Effect of Nitrate on Nodule and Root Growth of Soybean (Glycine max (L.) Merr.)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
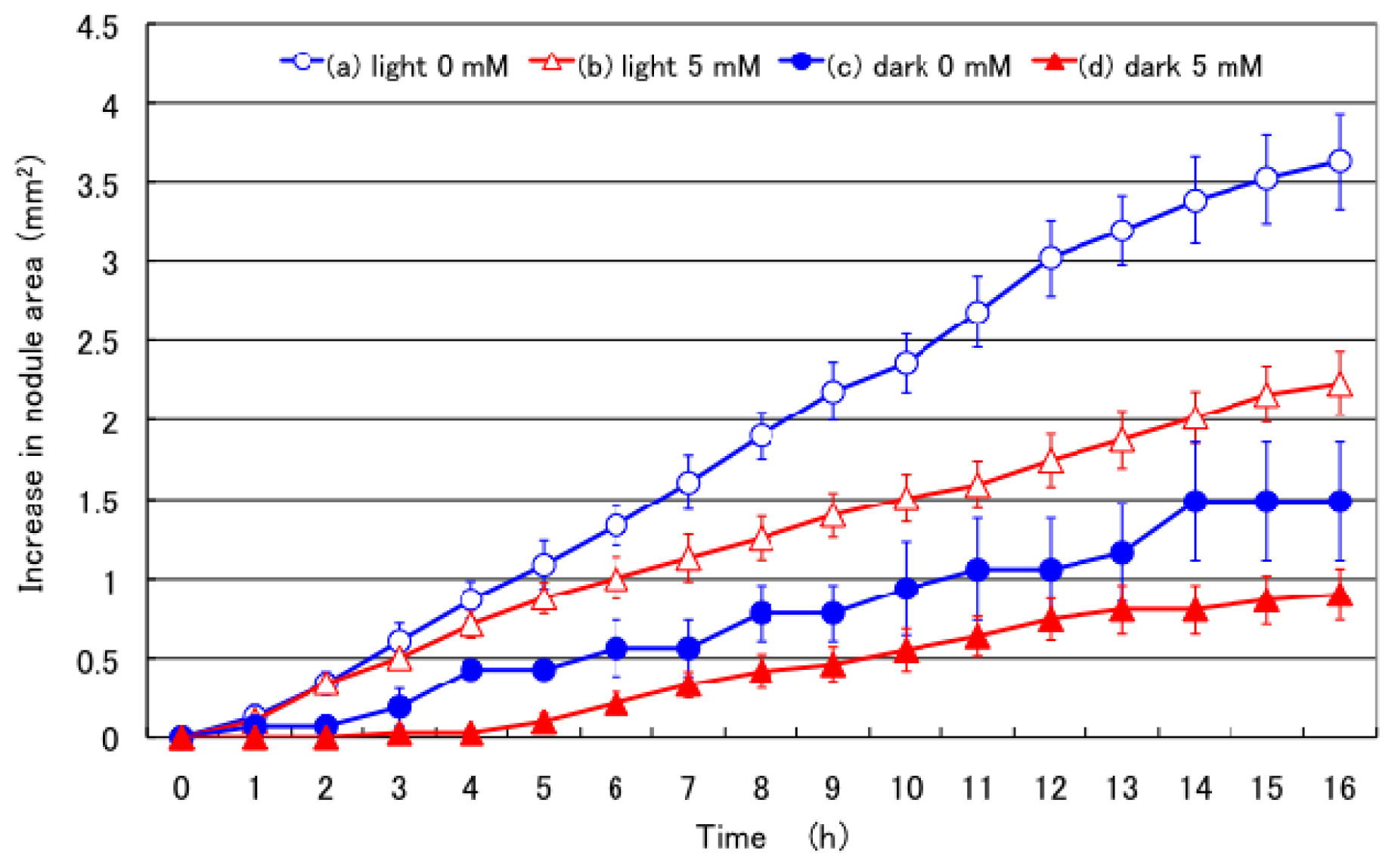
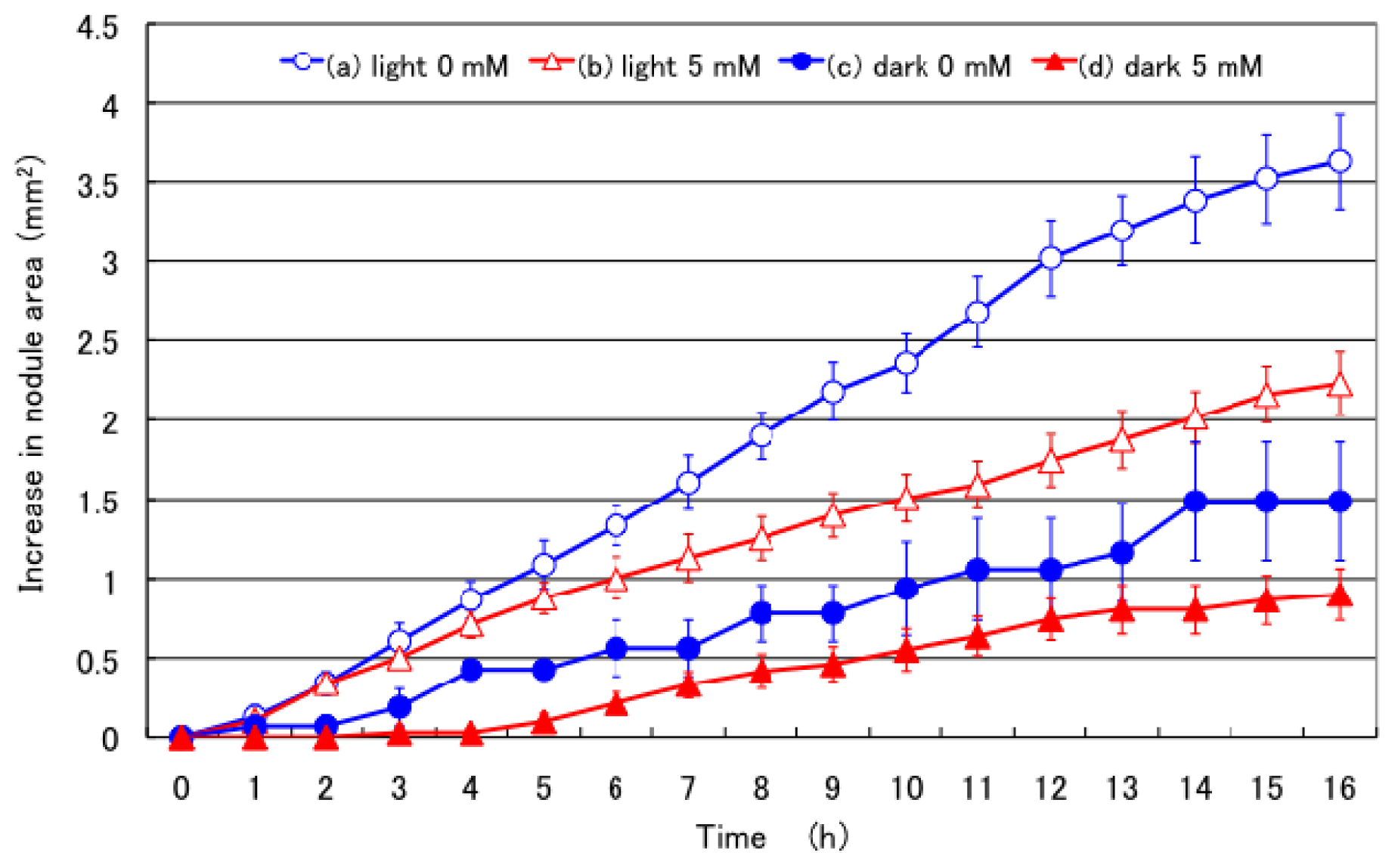
2.1. Effect of 5 mM Nitrate Supply in the Culture Solution on Nodule Growth under Light and Dark Conditions
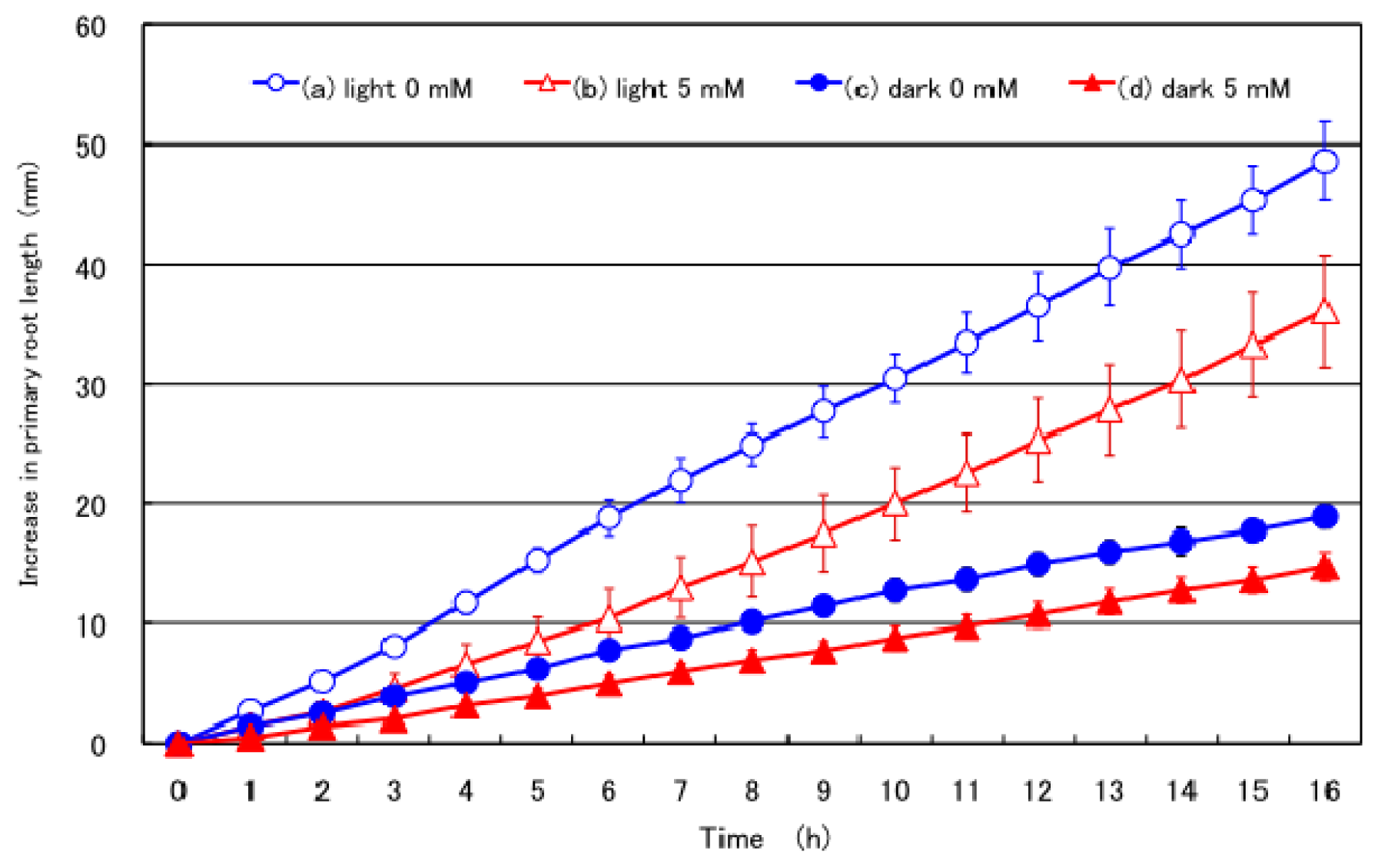
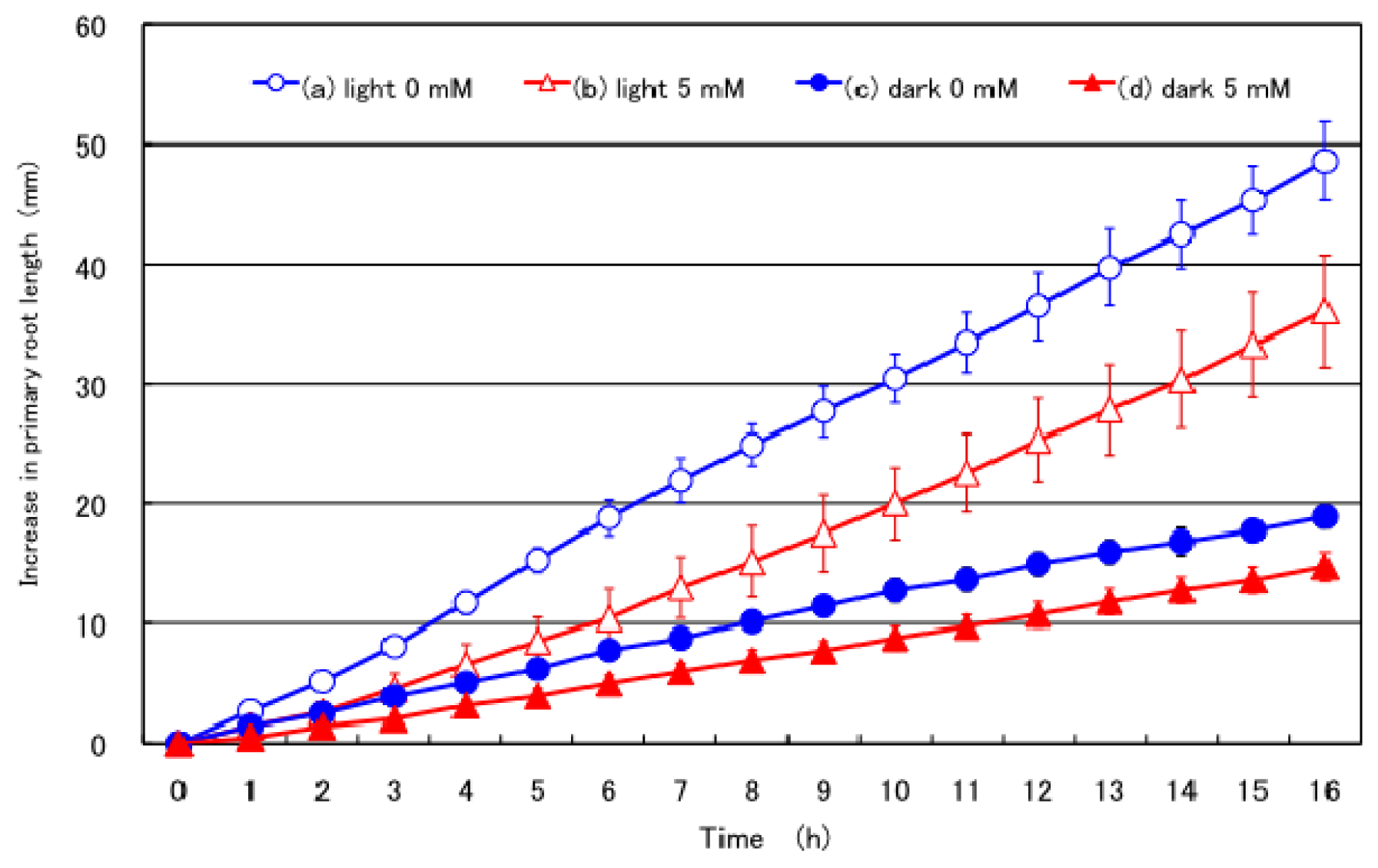
2.2. Effect of Nitrate Supply to the Culture Solution on the Growth of the Primary Root under Light and Dark Conditions
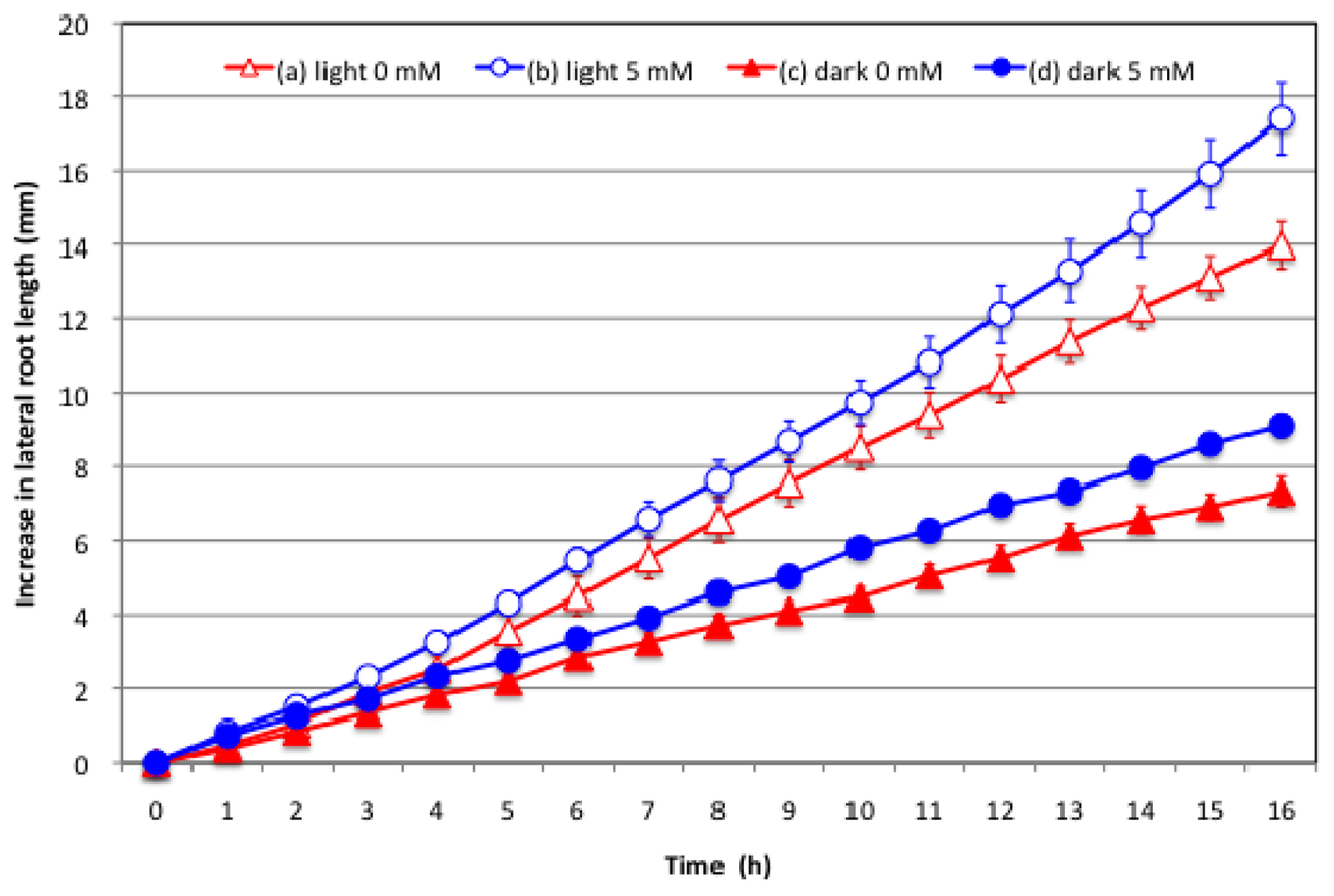
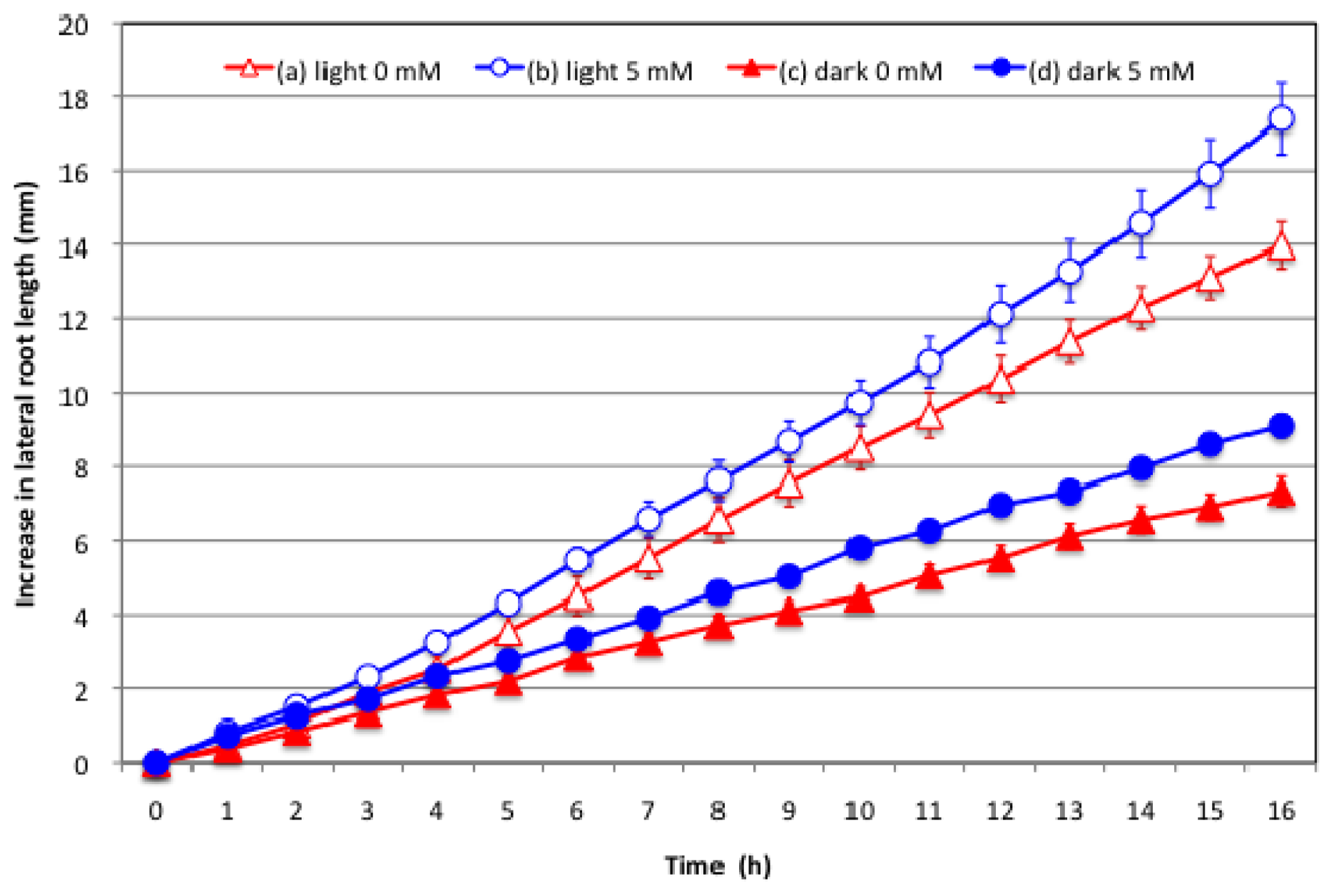
2.3. Effect of Nitrate Supply to the Culture Solution on the Growth of Lateral Roots under Light and Dark Conditions
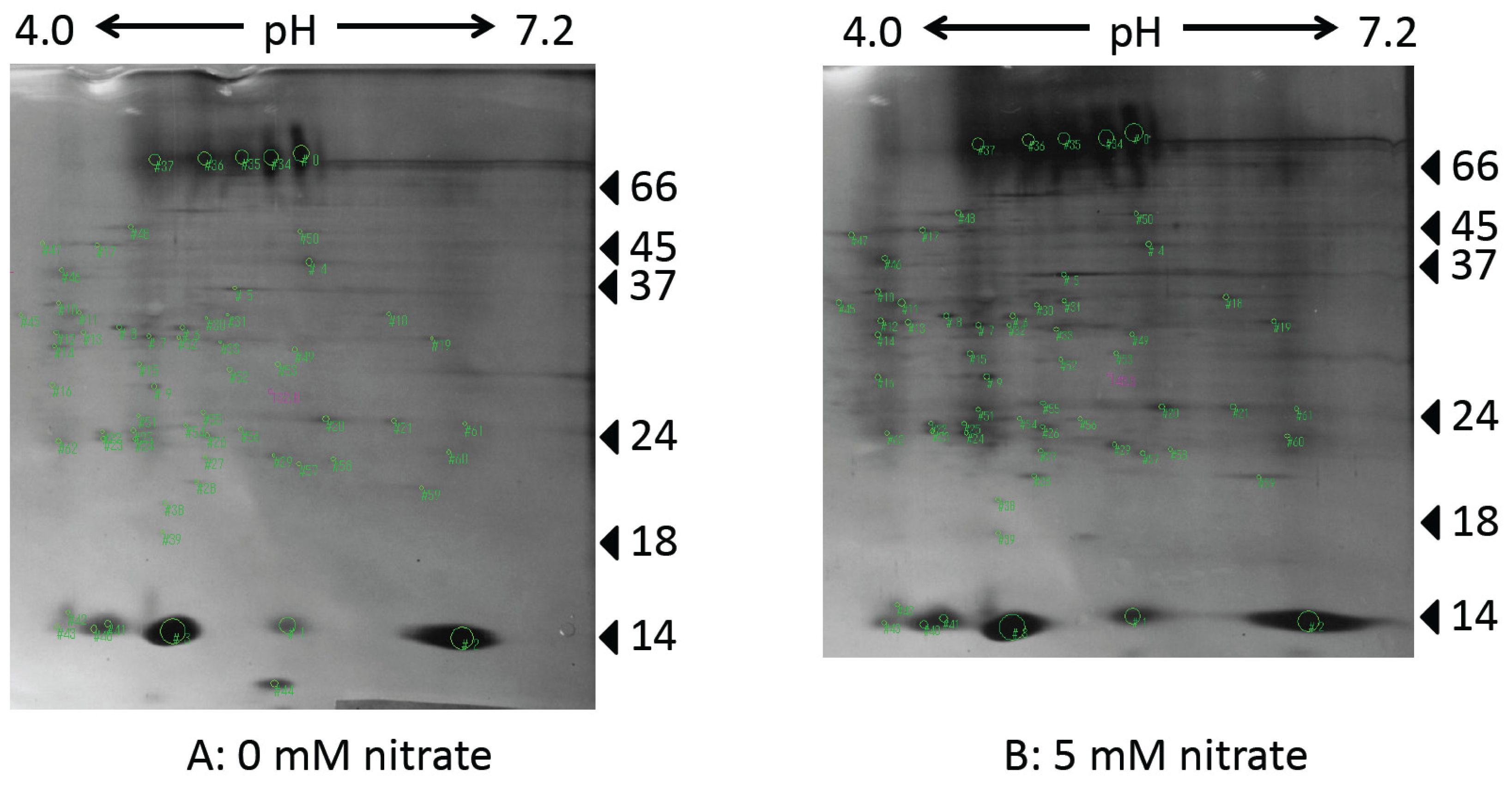
2.4. Effect of Supplying Nitrate to the Culture Solution on Nodule Protein
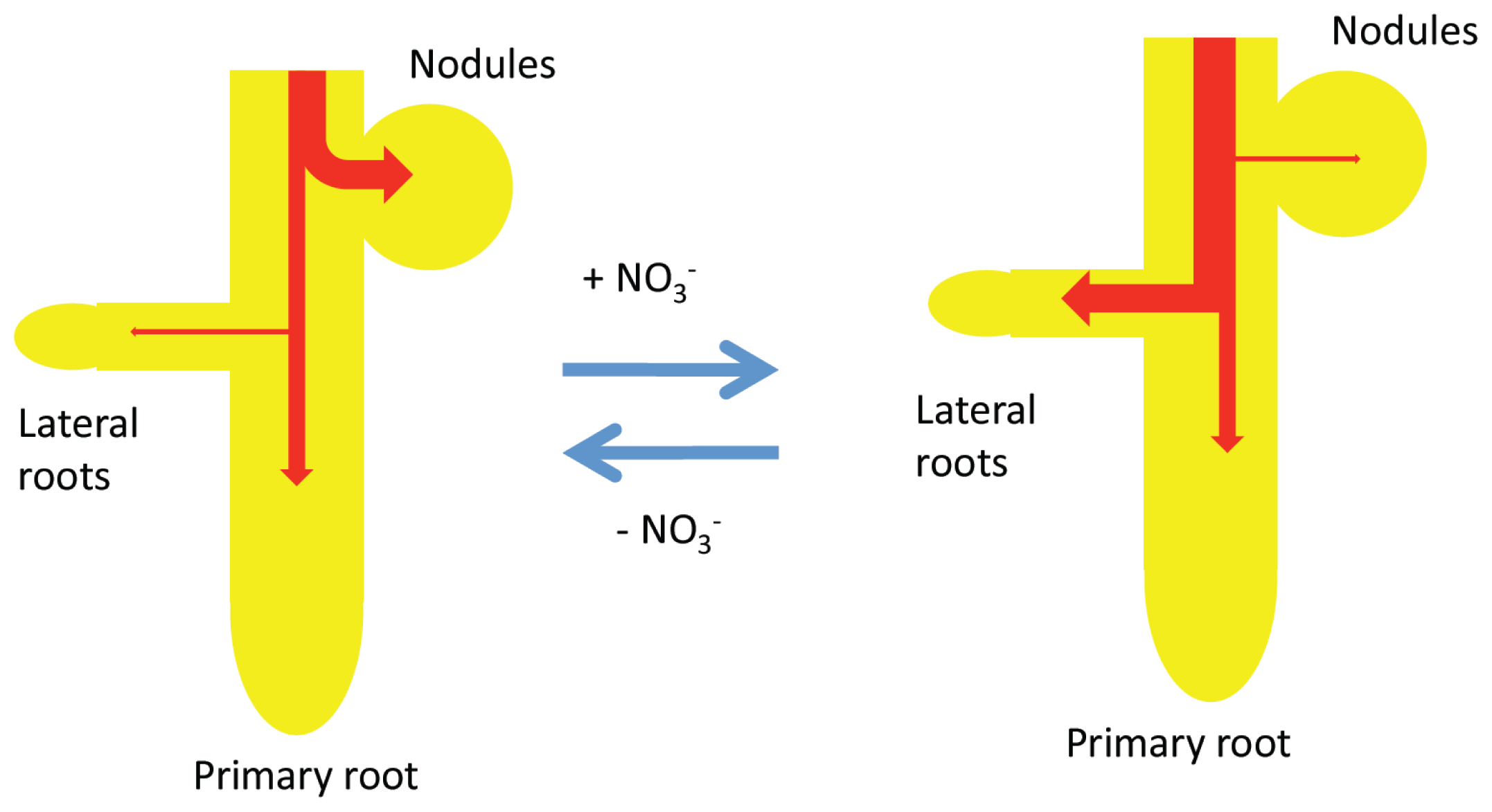
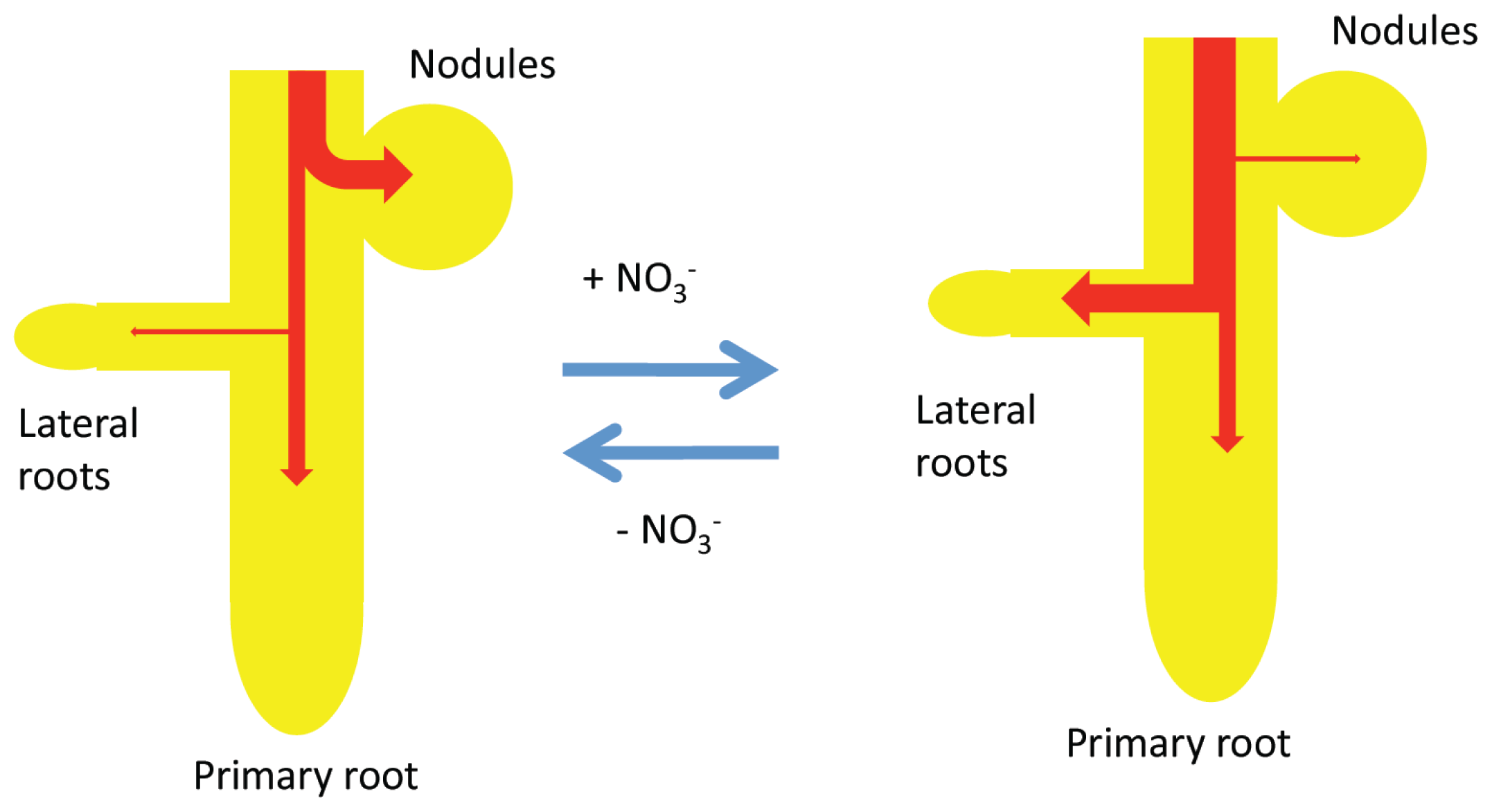
2.5. A Model for the Effects of a Short-Term Supply of Nitrate on the Growth of Nodules, Primary Roots, and Lateral Roots under Day/Night Conditions
3. Experimental Section
3.1. Plant Material and Growth Conditions
3.2. Monitoring Nodule Growth by the Addition of 5 mM Nitrate under Light or Dark Conditions
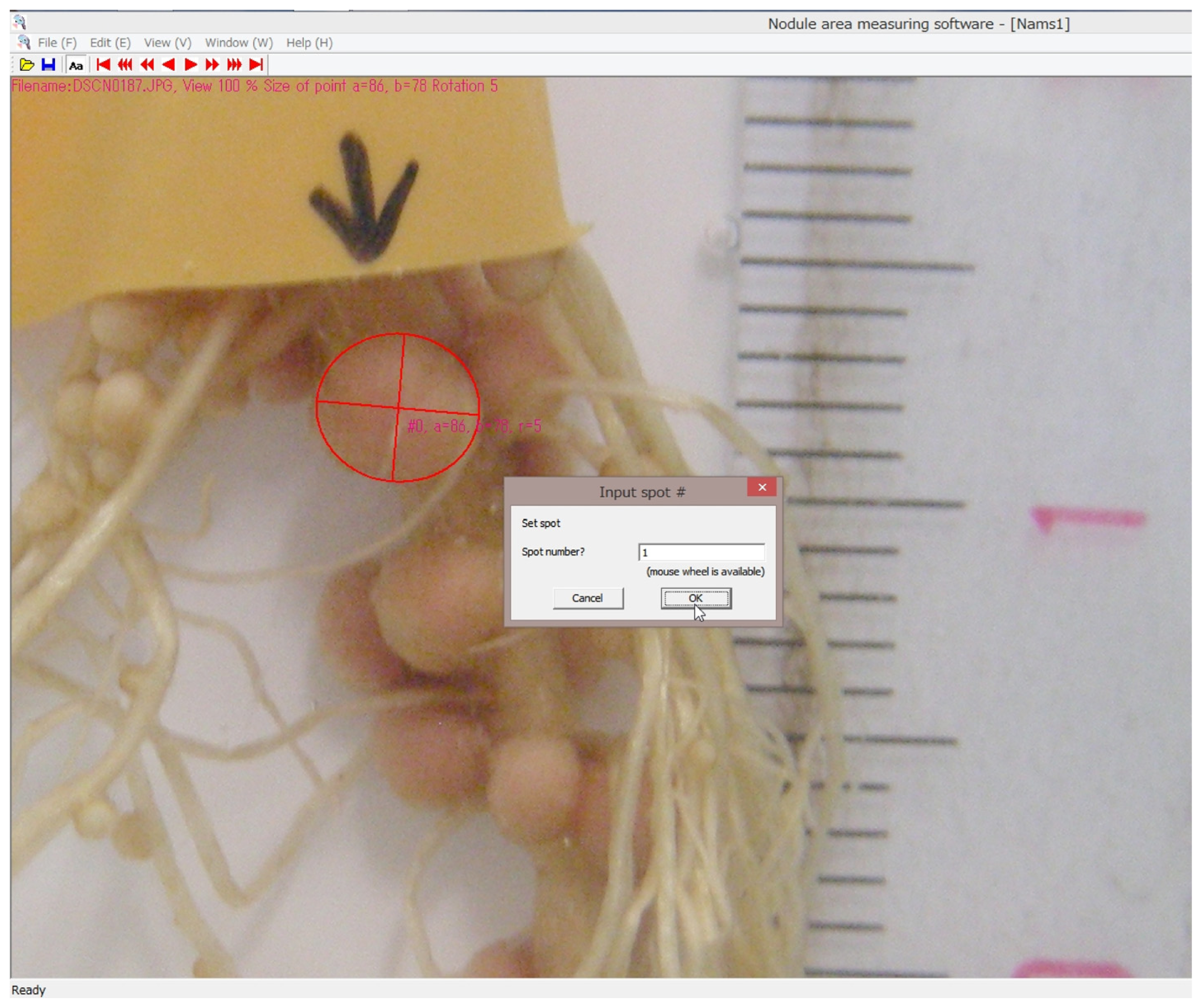
3.3. Calculation of Nodule Growth by a Computer Program
3.4. Monitoring Root Growth by the Addition of 5 mM Nitrate under Light or Dark Conditions
3.5. 2D-PAGE Analysis of Nodule Protein with or without Addition of 5 mM Nitrate
