Relaxation of Rat Aorta by Farrerol Correlates with Potency to Reduce Intracellular Calcium of VSMCs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
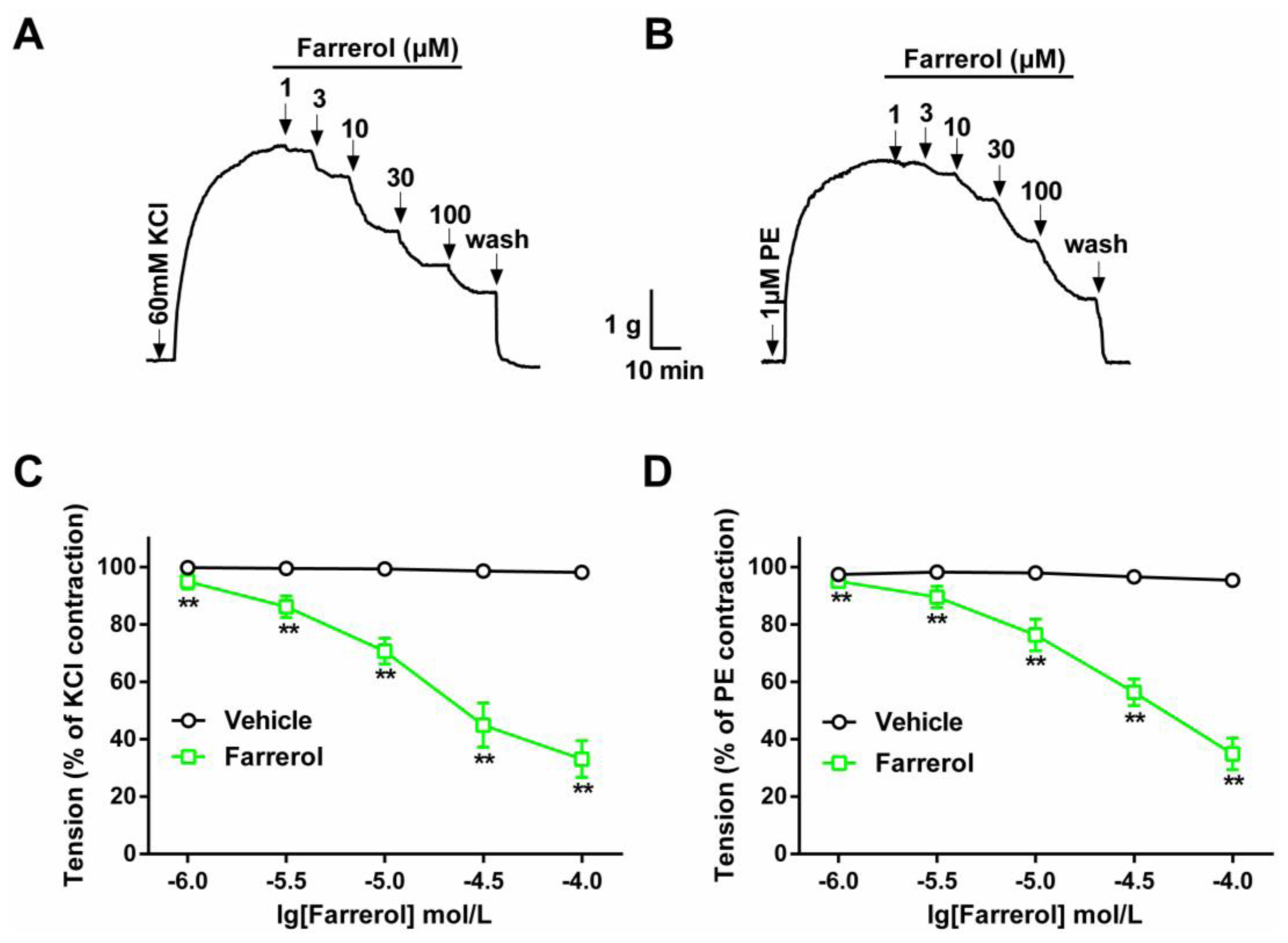
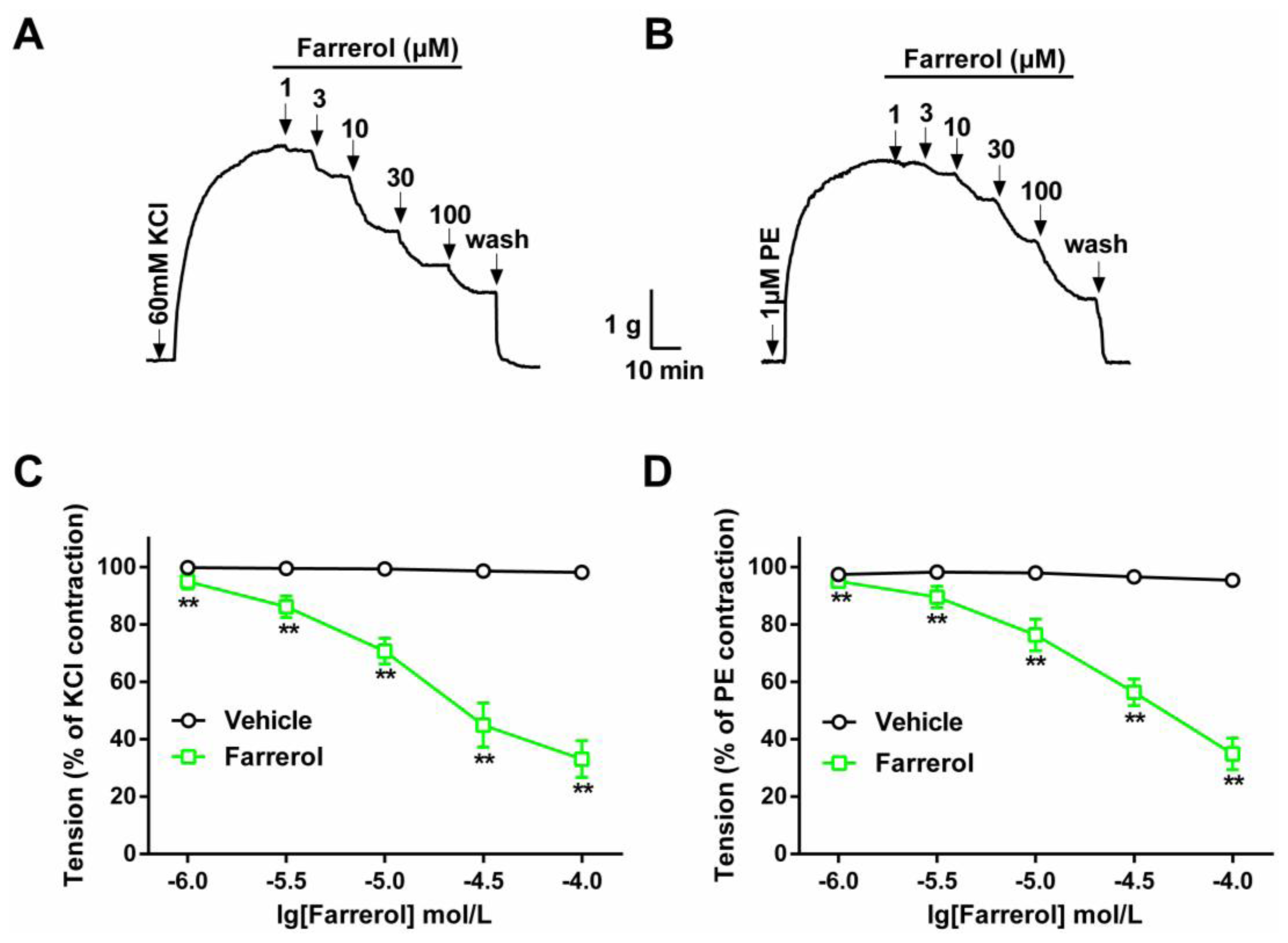
2.1.1. Dose-Dependent Relaxation Effect of Farrerol
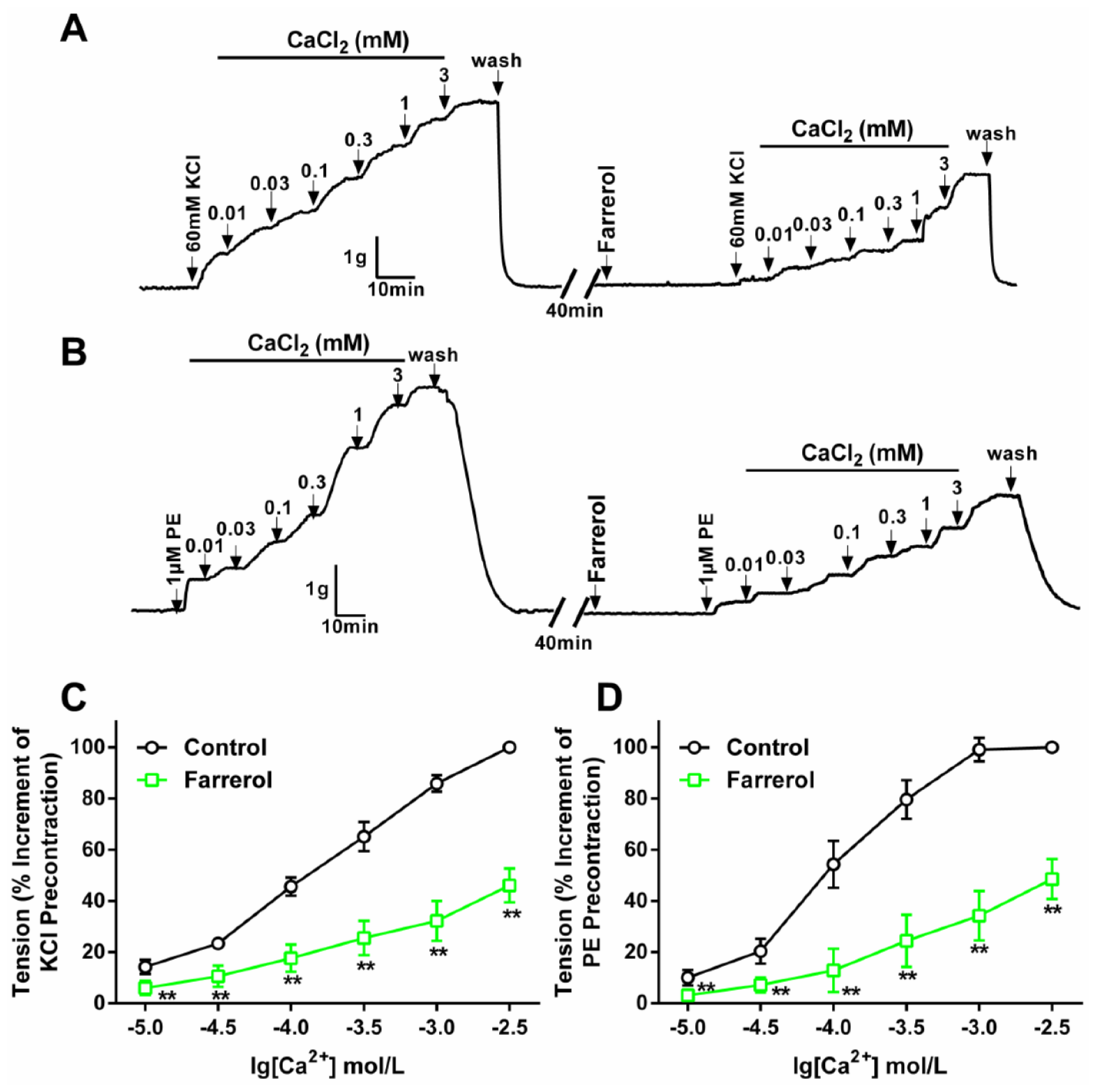
