Nrf2 and Redox Status in Prediabetic and Diabetic Patients



Abstract
:1. Introduction
2. Results
2.1. Clinical and Anthropometrical Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | CTRL n = 260 | PRE n = 116 | DM2 n = 183 | p Value |
|---|---|---|---|---|
| Age (years) | 48.25 (41–52.5) | 49.97 (43–54) | 58.87 (50–64) a,b | <0.001 |
| Male, n (%) | 127 (47.6) | 47 (18.3) | 87 (34) | 0.4360 |
| Female, n (%) | 138 (45.54) | 69 (22.77) | 96 (31.6) | - |
| BMI (kg·m−2) | 27.7 (25.3–29.8) | 31.37 (27.5–34.4) a | 29.14 (26–31.4) a,b | <0.001 |
| SBP (mmHg) | 113.9 (110–120) | 117.1 (110–120) | 120.6 (110–130) a | <0.001 |
| DBP (mmHg) | 74.6 (70–80) | 77.9 (70–80) a | 77.5 (70–81) a | <0.001 |
| Smoking, n (%) | 61 (23.6) | 29 (25) | 35 (19.1) | 0.4050 |
| Glucose (mg·dL−1) | 91.1 (86–97) | 106.8 (99–113) a | 153.8 (108-169) a,b | <0.001 |
| HbA1c (%) | 4.6 (4.2–5.1) | 5.9 (5.7–6.1) a | 7.4 (5.8–8.4) a,b | <0.001 |
| HOMA-IR | 1.8 (1.3–2.3) | 4.8 (3.2–5.9) a | - | <0.001 |
| TG (mg·dL−1) | 167.6 (106–210) | 202 (130–242) a | 214.2 (126–244) a | 0.0059 |
| T-C (mg·dL−1) | 194.1 (167–217) | 195.3 (168.5–218.5) | 185.3 (150–212) | 0.0910 |
| HDL-C (mg·dL−1) | 50.9 (43–58) | 46.1 (39–53) a | 43.7 (35–55) a | <0.001 |
| LDL-C (mg·dL−1) | 112.7 (94.5–132.5) | 110.1 (90–130) | 110.3 (84–129.5) | 0.8800 |
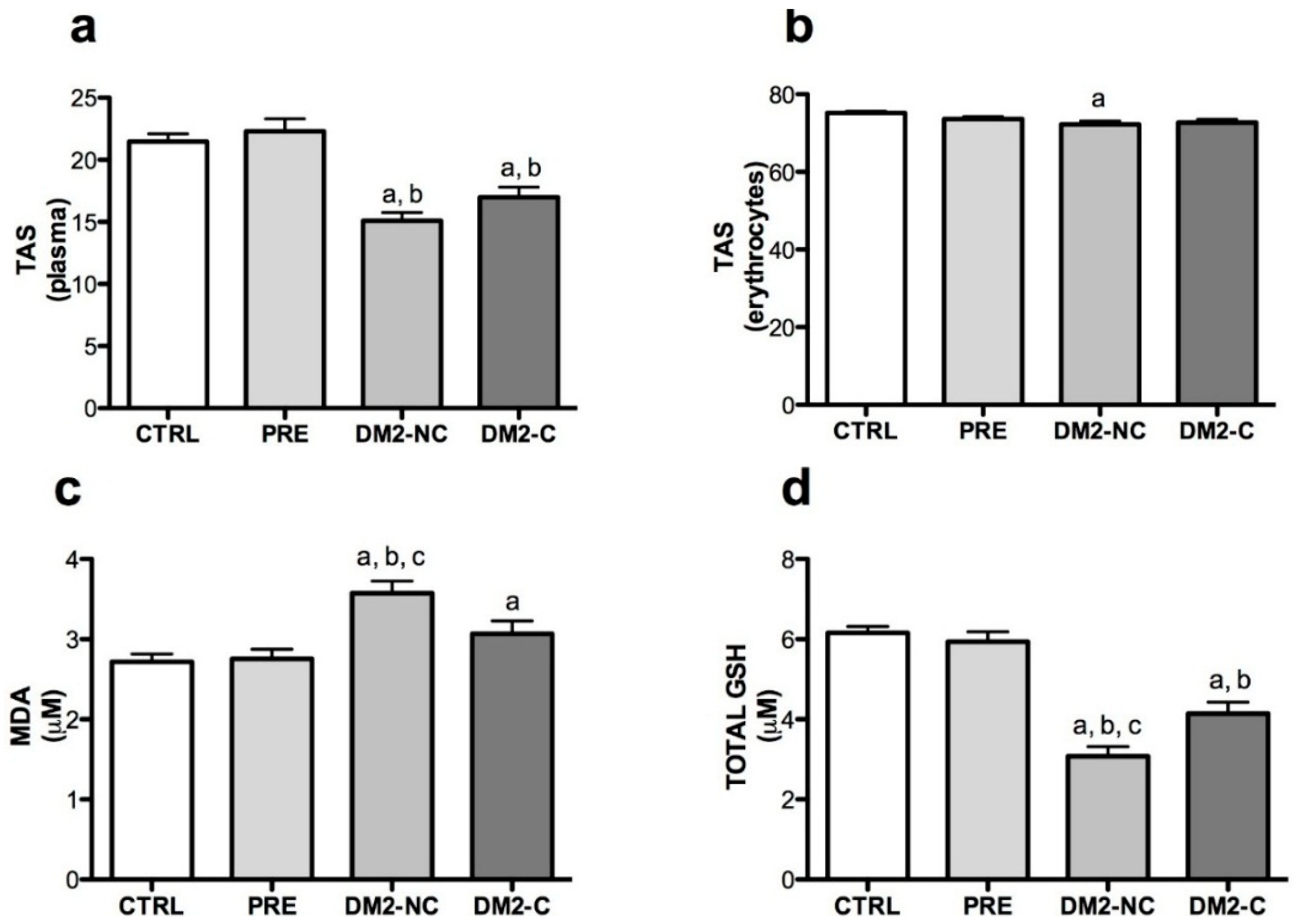
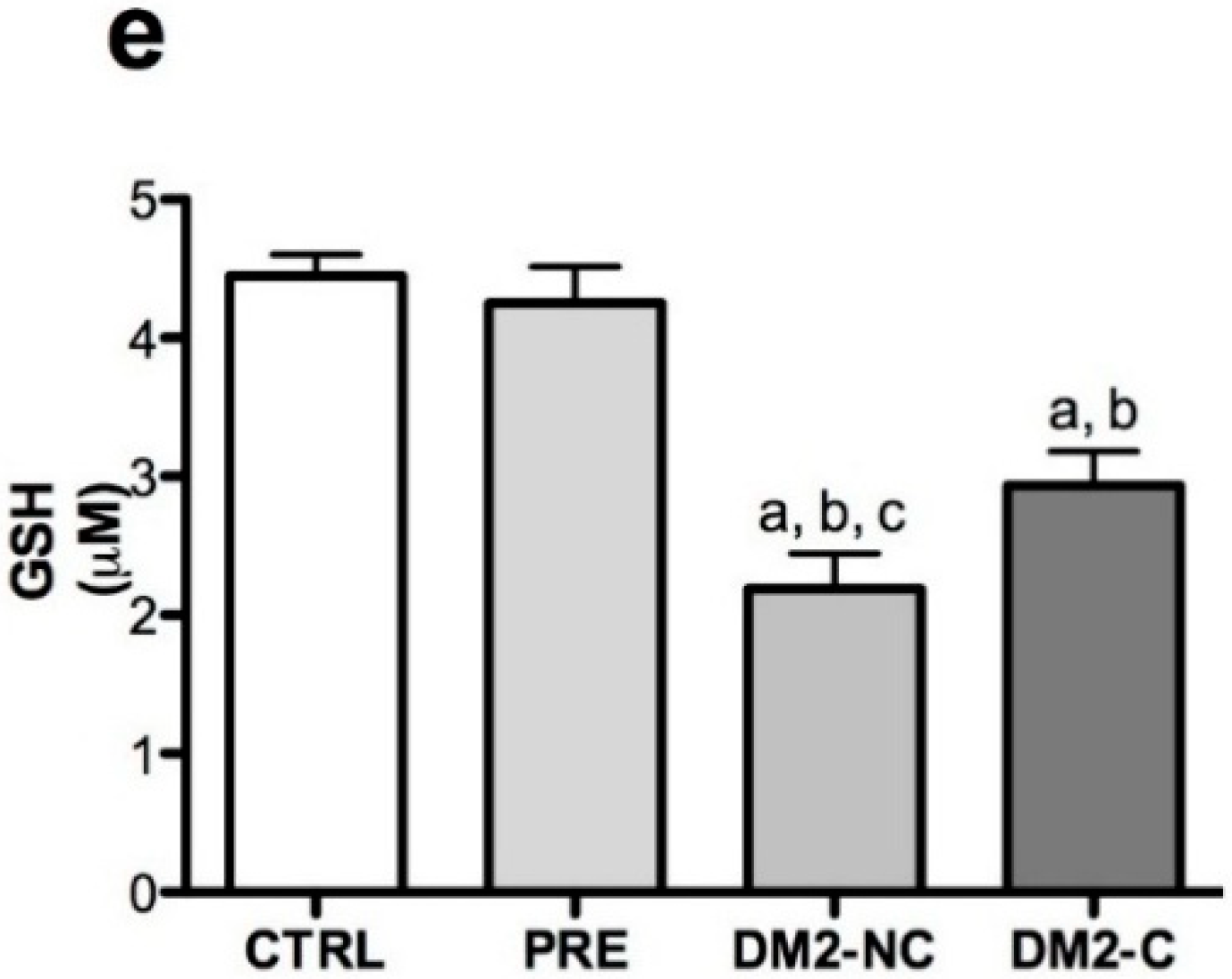
2.2. Oxidative Markers
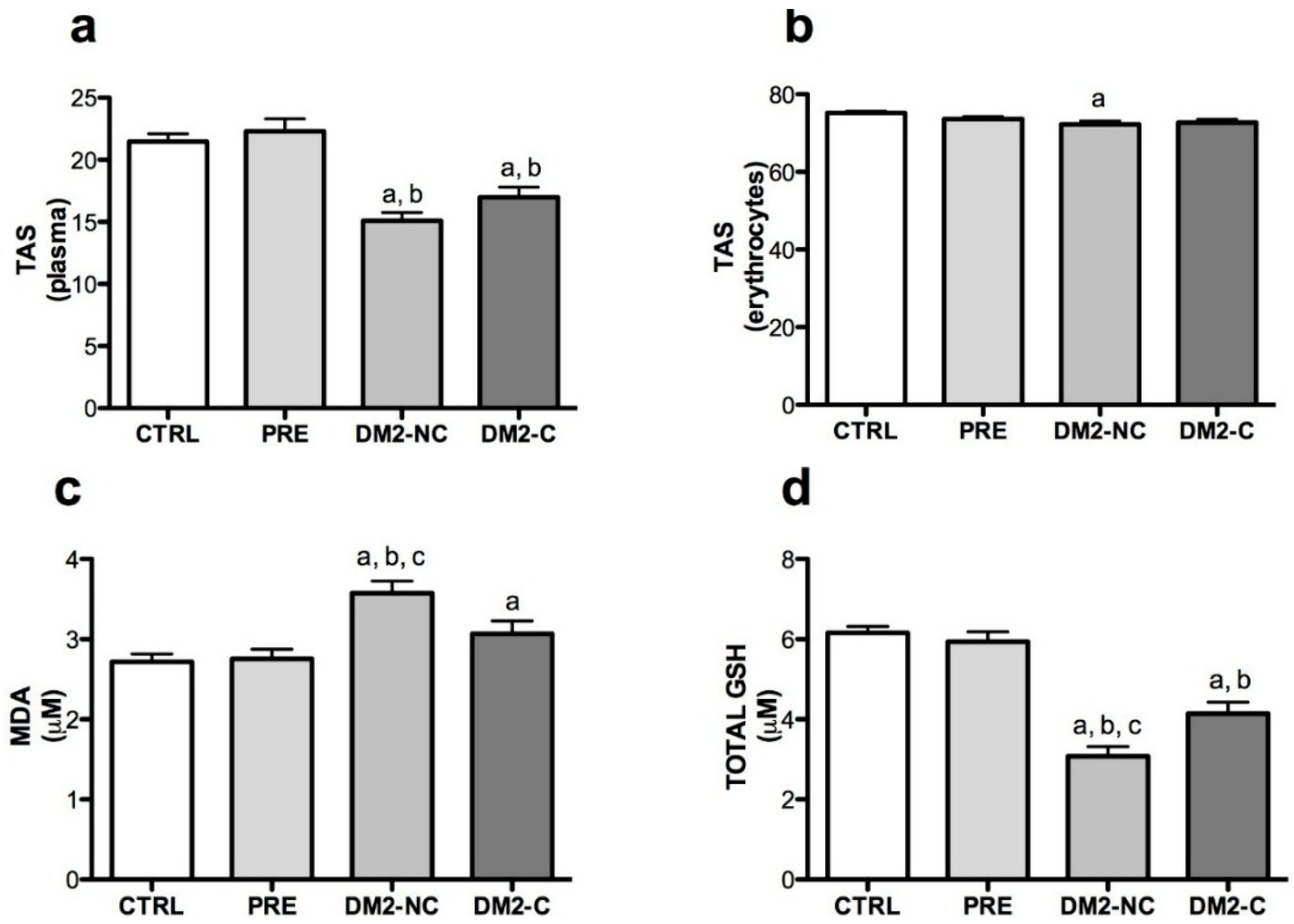
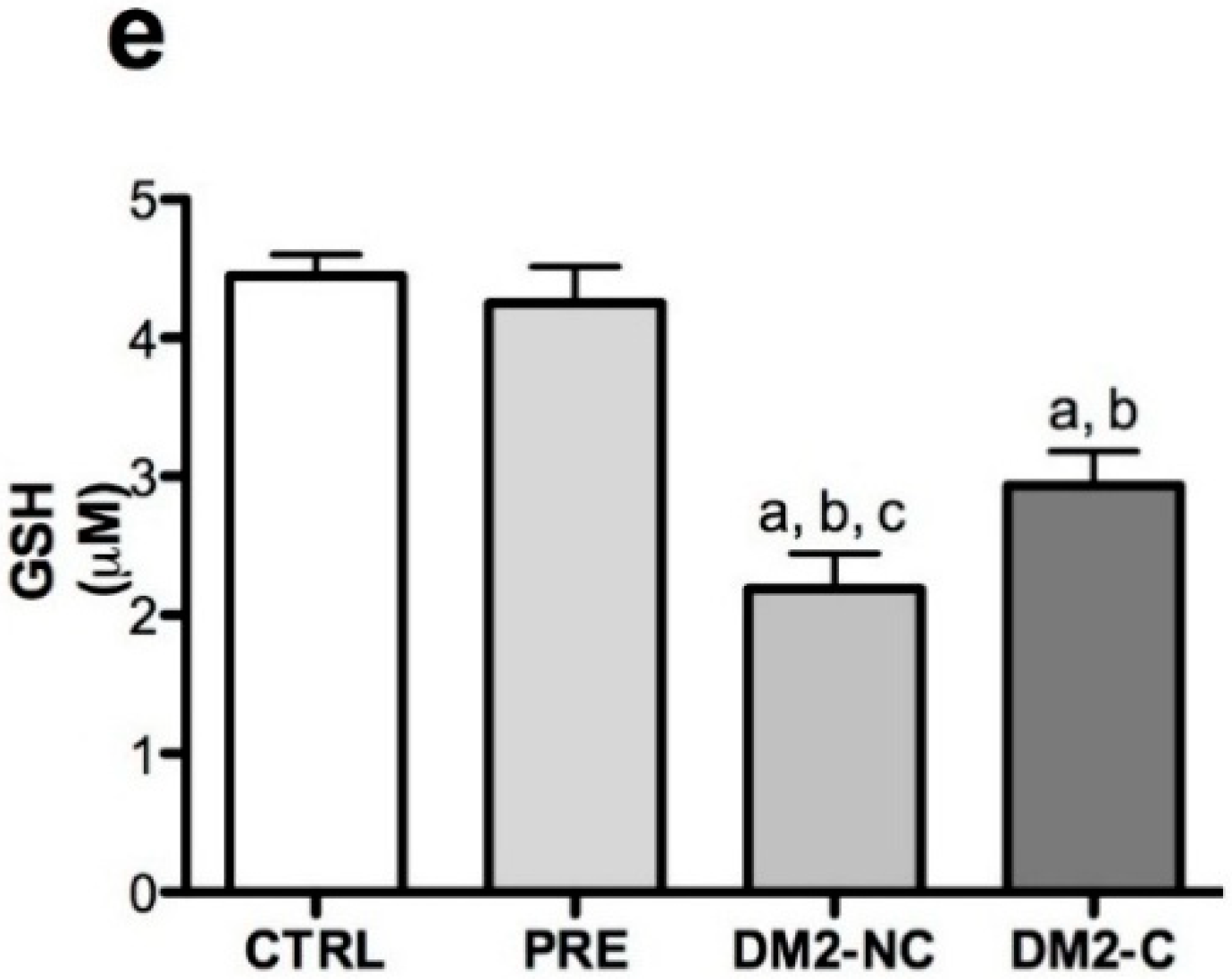
| Variables | Age | Gender | BMI | Glucose | HbA1c | |
|---|---|---|---|---|---|---|
| Plasma TAS | β | −0.21 | −2.28 | 0.012 | −0.019 | −0.98 |
| 95% CI | −0.28 to −0.13 | −3.82 to −0.19 | −0.16 to 0.18 | −0.037 to 0.076 | −1.7 to −0.2 | |
| p | <0.001 | 0.004 | 0.894 | 0.511 | 0.014 | |
| R2 | 0.190 | - | - | - | - | |
| Erythocyte TAS | β | −0.26 | −1.59 | 0.086 | −0.012 | −1.2 |
| 95% CI | −0.36 to −0.17 | −3.40 to 0.20 | −0.11 to 0.29 | −0.015 to 0.040 | −1.95 to −0.44 | |
| p | <0.001 | 0.084 | 0.411 | 0.381 | 0.002 | |
| R2 | 0.221 | - | - | - | - | |
