Role of Aeromonas hydrophila Flagella Glycosylation in Adhesion to Hep-2 Cells, Biofilm Formation and Immune Stimulation



Abstract
:1. Introduction
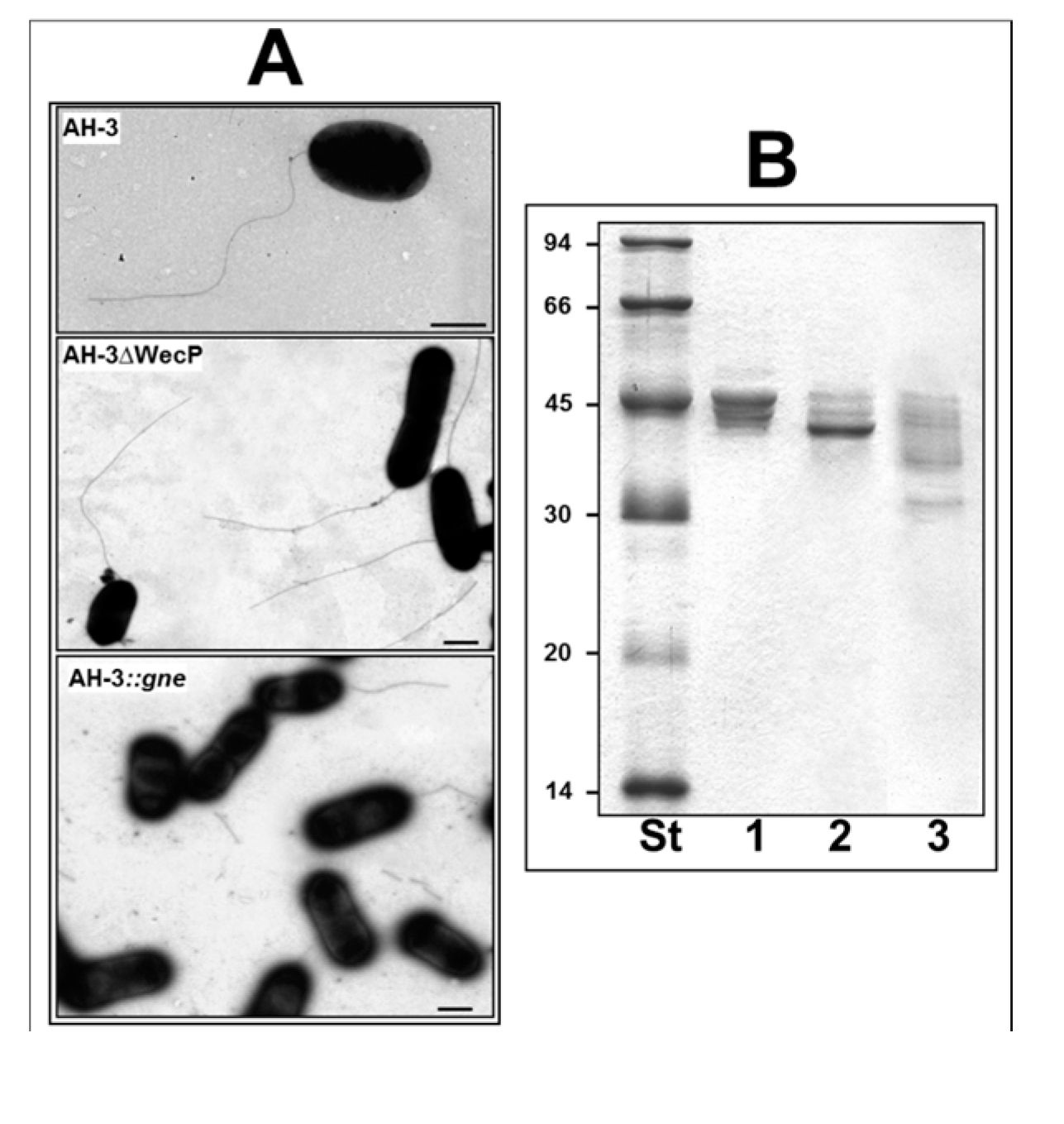
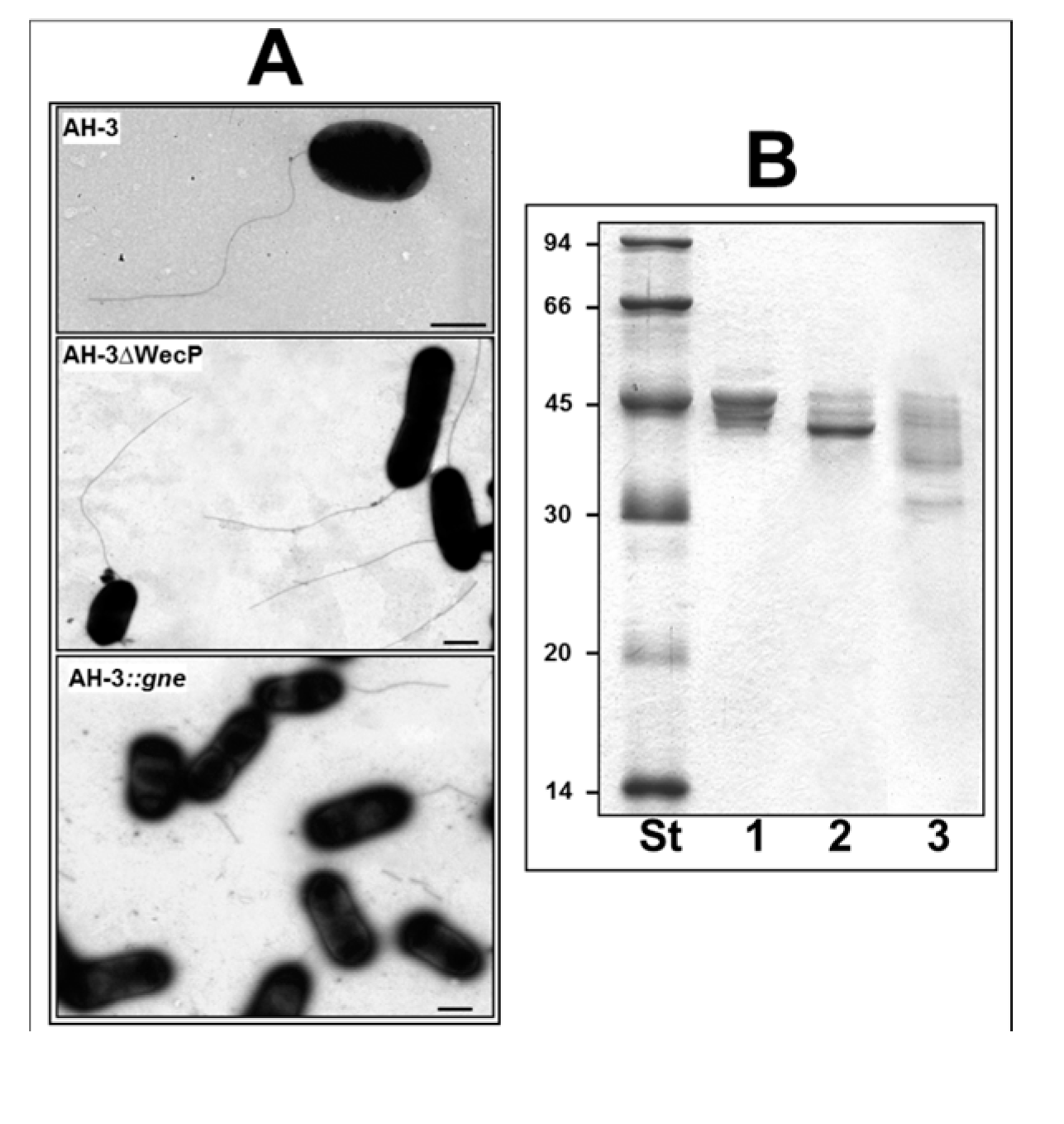
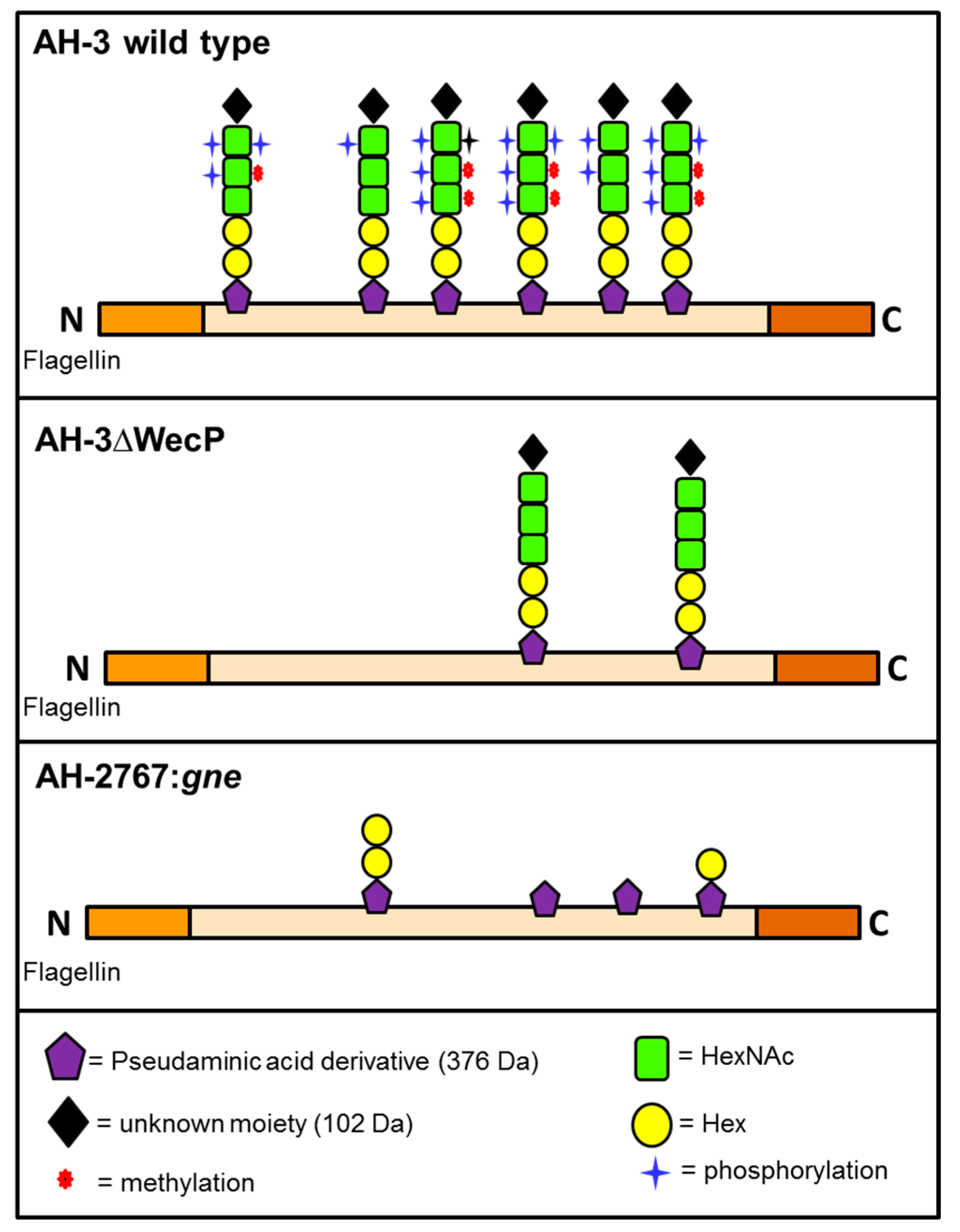
2. Results
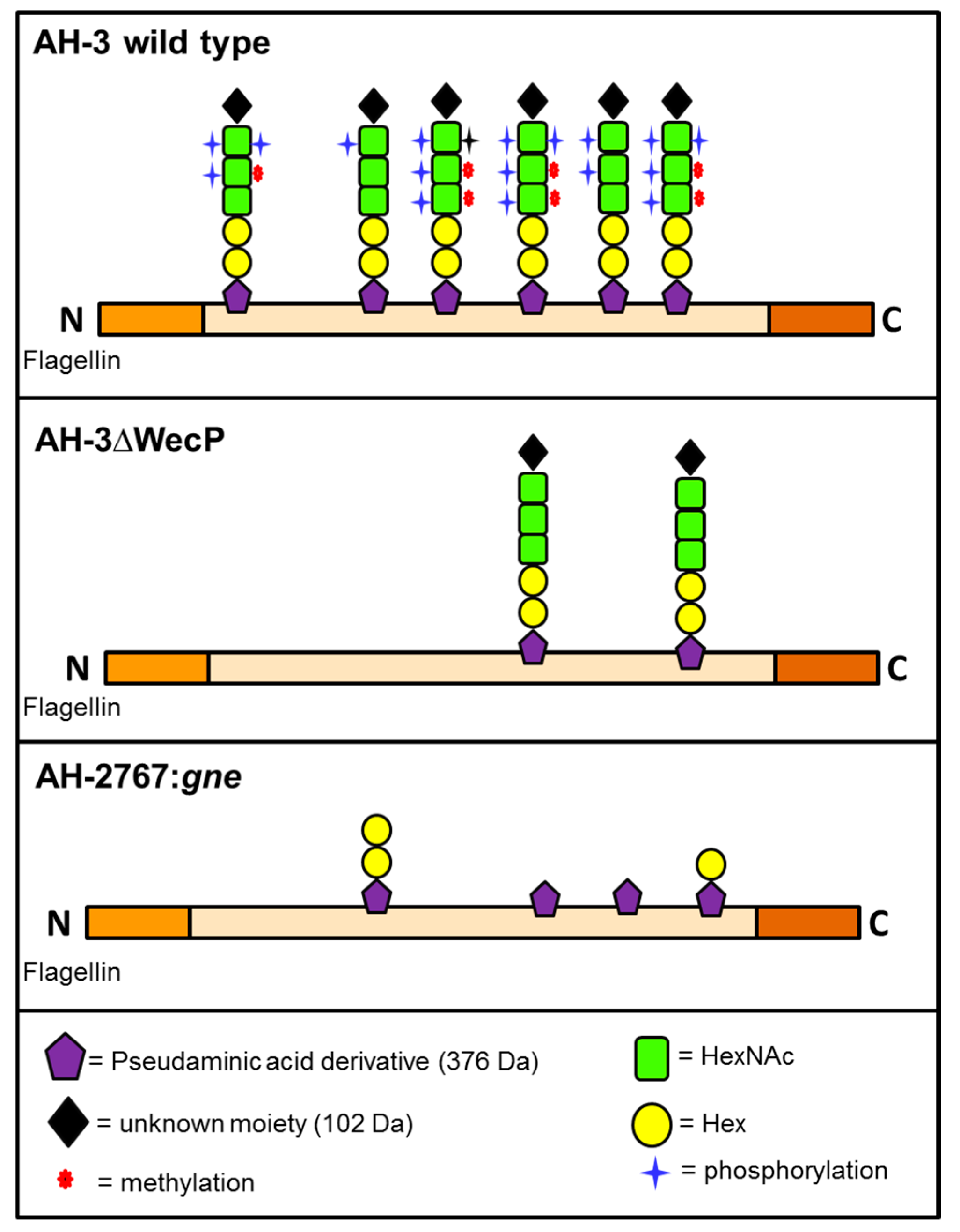
2.1. Adhesion to HEp-2 Cells and Biofilm Formation

{kind=link}
{kind=link}
{kind=link}
| Strain and Characteristics | Mean No. of Bacteria/ HEp-2 Cell +/− SD | % Reduction in Adhesion a |
|---|---|---|
| AH-3 (wild type) | 18.3 +/− 2.2 | - |
| AH-2767: gne mutant (O34−; flagella−−/+) | 7.2 +/− 0.7 | 61 b |
| AH-2767 + pACYC-GNE | 18.0 +/− 2.5 | <5 |
| AH-3ΔWecP (WecP mutant) (O34−; flagella−/+) | 9.8 +/− 1.0 | 46 |
| AH-3ΔWecP + pBAD-WecP | 17.9 +/− 1.7 | <5 |
| AH-3ΔManC (O34− mutant)→(O34−; flagella+) | 14.2 +/− 1.5 | 23 |
| AH-3ΔManC + pBAD-ManC | 18.7 +/− 1.9 | <5 |
| AH-3ΔPseB (flagella−) (O34+; flagella−) | 2.2 +/− 0.6 | 88 |
| AH-3ΔPseB + pBAD-PseB | 17.6 +/− 2.1 | <5 |
| AH-3ΔPseBΔManC (flagella−, O34− mutant) | <1.5 | >90 |
| AH-3:WecX (flagella−, O34+) | 2.3 +/− 0.4 | 87 |
| AH-3:WecX + pBAD-WecX | 18.1 +/− 1.6 | <5 |
| Strain and Characteristics | Value (OD570) |
|---|---|