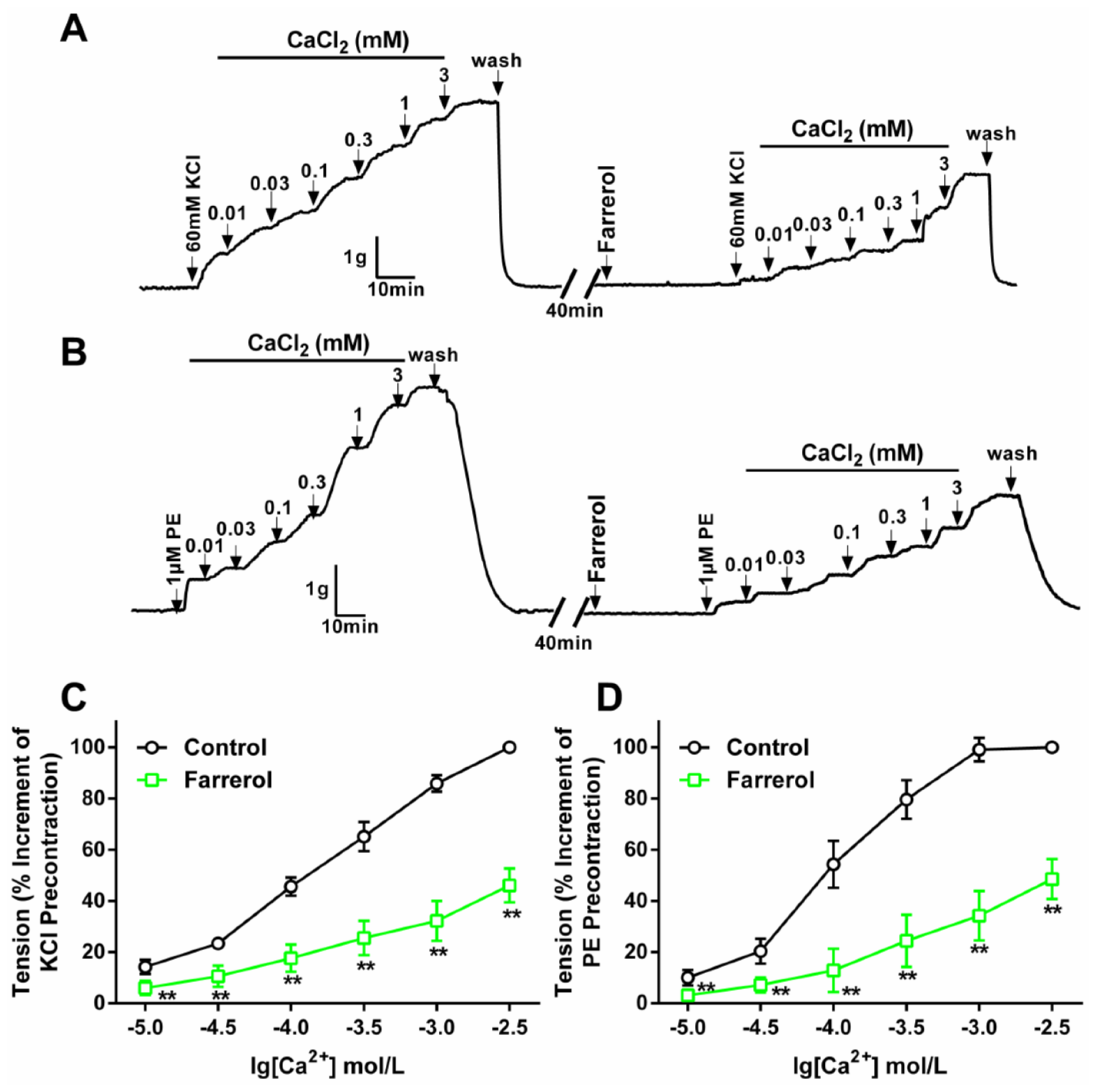
2.1.2. Effect of Farrerol on Extracellular Ca2+-Induced Contraction
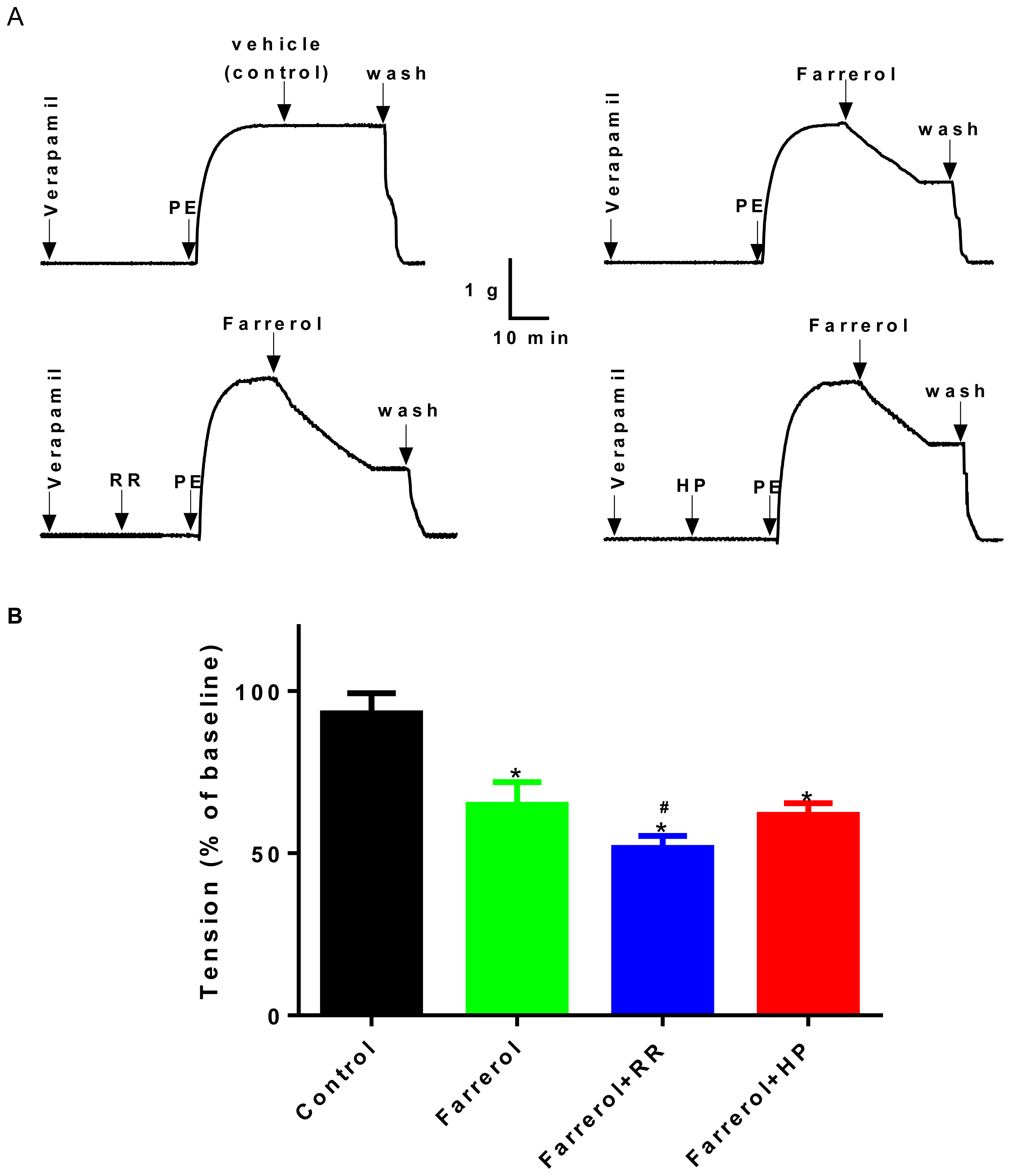
2.1.3. Relationship between Vasoactive Effects of Farrerol and [Ca2+]in Release
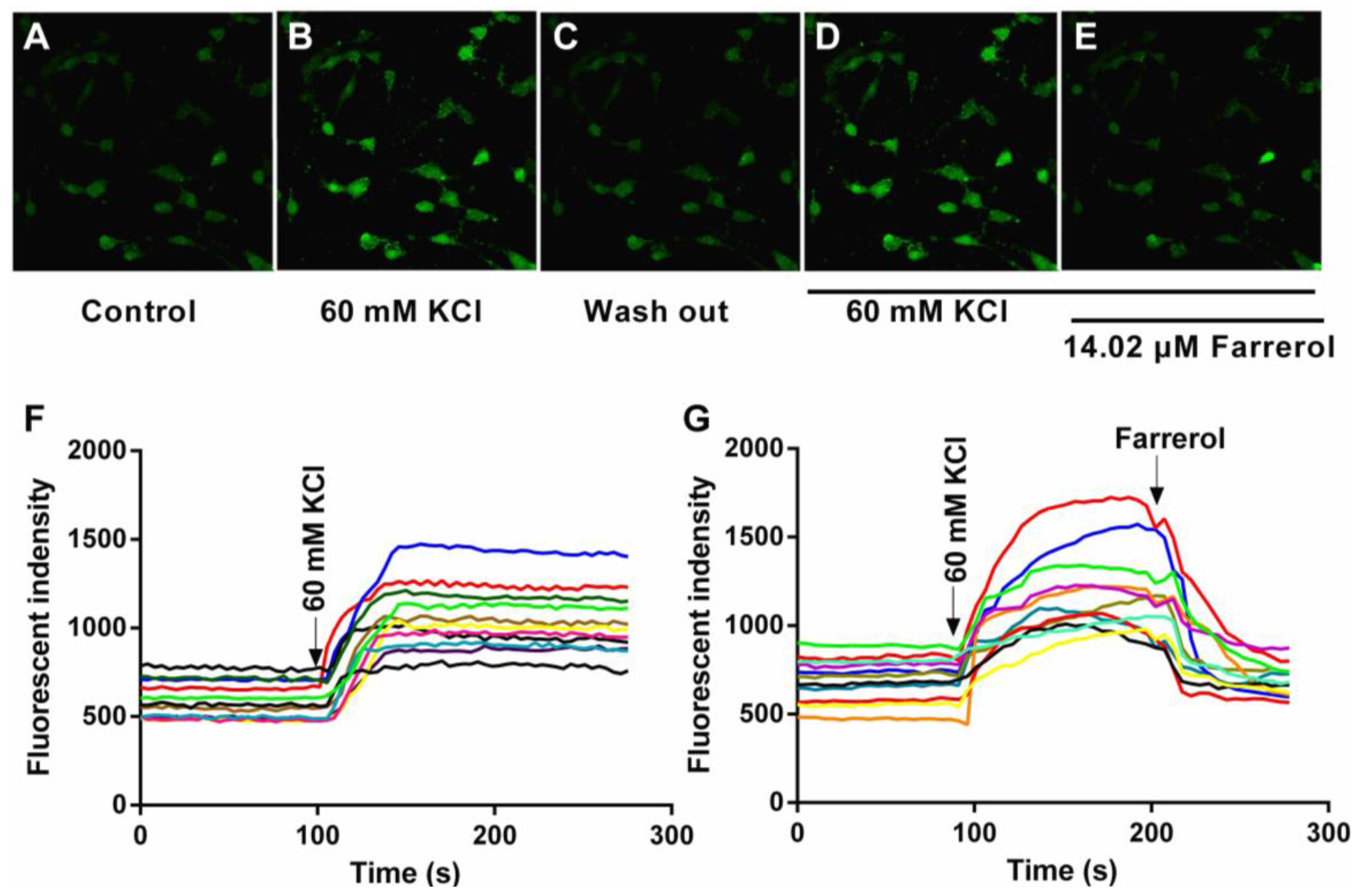
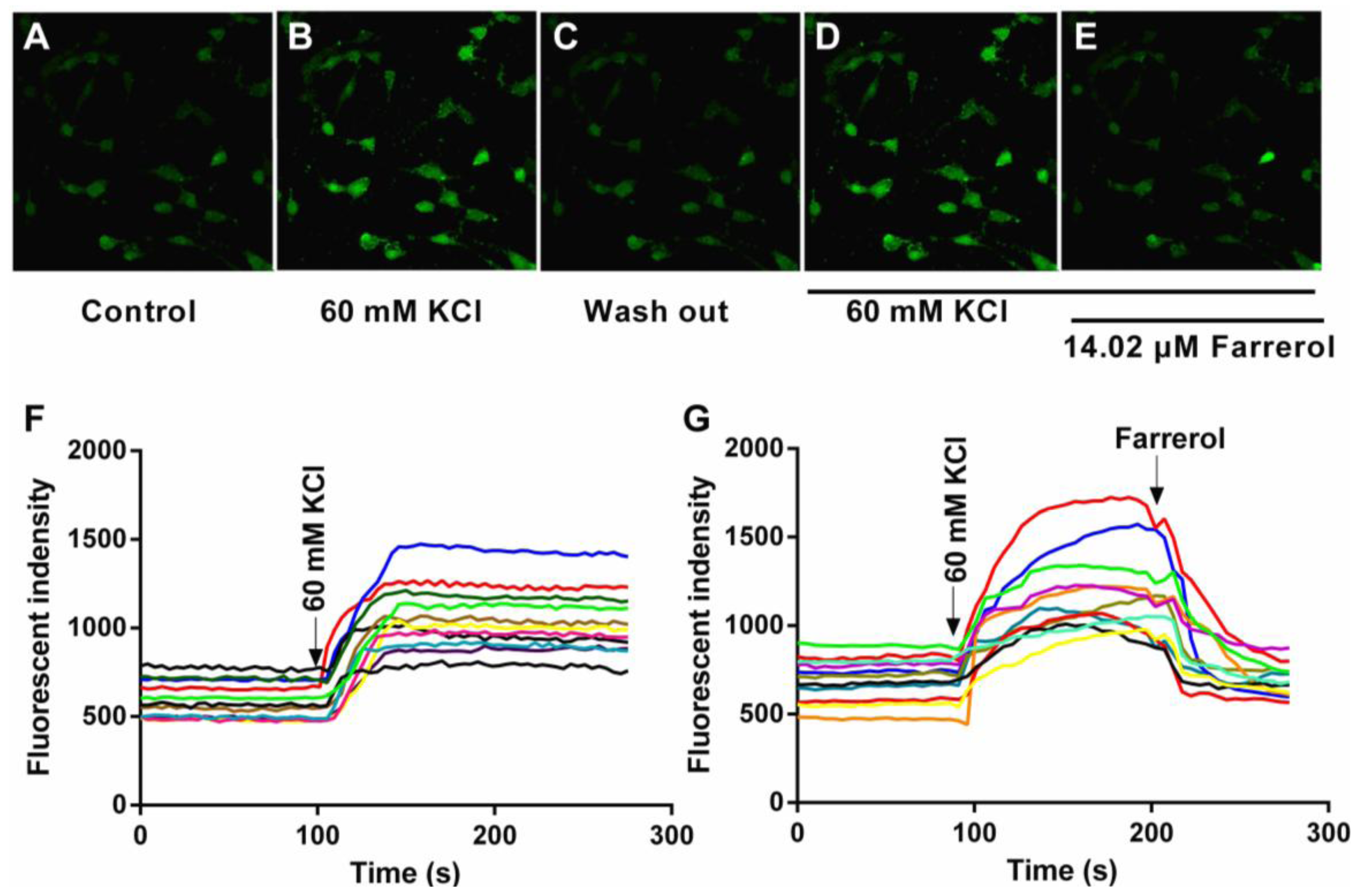
