Damage of Neuroblastoma Cell SH-SY5Y Mediated by MPP+ Inhibits Proliferation of T-Cell Leukemia Jurkat by Co-Culture System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
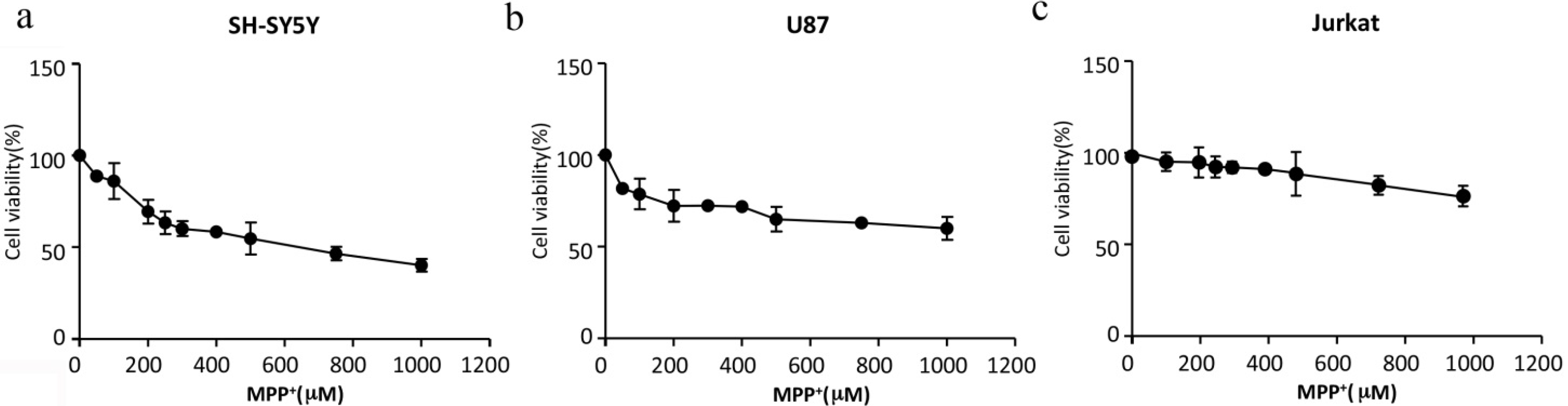
2.1. Cell Viability after MPP+ Treatment

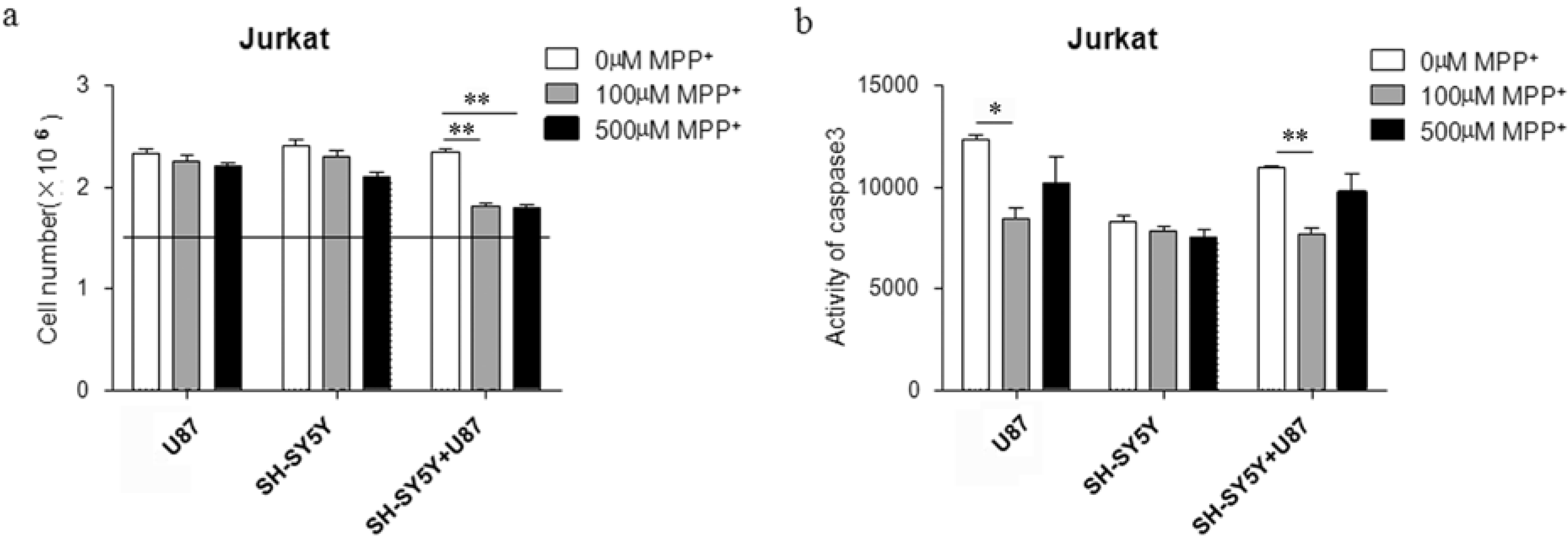
2.2. Proliferation of Jurkat Cells Was Inhibited in MPP+-Treated SH-SY5Y and U87 Cells Co-Culture Medium
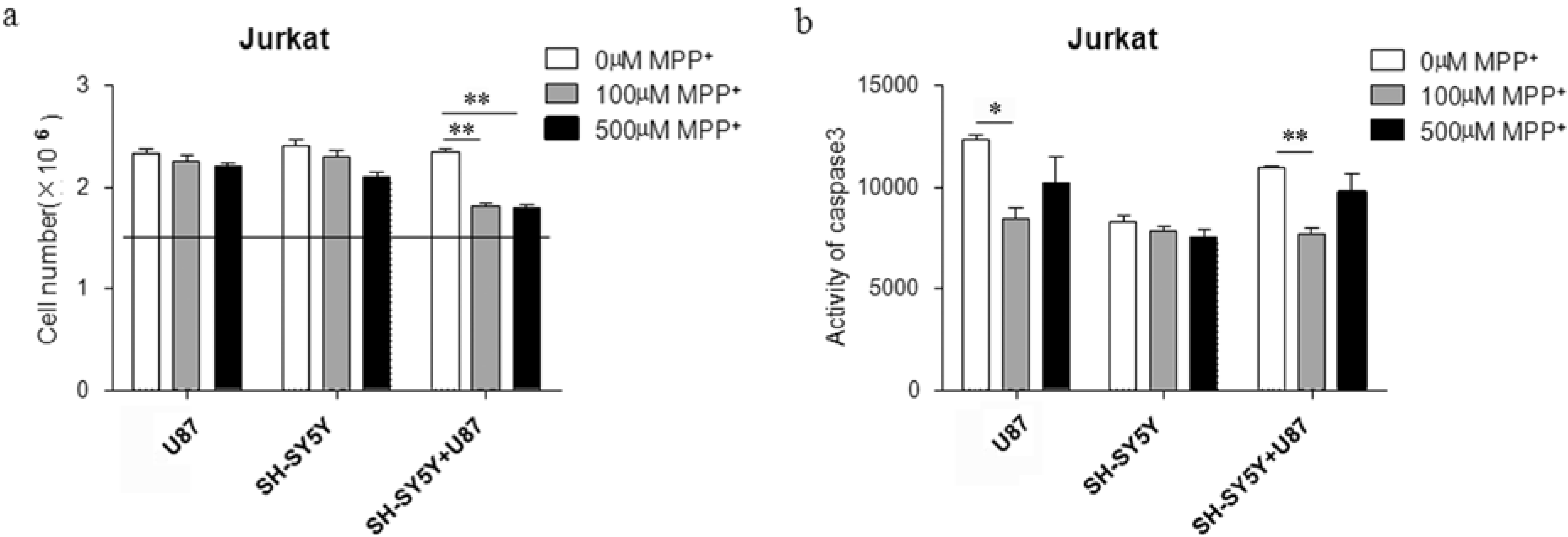
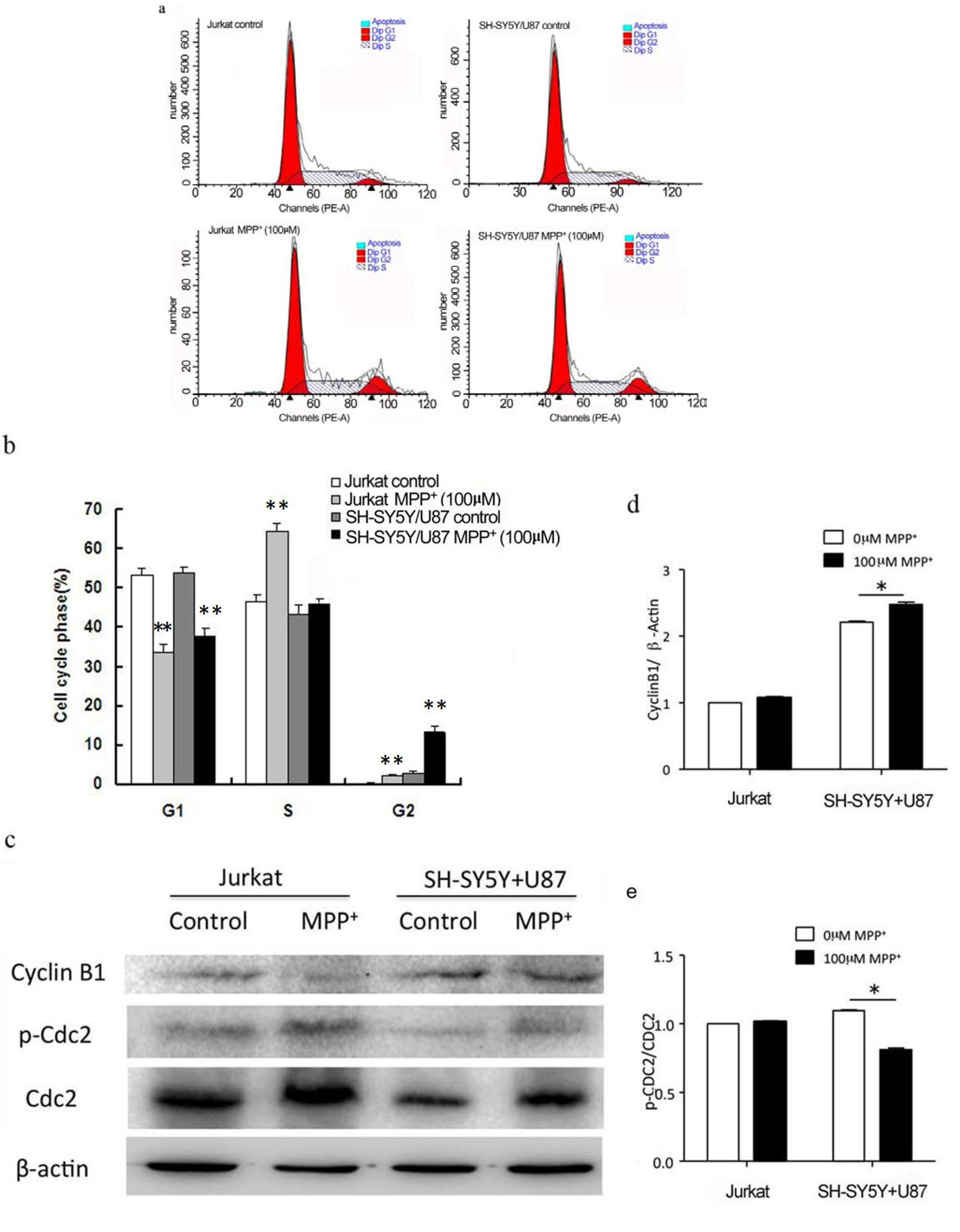
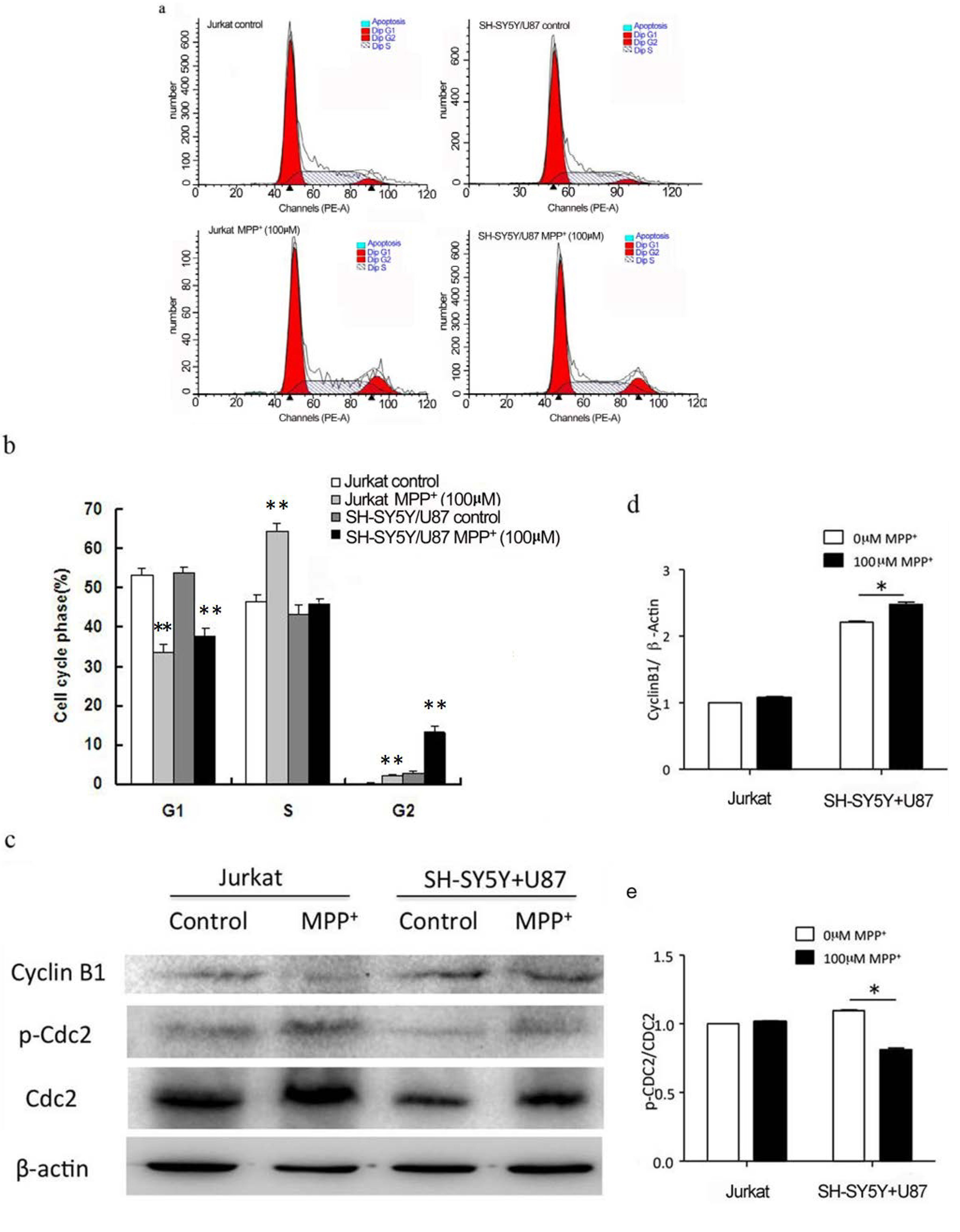
2.3. Low Concentration of MPP+-Treated SH-SY5Y and U87 Cells Co-Culture Medium-Induced Accumulation of G2/M Phase in Jurkat Cells
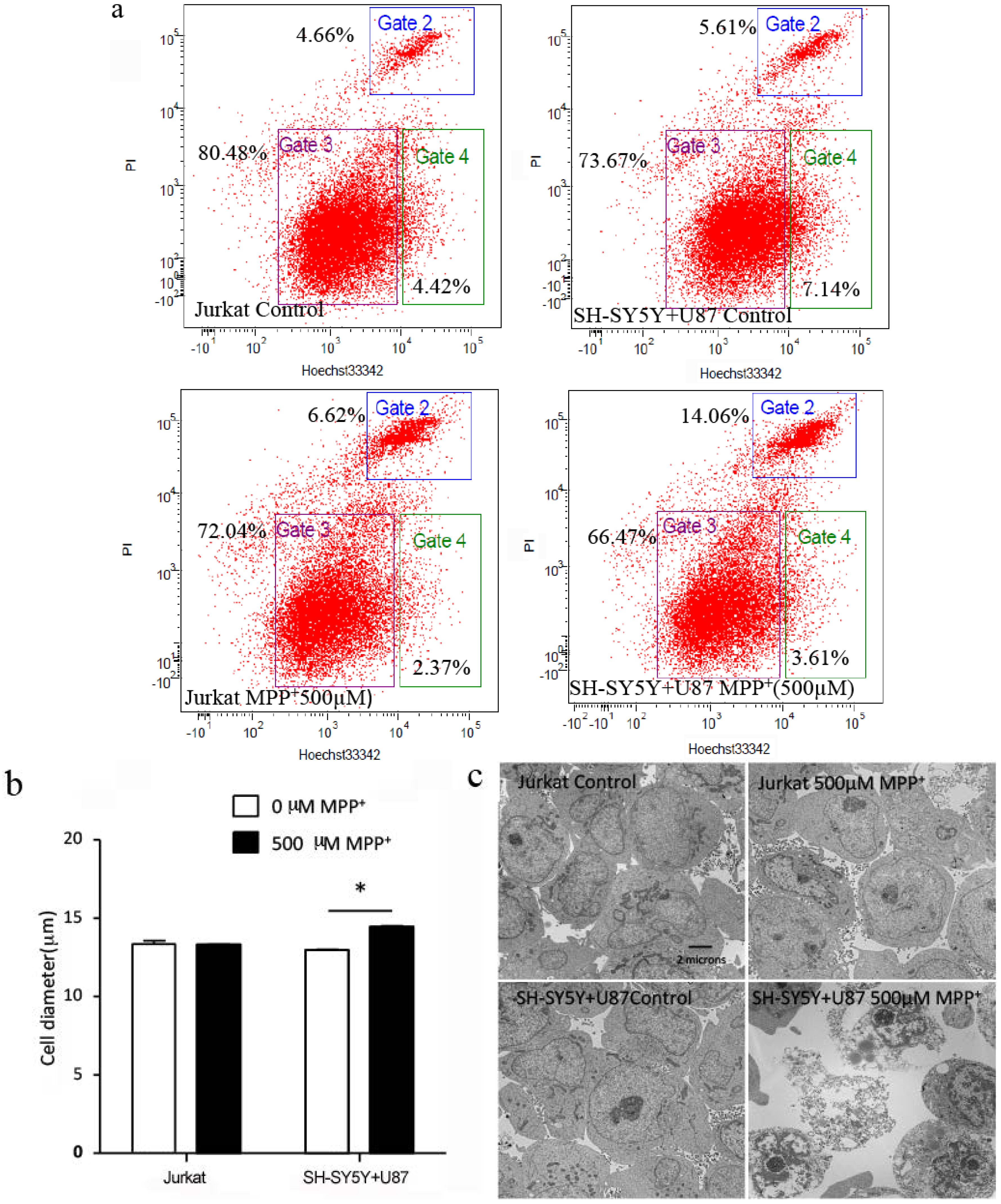
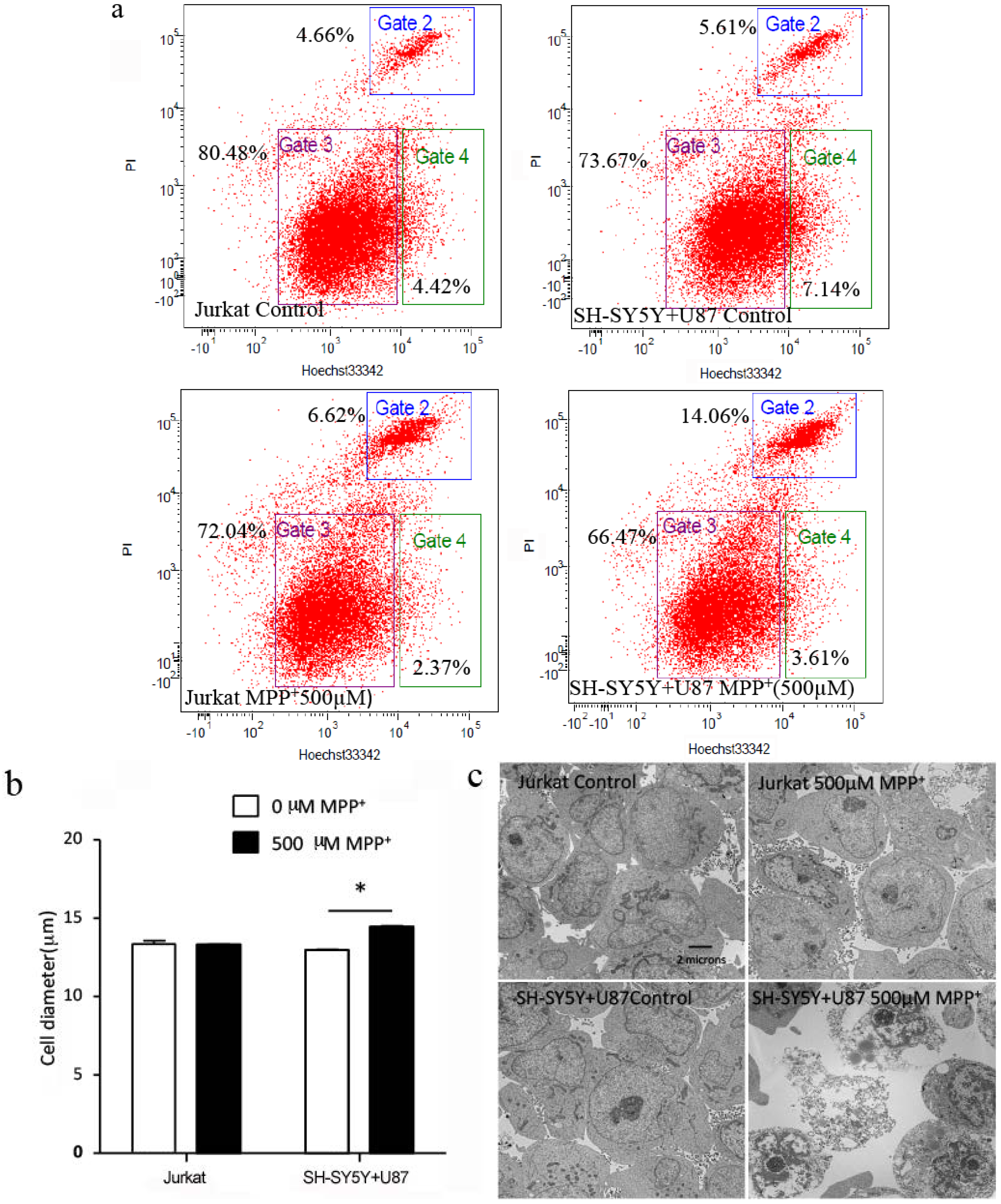
2.4. High Concentration of MPP+-Treated SH-SY5Y and U87 Cells Co-Culture Medium-Induced Jurkat Necrosis
3. Experimental Section
3.1. Chemicals and Reagents
3.2. Cell Culture
3.3. Cell Co-Culture
3.4. Caspase3/8 Activity Assays
3.5. Cell Viability Assay
3.6. Western Blot
3.7. Flow Cytometry Analysis
3.8. Transmission Electron Microscopy (TEM) Assay
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Abbreviations