| AH-3 (wild type) | 1.32 +/− 0.11 |
| AH-2767: gne mutant (O34−; flagella−−/+) | <0.2 |
| AH-3ΔWecP (O34−; flagella−/+) | 0.47 +/− 0.05 |
| AH-3ΔManC (O34−; flagella+) | 0.65 +/− 0.07 |
| AH-3ΔPseB (flagella−) | <0.2 |
| AH-3ΔPseBΔManC (O34−; flagella−) | <0.2 |
| AH-3:WecX (flagella−) | <0.2 |
| AH-2767: gne mutant + pACYC-GNE | 1.30 +/− 0.15 |
| AH-3ΔWecP + pBAD-WecP | 1.29 +/− 0.13 |
| AH-3ΔManC + pBAD-ManC | 1.31 +/− 0.10 |
| AH-3ΔPseB + pBAD-PseB | 1.27 +/− 0.12 |
| AH-3:WecX + pBAD-WecX | 1.29 +/− 0.08 |
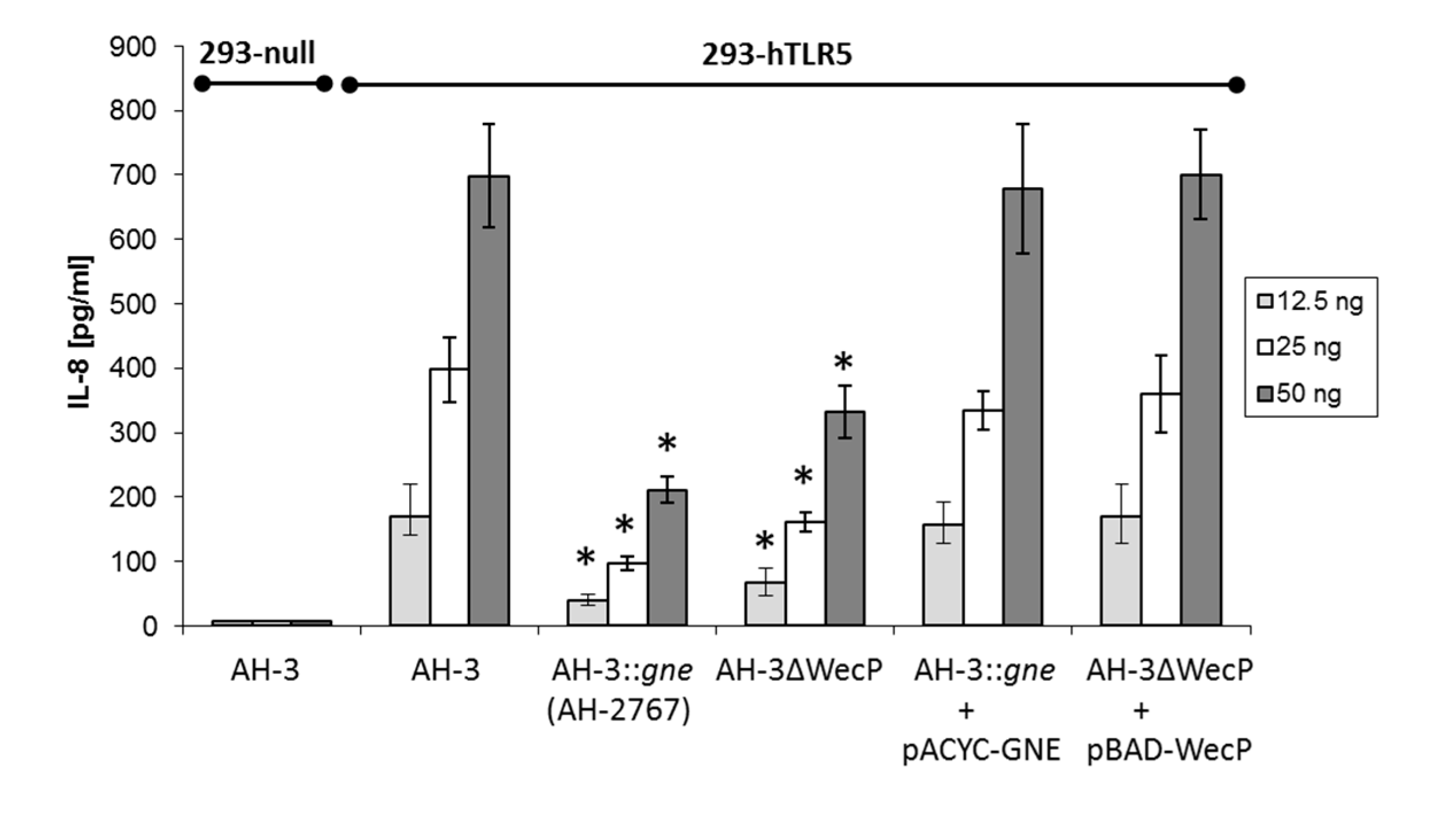
2.2. IL-8 Immune Stimulation
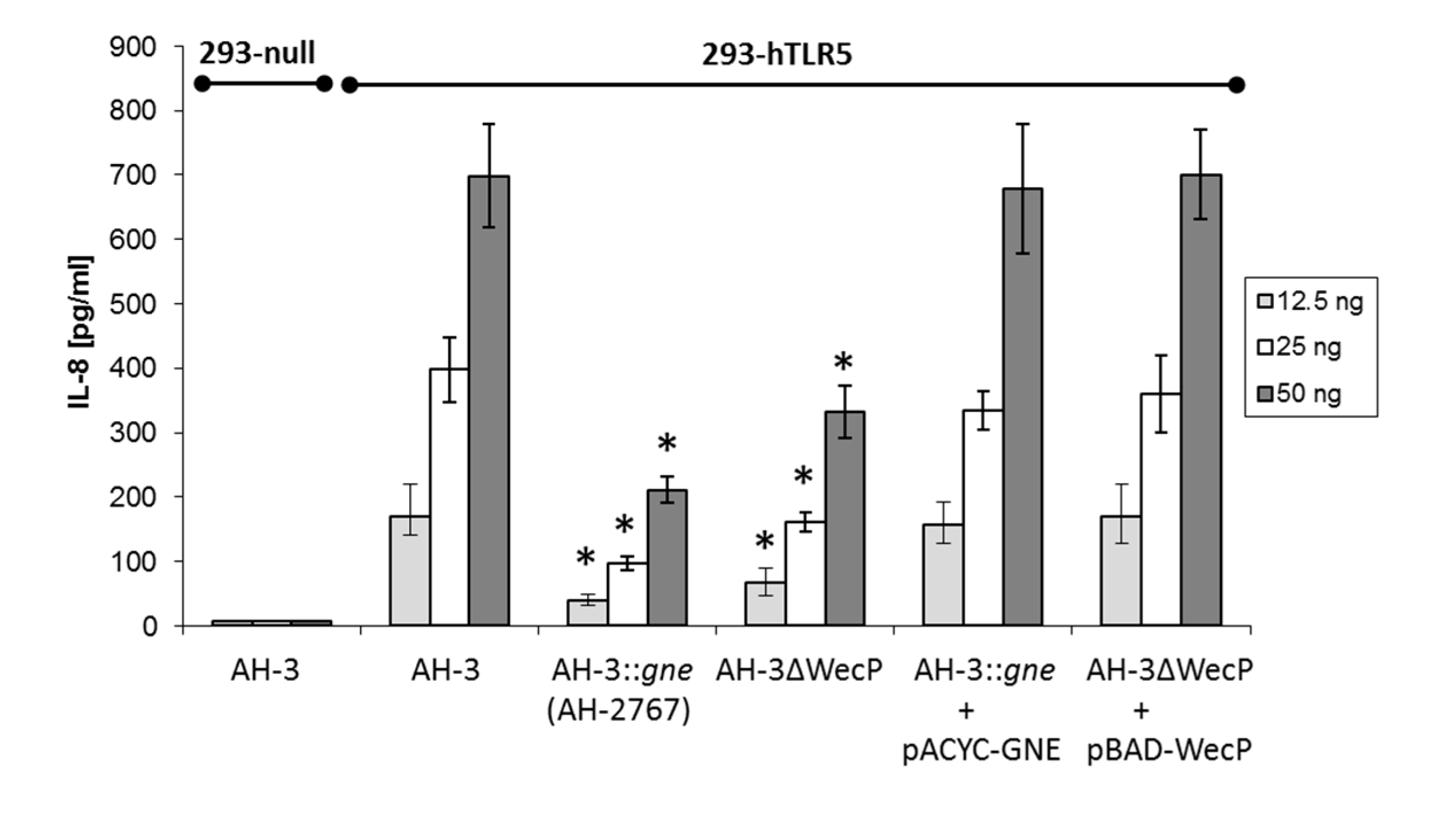
3. Discussion
4. Experimental Section
4.1. Bacterial Strains, Plasmids and Growth Conditions
| Strain or Plasmid | Relevant Characteristics | Reference or Source |
|---|---|---|
| E. coli strains | ||
| DH5α | F− end A hsdR17 (rK− mK+) supE44 thi-1 recA1 gyr-A96 _80lacZM15 | [23] |
| BL21(λD3) | F− ompT hsdSB (rB− mB−) gal dcm(λD3) | Novagen |
| A. hydrophila strains | ||
| AH-3 | O34, Wild type | [12] |
| AH-2767 | AH-3:gne mutant KmR | [13] |
| AH-3ΔWecP | AH-3 WecP mutant | [12] |
| AH-3ΔManC | AH-3 mutant in frame unable produce O34-antigen LPS | [11] |
| AH-3ΔPseB | AH-3 pseB mutant in frame with pDM4 | [9] |
