Genetic Variations of TAP1 Gene Exon 3 Affects Gene Expression and Escherichia coli F18 Resistance in Piglets


Abstract
:1. Introduction
2. Results
2.1. Identification of the Escherichia coli F18-Resistant and E. coli F18-Susceptible Sutai Piglets
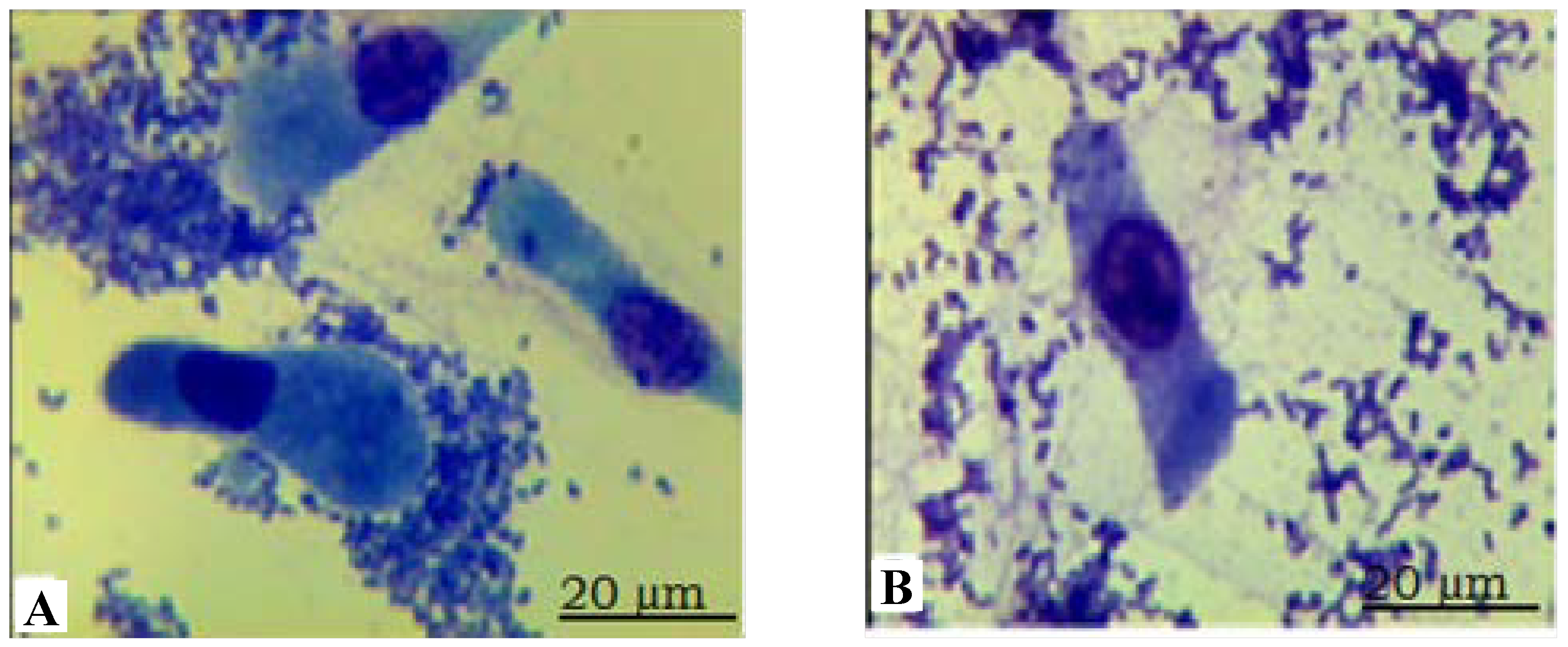

{kind=link}
{kind=link}
| Tissues | Identified Groups (Number) | Difference Multiples | |
|---|---|---|---|
| Resistant (8) | Susceptible (8) | ||
| Heart | 2.49 ± 0.94 | 1.74 ± 0.81 | 1.43 |
| Liver | 5.98 ± 1.65 | 4.75 ± 1.37 | 1.25 |
| Spleen | 25.86 ± 6.52 a | 13.21 ± 5.28 b | 1.96 |
| Lung | 34.65 ± 8.24 a | 18.28 ± 6.64 b | 1.90 |
| Kidney | 12.14 ± 2.51 | 9.19 ± 2.01 | 1.32 |
| Stomach | 9.85 ± 2.32 | 7.21 ± 2.74 | 1.37 |
| Muscle | 1.21 ± 0.45 | 1.00 ± 0.00 | 1.21 |
| Lymph | 36.85 ± 7.55 a | 16.44 ± 4.38 b | 2.24 |
| Thymus | 27.21 ± 4.21 a | 11.52 ± 3.21 b | 2.37 |
| Duodenum | 28.52 ± 9.88 a | 15.48 ± 6.58 b | 1.84 |
| Jejunum | 32.19 ± 4.58 a | 16.59 ± 7.05 b | 1.94 |
2.2. TAP1 Gene mRNA Expression Levels in 11 Tissues of the E. coli F18-Resistant and E. coli F18-Susceptible Sutai Piglets
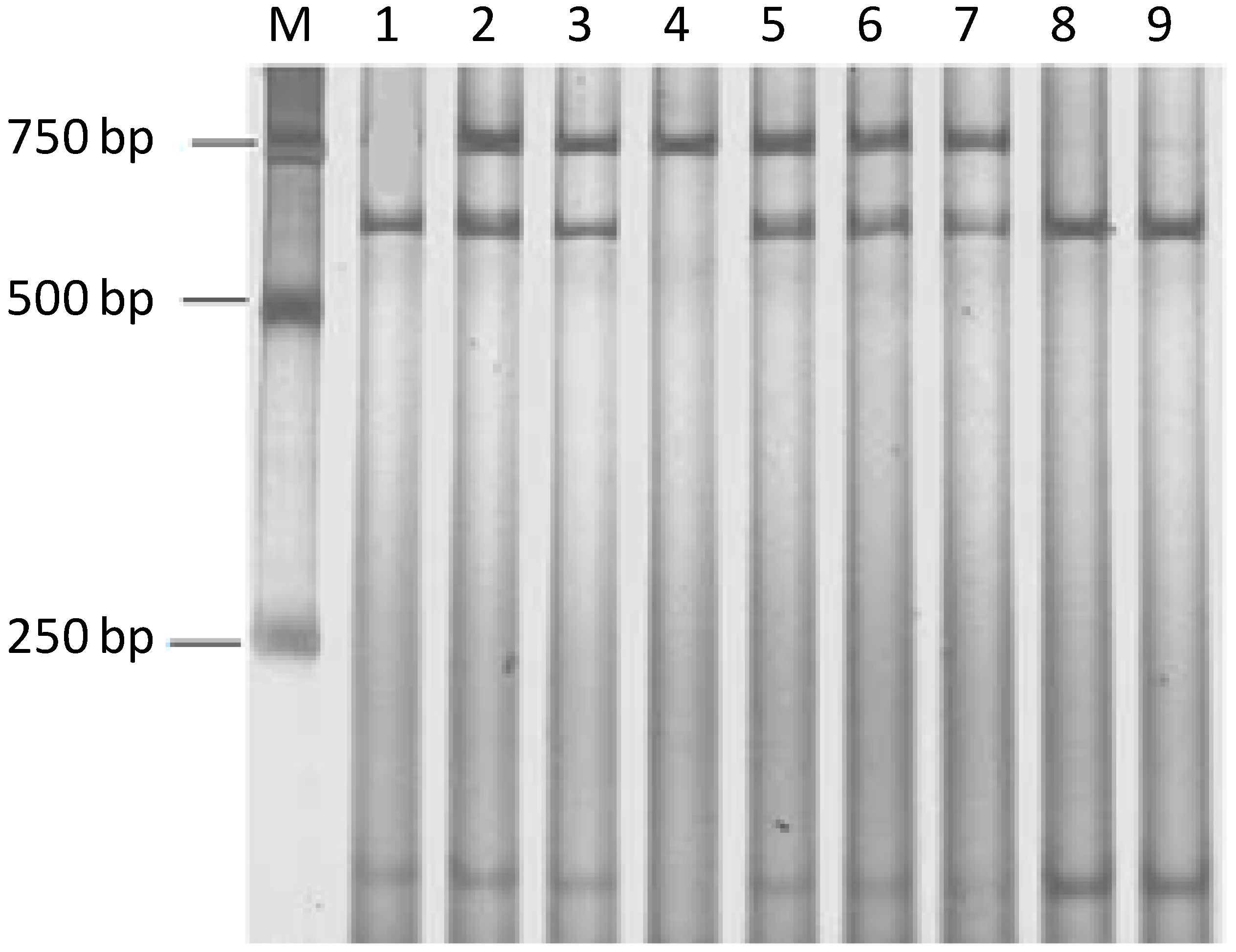
2.3. Analysis of PCR-RFLP and Sequencing Results

2.4. Analysis of Gene Frequency Distribution and Hardy-Weinberg Equilibrium of the TAP1 Gene Exon 3 G729A Mutation
| Sample Number | Genotype Frequency | Allele Frequency | χ2 Value | |||
|---|---|---|---|---|---|---|
| AA | AG | GG | A | G | ||
| 196 | 0.602 (118) | 0.276 (54) | 0.122 (24) | 0.740 | 0.260 | 15.851 |
2.5. Levels of TAP1 mRNA Relative Expression in Different Genotypes
| Tissues | Genotype (Number) | ||
|---|---|---|---|
| AA (8) | AG (8) | GG (8) | |
| Heart | 0.69 ± 0.09 a | 3.19 ± 1.73 b | 3.40 ± 2.01 b |
| Liver | 3.21 ± 1.45 a | 7.76 ± 5.49 a | 19.38 ± 12.32 b |
| Spleen | 10.91 ± 0.30 | 23.82 ± 13.68 | 65.28 ± 23.47 |
| Lung | 26.77 ± 7.40 a | 25.37 ± 5.94 a | 101.39 ± 41.12 b |
| Kidney | 6.75 ± 0.82 a | 11.46 ± 5.98 a | 25.81 ± 23.31 b |
| Stomach | 8.27 ± 1.39 | 21.13 ± 12.29 | 51.87 ± 20.91 |
| Muscle | 1.00 ± 0.00 | 1.07 ± 0.23 | 1.03 ± 0.15 |