Conflicts of Interest
References
- Marras, C.; Lang, A. Invited article: Changing concepts in Parkinson disease: Moving beyond the decade of the brain. Neurology 2008, 70, 1996–2003. [Google Scholar]
- Lin, X.; Parisiadou, L.; Sgobio, C.; Liu, G.; Yu, J.; Sun, L.; Shim, H.; Gu, X.L.; Luo, J.; Long, C.X.; et al. Conditional expression of Parkinson’s disease-related mutant α-Synuclein in the midbrain dopaminergic neurons causes progressive neurodegeneration and degradation of transcription factor nuclear receptor related 1. J. Neurosci. 2012, 32, 9248–9264. [Google Scholar]
- Ribeiro, R.P.; Moreira, E.L.; Santos, D.B.; Colle, D.; dos Santos, A.A.; Peres, K.C.; Figueiredo, C.P.; Farina, M. Probucol affords neuroprotection in a 6-OHDA mouse model of Parkinson’s disease. Neurochem. Res. 2013, 38, 660–668. [Google Scholar]
- Lu, C.; Zhang, J.; Shi, X.; Miao, S.; Bi, L.; Zhang, S.; Yang, Q.; Zhou, X.; Zhang, M.; Xie, Y.; et al. Neuroprotective effects of tetramethylpyrazine against dopaminergic neuron injury in a rat model of Parkinson’s disease induced by MPTP. Int. J. Biol. Sci. 2014, 10, 350–357. [Google Scholar]