| AH-3ΔPseFΔManC | AH-3 double mutant pseB and in frame unable produce O34-antigen LPS | [11] |
| AH-3:WecX | AH-3 wecX defined insertion mutant, CmR | [11] |
| Plasmids | ||
| pET-30 Xa/LIC | IPTG inducible expression vector KmR | Novagen |
| pET-30-FlaA | pET-30 Xa/LIC with A. hydrophila AH-3 flaA | This study |
| pET-30-FlaB | pET-30 Xa/LIC with A. hydrophila AH-3 flaB | This study |
| pBAD-ManC | pBAD33 with AH-3 manC | [11] |
| pBAD-WecP | pBAD33 with AH-3 wecP | [12] |
| pBAD-WecX | pBAD33 with AH-3 wecX | [11] |
| pACYC-GNE | pACYC184 with AH-3 gne | [13] |
4.2. Motility Assays
4.3. Transmission Electron Microscopy (TEM)
4.4. Flagella Purification
4.5. Adherence Assay to HEp-2 Cell
4.6. Biofilm Formation
4.7. Non-Glycosylated A. hydrophila Flagellins
4.8. Interleukin-8 (IL-8) Assay
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Frenchel, T. Microbial behavior in a heterogeneous world. Science 2002, 296, 1068–1071. [Google Scholar]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar]
- Shimada, T.; Sakazaki, R.; Suzuki, K. Peritrichous flagella in mesophilic strains of Aeromonas. Jpn. J. Med. Sci. Biol. 1985, 38, 141–145. [Google Scholar]