2.1.4. Effects of Farrerol on [Ca2+]in in Rat Aortic Vascular Smooth Muscle Cells (VSMCs)
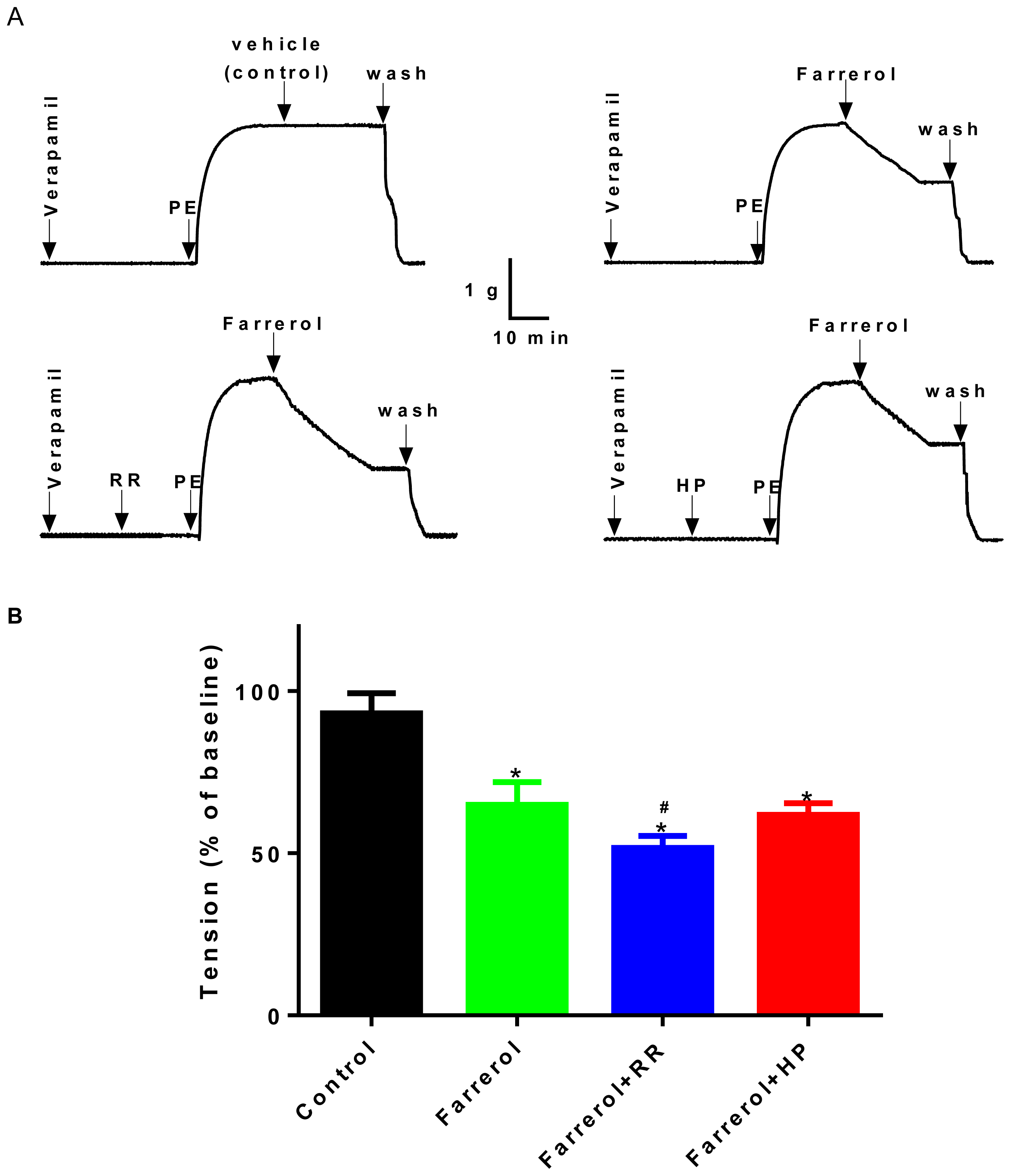
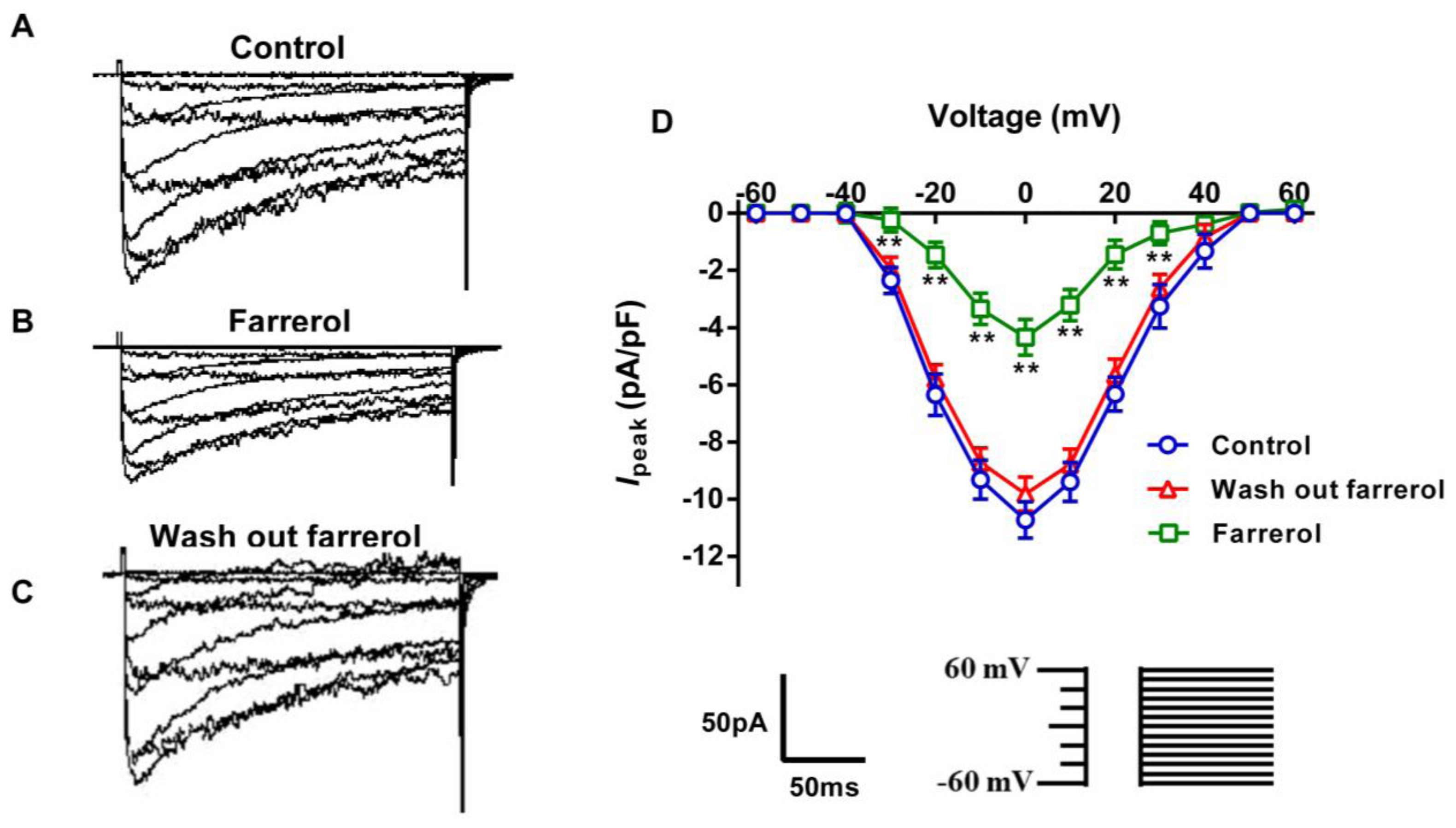
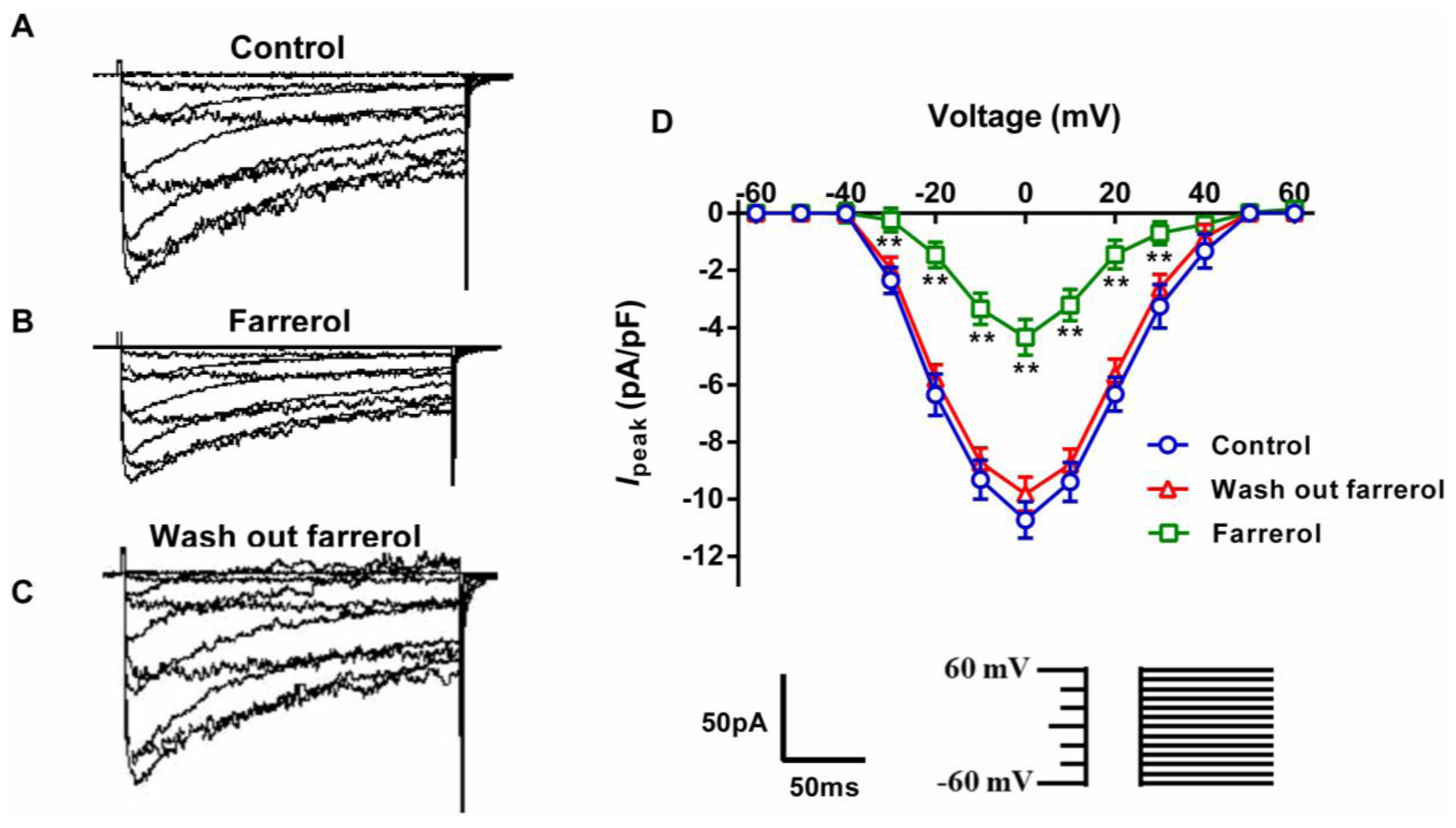
2.1.5. Effects of Farrerol on [Ca2+]ex Influx Current through L-Type Voltage-Gated Ca2+ Channels (LVGC) in Rat Aortic VSMCs