- Ransom, B.R.; Kunis, D.M.; Irwin, I.; Langston, J.W. Astrocytes convert the parkinsonism inducing neurotoxin, MPTP, to its active metabolite, MPP+. Neurosci. Lett. 1987, 75, 323–328. [Google Scholar]
- Williams, C.; Ramsden, D.B. Autotoxicity, methylation and a road to the prevention of Parkinson’s disease. J. Clin. Neurosci. 2005, 12, 6–11. [Google Scholar]
- McGeer, P.L.; Itagaki, S.; Tago, H.; McGeer, E.G. Reactive microglia in patients with senile dementia of the Alzheimer type are positive for the histocompatibility glycoprotein HLA-DR. Neurosci. Lett. 1987, 79, 195–200. [Google Scholar]
- McGeer, P.L.; Itagaki, S.; Boyes, B.E.; McGeer, E.G. Reactive microglia are positive for HLA-DR in the substantia nigra of Parkinson’s and Alzheimer’s disease brains. Neurology 1988, 38, 1285–1291. [Google Scholar]
- Farkas, E.; de Jong, G.I.; de Vos, R.A.; Jansen Steur, E.N.; Luiten, P.G. Pathological features of cerebral cortical capillaries are doubled in Alzheimer’s disease and Parkinson’s disease. Acta Neuropathol. 2000, 100, 395–402. [Google Scholar]
