Therapeutic Effect of Bone Marrow Mesenchymal Stem Cells on Laser-Induced Retinal Injury in Mice

Abstract
:1. Introduction
2. Results
2.1. Characterization of Green Fluorescent Protein Labeled Marrow Mesenchymal Stem Cells
2.2. GFP-MSC Migration
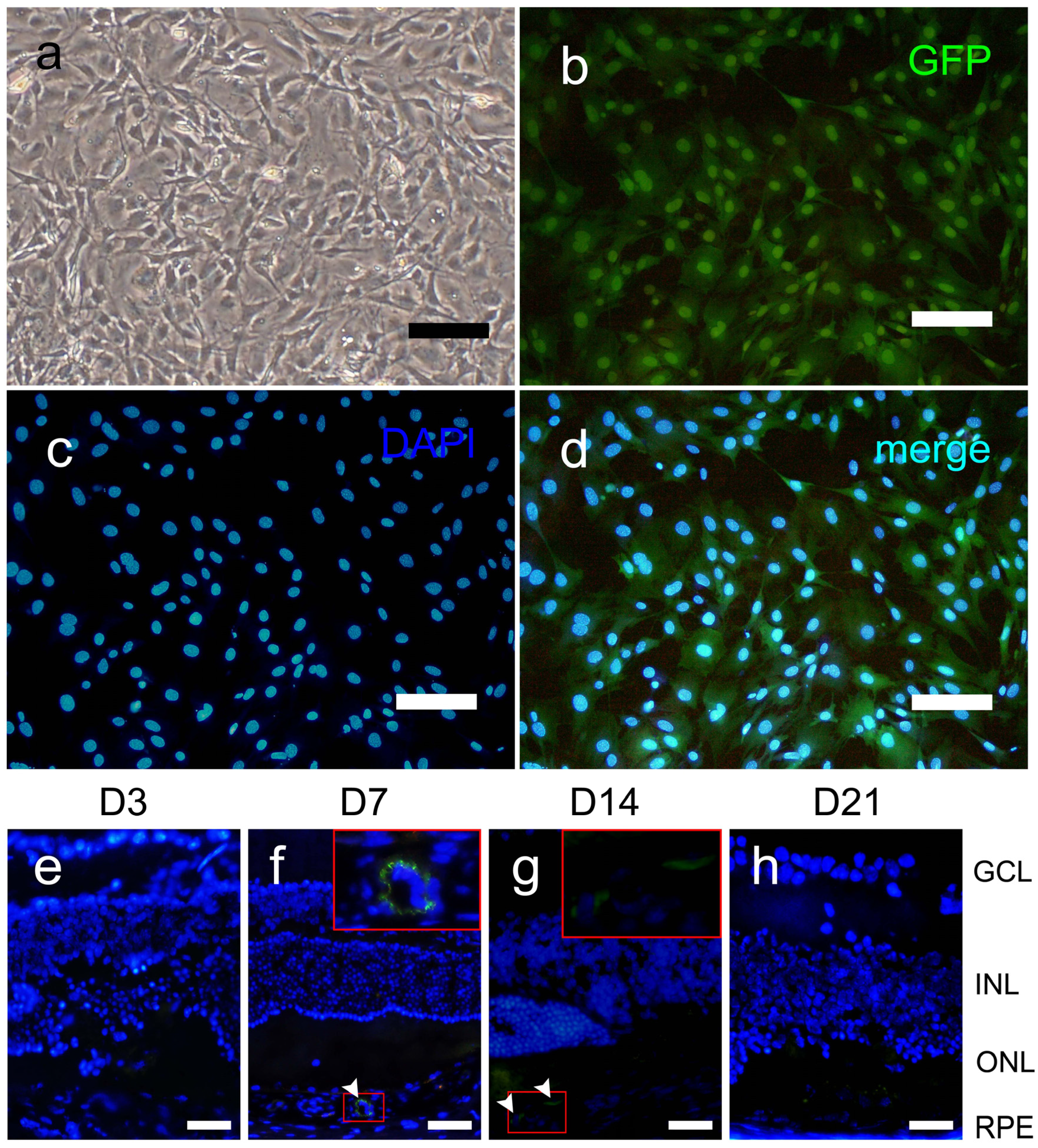
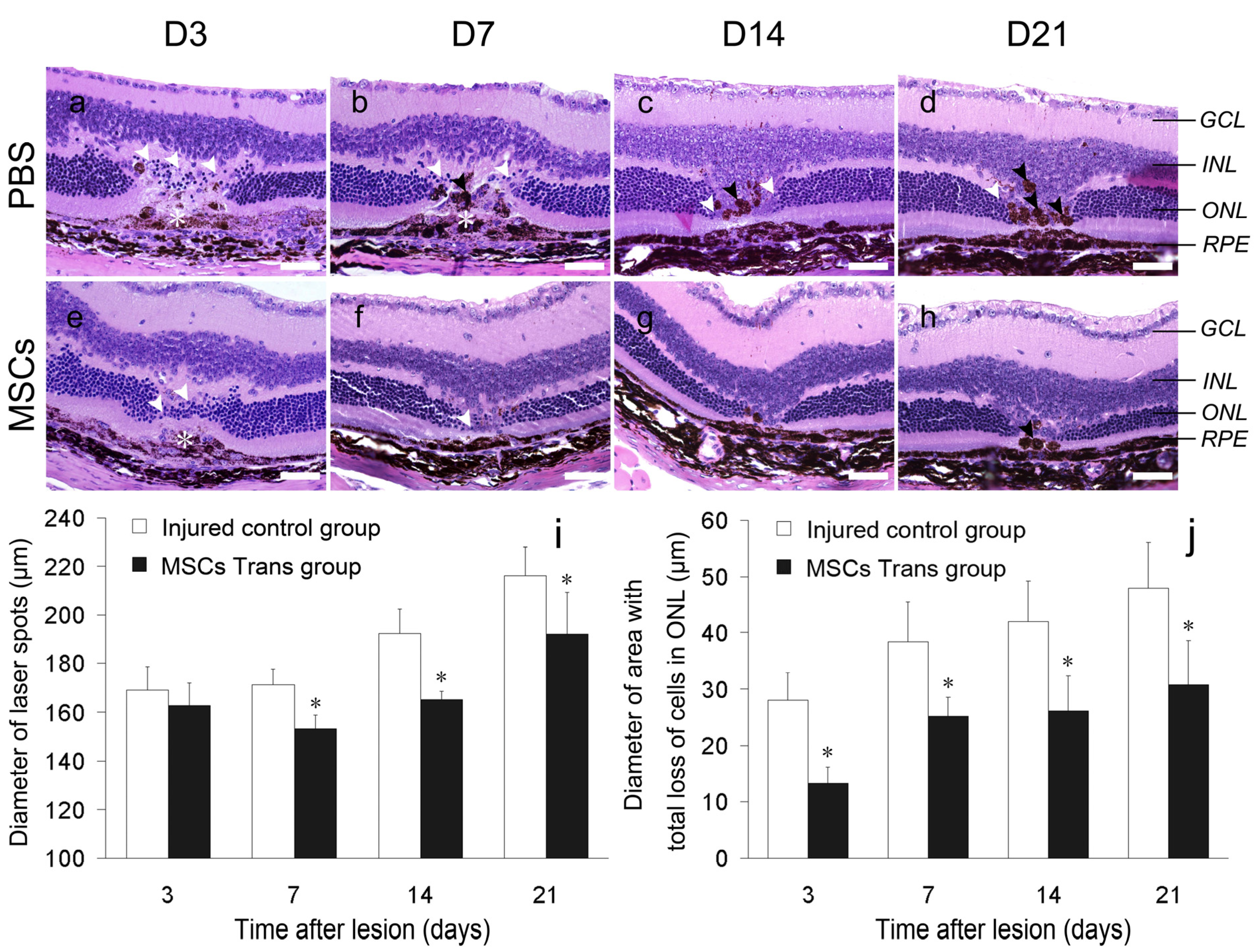
2.3. Histopathological Changes in the Retina after Laser Injury
2.4. Apoptosis of Retinal Cells
2.5. GFAP and MMP-2 mRNA Expression


3. Discussion
4. Experimental Section
4.1. Animals
4.2. Study Design
4.3. Laser-Induced Retinal Injury Model
4.4. GFP Labeled MSCs
4.5. Evaluation of GFP Expression in MSCs
4.6. Tracing GFP-Labeled MSCs
4.7. Histopathological Analysis of the Laser-Injured Retina
4.8. Apoptosis Analysis of the Laser-Injured Retina
4.9. Real-Time Quantitative RT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| GFAP | TGGAGGTGGAGAGGGACAAC | TGGTTTCATCTTGGAGCTTCTG |
| MMP-2 | GAGGACTATGACCGGGATAAGAAGT | GGGCACCTTCTGAATTTCCA |
| GAPDH | TGTGTCCGTCGTGGATCTGA | CCTGCTTCACCACCTTCTTGA |
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Abbreviations
Conflicts of Interest
References
- Cottet, S.; Schorderet, D.F. Mechanisms of apoptosis in retinitis pigmentosa. Curr. Mol. Med. 2009, 9, 375–383. [Google Scholar] [CrossRef]
- Eckmiller, M.S. Defective cone photoreceptor cytoskeleton, alignment, feedback, and energetics can lead to energy depletion in macular degeneration. Prog. Retin. Eye Res. 2004, 23, 495–522. [Google Scholar] [CrossRef]
- Nickells, R.W. Apoptosis of retinal ganglion cells in glaucoma: An update of the molecular pathways involved in cell death. Surv. Ophthalmol. 1999, 43, S151–S161. [Google Scholar] [CrossRef]
- Gregory, C.Y.; Bird, A.C. Cell loss in retinal dystrophies by apoptosis-death by informed consent! Br. J. Ophthalmol. 1995, 79, 186–190. [Google Scholar] [CrossRef]
- Maltman, D.J.; Hardy, S.A.; Przyborski, S.A. Role of mesenchymal stem cells in neurogenesis and nervous system repair. Neurochem. Int. 2011, 59, 347–356. [Google Scholar]
- Joyce, N.; Annett, G.; Wirthlin, L.; Olson, S.; Bauer, G.; Nolta, J.A. Mesenchymal stem cells for the treatment of neurodegenerative disease. Regen. Med. 2010, 5, 933–946. [Google Scholar] [CrossRef]