- Canals, R.; Altarriba, M.; Vilches, S.; Horsburg, G.; Shaw, J.S.; Tomas, J.M.; Merino, S. Analysis of the lateral flagella gene system of Aeromonas hydrophila AH-3. J. Bacteriol. 2006, 188, 852–862. [Google Scholar]
- Kirov, S.M.; Castrisios, M.; Shaw, J.G. Aeromonas. flagella (polar and lateral) are enterocyte adhesins that contribute to biofilm formation on surfaces. Infect. Immun. 2004, 72, 1939–1945. [Google Scholar]
- Logan, S.M. Flagellar glycosylation—A new component of the motility repertoire? Microbiology. 2006, 152, 1249–1262. [Google Scholar]
- Nothaft, H.; Symanski, C.M. Protein glycosylation in bacteria: Sweeter than ever. Nat. Rev. Microbiol. 2010, 8, 765–778. [Google Scholar]
- Twine, S.M.; Logan, S.M. Bacterial glycomic: Current research technology and applications. In Current Research Technology and Applications; Reid, S.W., Reid, A.N., Twine, S.M., Eds.; Caister Academic Press: Norwich, UK, 2011. [Google Scholar]
- Wilhelms, M.; Fulton, K.M.; Twine, S.M.; Tomás, J.M.; Merino, S. Differential glycosylation of polar and lateral flagellins in Aeromonas hydrophila AH-3. J. Biol. Chem. 2012, 287, 27851–27862. [Google Scholar]
- Valvano, M.A. Common themes in glyconjugate assembly. Using the biogenesis of O-antigen lipopolysaccharide as a model system. Biochemistry 2011, 76, 729–735. [Google Scholar]