2.2. Discussion
3. Experimental Section
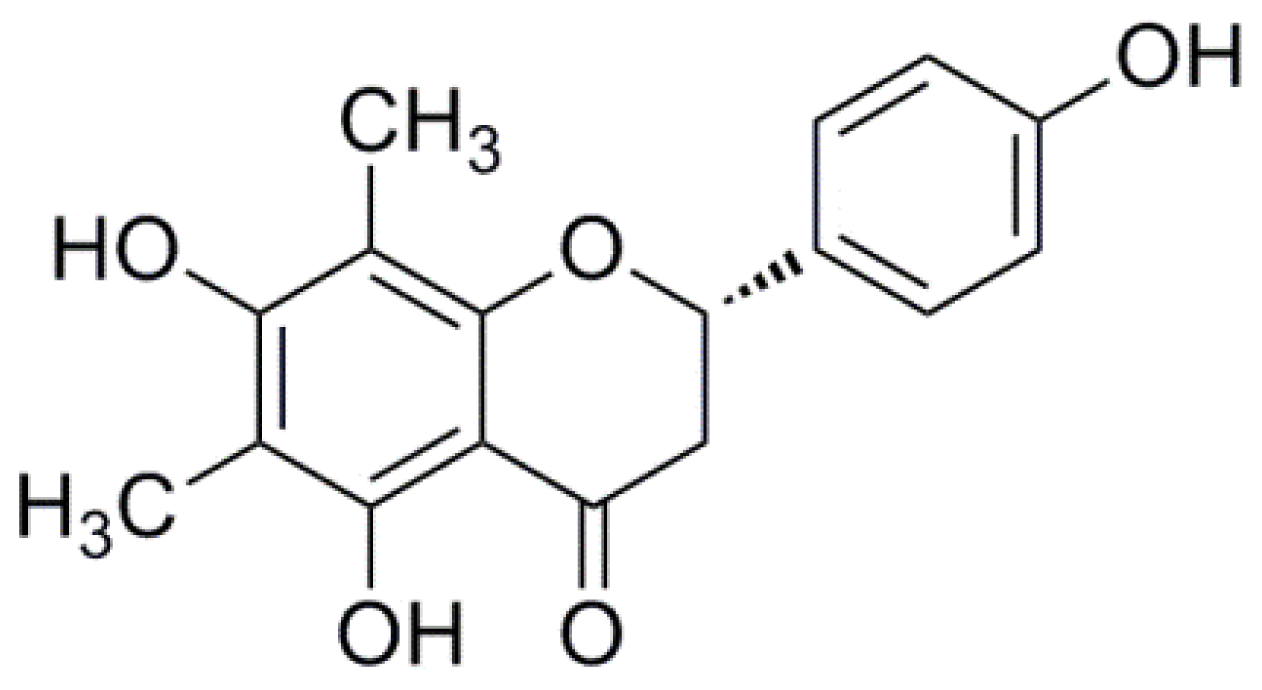
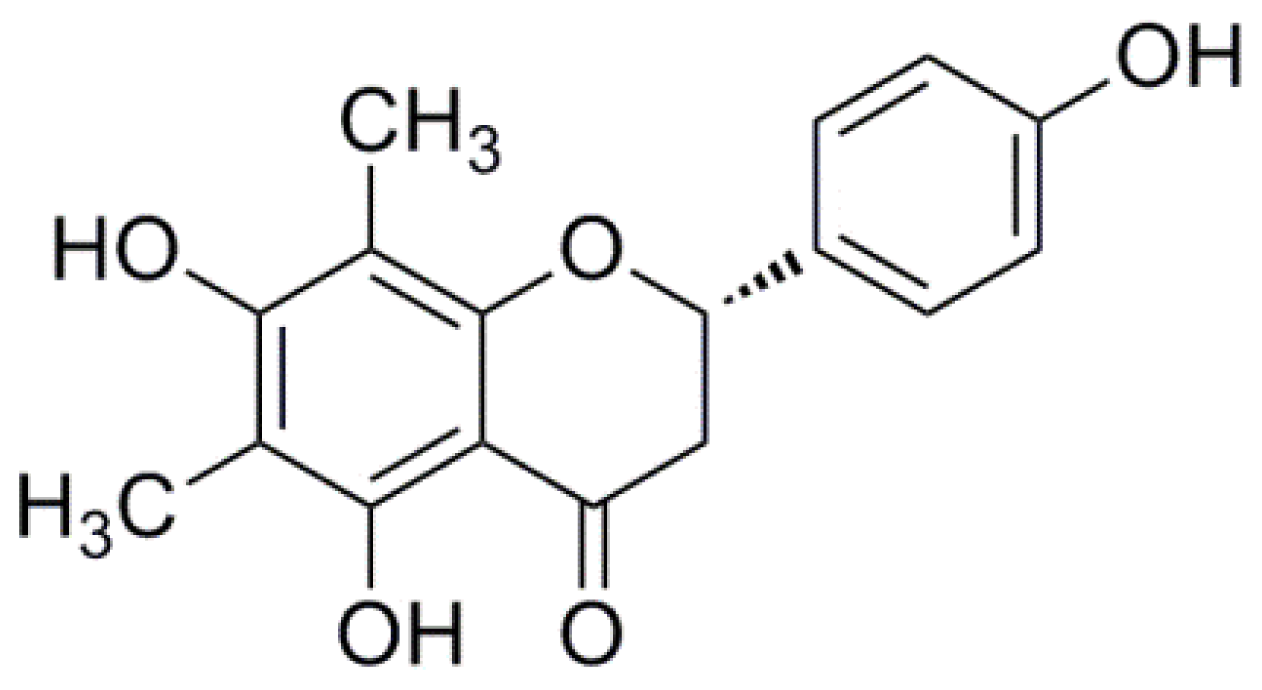
3.1. Chemicals and Drugs
3.2. Animals
3.3. Aorta Preparations and General Protocols
3.3.1. Effect of Farrerol on KCl- and PE-Induced Contraction of Rat Aortic Rings