- Parr, A.M.; Tator, C.H.; Keating, A. Bone marrow-derived mesenchymal stromal cells for the repair of central nervous system injury. Bone Marrow Transplant. 2007, 40, 609–619. [Google Scholar] [CrossRef]
- Karussis, D.; Kassis, I.; Kurkalli, B.G.; Slavin, S. Immunomodulation and neuroprotection with mesenchymal bone marrow stem cells (MSCs): A proposed treatment for multiple sclerosis and other neuroimmunological/neurodegenerative diseases. J. Neurol. Sci. 2008, 265, 131–135. [Google Scholar] [CrossRef]
- Torrente, Y.; Polli, E. Mesenchymal stem cell transplantation for neurodegenerative diseases. Cell Transplant. 2008, 17, 1103–1113. [Google Scholar] [CrossRef]
- Johnson, T.V.; Bull, N.D.; Hunt, D.P.; Marina, N.; Tomarev, S.I.; Martin, K.R. Neuroprotective effects of intravitreal mesenchymal stem cell transplantation in experimental glaucoma. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2051–2059. [Google Scholar] [CrossRef]
- Inoue, Y.; Iriyama, A.; Ueno, S.; Takahashi, H.; Kondo, M.; Tamaki, Y.; Araie, M.; Yanagi, Y. Subretinal transplantation of bone marrow mesenchymal stem cells delays retinal degeneration in the RCS rat model of retinal degeneration. Exp. Eye Res. 2007, 85, 234–241. [Google Scholar] [CrossRef]
- Chung, J.K.; Park, T.K.; Ohn, Y.H.; Park, S.K.; Hong, D.S. Modulation of retinal wound healing by systemically administered bone marrow-derived mesenchymal stem cells. Korean J. Ophthalmol. 2011, 25, 268–274. [Google Scholar] [CrossRef]
- Belokopytov, M.; Belkin, M.; Dubinsky, G.; Epstein, Y.; Rosner, M. Development and recovery of laser-induced retinal lesion in rats. Retina 2010, 30, 662–670. [Google Scholar] [CrossRef]
- Schulmeister, K.; Stuck, B.E.; Lund, D.J.; Sliney, D.H. Review of thresholds and recommendations for revised exposure limits for laser and optical radiation for thermally induced retinal injury. Health Phys. 2011, 100, 210–220. [Google Scholar] [CrossRef]
- Kasaoka, M.; Ma, J.; Lashkari, K. c-Met modulates RPE migratory response to laser-induced retinal injury. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Müther, P.S.; Semkova, I.; Schmidt, K.; Abari, E.; Kuebbeler, M.; Beyer, M.; Abken, H.; Meyer, K.L.; Kociok, N.; Joussen, A.M. Conditions of retinal glial and inflammatory cell activation after irradiation in a GFP-chimeric mouse model. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4831–4839. [Google Scholar] [CrossRef]