- Merino, S.; Fulton, K.M.; Twine, S.M.; Wilhelms, M.; Molero, R.; Tomás, J.M. Aeromonas hydrophila flagella glycosylation: Involvement of a lipid carrier. PLoS One 2014, 9, e89630. [Google Scholar]
- Merino, S.; Jimenez, N.; Molero, R.; Bouamama, L.; Regué, M.; Tomás, J.M. A UDP-HexNAc: Polyprenol-P GalNAc-1-P transferase (WecP) representing a new subgroup of this enzyme family. J. Bacteriol. 2011, 193, 1943–1952. [Google Scholar]
- Canals, R.; Jiménez, N.; Vilches, S.; Regué, M.; Merino, S.; Tomás, J.M. The UDP N-acetylgalactosamine 4-epimerase gene is essential for mesophilic Aeromonas serotype O34 virulence. Infect. Immun. 2006, 74, 537–548. [Google Scholar]
- Knirel, Y.A.; Shaskov, A.S.; Sechenkova, S.N.; Merino, S.; Tomás, J.N. Structure of the O-polysaccharide of Aeromonas hydrophila O34: A case of random O-acetylation of 6-deoxy-l-talose. Carbohydr. Res. 2002, 337, 1381–1386. [Google Scholar]
- Kleta, S.; Nordhoff, M.; Tedin, K.; Wieler, L.H.; Kolenda, R.; Oswald, S.; Oelschlaeger, T.A.; Bleiß, W.; Schierack, P. Role of F1C fimbriae, flagella, and secreted bacterial components in the inhibitory effect of probiotic Escherichia coli Nissle 1917 on atypical enteropathogenic E. coli infection. Infect. Immun. 2014, 82, 1801–1812. [Google Scholar]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002, 56, 187–210. [Google Scholar]