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Ohyama, T.; Minagawa, R.; Ishikawa, S.; Yamamoto, M.; Hung, N.V.P.; Ohtake, N.; Sueyoshi, K.; Sato, T.; Nagumo, Y.; Takahashi, Y. Soybean Seed Production and Nitrogen Nutrition. In A Comprehensive Survey of International Soybean Research-Genetics, Physiology, Agronomy and Nitrogen Relationships; Board, J.E., Ed.; InTech: Rijeka, Croatia, 2013; pp. 115–157. [Google Scholar]
- Harper, J.E. Soil and symbiotic nitrogen requirements for optimum soybean production. Crop Sci. 1974, 14, 255–260. [Google Scholar]
- Gibson, A.H.; Harper, J.E. Nitrate effect on nodulation of soybean by Bradyrhizobium japonicum. Crop Sci. 1985, 25, 497–501. [Google Scholar]
- Imsande, J. Inhibition of nodule development in soybean by nitrate or reduced nitrogen. J. Exp. Bot. 1986, 37, 348–355. [Google Scholar]
- Ohyama, T.; Fujikake, H.; Yashima, H.; Tanabata, S.; Ishikawa, S.; Sato, T.; Nishiwaki, T.; Ohtake, N.; Sueyoshi, K.; Ishii, S.; et al. Effect of Nitrate on Nodulation and Nitrogen Fixation of Soyeban. In Soybean Physiology and Biochemistry; El-Shemy, H.A., Ed.; InTech: Rijeka, Croatia, 2012; pp. 333–364. [Google Scholar]
- Streeter, J.G. Inhibition of legume nodule formation and N2 fixation by nitrate. CRC Crit. Rev. Plant Sci. 1988, 7, 1–23. [Google Scholar]
- Harper, J.E. Nitrogen Metabolism. In Soybeans: Improvement, Production and Uses, 2nd ed.; Wilcox, J.R., Ed.; American Society of Agronomy, Inc.-Crop Science Society of America, Inc.-Soil Science Society of America, Inc.: Madison, WI, USA, 1987; pp. 497–533. [Google Scholar]