- Brochard, V.; Combadiere, B.; Prigent, A.; Laouar, Y.; Perrin, A.; Beray-Berthat, V.; Bonduelle, O.; Alvarez-Fischer, D.; Callebert, J.; Launay, J.M.; et al. Infiltration of CD4+ lymphocytes into the brain contributes to neurodegeneration in a mouse model of Parkinson disease. J. Clin. Investig. 2009, 119, 182–192. [Google Scholar]
- Faucheux, B.A.; Bonnet, A.M.; Agid, Y.; Hirsch, E.C. Blood vessels change in the mesencephalon of patients with Parkinson’s disease. Lancet 1999, 353, 981–982. [Google Scholar]
- Baba, Y.; Kuroiwa, A.; Uitti, R.J.; Wszolek, Z.K.; Yamada, T. Alterations of T-lymphocyte populations in Parkinson disease. Parkinsonism Relat. Disord. 2005, 11, 493–498. [Google Scholar]
- Hisanaga, K.; Asagi, M.; Itoyama, Y.; Iwasaki, Y. Increase in peripheral CD4+ bright CD8+ dull T cells in Parkinson disease. Arch. Neurol. 2001, 58, 1580–1583. [Google Scholar]
- Calopa, M.; Bas, J.; Callen, A.; Mestre, M. Apoptosis of peripheral blood lymphocytes in Parkinson patients. Neurobiol. Dis. 2010, 38, 1–7. [Google Scholar]
- Abraham, R.T.; Weiss, A. Jurkat T cells and development of the T-cell receptor signalling paradigm. Nat. Rev. Immunol. 2004, 4, 301–308. [Google Scholar]