- Tso, M.O.; Fine, B.S. Repair and late degeneration of the primate foveola after injury by argon laser. Investig. Ophthalmol. Vis. Sci. 1979, 18, 447–461. [Google Scholar]
- Wang, H.C.; Brown, J.; Alayon, H.; Stuck, B.E. Transplantation of quantum dot-labelled bone marrow-derived stem cells into the vitreous of mice with laser-induced retinal injury: Survival, integration and differentiation. Vision Res. 2010, 50, 665–673. [Google Scholar] [CrossRef]
- Castanheira, P.; Torquetti, L.; Nehemy, M.B.; Goes, A.M. Retinal incorporation and differentiation of mesenchymal stem cells intravitreally injected in the injured retina of rats. Arq. Bras. Oftalmol. 2008, 71, 644–650. [Google Scholar] [CrossRef]
- Castanheira, P.; Torquetti, L.T.; Magalhas, D.R.; Nehemy, M.B.; Goes, A.M. DAPI diffusion after intravitreal injection of mesenchymal stem cells in the injured retina of rats. Cell Transplant. 2009, 18, 423–431. [Google Scholar] [CrossRef]
- Sasaki, M.; Shinoda, H.; Koto, T.; Uchida, A.; Tsubota, K.; Ozawa, Y. Use of micronutrient supplement for preventing advanced age-related macular degeneration in Japan. Arch. Ophthalmol. 2012, 130, 254–255. [Google Scholar] [CrossRef]
- Girmens, J.F.; Picaud, S.; Sahel, J.A. Therapeutic innovation in AMD and other retinal diseases. Rev. Prat. 2013, 63, 68–73. (In French) [Google Scholar]
- Thumann, G. Prospectives for gene therapy of retinal degenerations. Curr. Genomics 2012, 13, 350–362. [Google Scholar] [CrossRef]
- Fernandes, R.A.; Diniz, B.; Ribeiro, R.; Humayun, M. Artificial vision through neuronal stimulation. Neurosci. Lett. 2012, 519, 122–128. [Google Scholar] [CrossRef]
- Turlea, C. Future methods of treatment in age related macular degeneration. Oftalmologia 2012, 56, 30–35. (In Romanian) [Google Scholar]
- Stern, J.H.; Temple, S. Stem cells for retinal replacement therapy. Neurotherapeutics 2011, 8, 736–743. [Google Scholar]
- Joe, A.W.; Gregory-Evans, K. Mesenchymal stem cells and potential applications in treating ocular disease. Curr. Eye Res. 2010, 35, 941–952. [Google Scholar] [CrossRef]
- Tibbetts, M.D.; Samuel, M.A.; Chang, T.S.; Ho, A.C. Stem cell therapy for retinal disease. Curr. Opin. Ophthalmol. 2012, 23, 226–234. [Google Scholar] [CrossRef]