- Mizukoshi, K.; Nishiwaki, T.; Ohtake, N.; Minagawa, R.; Ikarashi, T.; Ohyama, T. Nitrate transport pathway into soybean nodules traced by tungstate and 15NO3−. Soil Sci. Plant Nutr 1995, 41, 75–88. [Google Scholar]
- Ohyama, T.; Ito, S.; Nagumo, Y.; Ohtake, N.; Sueyoshi, K.; Takahashi, Y.; Sato, T. Symbiotic Nitrogen Fixation and Its Assimilation in Soybean. In Nitrogen Assimilation in Plants; Research Signpost: Kerala, India, 2010; pp. 175–203. [Google Scholar]
- Caetano-Anollés, G.; Gresshoff, P. Plant genetic control of nodulation. Annu. Rev. Microbiol. 1991, 45, 345–382. [Google Scholar]
- Reid, D.E.; Ferguson, B.J.; Hayashi, S.; Lin, Y.-H.; Gresshoff, P.M. Molecular mechanisms controlling legume autoregulation of nodulation. Ann. Bot. 2011, 108, 789–795. [Google Scholar]
- Soyano, T.; Kawaguchi, M. Systemic Regulation of Root Nodule Formation. In Advances in Biology and Ecology of Nitrogen Fixation; Ohyama, T., Ed.; InTech: Rijeka, Croatia, 2014; pp. 89–109. [Google Scholar]
- Akao, S.; Kouchi, H. A supernodulation mutant isolated from soybean cultivar Enrei. Soil Sci. Plant Nutr 1992, 38, 183–187. [Google Scholar]
- Delves, A.C.; Mathews, A.; Day, D.A.; Carter, A.S.; Carroll, B.J.; Gresshoff, P. Regulation of the soybean-Rhizobium nodule symbiosis by shoot and root factors. Plant Physiol. 1986, 82, 588–590. [Google Scholar]
- Gremaud, M.F.; Harper, J.E. Selection and initial characterization of partially nitrate tolerant nodulation mutants of soybean. Plant Physiol. 1989, 89, 169–173. [Google Scholar]
