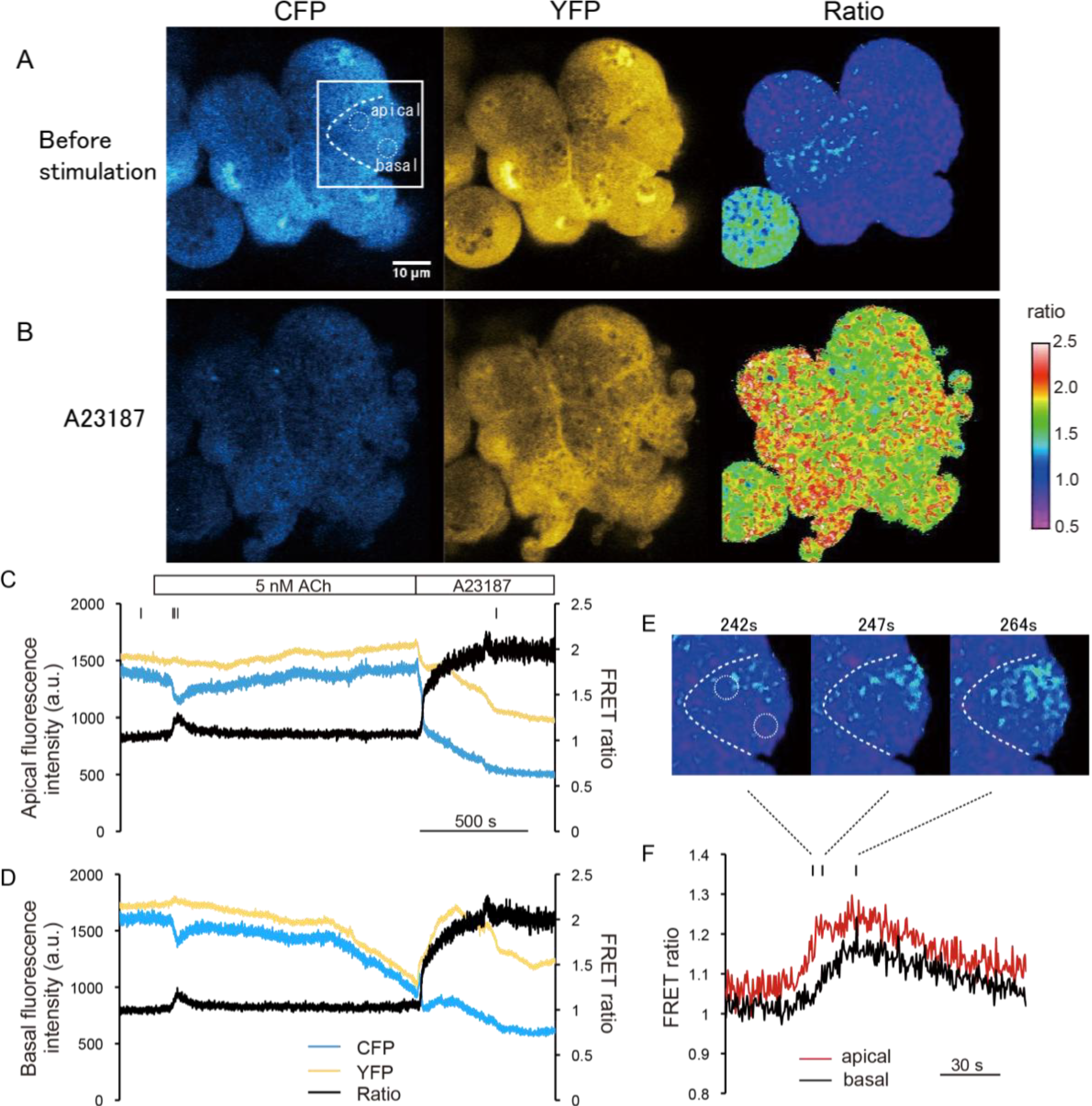
2.2. Evaluation of Dynamic Range of YC-Nano15 as an Ultrasensitive Ca2+ Indicator by Two-Photon Excitation FRET Ratio Imaging
Using two-photon excitation microscopy, we performed FRET ratio imaging of pancreatic acinar cells of YC-Nano15-205 mice. Two-photon excitation microscopy can visualize deep regions of tissues and excite multiple fluorescent proteins simultaneously [
3,
14,
15,
16]. To perform the quantitative analysis of Ca
2+ dynamics induced by the agonist stimulation, we employed ratiometric FRET imaging, and the acceptor/donor (YFP/CFP) ratio value was normalized to be 1 under normal conditions (before stimulation). First, we collected two-photon excitation fluorescence images of pancreatic acini under superfusing Solution A buffer in the absence of stimulation (
Figure 2A). Both CFP and YFP signal were detected at an excitation wavelength of 850 nm. For spatiotemporal FRET analysis, we calculated FRET ratio (YFP/CFP) images. After we increased the intracellular [Ca
2+] to 1 mM by treatment with a Ca
2+ ionophore, the FRET ratio increased as expected (
Figure 2B). Under these conditions, the morphology of these cells was significantly altered (e.g., by blebbing) probably due to the cytotoxicity of the Ca
2+ ionophore.