- Zhou, X.; Xia, X.B.; Xiong, S.Q. Neuro-protection of retinal stem cells transplantation combined with copolymer-1 immunization in a rat model of glaucoma. Mol. Cell Neurosci. 2013, 54, 1–8. [Google Scholar] [CrossRef]
- Johnson, T.V.; Martin, K.R. Cell transplantation approaches to retinal ganglion cell neuroprotection in glaucoma. Curr. Opin. Pharmacol. 2013, 13, 78–82. [Google Scholar] [CrossRef]
- Brancato, R.; Pece, A.; Avanza, P.; Radrizzani, E. Photocoagulation scar expansion after laser therapy for choroidal neovascularization in degenerative myopia. Retina 1990, 10, 239–243. [Google Scholar]
- Dastgheib, K.; Bressler, S.B.; Green, W.R. Clinicopathologic correlation of laser lesion expansion after treatment of choroidal neovascularization. Retina 1993, 13, 345–352. [Google Scholar] [CrossRef]
- Ghazi-Nouri, S.M.; Assi, A.; Limb, G.A.; Scott, R.A.; von Bussmann, K.; Humphrey, I.; Luthert, P.J.; Charteris, D.G. Laser photocoagulation alters the pattern of staining for neurotrophin-4, GFAP, and CD68 in human retina. Br. J. Ophthalmol. 2003, 87, 488–492. [Google Scholar] [CrossRef]
- Wong, T.T.; Sethi, C.; Daniels, J.T.; Limb, G.A.; Murphy, G.; Khaw, P.T. Matrix metalloproteinases in disease and repair processes in the anterior segment. Surv. Ophthalmol. 2002, 47, 239–256. [Google Scholar] [CrossRef]
- Vaughan-Thomas, A.; Gilbert, S.J.; Duance, V.C. Elevated levels of proteolytic enzymes in the aging human vitreous. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3299–3304. [Google Scholar]
- Brown, D.; Hamdi, H.; Bahri, S.; Kenney, M.C. Characterization of an endogenous metalloproteinase in human vitreous. Curr. Eye Res. 1994, 13, 639–647. [Google Scholar] [CrossRef]
- Plantner, J.J.; Smine, A.; Quinn, T.A. Matrix metalloproteinases and metalloproteinase inhibitors in human interphotoreceptor matrix and vitreous. Curr. Eye Res. 1998, 17, 132–140. [Google Scholar] [CrossRef]
- Hofmaier, F.; Hauck, S.M.; Amann, B.; Degroote, R.L.; Deeg, C.A. Changes in matrix metalloproteinase network in a spontaneous autoimmune uveitis model. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2314–2320. [Google Scholar]