- Ohyama, T.; Nicholas, J.C.; Harper, J.E. Assimilation of 15N2 and 15NO3− by partially nitrate-tolerant nodulation mutants of soybean. J. Exp. Bot. 1993, 44, 1739–1747. [Google Scholar]
- Reid, D.E.; Ferguson, B.J.; Gresshoff, P.M. Inoculation- and nitrate-induced CLE peptides of soybean control NARK-dependent nodule formation Molecular mechanisms controlling legume autoregulation of nodulation. MPMI 2011, 24, 606–618. [Google Scholar]
- Okamoto, S.; Ohnishi, E.; Sato, S.; Takahashi, H.; Nakazono, M.; Tabata, S.; Kawaguchi, M. Nod factor/nitrate-inducible CLE genes that drive systemic regulation of nodulation. Plant Cell Physiol. 2009, 50, 67–77. [Google Scholar]
- Reid, D.E.; Li, D.; Ferguson, B.J.; Gresshoff, P.M. Structure-function analysis of the GmRIC1 signal peptide and CLE domain required for nodulation control in soybean. J. Exp. Bot. 2013, 64, 1575–1585. [Google Scholar]
- Davidson, I.A.; Robson, M.J. Effect of contrasting patterns of nitrate application on the nitrate uptake N2-fixation nodulation and growth of white clover. Ann. Bot. 1986, 57, 331–338. [Google Scholar]
- Harper, J.E.; Gibson, A.H. Differential nodulation tolerance to nitrate among legume species. Crop Sci. 1984, 24, 797–801. [Google Scholar]
- Yashima, H.; Fujikake, H.; Sato, T.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Systemic and local effects of long term application of nitrate on nodule growth and N2 fixation in soybean (Glycine max [L] Merr). Soil Sci. Plant Nutr. 2003, 49, 825–834. [Google Scholar]