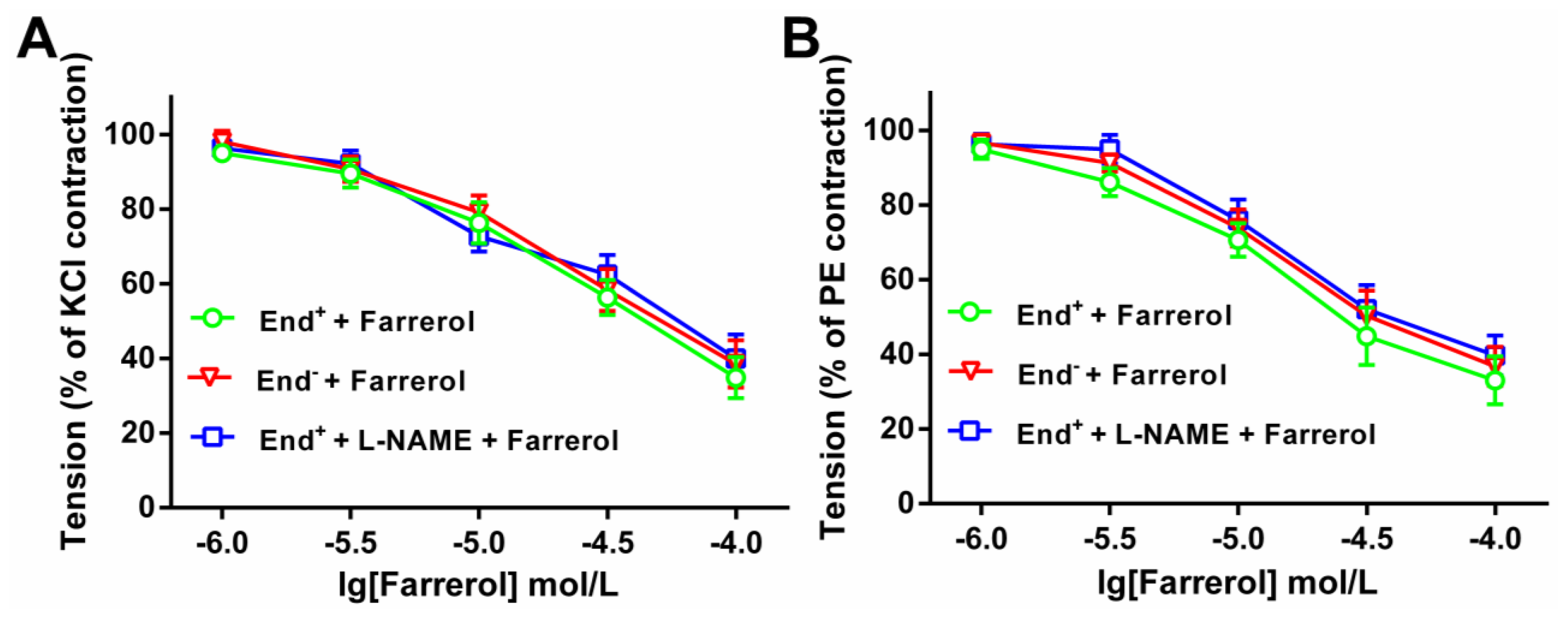
3.3.2. Effect of Farrerol on the Endothelium-Denuded Aortic Rings Contraction Induced by [Ca2+]in Release and [Ca2+]ex Influx
3.4. VSMCs Preparation
3.4.1. [Ca2+]in Measurements
3.4.2. Whole-Cell Patch Clamp Recordings
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsConceived and designed the experiments: Xiaojiang Qin and Qingshan Li. Performed the experiments: Xiaojiang Qin and Xiaomin Hou. Analyzed the data: Xiaojiang Qin, Jianmin Zhi and Mingsheng Zhang. Contributed reagents/materials/analysis tools: Lingge Han and Mingsheng Zhang. Wrote the paper: Xiaojiang Qin, Xiaomin Hou and Taigang Liang.