- Guerry, P. Campylobacter flagella: Not just for motility. Trends Microbiol. 2007, 15, 456–461. [Google Scholar]
- Howard, S.L.; Jagannathan, A.; Soo, E.C.; Hui, J.P.M.; Aubry, A.J.; Ahmed, I.; Karlyshev, A.; Kelly, J.F.; Jones, M.A.; Stevens, M.P.; et al. Campylobacter jejuni glycosylation island important in cell charge, legionaminic acid biosynthesis, and colonization of chickens. Infect. Immun. 2009, 77, 2544–2556. [Google Scholar]
- Asakura, H.; Churin, Y.; Bauer, B.; Boettcher, J.P.; Bartfeld, S.; Hashii, N.; Kawasaki, N.; Mollenkopf, H.J.; Jungblut, P.R.; Brinkmann, V.; et al. Helicobacter pylori HP0518 affects flagellin glycosylation to alter bacterial motility. Mol. Microbiol. 2010, 78, 1130–1144. [Google Scholar]
- Huang, F.C. Regulation of Salmonella flagellin-induced interleukin-8 in intestinal epithelial cells by muramyl dipeptide. Cell. Immunol. 2012, 278, 1–9. [Google Scholar]
- Basu, M.; Swain, B.; Maiti, N.K.; Routray, P.; Samanta, M. Inductive expression of toll-like receptor 5 (TLR5) and associated downstream signaling molecules following ligand exposure and bacterial infection in the Indian major carp, mrigal (Cirrhinus mrigala). Fish Shellfish Immunol. 2012, 32, 121–131. [Google Scholar]