| MDA | β | 0.006 | −0.14 | 0.04 | −0.019 | 0.17 |
| 95% CI | −0.01 to 0.23 | −0.43 to 0.15 | 0.004 to 0.074 | −0.004 to 0.003 | 0.06 to 0.28 | |
| p | 0.45 | 0.34 | 0.026 | 0.941 | 0.003 | |
| R2 | 0.08 | - | - | - | - | |
| GSH | β | −0.074 | −0.46 | 0.04 | 0.002 | −0.25 |
| 95% CI | −0.1 to −0.04 | −1.01 to 0.08 | −0.02 to 0.11 | −0.0007 to 0.004 | −0.45 to −0.055 | |
| p | <0.001 | 0.097 | 0.19 | 0.159 | 0.013 | |
| R2 | 0.13 | - | - | - | - | |
| SOD-1 | β | 0.29 | 7.04 | 0.052 | −0.005 | 2.99 |
| 95% CI | 0.06 to 0.51 | 3.1 to 10.9 | −0.42 to 0.53 | −0.07 to 0.06 | 1.14 to 4.84 | |
| p | 0.011 | <0.001 | 0.831 | 0.89 | 0.002 | |
| R2 | 0.124 | - | - | - | - | |
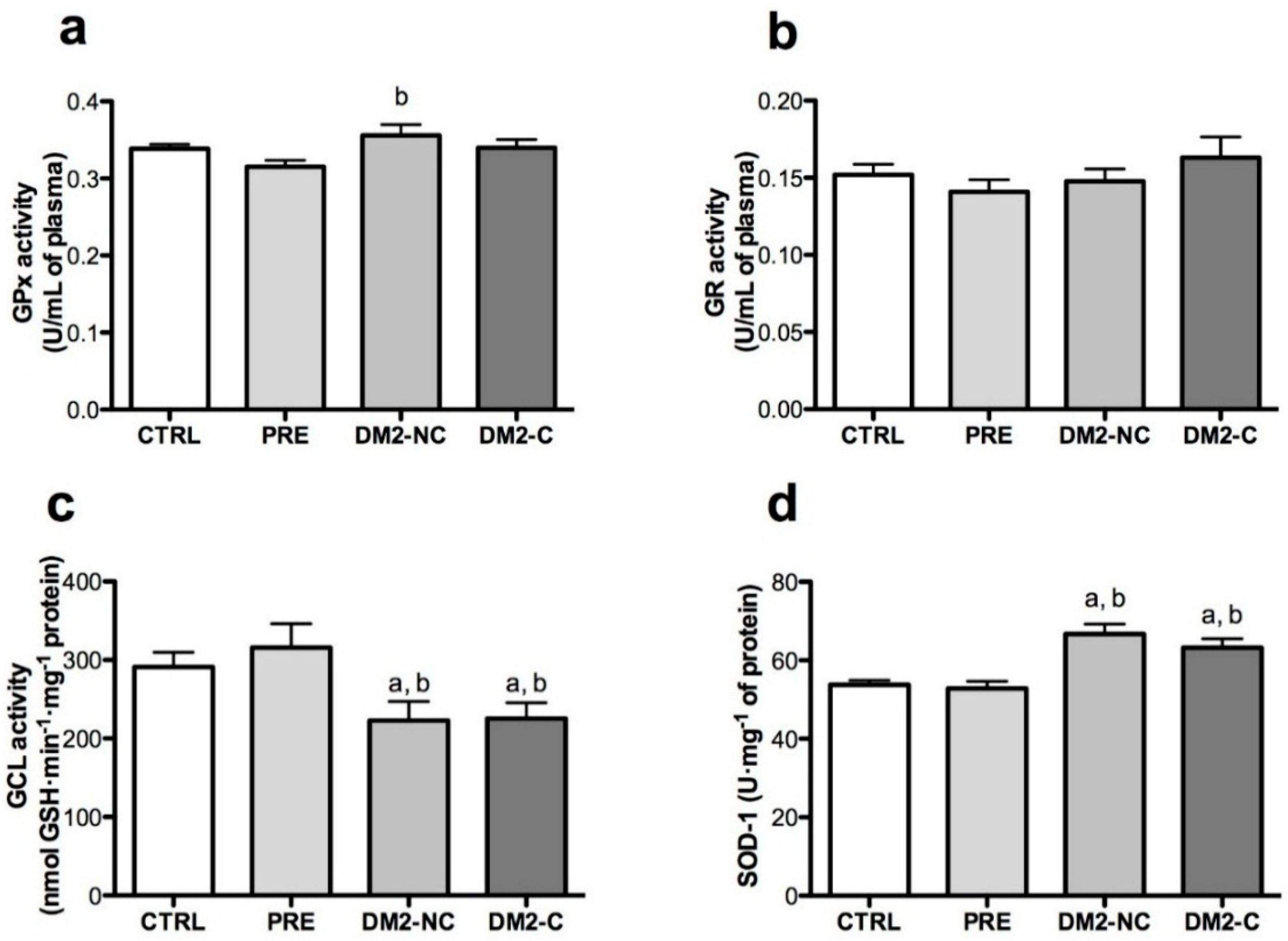
2.3. Activities of Antioxidant Enzymes
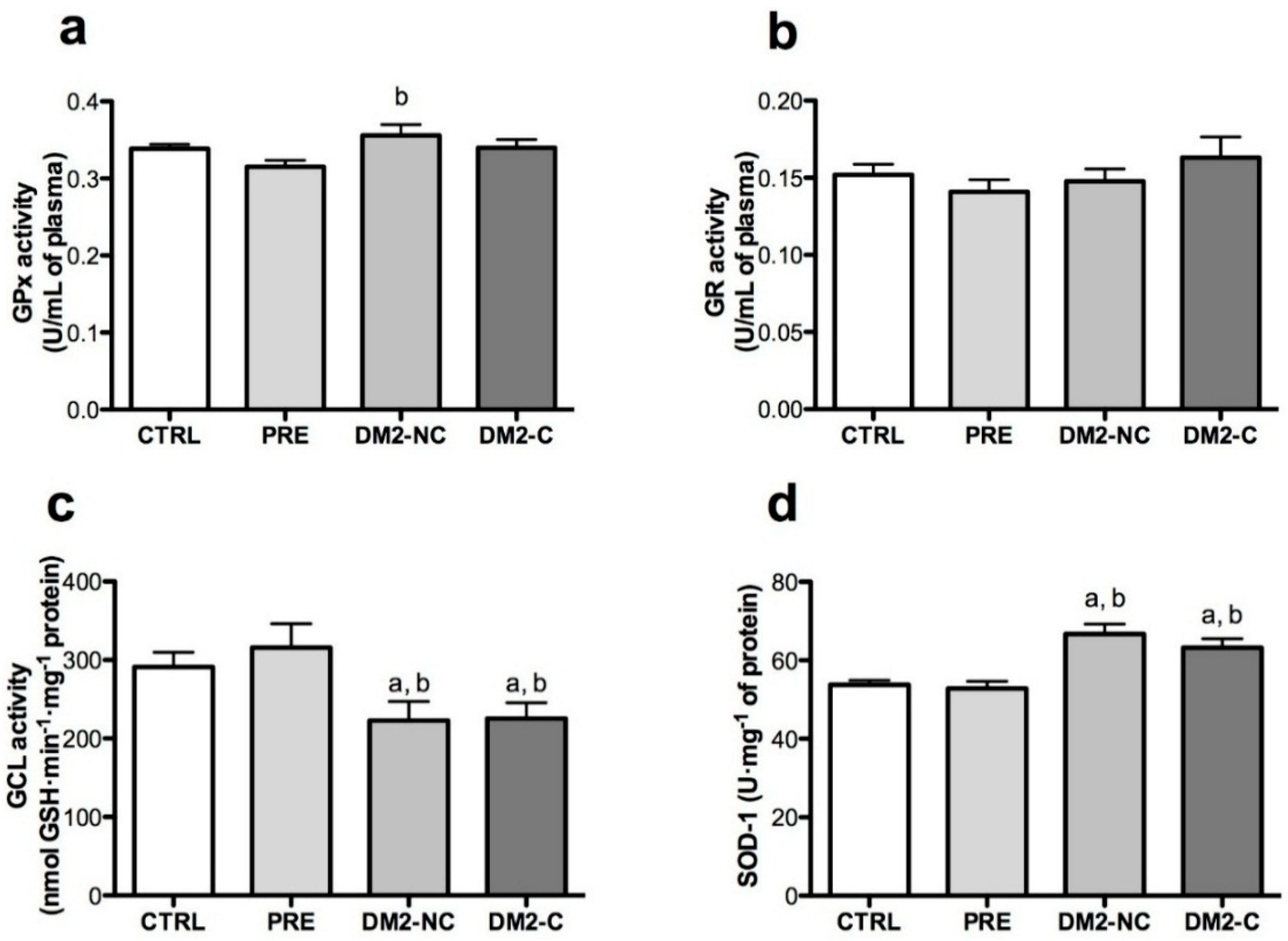
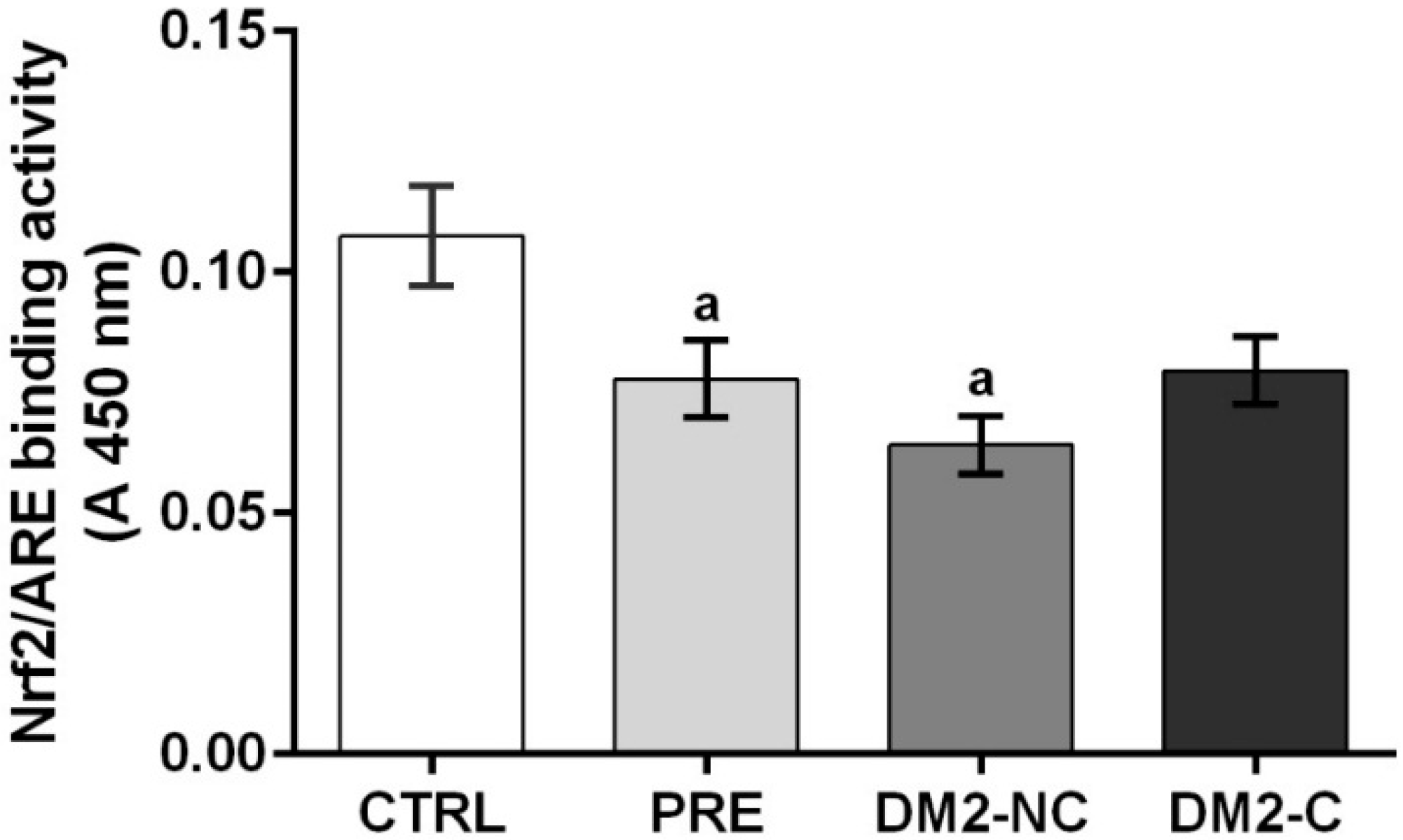
2.4. Nrf2/ARE Binding Activity Assay

| Variables | β ± S.E. of β | p | R2 | ||
|---|---|---|---|---|---|
| HbA1c | −0.097 | ± | 0.04 | 0.042 | 0.194 |
| Age | −0.011 | ± | 0.005 | 0.045 | - |
| Hospitalization | 0.48 | ± | 0.19 | 0.013 | - |
| DM2-NC | −1.06 | ± | 0.23 | <0.001 | - |
| DM2-C | −0.48 | ± | 0.14 | 0.001 | - |
| TAS * GSH | −0.008 | ± | 0.0004 | 0.044 | - |
3. Discussion
4. Experimental Section
4.1. Study Design and Subjects
4.2. Sample Collection and Biochemical Analysis
4.3. PBMC and Nuclear Extract Isolations
4.4. Redox Status Markers
4.4.1. Total Antioxidant Status (TAS)
4.4.2. MDA and GSH Content in Plasma
4.4.3. Activity of Antioxidant Enzymes in Plasma
4.4.4. Activity of Antioxidant Enzymes in Erythrocyte
4.5. Nrf2 Binding Activity Assay
4.6. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chen, L.; Magliano, D.J.; Zimmet, P. The worldwide epidemiology of type 2 diabetes mellitus-present and future perspectives. Nat. Rev. Endocrinol. 2012, 8, 228–236. [Google Scholar] [CrossRef]
- Arredondo, A.; de Icaza, E. The cost of diabetes in Latin America: Evidence from Mexico. Value Health 2011, 14, S85–S88. [Google Scholar]