| Thymus | 17.54 ± 6.00 a | 18.74 ± 7.59 a | 61.16 ± 19.34 b |
| Lymph node | 31.18 ± 21.72 a | 25.39 ± 15.01 a | 75.15 ± 21.3.8 b |
| Duodenum | 43.14 ± 4.68 a | 30.22 ± 12.18 a | 59.97 ± 15.26 b |
| Jejunum | 30.16 ± 0.28 a | 31.14 ± 15.51 a | 89.91 ± 15.07 b |
3. Discussion
4. Experimental Section
4.1. Experimental Materials and Sample Collection
4.1.1. Escherichia coli F18-Resistant and E. coli F18-Susceptible Sutai Pigs
4.1.2. Other Experimental Samples
4.2. Primer Design
| Gene | Method | Sequence (5'–3') | Annealing Temperature | Length |
|---|---|---|---|---|
| TAP1 | PCR-RFLP | GAAATGTGGATAAGAGCA | 63 °C | 767 bp |
| AAACAGACGGATAATGAAAGAGG | ||||
| TAP1 | Real-time PCR | CCACTGCTTTTCCTTCTGCCT | 60 °C | 109 bp |
| ACAGAACCTCAATGGCCACCT | ||||
| GAPDH | Real-time PCR | ACATCATCCCTGCTTCTACTGG | 60 °C | 187 bp |
| CTCGGACGCCTGCTTCAC |
4.3. PCR-RFLP Analysis
4.4. Total RNA Extraction and Real-Time PCR
4.5. Data Processing and Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Paulsson, K.M. Evolutionary and functional perspectives of the major histocompatibility complex class I antigen processing machinery. Cell. Mol. Life Sci. 2004, 61, 2446–2460. [Google Scholar]
- Cresswell, P.; Bangia, N.; Dick, T.; Diedrich, G. The nature of the MHC class I peptide loading complex. Immun. Rev. 1999, 172, 21–28. [Google Scholar] [CrossRef]
- Powis, S.J.; Deverson, E.V.; Coadwell, W.J.; Ciruela, A.; Huskisson, N.S.; Smith, H.; Butcher, G.W.; Howard, J.C. Effect of polymorphism of an MHC-linked transporter on the peptides assembled in a class I molecule. Nature 1992, 357, 211–215. [Google Scholar]
- Witkowska-Tobola, A.M.; Szczerkowska-Dobosz, A.; Nedoszytko, B.; Roszkiewicz, J. Polymorphism of the TAP1 gene in Polish patients with psoriasis vulgaris. J. Appl. Genet. 2004, 45, 391–393. [Google Scholar]
- Vaske, D.A.; Ruohonen-Lehto, M.K.; Larson, R.G.; Rothschild, M.F. Rapid communication: Restriction fragment length polymorphisms at the porcine transporter associated with antigen processing 1 (TAP1) locus. J. Anim. Sci. 1994, 72, 255. [Google Scholar]