References
- Yang, Z.J.; Liu, J.; Ge, J.P.; Chen, L.; Zhao, Z.G.; Yang, W.Y. Prevalence of cardiovascular disease risk factor in the Chinese population: The 2007–2008 china national diabetes and metabolic disorders study. Eur. Heart J 2012, 33, 213–220. [Google Scholar]
- Gao, Y.; Chen, G.; Tian, H.; Lin, L.; Lu, J.; Weng, J.; Jia, W.; Ji, L.; Xiao, J.; Zhou, Z. Prevalence of hypertension in China: A cross-sectional study. PLoS One 2013. [Google Scholar] [CrossRef]
- Wang, L.; Kong, L.; Wu, F.; Bai, Y.; Burton, R. Preventing chronic diseases in China. Lancet 2005, 366, 1821–1824. [Google Scholar]
- Kearney, P.M.; Whelton, M.; Reynolds, K.; Muntner, P.; Whelton, P.K.; He, J. Global burden of hypertension: Analysis of worldwide data. Lancet 2005, 365, 217–223. [Google Scholar]
- Ezzati, M.; Lopez, A.D.; Rodgers, A.; Vander Hoorn, S.; Murray, C.J. Comparative risk assessment collaborating group. Selected major risk factors and global and regional burden of disease. Lancet 2002, 360, 1347–1360. [Google Scholar]
- Neal, B.; MacMahon, S.; Chapman, N. Blood pressure lowering treatment trialists’ collaboration. Effects of ACE inhibitors, calcium antagonists, and other blood-pressure-lowering drugs: Results of prospectively designed overviews of randomised trials. Lancet 2000, 356, 1955–1964. [Google Scholar]
- Curb, J.D.; Borhani, N.O.; Blaszkowski, T.P.; Zimbaldi, N.; Fotiu, S.; Williams, W. Long-term surveillance for adverse effects of antihypertensive drugs. JAMA 1985, 253, 3263–3268. [Google Scholar]
- Düsing, R.; Weisser, B.; Mengden, T.; Vetter, H. Changes in antihypertensive therapy-the role of adverse effects and compliance. Blood Press 1998, 7, 313–315. [Google Scholar]
- Husserl, F.E.; Messerli, F.H. Adverse effects of antihypertensive drugs. Drugs 1981, 22, 188–210. [Google Scholar]
- Waeber, B. Fixed low-dose combination therapy for hypertension. Curr. Hypertens. Rep 2002, 4, 298–306. [Google Scholar]
- Harapu, C.; Miron, A.; Cuciureanu, M.; Cuciureanu, R. Flavonoids-bioactive compounds in fruits juice. Rev. Med. Chir. Soc. Med. Nat. Iasi 2010, 114, 1209–1214. [Google Scholar]
- Chen, Y.; Fu, D.; He, Y.; Fu, X.; Xu, Y.; Liu, Y.; Feng, X.; Zhang, T.; Wang, W. Effects of Chinese herbal medicine Yiqi Huaju Formula on hypertensive patients with metabolic syndrome: A randomized, placebo-controlled trial. J. Integr. Med 2013, 11, 184–194. [Google Scholar]
- Wang, G.J.; Shan, J.; Pang, P.; Yang, M.; Chou, C.J.; Chen, C.F. The vasorelaxing action of rutaecarpine: Direct paradoxical effects on intracellular calcium concentration of vascular smooth muscle and endothelial cells. J. Pharmacol. Exp. Ther 1996, 276, 1016–1021. [Google Scholar]
- Crow, J.M. Microbiome: That healthy gut feeling. Nature 2011, 480, S88–S89. [Google Scholar]
- Xiong, X.; Yang, X.; Liu, Y.; Zhang, Y.; Wang, P.; Wang, J. Chinese herbal formulas for treating hypertension in traditional Chinese medicine: Perspective of modern science. Hypertens. Res 2013, 36, 570–579. [Google Scholar]
- Hertog, M.; Sweetnam, P.M.; Fehily, A.M.; Elwood, P.C.; Kromhout, D. Antioxidant flavonols and ischemic heart disease in a Welsh population of men: The caerphilly study. Am. J. Clin. Nutr 1997, 65, 1489–1494. [Google Scholar]
- Perez-Vizcaino, F.; Duarte, J. Flavonols and cardiovascular disease. Mol. Aspects. Med 2010, 31, 478–494. [Google Scholar]
- Chan, E.C.; Pannangpetch, P.; Woodman, O.L. Relaxation to flavones and flavonols in rat isolated thoracic aorta: Mechanism of action and structure-activity relationships. J. Cardiovasc. Pharmacol 2000, 35, 326–333. [Google Scholar]
- Sonia de, P.-T.; Diego, A.M.; Cristina, G.-V. Flavanols and anthocyanins in cardiovascular health: A review of current evidence. Int. J. Mol. Sci 2010, 11, 1679–1703. [Google Scholar]
- Knekt, P.; Jarvinen, R.; Reunanen, A.; Maatela, J. Flavonoid intake and coronary mortality in Finland: A cohort study. Br. Med. J 1996, 312, 478–481. [Google Scholar]
- Perez-Vizcaino, F.; Duarte, J.; Jimenez, R.; Santos-Buelga, C.; Osuna, A. Antihypertensive effects of the flavonoid quercetin. Pharmacol. Rep 2009, 61, 67–75. [Google Scholar]
- Ajay, M.; Gilani, A.U.; Mustafa, M.R. Effects of flavonoids on vascular smooth muscle of the isolated rat thoracic aorta. Life Sci 2003, 74, 603–612. [Google Scholar]
- Xia, E.-Q.; Deng, G.-F.; Guo, Y.-J.; Li, H.-B. Biological activities of polyphenols from grapes. Int. J. Mol. Sci 2010, 11, 622–646. [Google Scholar]
- Herrera, M.D.; Zarzuelo, A.; Jiménez, J.; Marhuenda, E.; Duarte, J. Effects of flavonoids on rat aortic smooth muscle contractility: Structure-activity relationships. Gen. Pharmacol 1996, 27, 273–277. [Google Scholar]
- Cazarolli, L.H.; Zanatta, L.; Alberton, E.H.; Figueiredo, B.; Reis, M.S.; Folador, P.; Damazio, R.G.; Pizzolatti, M.G.; Silva, B.; Mena, F.R. Flavonoids: Prospective drug candidates. Mini Rev. Med. Chem 2008, 8, 1429–1440. [Google Scholar]
- Gonçalves, R.L.; Lugnier, C.; Keravis, T.; Lopes, M.J.; Fantini, F.A.; Schmitt, M.; Cortes, S.F.; Lemos, V.S. The flavonoid dioclein is a selective inhibitor of cyclic nucleotide phosphodiesterase type 1 (PDE1) and a cGMP-dependent protein kinase (PKG) vasorelaxant in human vascular tissue. Eur. J. Pharmacol 2009, 620, 78–83. [Google Scholar]
- Li, J.K.; Ge, R.; Tang, L.; Li, Q.S. Protective effects of farrerol against hydrogen peroxide-induced apoptosis in human endothelium-derived EA. hy926 Cells. Can. J. Physiol. Pharmacol 2013, 91, 733–740. [Google Scholar]