- National Institute of Public Health. National Health and Nutrition Survery-2010. Damages and Health Services, Cuernavaca, M.I.S. Available online: http://consulta.mx/web/index.php/estudios-e-investigaciones/otros-estudios/502-encuesta-nacional-de-salud-y-nutricion-2012-ensanut (accessed on 8 August 2014).
- American Diabetes Association. Standards of medical care in diabetes-2014. Diabetes Care 2014, 37, 14–80. [Google Scholar]
- Weyer, C.; Bogardus, C.; Mott, D.; Pratley, R. The natural history of insulin secretory dysfunction and insulin resistance in the pathogenesis of type 2 diabetes mellitus. J. Clin. Investig. 1999, 104, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Kaneto, H.; Katakami, N.; Kawamori, D.; Miyatsuka, T.; Sakamoto, K.; Matsuoka, T.A.; Matsuhisa, M.; Yamasaki, Y. Involvement of oxidative stress in the pathogenesis of diabetes. Antioxid. Redox Signal. 2007, 9, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Haber, E.P.; Hirabara, S.M.; Rebelato, E.L.; Procopio, J.; Morgan, D.; Oliveira-Emilio, H.C.; Carpinelli, A.R.; Curi, R. Diabetes associated cell stress and dysfunction: Role of mitochondrial and non-mitochondrial ROS production and activity. J. Physiol. 2007, 583, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Perner, A.; Nielsen, S.; Rask-Madsen, J. High glucose impairs superoxide production from isolated blood neutrophils. Intensiv. Care Med. 2003, 29, 642–645. [Google Scholar]
- Wolff, S.; Dean, R. Glucose autoxidation and protein modification. The potential role of “autoxidative glycosylation” in diabetes. Biochem. J. 1987, 245, 243–250. [Google Scholar]
- Hybertson, B.; Gao, B.; Bose, S.; McCord, J. Oxidative stress in health and disease: The therapeutic potential of Nrf2 activation. Mol. Aspects Med. 2011, 32, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Kodiha, M.; Stochaj, U. Nuclear transport: A switch for the oxidative stress-signaling circuit? J. Signal Transduct. 2012, 2012. [Google Scholar] [CrossRef]