- Yashima, H.; Fujikake, H.; Yamazaki, A.; Ito, S.; Sato, T.; Tewari, K.; Ohtake, N.; Sueyoshi, K.; Takahashi, Y.; Ohyama, T. Long-term effect of nitrate application from lower part of roots on nodulation and N2 fixation in upper part of roots of soybean (Glycine max [L] Merr) in two-layered pot experiment. Soil Sci. Plant Nutr. 2005, 51, 981–990. [Google Scholar]
- Carroll, B.J.; Mathews, A. Nitrate Inhibition of Nodulation in Legumes. In Molecular Biology of Symbiotic Nitrogen Fixation; CRC Press: Boca Raton, FL, USA, 1990; pp. 159–180. [Google Scholar]
- Fujikake, H.; Yamazaki, A.; Ohtake, N.; Sueyoshi, K.; Matsuhashi, S.; Ito, T.; Mizuniwa, C.; Kume, T.; Hashimoto, S.; Ishioka, N.S.; et al. Quick and reversible inhibition of soybean root nodule growth by nitrate involves a decrease in sucrose supply to nodules. J. Exp. Bot. 2003, 54, 1379–1388. [Google Scholar]
- Thorpe, M.R.; Walsh, K.B.; Minchin, P.E.H. Photoassimilate partitioning in nodulated soybean I 11C methodology. J. Exp. Bot. 1998, 49, 1805–1815. [Google Scholar]
- Vessey, J.K.; Walsh, K.B.; Layzell, D.B. Can a limitation in phloem supply to nodules account for the inhibitory effect of nitrate on nitrogenase activity in soybean? Physiol. Plant 1988, 74, 137–146. [Google Scholar]
- Neo, H.H.; Layzell, D.B. Phloem glutamine and the regulation of O2 diffusion in legume nodules. Plant Physiol. 1997, 113, 259–267. [Google Scholar]
- Bacanamwo, M.; Harper, J.E. The feedback mechanism of nitrate inhibition of nitrogenase activity in soybean may involve asparagines and/or products of its metabolism. Physiol. Plant 1997, 100, 371–377. [Google Scholar]