- Li, Q.Y.; Chen, L.; Zhu, Y.H.; Zhang, M.; Wang, Y.P.; Wang, M.W. Involvement of estrogen receptor-β in farrerol inhibition of rat thoracic aorta vascular smooth muscle cell proliferation. Acta Pharmacol. Sin 2011, 32, 433–440. [Google Scholar]
- Shi, L.; Feng, X.E.; Cui, J.R.; Fang, L.H.; Du, G.H.; Li, Q.S. Synthesis and biological activity of flavanone derivatives. Bioorg. Med. Chem. Lett 2010, 20, 5466–5468. [Google Scholar]
- Karaki, H.; Ozaki, H.; Hori, M.; Mitsui-Saito, M.; Amano, K.I.; Harada, K.I.; Miyamoto, S.; Nakazawa, H.; Won, K.J.; Sato, K. Calcium movements, distribution, and functions in smooth muscle. Pharmacol. Rev 1997, 49, 157–230. [Google Scholar]
- Jones, R.D.; Pugh, P.J.; Jones, T.H.; Channer, K.S. The vasodilatory action of testosterone: A potassium-channel opening or a calcium antagonistic action? Br. J. Pharmacol 2003, 138, 733–744. [Google Scholar]
- Lee, K.; Park, G.; Ham, I.; Yang, G.; Lee, M.; Bu, Y.; Kim, H.; Choi, H.Y. Vasorelaxant effect of osterici radix ethanol extract on rat aortic rings. Evid-Based Complement. Altern 2013. [Google Scholar] [CrossRef]
- Li, X.; Chen, G.P.; Li, L.; Wang, K.J.; Zhang, B.Q.; Hu, S.J. Dual effects of sodium aescinate on vascular tension in rat thoracic aorta. Microvasc. Res 2010, 79, 63–69. [Google Scholar]
- Somlyo, A.P.; Himpens, B. Cell calcium and its regulation in smooth muscle. FASEB J 1989, 3, 2266–2276. [Google Scholar]
- Fleischer, S.; Inui, M. Biochemistry and biophysics of excitation-contraction coupling. Annu. Rev. Biophys. Biophys. Chem 1989, 18, 333–364. [Google Scholar]
- Stull, J.T.; Gallagher, P.J.; Herring, B.P.; Kamm, K.E. Vascular smooth muscle contractile elements. Cellular regulation. Hypertension 1991, 17, 723–732. [Google Scholar]
- Rembold, C.M. Regulation of contraction and relaxation in arterial smooth muscle. Hypertension 1992, 20, 129–137. [Google Scholar]
- Buraei, Z.; Yang, J. The β subunit of voltage-gated Ca2+ channels. Physiol. Rev 2010, 90, 1461–1506. [Google Scholar]
- Xia, M.; Qian, L.; Zhou, X.; Gao, Q.; Bruce, I.C.; Xia, Q. Endothelium-independent relaxation and contraction of rat aorta induced by ethyl acetate extract from leaves of Morus alba (L.). J. Ethnopharmacol 2008, 120, 442–446. [Google Scholar]
- Karaki, H.; Weiss, G. Calcium channels in smooth muscle. Gastroenterology 1984, 87, 960–970. [Google Scholar]
- Dellis, O.; Dedos, S.G.; Tovey, S.C.; Dubel, S.J.; Taylor, C.W. Ca2+ entry through plasma membrane IP3 receptors. Science 2006, 313, 229–233. [Google Scholar]
- Wier, W.; Morgan, K. α1-Adrenergic signaling mechanisms in contraction of resistance arteries. Rev. Phys. Biochem. Pharmacol 2004, 150, 91–139. [Google Scholar]
- Hester, R.K. Effects of 2-nicotinamidoethyl nitrate on agonist-sensitive Ca++ release and Ca++ entry in rabbit aorta. J. Pharmacol. Exp. Ther 1985, 233, 100–111. [Google Scholar]
- Qian, L.B.; Wang, H.P.; Qiu, W.L.; Huang, H.; Bruce, I.C.; Xia, Q. Interleukin-2 protects against endothelial dysfunction induced by high glucose levels in rats. Vasc. Pharmacol 2006, 45, 374–382. [Google Scholar]
- Li, H.; Chai, Q.; Gutterman, D.D.; Liu, Y. Elevated glucose impairs cAMP-mediated dilation by reducing Kv channel activity in rat small coronary smooth muscle cells. Am. J. Physiol. Heart Circ. Physiol 2003, 285, H1213–H1219. [Google Scholar]
- Niu, L.G.; Zhang, M.S.; Liu, Y.; Xue, W.X.; Liu, D.B.; Zhang, J.; Liang, Y.Q. Vasorelaxant effect of taurine is diminished by tetraethylammonium in rat isolated arteries. Eur. J. Pharmacol 2008, 580, 169–174. [Google Scholar]
- Franceschelli, S.; Ferrone, A.; Pesce, M.; Riccioni, G.; Speranza, L. Biological functional relevance of asymmetric dimethylarginine (adma) in cardiovascular disease. Int. J. Mol. Sci 2013, 14, 24412–24421. [Google Scholar]
- Klöckner, U.; Isenberg, G. Intracellular pH modulates the availability of vascular L-type Ca2+ channels. J. Gen. Physiol 1994, 103, 647–663. [Google Scholar]
- Kim, S.H.; Kang, K.W.; Kim, K.W.; Kim, N.D. Procyanidins in crataegus extract evoke endothelium-dependent vasorelaxation in rat aorta. Life Sci 2000, 67, 121–131. [Google Scholar]
- Maggi, C.A.; Patacchini, R.; Perretti, F.; Tramontana, M.; Manzini, S.; Geppetti, P.; Santicioli, P. Sensory nerves, vascular endothelium and neurogenic relaxation of the guinea-pig isolated pulmonary artery. Naunyn. Schmiedebergs. Arch. Pharmacol 1990, 342, 78–84. [Google Scholar]
- Karashima, Y.; Prenen, J.; Talavera, K.; Janssens, A.; Voets, T.; Nilius, B. Agonist-induced changes in ca2+ permeation through the nociceptor cation channel trpa1. Biophys. J 2010, 98, 773–783. [Google Scholar]
- Mohanty, M.J.; Li, X. Stretch-induced Ca2+ release via an IP3-insensitive Ca2+ channel. Am. J. Physiol. Cell Physiol 2002, 283, C456–C462. [Google Scholar]

© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Qin, X.; Hou, X.; Zhang, M.; Liang, T.; Zhi, J.; Han, L.; Li, Q. Relaxation of Rat Aorta by Farrerol Correlates with Potency to Reduce Intracellular Calcium of VSMCs. Int. J. Mol. Sci. 2014, 15, 6641-6656. https://doi.org/10.3390/ijms15046641
Qin X, Hou X, Zhang M, Liang T, Zhi J, Han L, Li Q. Relaxation of Rat Aorta by Farrerol Correlates with Potency to Reduce Intracellular Calcium of VSMCs. International Journal of Molecular Sciences. 2014; 15(4):6641-6656. https://doi.org/10.3390/ijms15046641
Chicago/Turabian StyleQin, Xiaojiang, Xiaomin Hou, Mingsheng Zhang, Taigang Liang, Jianmin Zhi, Lingge Han, and Qingshan Li. 2014. "Relaxation of Rat Aorta by Farrerol Correlates with Potency to Reduce Intracellular Calcium of VSMCs" International Journal of Molecular Sciences 15, no. 4: 6641-6656. https://doi.org/10.3390/ijms15046641