- Magesh, S.; Chen, Y.; Hu, L. Small molecule modulators of Keap1-Nrf2-ARE pathway as potential preventive and therapeutic agents. Med. Res. Rev. 2012, 32, 687–726. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Qian, Q.; Adaikalakoteswari, A.; Rabbani, N.; Babaei-Jadidi, R.; Thornalley, P. Activation of NF-E2-related factor-2 reverses biochemical dysfunction of endothelial cells induced by hyperglycemia linked to vascular disease. Diabetes 2008, 57, 2809–2817. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Hur, E.; Ryoo, I.; Jung, K.; Kwak, J.; Kwak, M. Involvement of the Nrf2-proteasome pathway in the endoplasmic reticulum stress response in pancreatic beta-cells. Toxicol. Appl. Pharmacol. 2012, 264, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Yagishita, Y.; Fukutomi, T.; Sugawara, A.; Kawamura, H.; Takahashi, T.; Pi, J.; Uruno, A.; Yamamoto, M. Nrf2 protects pancreatic beta-cells from oxidative and nitrosative stress in diabetic model mice. Diabetes 2014, 63, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Shao, W.; Chiang, Y.; Foltz, W.; Zhang, Z.; Ling, W.; Fantus, I.G.; Jin, T. Oltipraz upregulates the nuclear factor (erythroid-derived 2)-like 2 [corrected] (NRF2) antioxidant system and prevents insulin resistance and obesity induced by a high-fat diet in C57BL/6J mice. Diabetologia 2011, 54, 922–934. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Li, Q.; Liu, Z.; Wang, J.; Wang, X.; Qin, M.; Wen, Q. Curcumin attenuates insulin resistance in hepatocytes by inducing Nrf2 nuclear translocation. Hepatogastroenterology 2011, 58, 2106–2111. [Google Scholar] [PubMed]
- He, X.; Kan, H.; Cai, L.; Ma, Q. Nrf2 is critical in defense against high glucose-induced oxidative damage in cardiomyocytes. J. Mol. Cell. Cardiol. 2009, 46, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Yoh, K.; Hirayama, A.; Ishizaki, K.; Yamada, A.; Takeuchi, M.; Yamagishi, S.; Morito, N.; Nakano, T.; Ojima, M.; Shimohata, H.; Itoh, K.; Takahashi, S.; Yamamoto, M. Hyperglycemia induces oxidative and nitrosative stress and increases renal functional impairment in Nrf2-deficient mice. Genes Cells 2008, 13, 1159–1170. [Google Scholar] [PubMed]