- Gordon, A.J.; Skot, L.; James, C.L.; Minchin, F.R. Short-term metabolic response of soybean root nodules to nitrate. J. Exp. Bot. 2002, 53, 423–428. [Google Scholar]
- Schuller, K.A.; Minchin, F.R.; Gresshoff, P. Nitrogenase activity and oxygen diffusion in nodules of soybean cv Bragg and a supernodulating mutant: Effect of nitrate. J. Exp. Bot. 1988, 39, 865–877. [Google Scholar]
- Fujikake, H.; Yashima, H.; Sato, T.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Rapid and reversible nitrate inhibition of nodule growth and N2 fixation activity in soybean (Glycine max (L) Merr). Soil Sci. Plant Nutr. 2002, 48, 211–217. [Google Scholar]
- Tanabata, S.; Tanabata, T.; Saito, A.; Tajima, S.; Watanabe, S.; Ishikawa, K.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Computational imaging analysis method for measuring size of nodule growth in soybean. Jpn. J. Soil Sci. Plant Nutr. 2014, 85, 43–47. [Google Scholar]
- Drew, M.C. Comparison of the effects of a localized supply of phosphate nitrate ammonium and potassium on the growth of the seminal root system and the shoot in barley. New Phytol. 1975, 75, 479–490. [Google Scholar]
- Fukai, H.; Tameda, S.; Masuda, H.; Tasaka, M. Lateral root formation is blocked by a gain-of-function mutation in the SOLITARY-ROOT/IAA14 gene of Arabidopsis. Plant J. 2002, 29, 153–168. [Google Scholar]
- Malamy, J.E.; Benfey, P.N. Down and out in Arabidopsis: The formation of lateral roots. Trends Plant Sci. 1997, 2, 390–396. [Google Scholar]
- Nodame: Nodule Area Measuring Software. Available online: http://phenotyping.image.coocan.jp/nodame/ (accessed on 11 March 2014).
