; et al. Diallyl disulfide induces apoptosis in human colon cancer cell line (COLO 205) through the induction of reactive oxygen species, endoplasmic reticulum stress, caspases casade and mitochondrial-dependent pathways. Food Chem. Toxicol. 2009, 47, 171–179. [Google Scholar] [CrossRef]
- Nagaraj, N.S.; Anilakumar, K.R.; Singh, O.V. Diallyl disulfide causes caspase-dependent apoptosis in human cancer cells through a Bax-triggered mitochondrial pathway. J. Nutr. Biochem. 2010, 21, 405–412. [Google Scholar] [CrossRef]
- Altonsy, M.O.; Habib, T.N.; Andrews, S.C. Diallyl disulfide-induced apoptosis in a breast-cancer cell line (MCF-7) may be caused by inhibition of histone deacetylation. Nutr. Cancer 2012, 64, 1251–1260. [Google Scholar] [CrossRef]
- Gayathri, R.; Gunadharini, D.N.; Arunkumar, A.; Senthilkumar, K.; Krishnamoorthy, G.; Banudevi, S.; Vignesh, R.C.; Arunakaran, J. Effects of diallyl disulfide (DADS) on expression of apoptosis associated proteins in androgen independent human prostate cancer cells (PC-3). Mol. Cell. Biochem. 2009, 320, 197–203. [Google Scholar] [CrossRef]
- Sun, K.W.; Ma, Y.Y.; Guan, T.P.; Xia, Y.J.; Shao, C.M.; Chen, L.G.; Ren, Y.J.; Yao, H.B.; Yang, Q.; He, X.J. Oridonin induces apoptosis in gastric cancer through Apaf-1, cytochrome c and caspase-3 signaling pathway. World J. Gastroenterol. 2012, 18, 7166–7174. [Google Scholar] [CrossRef]
- Liu, J.; Qin, C.K.; Lv, W.; Zhao, Q.; Qin, C.Y. OSU-03012, a non-Cox inhibiting celecoxib derivative, induces apoptosis of human esophageal carcinoma cells through a p53/Bax/cytochrome c/caspase-9-dependent pathway. Anti-Cancer Drugs 2013, 24, 690–698. [Google Scholar] [CrossRef]
- Produit-Zengaffinen, N.; Pournaras, C.J.; Schorderet, D.F. Retinal ischemia-induced apoptosis is associated with alteration in Bax and Bcl-x(L) expression rather than modifications in Bak and Bcl-2. Mol. Vis. 2009, 15, 2101–2110. [Google Scholar]
- Pratheeshkumar, P.; Thejass, P.; Kutan, G. Diallyl disulfide induces caspase-dependent apoptosis via mitochondria-mediated intrinsic pathway in B16F-10 melanoma cells by up-regulating p53, caspase-3 and down-regulating pro-inflammatory cytokines and nuclear factor-kappabeta-mediated Bcl-2 activation. J. Environ. Pathol. Toxicol. Oncol. 2010, 29, 113–125. [Google Scholar] [CrossRef]
- Lin, Y.T.; Yang, J.S.; Lin, S.Y.; Tan, T.W.; Ho, C.C.; Hsia, T.C.; Chiu, T.H.; Yu, C.S.; Lu, H.F.; Weng, Y.S.; et al. Diallyl disulfide (DADS) induces apoptosis in human cervical cancer Ca Ski cells via reactive oxygen species and Ca2+-dependent mitochondria-dependent pathway. Anticancer Res. 2008, 28, 2791–2799. [Google Scholar]
- Masuelli, L.; Marzocchella, L.; Focaccetti, C.; Tresoldi, I.; Palumbo, C.; Izzi, V.; Benvenuto, M.; Fantini, M.; Lista, F.; Tarantino, U.; et al. Resveratrol and diallyl disulfide enhance curcumin-induced sarcoma cell apoptosis. Front. Biosci. 2012, 17, 498–508. [Google Scholar]