- Negi, G.; Kumar, A.; Joshi, R.P.; Sharma, S.S. Oxidative stress and Nrf2 in the pathophysiology of diabetic neuropathy: Old perspective with a new angle. Biochem. Biophys. Res. Commun. 2011, 408, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Palsamy, P.; Subramanian, S. Resveratrol protects diabetic kidney by attenuating hyperglycemia-mediated oxidative stress and renal inflammatory cytokines via Nrf2-Keap1 signaling. BBA Mol. Basis Dis. 2011, 1812, 719–731. [Google Scholar] [CrossRef]
- Pergola, P.; Raskin, P.; Toto, R.D.; Meyer, C.J.; Huff, J.W.; Grossman, E.B.; Krauth, M.; Ruiz, S.; Audhya, P.; Christ-Schmidt, H.; et al. BEAM Study Investigators. Bardoxolone methyl and kidney function in CKD with type 2 diabetes. N. Engl. J. Med. 2011, 365, 327–333. [Google Scholar]
- Dziegielewska-Gesiak, S.; Wysocka, E.; Michalak, S.; Nowakowska-Zajdel, E.; Kokot, T.; Muc-Wierzgoń, M. Role of lipid peroxidation products, plasma total antioxidant status, and Cu-, Zn-superoxide dismutase activity as biomarkers of oxidative stress in elderly prediabetics. Oxid. Med. Cell Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Al-Aubaidy, H.; Jelinek, H. 8-Hydroxy-2-deoxy-guanosine identifies oxidative DNA damage in a rural prediabetes cohort. Redox Rep. 2010, 15, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Zengi, A.; Ercan, G.; Caglayan, O.; Tamsel, S.; Karadeniz, M.; Simsir, I.; Harman, E.; Kahraman, C.; Orman, M.; Cetinkalp, S.; et al. Increased oxidative DNA damage in lean normoglycemic offspring of type 2 diabetic patients. Exp. Clin. Endocrinol. Diabetes 2011, 119, 467–471. [Google Scholar]
- Yoshida, K.; Hirokawa, J.; Tagami, S.; Kawakami, Y.; Urata, Y.; Kondo, T. Weakened cellular scavenging activity against oxidative stress in diabetes mellitus: Regulation of glutathione synthesis and efflux. Diabetologia 1995, 38, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Seghieri, G.; di Simplicio, P.; de Giorgio, L.; Anichini, R.; Alberti, L.; Franconi, F. Relationship between metabolic glycaemic control and platelet content of glutathione and its related