- Ito, S.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Allocation of photosynthetic products in soybean during the early stages of nodule formation. Soil Sci. Plant Nutr. 2006, 52, 438–443. [Google Scholar]
- Ito, S.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Allocation of photosynthetic products in hypernodulation mutant of soybean NOD1–3 in the early stages of nodule formation. Bull. Fac. Agric. Niigata Univ. 2006, 59, 33–37. [Google Scholar]
- Oehrle, N.W.; Sarma, A.D.; Waters, J.K.; Emerich, D.W. Proteomic analysis of soybean nodule cytosol. Phytochemistry 2008, 69, 2426–2438. [Google Scholar]
- Appleby, C.A. Leghemoglobin. In The Biology of Nitrogen Fixation; Quispel, A., Ed.; North Holland Publishing: Amsterdam, The Netherlands, 1974; pp. 521–554. [Google Scholar]
- Appleby, C.A. Leghemoglobin and rhizobium respiration. Ann. Rev. Plant Physiol. 1984, 35, 443–478. [Google Scholar]
- Nishiwaki, T.; Sato, T.; Yashima, H.; Ikarashi, T.; Ohyama, T.; Harper, J.E.; Akao, S.; Kouchi, H. Changes in concentration of leghemoglobin components in hypernodulation mutants of soybean. Soil Sci. Plant Nutr. 1997, 43, 1091–1096. [Google Scholar]
- Sato, T.; Yashima, H.; Ohtake, N.; Sueyoshi, K.; Akao, S.; Ohyama, T. Possible Involvement of photosynthetic supply in changes of nodule characteristics of hypernodulating soybeans. Soil Sci. Plant Nutr. 1999, 45, 187–196. [Google Scholar]
- Bisseling, T.; van Den Bos, R.C.; van Kammen, A. The effect of ammonium nitrate on the synthesis of nitrogenase and the concentration of leghemoglobin in pea root nodules induced by Rhizomium leguminosarum. Biochem. Biophys. Acta 1978, 539, 1–11. [Google Scholar]