- Sun, N.N.; Liu, D.W.; Chen, H.B.; Liu, X.D.; Meng, F.M.; Zhang, X.W.; Chen, H.Y.; Xie, S.X.; Li, X.Y.; Zhang, Z.F. Localization, expression change in PRRSV infection and association analysis of the porcine TAP1 gene. Int. J. Biol. Sci. 2012, 8, 49–58. [Google Scholar]
- Bao, W.B.; Ye, L.; Pan, Z.Y.; Zhu, J.; Du, Z.D.; Zhu, G.Q.; Huang, X.G.; Wu, S.L. Microarray analysis of differential gene expression in sensitive and resistant pig to Escherichia coli F18. Anim. Genet. 2012, 43, 525–534. [Google Scholar] [CrossRef]
- Wysocki, M.; Chen, H.; Steibel, J.P.; Kuhar, D.; Petry, D.; Bates, J.; Johnson, R.; Ernst, C.W.; Lunney, J.K. Identifying putative candidate genes and pathways involved in immune responses to porcine reproductive and respiratory syndrome virus (PRRSV) infection. Anim. Genet. 2012, 43, 328–332. [Google Scholar] [CrossRef]
- Uthe, J.J.; Bearson, S.M.D.; Qu, L.; Dekkers, J.C.; Nettleton, D.; Torres, Y.R.; O’Connor, A.M.; Mckean, J.D.; Tuggle, C.K. Integrating comparative expression profiling data and association of SNPs with Salmonella shedding for improved food safety and porcine disease resistance. Anim. Genet. 2011, 42, 521–534. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Dong, W.H.; Huo, Y.J.; Huang, X.G.; Wu, S.L.; Bao, W.B. Dynamic changes in TAP1 expression levels in newborn to weaning piglets, and its association with Escherichia coli F18 resistance. Genet. Mol. Res. 2014, 13, 3686–3692. [Google Scholar] [CrossRef]
- Liu, L.; Wang, J.; Zhao, Q.H.; Zi, C.; Wu, Z.C.; Su, X.M.; Huo, Y.J.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Genetic variation in exon 10 of the BPI gene is associated with Escherichia coli F18 susceptibility in Sutai piglets. Gene 2013, 523, 70–75. [Google Scholar] [CrossRef]
- Wu, S.L.; Yuan, Z.W.; Ju, H.P.; Huang, X.G.; Hua, J.D.; Shen, J.L.; Zhou, G.Y.; Wang, J.Y.; Xie, K.Z.; Chen, G.H.; et al. Polymorphisms of the FUT1 gene M307 locus in post-weaning Sutai breed piglet and resistance to F18 fimbrial Escherichia coli in vitro. Chin. J. Prev. Vet. Med. 2007, 29, 783–787. [Google Scholar]