- Alberio, T.; Anchieri, C.; Piacentini, L.; Gentile, G.; Simmaco, M.; Biasin, M.; Fasano, M. Proteomic characterization of Jurkat T leukemic cells after dopamine stimulation: A model of circulating dopamine-sensitive cells. Biochimie 2011, 93, 892–898. [Google Scholar]
- Terashvili, M.; Sarkar, P.; Nostrand, M.V.; Falck, J.R.; Harder, D.R. The protective effect of astrocyte-derived 14,15-epoxyeicosatrienoic acid on hydrogen peroxide-induced cell injury in astrocyte-dopaminergic neuronal cell line co-culture. Neuroscience 2012, 223, 68–76. [Google Scholar]
- Ma, H.; Rao, L.; Wang, H.L.; Mao, Z.W.; Lei, R.H.; Yang, Z.Y.; Qing, H.; Deng, Y.L. Transcriptome analysis of Glioma cells for the dynamic response to γ-irradiation and dual regulation of apoptosis genes: A new insight into radiotherapy for glioblastomas. Cell Death Dis. 2013, 4, e895. [Google Scholar]
- Langston, J.W.; Ballard, P.; Tetrud, J.W.; Irwin, I. Chronic Parkinsonism in humans due to a product of meperidine-analog synthesis. Science 1983, 219, 979–980. [Google Scholar]