- Sprent, J.I.; James, E.K. Legume Evolution: Where do nodules and mycorrhizas fit in? Plant Physiol. 2007, 144, 575–581. [Google Scholar]
- Sano, N.; Permana, H.; Kumada, R.; Shinozaki, Y.; Tanabata, T.; Yamada, T.; Hirasawa, T.; Kanekatsu, M. Proteomic analysis of embryonic protein synthesized from long-lived mRNAs during germination of rice seeds. Plant Cell Physiol 2012, 53, 687–698. [Google Scholar]
- Polyans2d. Available online: http://www.kazusa.or.jp/polyans2d/ (accessed on 11 March 2014).
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Saito, A.; Tanabata, S.; Tanabata, T.; Tajima, S.; Ueno, M.; Ishikawa, S.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Effect of Nitrate on Nodule and Root Growth of Soybean (Glycine max (L.) Merr.). Int. J. Mol. Sci. 2014, 15, 4464-4480. https://doi.org/10.3390/ijms15034464
Saito A, Tanabata S, Tanabata T, Tajima S, Ueno M, Ishikawa S, Ohtake N, Sueyoshi K, Ohyama T. Effect of Nitrate on Nodule and Root Growth of Soybean (Glycine max (L.) Merr.). International Journal of Molecular Sciences. 2014; 15(3):4464-4480. https://doi.org/10.3390/ijms15034464
Chicago/Turabian StyleSaito, Akinori, Sayuri Tanabata, Takanari Tanabata, Seiya Tajima, Manabu Ueno, Shinji Ishikawa, Norikuni Ohtake, Kuni Sueyoshi, and Takuji Ohyama. 2014. "Effect of Nitrate on Nodule and Root Growth of Soybean (Glycine max (L.) Merr.)" International Journal of Molecular Sciences 15, no. 3: 4464-4480. https://doi.org/10.3390/ijms15034464