- Eyal, R. Organ-specific regulation of innate immunity. Nat. Immun. 2007, 8, 3–4. [Google Scholar]
- Du, S.D.; Mao, Y.L.; He, G.Z. New understanding on pathway of intestinal infection: Lymphatic immune passage. Chin. J. Pract. Surg. 2002, 22, 549–551. [Google Scholar]
- Kamarashev, J.; Ferrone, S.; Seifert, B.; Boni, R.; Nestle, F.; Burg, G.; Dummer, R. TAP1 down-regulation in primary melanoma lesions: An independent marker of poor prognosis. Int. J. Cancer 2001, 95, 23–28. [Google Scholar] [CrossRef]
- Durgeau, A.; Hage, F.E.; Vergnon, I.; Validire, P.; Montpreville, V.D.; Besse, B.; Soria, J.C.; Hall, T.V.; Mami-Chouaib, F. Different expression levels of the TAP peptide transporter lead to recognition of different antigenic peptides by tumor-specific CTL. J. Immun. 2011, 187, 5532–5539. [Google Scholar]
- Zhang, Q.J.; Seipp, R.P.; Chen, S.S.; Vitalis, T.Z.; Li, X.L.; Choi, K.B.; Jeffries, A.; Jefferies, W.A. TAP expression reduces IL-10 expressing tumor infiltrating lymphocytes and restores immunosurveillance against melanoma. Int. J. Cancer 2007, 120, 1935–1941. [Google Scholar] [CrossRef]
- Zhao, S.H.; Kuhar, D.; Lunney, J.K.; Dawson, H.; Guidry, C.; Uthe, J.J.; Bearson, S.M.D.; Recknor, J.; Nettleton, D.; Tuggle, C.K. Gene expression profiling in Salmonella Choleraesuis-infected porcine lung using a long oligonucleotide microarray. Mamm. Genome 2006, 17, 777–789. [Google Scholar] [CrossRef]
- Li, Y.T.; Zhou, H.B.; Wen, Z.B.; Wu, S.J.; Huang, C.H.; Jia, G.M.; Chen, H.C.; Jin, M.L. Transcription analysis on response of swine lung to H1N1 swine influenza virus. BMC Genomic. 2011, 12, 398–412. [Google Scholar] [CrossRef]
- Schultz, H.; Schinke, S.; Weiss, J.; Cerundolo, V.; Gross, W.L.; Gadola, S. BPI-ANCA intransporter associated with antigen presentation (TAP) deficiency: Possible role in susceptibility to Gram-negative bacterial infections. Clin. Exp. Immun. 2003, 133, 252–259. [Google Scholar] [CrossRef]
- Salle, H.D.L.; Hanau, D.; Fricker, D.; Urlacher, A.; Kelly, A.; Salamero, J.; Powis, S.H.; Donato, L.; Bausinger, H.; Laforet, M.L.; et al. Homozygous human TAP peptide transporter mutation in HLA class I deficiency. Science 1994, 265, 237–241. [Google Scholar]