- Nicklas, W.J.; Vyas, I.; Heikkila, R.E. Inhibition of NADH-linked oxidation in brain mitochondria by 1-methyl-4-phenyl-pyridine, a metabolite of the neurotoxin, 1-methyl-4-phenyl-1,2,5,6-tetrahydropyridine. Life Sci. 1985, 36, 2503–2508. [Google Scholar]
- Mogi, M.; Harada, M.; Riederer, P.; Narabayashi, H.; Fujita, K.; Nagatsu, T. Tumor necrosis factor-alpha (TNF-α) increases both in the brain and in the cerebrospinal fluid from parkinsonian patients. Neurosci. Lett. 1994, 165, 208–210. [Google Scholar]
- Mogi, M.; Harada, M.; Kondo, T.; Riederer, P.; Inagaki, H.; Minami, M.; Nagatsu, T. Interleukin-1β, interleukin-6, epidermal growth factor and transforming growth factor-α are elevated in the brain from parkinsonian patients. Neurosci. Lett. 1994, 180, 147–150. [Google Scholar]
- Hunot, S.; Hirsch, E.C. Neuroinflammatory processes in Parkinson’s disease. Ann. Neurol. 2003, 53, S49–S60. [Google Scholar]
- Koziorowski, D.; Tomasiuk, R.; Szlufik, S.; Friedman, A. Inflammatory cytokines and NT-proCNP in Parkinson’s disease patients. Cytokine 2012, 60, 762–766. [Google Scholar]
- Bas, J.; Calopa, M.; Mestre, M.; Mollevi, D.G.; Cutillas, B.; Ambrosio, S.; Buendia, E. Lymphocyte populations in Parkinson’s disease and in rat models of parkinsonism. J. Neuroimmunol. 2001, 113, 146–152. [Google Scholar]