- El-Shabrawi, Y.G.; Christen, W.G.; Foster, S.C. Correlation of metalloproteinase-2 and -9 with proinflammatory cytokines interleukin-1b, interleukin-12 and the interleukin-1 receptor antagonist in patients with chronic uveitis. Curr. Eye Res. 2000, 20, 211–214. [Google Scholar] [CrossRef]
- Ahuja, S.; Ahuja, P.; Caffe, A.R.; Ekstrom, P.; Abrahamson, M.; van Veen, T. RD1 mouse retina shows imbalance in cellular distribution and levels of TIMP-1/MMP-9, TIMP-2/MMP-2 and sulfated glycosaminoglycans. Ophthalmic Res. 2006, 38, 125–136. [Google Scholar] [CrossRef]
- Mohammad, G.; Kowluru, R.A. Matrix metalloproteinase-2 in the development of diabetic retinopathy and mitochondrial dysfunction. Lab. Investig. 2010, 90, 1365–1372. [Google Scholar] [CrossRef]
- Cao, L.; Wang, H.; Wang, F. Amyloid-beta-induced matrix metalloproteinase-9 secretion is associated with retinal pigment epithelial barrier disruption. Int. J. Mol. Med. 2013, 31, 1105–1112. [Google Scholar]
- Tasso, R.; Ilengo, C.; Quarto, R.; Cancedda, R.; Caspi, R.R.; Pennesi, G. Mesenchymal stem cells induce functionally active T-regulatory lymphocytes in a paracrine fashion and ameliorate experimental autoimmune uveitis. Investig. Ophthalmol. Vis. Sci. 2012, 53, 786–793. [Google Scholar] [CrossRef]
- Kimbrel, E.A.; Kouris, N.A.; Yavanian, G.; Chu, J.; Qin, Y.; Chan, A.; Singh, R.P.; McCurdy, D.; Gordon, L.; Levinson, R.D.; et al. Mesenchymal stem cell population derived from human pluripotent stem cells displays potent immunomodulatory and therapeutic properties. Stem Cells Dev. 2014, in press. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jiang, Y.; Zhang, Y.; Zhang, L.; Wang, M.; Zhang, X.; Li, X. Therapeutic Effect of Bone Marrow Mesenchymal Stem Cells on Laser-Induced Retinal Injury in Mice. Int. J. Mol. Sci. 2014, 15, 9372-9385. https://doi.org/10.3390/ijms15069372
Jiang Y, Zhang Y, Zhang L, Wang M, Zhang X, Li X. Therapeutic Effect of Bone Marrow Mesenchymal Stem Cells on Laser-Induced Retinal Injury in Mice. International Journal of Molecular Sciences. 2014; 15(6):9372-9385. https://doi.org/10.3390/ijms15069372
Chicago/Turabian StyleJiang, Yuanfeng, Yan Zhang, Lingjun Zhang, Meiyan Wang, Xiaomin Zhang, and Xiaorong Li. 2014. "Therapeutic Effect of Bone Marrow Mesenchymal Stem Cells on Laser-Induced Retinal Injury in Mice" International Journal of Molecular Sciences 15, no. 6: 9372-9385. https://doi.org/10.3390/ijms15069372