- Salle, H.D.L.; Zimmer, J.; Fricher, D.; Angenieux, C.; Cazenave, J.P.; Okubo, M.; Maeda, H.; Plebani, A.; Tongio, M.M.; Dormoy, A.; et al. HLA class I deficiencies due to mutations in subunit 1 of the peptide transporter TAP1. J. Clin. Investig. 1999, 103, 9–13. [Google Scholar] [CrossRef]
- Kimchi-Sarfaty, C.; Oh, J.M.; Kim, I.W.; Saune, Z.E.; Calcagno, A.M.; Ambudkar, S.V.; Gottesman, M.M. A “silent” polymorphism in the MDR1 gene changes substrate specificity. Science 2007, 315, 525–528. [Google Scholar] [CrossRef]
- Benin, A.M.; Ducher-Suchaux, M.F. Relationship between virulence and adherence of various enterotoxigenic Escherichia coli: Strains to isolated intestinal epithelial cells from Chinese Meishan and European large white pigs. Am. J. Vet. Res. 1991, 52, 45–49. [Google Scholar]
- Chen, H.Y.; Wu, Z.F. Prospective of BF, DRB, DQB, TAP1 and IFN-γ genes used as porcine marker assisted disease resistance and breeding. Biotechnol. Bull. 2009, 1, 103–106. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; CSHL Press: New York, NY, USA, 2001; pp. 14–19. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhao, Q.; Liu, Y.; Dong, W.; Zhu, S.; Huo, Y.; Wu, S.; Bao, W. Genetic Variations of TAP1 Gene Exon 3 Affects Gene Expression and Escherichia coli F18 Resistance in Piglets. Int. J. Mol. Sci. 2014, 15, 11161-11171. https://doi.org/10.3390/ijms150611161
Zhao Q, Liu Y, Dong W, Zhu S, Huo Y, Wu S, Bao W. Genetic Variations of TAP1 Gene Exon 3 Affects Gene Expression and Escherichia coli F18 Resistance in Piglets. International Journal of Molecular Sciences. 2014; 15(6):11161-11171. https://doi.org/10.3390/ijms150611161
Chicago/Turabian StyleZhao, Qiaohui, Ying Liu, Wenhua Dong, Shiping Zhu, Yongjiu Huo, Shenglong Wu, and Wenbin Bao. 2014. "Genetic Variations of TAP1 Gene Exon 3 Affects Gene Expression and Escherichia coli F18 Resistance in Piglets" International Journal of Molecular Sciences 15, no. 6: 11161-11171. https://doi.org/10.3390/ijms150611161