enzymes, in insulin-dependent diabetes mellitus. Clin. Chim. Acta 2000, 299, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Kalkan, H.; Suher, M. The relationship between the level of glutathione, impairment of glucose metabolism and complications of diabetes mellitus. Pak. J. Med. Sci. 2013, 29, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Franklin, C.; Backos, D.; Mohar, I.; White, C.; Forman, H.; Kavanagh, T. Structure, function, and post-translational regulation of the catalytic and modifier subunits of glutamate cysteine ligase. Mol. Aspects Med. 2009, 30, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, R.; Acharya, J.; Ghaskadbi, S.; Goel, P. Thresholds of oxidative stress in newly diagnosed diabetic patients on intensive glucose-control therapy. PLoS One 2014, 9, e100897. [Google Scholar]
- Pavlovic, D.; Kocić, R.; Kocić, G.; Jevtović, T.; Radenković, S.; Mikić, D. Effect of four-week metformin treatment on plasma and erythrocyte antioxidative defense enzymes in newly diagnosed obese patients with type 2 diabetes. Diabetes Obes. Metab. 2000, 2, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Bandeira, S.M.; Glaucevane, S.G; Fonseca, L.J; Pires, A.; Gelain, D.P.; Moreira, J.C.; Rabelo, L.A.; Vasconcelos, S.M.; Goulart, M.O. Characterization of blood oxidative stress in type 2 diabetes mellitus patients: Increase in lipid peroxidation and SOD activity. Oxid. Med. Cell Longev. 2012. [Google Scholar] [CrossRef]
- Aouacheri, O.; Saka, S.; Krim, M.; Messaadia, A.; Maidi, I. The investigation of the oxidative stress-related parameters in type 2 diabetes mellitus. Can. J. Diabetes 2014. [Google Scholar] [CrossRef]
- Soliman, G. Blood lipid peroxidation (superoxide dismutase, malondialdehyde, glutathione) levels in Egyptian type 2 diabetic patients. Singapore Med. J. 2008, 49, 129–136. [Google Scholar] [PubMed]