- Liao, Q.J.; Su, J.; He, J.; Song, Y.; Tang, H.L.; Su, Q. Effect of diallyl disulfide on cell cycle arrest of human colon cancer SW480 cells. Chin. J. Cancer 2009, 28, 138–141. (In Chinese) [Google Scholar]
- Wang, T.T.; Wang, S.K.; Huang, G.L.; Sun, G.J. Luteolin induced-growth inhibition and apoptosis of human esophageal squamous carcinoma cell line Eca109 cells in vitro. Asian Pac. J. Cancer Prev. 2012, 13, 5455–5461. [Google Scholar] [CrossRef]
- Ciccarelli, A.; Giustetto, M. Role of ERK signaling in activity-dependent modifications of histone proteins. Neuropharmacology 2014, 80C, 34–44. [Google Scholar] [CrossRef]
- Bhalla, S.; Evens, A.M.; Dai, B.; Prachand, S.; Gordon, L.I.; Gartenhaus, R.B. The novel anti-MEK small molecule AZD6244 induces BIM-dependent and AKT-independent apoptosis in diffuse large B-cell lymphoma. Blood 2011, 118, 1052–1061. [Google Scholar] [CrossRef]
- Lv, C.; Sun, W.; Sun, H.; Wei, S.; Chen, R.; Wang, B.; Huang, C. Asperolide A, a marine-derived tetranorditerpenoid, induces G2/M arrest in human NCI-H460 lung carcinoma cells, is mediated by p53–p21 stabilization and modulated by Ras/Raf/MEK/ERK signaling pathway. Mar. Drugs 2013, 11, 316–331. [Google Scholar] [CrossRef]
- Lai, K.C.; Hsu, S.C.; Kuo, C.L.; Yang, J.S.; Ma, C.Y.; Lu, H.F.; Tang, N.Y.; Hsia, T.C.; Ho, H.C.; Chung, J.G. Diallyl sulfide, diallyl disulfide, and diallyl trisulfide inhibit migration and invasion in human colon cancer colo 205 cells through the inhibition of matrix metalloproteinase-2, -7, and -9 expressions. Environ. Toxicol. 2013, 28, 479–488. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Wen, J.; Xiao, J.B.; Talbot, S.G.; Li, G.C.; Xu, M. Induction of apoptosis and transient increase of phosphorylated MAPKs by diallyl disulfide treatment in human nasopharyngeal carcinoma CNE2 cells. Arch. Pharm. Res. 2006, 29, 1125–1131. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yin, X.; Zhang, J.; Li, X.; Liu, D.; Feng, C.; Liang, R.; Zhuang, K.; Cai, C.; Xue, X.; Jing, F.; et al. DADS Suppresses Human Esophageal Xenograft Tumors through RAF/MEK/ERK and Mitochondria-Dependent Pathways. Int. J. Mol. Sci. 2014, 15, 12422-12441. https://doi.org/10.3390/ijms150712422
Yin X, Zhang J, Li X, Liu D, Feng C, Liang R, Zhuang K, Cai C, Xue X, Jing F, et al. DADS Suppresses Human Esophageal Xenograft Tumors through RAF/MEK/ERK and Mitochondria-Dependent Pathways. International Journal of Molecular Sciences. 2014; 15(7):12422-12441. https://doi.org/10.3390/ijms150712422
Chicago/Turabian StyleYin, Xiaoran, Jun Zhang, Xiaoning Li, Dong Liu, Cheng Feng, Rongrui Liang, Kun Zhuang, Chenlei Cai, Xinghuan Xue, Fuchun Jing, and et al. 2014. "DADS Suppresses Human Esophageal Xenograft Tumors through RAF/MEK/ERK and Mitochondria-Dependent Pathways" International Journal of Molecular Sciences 15, no. 7: 12422-12441. https://doi.org/10.3390/ijms150712422