- Tang, L.; Gao, Y.; Yan, F.; Tang, J. Evaluation of cyclin-dependent kinase-like 1 expression in breast cancer tissues and its regulation in cancer cell growth. Cancer Biother. Radiopharm. 2012, 27, 392–398. [Google Scholar]
- Zhang, X.; Jia, S.; Yang, S.; Yang, Y.; Yang, T. Arsenic trioxide induces G2/M arrest in hepatocellular carcinoma cells by increasing the tumor suppressor PTEN expression. J. Cell. Biochem. 2012, 113, 3528–3535. [Google Scholar]
- Oliveras-Ferraros, C.; Fernandez-Arroyo, S.; Vazquez-Martin, A.; Lozano-Sanchez, J.; Cufi, S.; Joven, J.; Micol, V.; Fernandez-Gutierrez, A.; Segura-Carretero, A.; Menendez, J.A. Crude phenolic extracts from extra virgin olive oil circumvent de novo breast cancer resistance to HER1/HER2-targeting drugs by inducing GADD45-sensed cellular stress, G2/M arrest and hyperacetylation of Histone H3. Int. J. Oncol. 2011, 38, 1533–1547. [Google Scholar]
- Zong, W.X.; Thompson, C.B. Necrotic death as a cell fate. Genes Dev. 2006, 20, 1–15. [Google Scholar]