- Merino, S.; Rubires, X.; Aguilar, A.; Tomás, J.M. The role of flagella and motility on the adherence and invasion to fish cell lines by Aeromonas hydrophila strains serogroup O34. FEMS 1997, 151, 213–217. [Google Scholar]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar]
- Canals, R.; Ramirez, S.; Vilches, S.; Horsburgh, G.; Shaw, J.G.; Tomás, J.M.; Merino, S. Polar flagellum biogenesis in Aeromonas hydrophila. J. Bacteriol. 2006, 188, 542–555. [Google Scholar]
- Carrello, A.; Silburn, K.A.; Budden, J.R.; Chang, B.J. Adhesion of clinical and environmental Aeromonas isolates to Hep-2 cells. J. Med. Microbiol. 1988, 26, 19–27. [Google Scholar]
- Pratt, L.A.; Kolter, R. Genetic analysis of Escherichia coli biofilm formation: Role of flagella, motility, chemotaxis and type I pilli. Mol. Microbiol. 1998, 30, 285–293. [Google Scholar]
- Canals, R.; Jiménez, N.; Vilches, S.; Regué, M.; Merino, S.; Tomás, J.M. The role of Gne and GalE in the virulence of Aeromonas hydrophila serotype O34. J. Bacteriol. 2007, 189, 540–550. [Google Scholar]