- Bhakkiyalakshmi, E.; Shalini, D.; Sekar, T.; Rajaguru, P.; Paulmurugan, R.; Ramkumar, K. Therapeutic potential of pterostilbene against pancreatic beta-cell apoptosis mediated through Nrf2. Br. J. Pharmacol. 2014, 171, 1747–1757. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, L.F.; Pedruzzi, L.M.; Stenvinkel, P.; Stockler-Pinto, M.B.; Daleprane, J.B.; Leite, M., Jr.; Mafra, D. Nutritional strategies to modulate inflammation and oxidative stress pathways via activation of the master antioxidant switch Nrf2. Biochimie 2013, 95, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; McDonald, P.; Liu, J.; Klaassen, C. Screening of natural compounds as activators of the keap1-nrf2 pathway. Planta Med. 2014, 80, 97–104. [Google Scholar] [PubMed]
- Chuengsamarn, S.; Rattanamongkolgul, S.; Luechapudiporn, R.; Phisalaphong, C.; Jirawatnotai, S. Curcumin extract for prevention of type 2 diabetes. Diabetes Care 2012, 35, 2121–2127. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, C.; Peng, H.; Ye, Z.; Zhang, J.; Liu, X.; Lou, T. Activation of the Nrf2-ARE pathway attenuates hyperglycemia-mediated injuries in mouse podocytes. Cell Physiol. Biochem. 2014, 34, 891–902. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Vaziri, N.D. Contribution of impaired Nrf2-Keap1 pathway to oxidative stress and inflammation in chronic renal failure. Am. J. Physiol. Renal Physiol. 2010, 298, F662–F671. [Google Scholar]
- Marzec, J.M.; Christie, J.D.; Reddy, S.P.; Jedlicka, A.E.; Vuong, H.; Lanken, P.N.; Aplenc, R.; Yamamoto, T.; Yamamoto, M.; Cho, H.Y.; et al. Functional polymorphisms in the transcription factor NRF2 in humans increase the risk of acute lung injury. FASEB J. 2007, 21, 2237–2246. [Google Scholar]
- Marczak, E.D.; Marzec, J.; Zeldin, D.C.; Kleeberger, S.R.; Brown, N.J.; Pretorius, M.; Lee, C.R. Polymorphisms in the transcription factor NRF2 and forearm vasodilator responses in humans. Pharmacogenet Genomics 2012, 22, 620–628. [Google Scholar] [PubMed]