- Benner, E.J.; Banerjee, R.; Reynolds, A.D.; Sherman, S.; Pisarev, V.M.; Tsiperson, V.; Nemachek, C.; Ciborowski, P.; Przedborski, S.; Mosley, R.L.; et al. Nitrated α-synuclein immunity accelerates degeneration of nigral dopaminergic neurons. PLoS One 2008, 3, e1376. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, F.; Awan, U.F.; Wang, Y.; Wang, L.; Qing, H.; Ma, H.; Deng, Y. Damage of Neuroblastoma Cell SH-SY5Y Mediated by MPP+ Inhibits Proliferation of T-Cell Leukemia Jurkat by Co-Culture System. Int. J. Mol. Sci. 2014, 15, 10738-10750. https://doi.org/10.3390/ijms150610738
Wang F, Awan UF, Wang Y, Wang L, Qing H, Ma H, Deng Y. Damage of Neuroblastoma Cell SH-SY5Y Mediated by MPP+ Inhibits Proliferation of T-Cell Leukemia Jurkat by Co-Culture System. International Journal of Molecular Sciences. 2014; 15(6):10738-10750. https://doi.org/10.3390/ijms150610738
Chicago/Turabian StyleWang, Fuli, Umer Farooq Awan, Yuanyuan Wang, Luna Wang, Hong Qing, Hong Ma, and Yulin Deng. 2014. "Damage of Neuroblastoma Cell SH-SY5Y Mediated by MPP+ Inhibits Proliferation of T-Cell Leukemia Jurkat by Co-Culture System" International Journal of Molecular Sciences 15, no. 6: 10738-10750. https://doi.org/10.3390/ijms150610738