- Jiménez, N.; Vilches, S.; Lacasta, A.; Regué, M.; Merino, S.; Tomás, J.M. A bifunctional enzyme in a single gene catalyzes the incorporation of GlcN into the Aeromonas core LPS. J. Biol. Chem. 2009, 284, 32995–33005. [Google Scholar]
- Al-Dabbagh, B.; Mengin-Lecreux, D.; Bouhss, A. Purification and characterization of the bacterial UDP-GlcNAC: Undecaprenyl-phosphate GlcNAc-1-phosphate transferase WecA. J. Bacteriol. 2008, 190, 7141–7146. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merino, S.; Wilhelms, M.; Tomás, J.M. Role of Aeromonas hydrophila Flagella Glycosylation in Adhesion to Hep-2 Cells, Biofilm Formation and Immune Stimulation. Int. J. Mol. Sci. 2014, 15, 21935-21946. https://doi.org/10.3390/ijms151221935
Merino S, Wilhelms M, Tomás JM. Role of Aeromonas hydrophila Flagella Glycosylation in Adhesion to Hep-2 Cells, Biofilm Formation and Immune Stimulation. International Journal of Molecular Sciences. 2014; 15(12):21935-21946. https://doi.org/10.3390/ijms151221935
Chicago/Turabian StyleMerino, Susana, Markus Wilhelms, and Juan M. Tomás. 2014. "Role of Aeromonas hydrophila Flagella Glycosylation in Adhesion to Hep-2 Cells, Biofilm Formation and Immune Stimulation" International Journal of Molecular Sciences 15, no. 12: 21935-21946. https://doi.org/10.3390/ijms151221935