- Chen, Y.; Wu, Y.; Wu, Y.; Lee-Chen, G.; Chen, C. Genetic analysis of NFE2L2 promoter variation in Taiwanese Parkinson’s disease. Parkinsonism Relat. Disord. 2013, 19, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, Y.; Mitsuda, Y.; Tsuruta, Y.; Hamajima, N.; Niwa, T. Polymorphism of Nrf2, an antioxidative gene, is associated with blood pressure and cardiovascular mortality in hemodialysis patients. Int. J. Med. Sci. 2014, 11, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Koren, E.; Kohen, R.; Ginsburg, I. Polyphenols enhance total oxidant-scavenging capacities of human blood by binding to red blood cells. Exp. Biol. Med. 2010, 235, 689–699. [Google Scholar] [CrossRef]
- Chirino, Y.; Sanchez-González, J.; Martínez-Martínez, C.; Cruz, C.; Pedraza-Chaverri, J. Protective effects of apocynin against cisplatin-induced oxidative stress and nephrotoxicity. Toxicology 2008, 245, 18–23. [Google Scholar] [CrossRef] [PubMed]
- González-Reyes, S.; Guzmán-Beltrán, S.; Medina-Campos, O.; Pedraza-Chaverri, J. Curcumin pretreatment induces Nrf2 and an antioxidant response and prevents hemin-induced toxicity in primary cultures of cerebellar granule neurons of rats. Oxid. Med. Cell Longev. 2013, 2013. [Google Scholar] [CrossRef]
- White, C.; Viernes, H.; Krejsa, C.; Botta, D.; Kavanagh, T. Fluorescence-based microtiter plate assay for glutamate-cysteine ligase activity. Anal. Biochem. 2003, 318, 175–180. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Osorio, A.S.; Picazo, A.; González-Reyes, S.; Barrera-Oviedo, D.; Rodríguez-Arellano, M.E.; Pedraza-Chaverri, J. Nrf2 and Redox Status in Prediabetic and Diabetic Patients. Int. J. Mol. Sci. 2014, 15, 20290-20305. https://doi.org/10.3390/ijms151120290
Jiménez-Osorio AS, Picazo A, González-Reyes S, Barrera-Oviedo D, Rodríguez-Arellano ME, Pedraza-Chaverri J. Nrf2 and Redox Status in Prediabetic and Diabetic Patients. International Journal of Molecular Sciences. 2014; 15(11):20290-20305. https://doi.org/10.3390/ijms151120290
Chicago/Turabian StyleJiménez-Osorio, Angélica S., Alejandra Picazo, Susana González-Reyes, Diana Barrera-Oviedo, Martha E. Rodríguez-Arellano, and José Pedraza-Chaverri. 2014. "Nrf2 and Redox Status in Prediabetic and Diabetic Patients" International Journal of Molecular Sciences 15, no. 11: 20290-20305. https://doi.org/10.3390/ijms151120290