Figure 1.
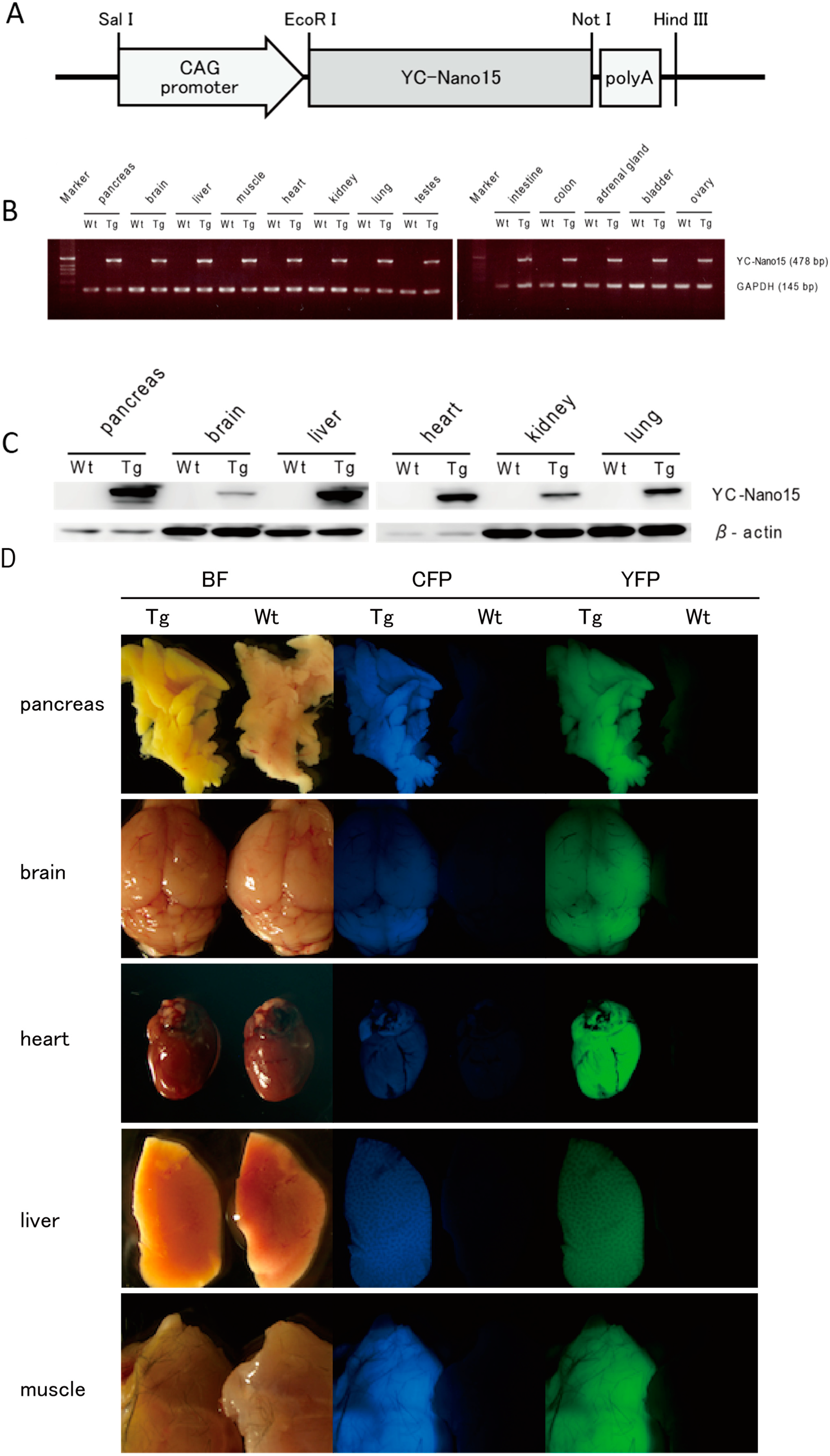
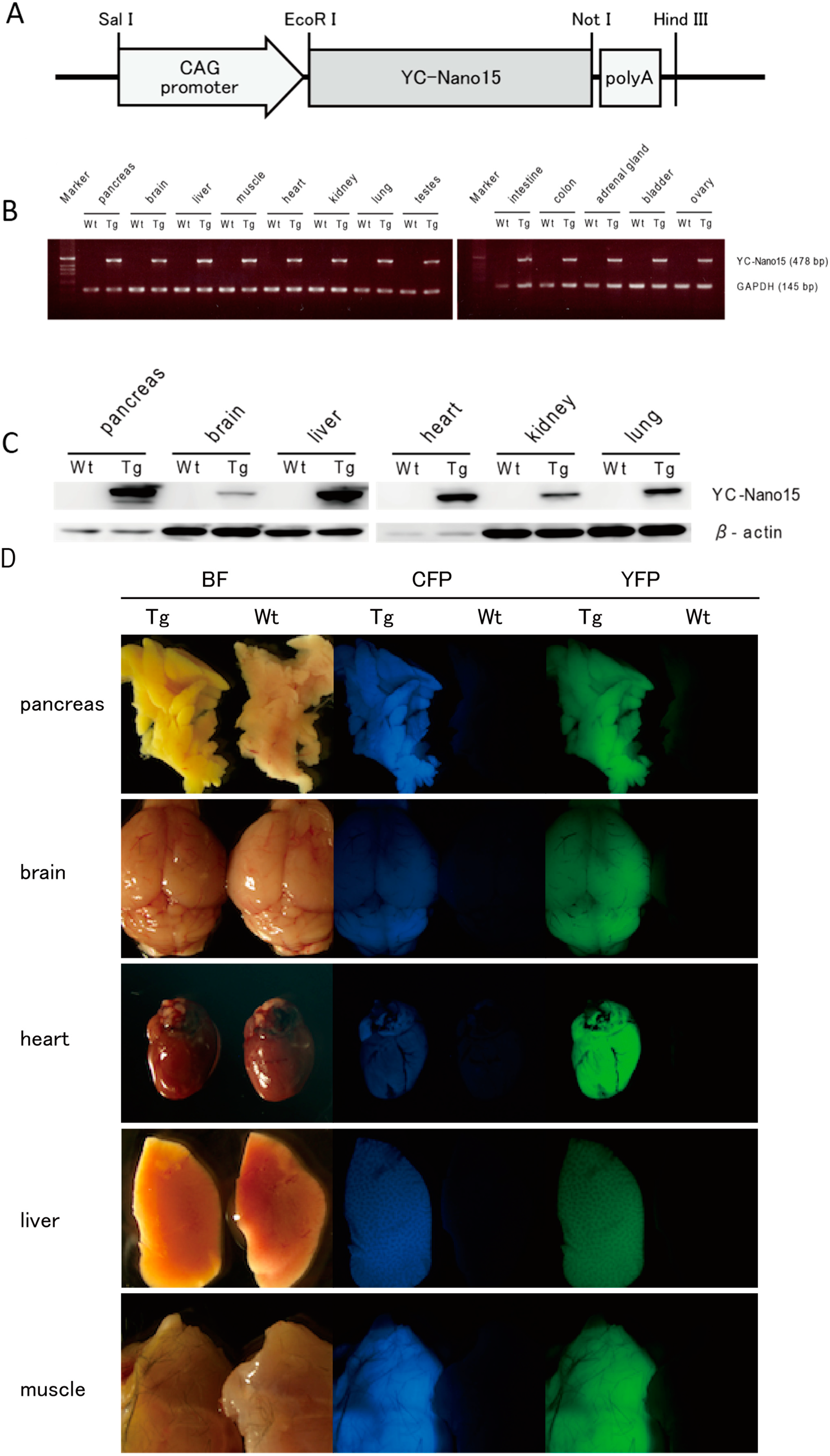
Yellow Cameleon-Nano 15 (YC-Nano15) is expressed ubiquitously in adult organs. (A) The YC-Nano15 transgene construct contains a 1.9-kb YC-Nano15 sequence downstream of the CAG promoter and upstream of the SV40 polyadenylation sequence; (B) Real time-PCR (RT-PCR) analysis of various tissues of YC-Nano15-205 mice revealed that mRNA of YC-Nano15 was expressed ubiquitously in the adult organs. In wild-type (C57BL/6J) mice, no band appeared in a PCR amplification using the YC-Nano15 primers (internal standard: mGAPDH); (C) Western-blot analysis. YC-Nano15 protein is expressed ubiquitously in adult organs; and (D) Fluorescence of various organs in a YC-Nano15-205 mouse. Bright-field (left), CFP fluorescence (middle), and YFP fluorescence images (right) of pancreas, brain, heart, liver, and muscle of a YC-Nano15-205 (Tg) mouse and a C57BL6J (wild-type) mouse are shown. The tissues of YC-Nano15-205 mice are yellower than those of wild-type mice due to expression of YC-Nano15.
Figure 1.
Yellow Cameleon-Nano 15 (YC-Nano15) is expressed ubiquitously in adult organs. (A) The YC-Nano15 transgene construct contains a 1.9-kb YC-Nano15 sequence downstream of the CAG promoter and upstream of the SV40 polyadenylation sequence; (B) Real time-PCR (RT-PCR) analysis of various tissues of YC-Nano15-205 mice revealed that mRNA of YC-Nano15 was expressed ubiquitously in the adult organs. In wild-type (C57BL/6J) mice, no band appeared in a PCR amplification using the YC-Nano15 primers (internal standard: mGAPDH); (C) Western-blot analysis. YC-Nano15 protein is expressed ubiquitously in adult organs; and (D) Fluorescence of various organs in a YC-Nano15-205 mouse. Bright-field (left), CFP fluorescence (middle), and YFP fluorescence images (right) of pancreas, brain, heart, liver, and muscle of a YC-Nano15-205 (Tg) mouse and a C57BL6J (wild-type) mouse are shown. The tissues of YC-Nano15-205 mice are yellower than those of wild-type mice due to expression of YC-Nano15.
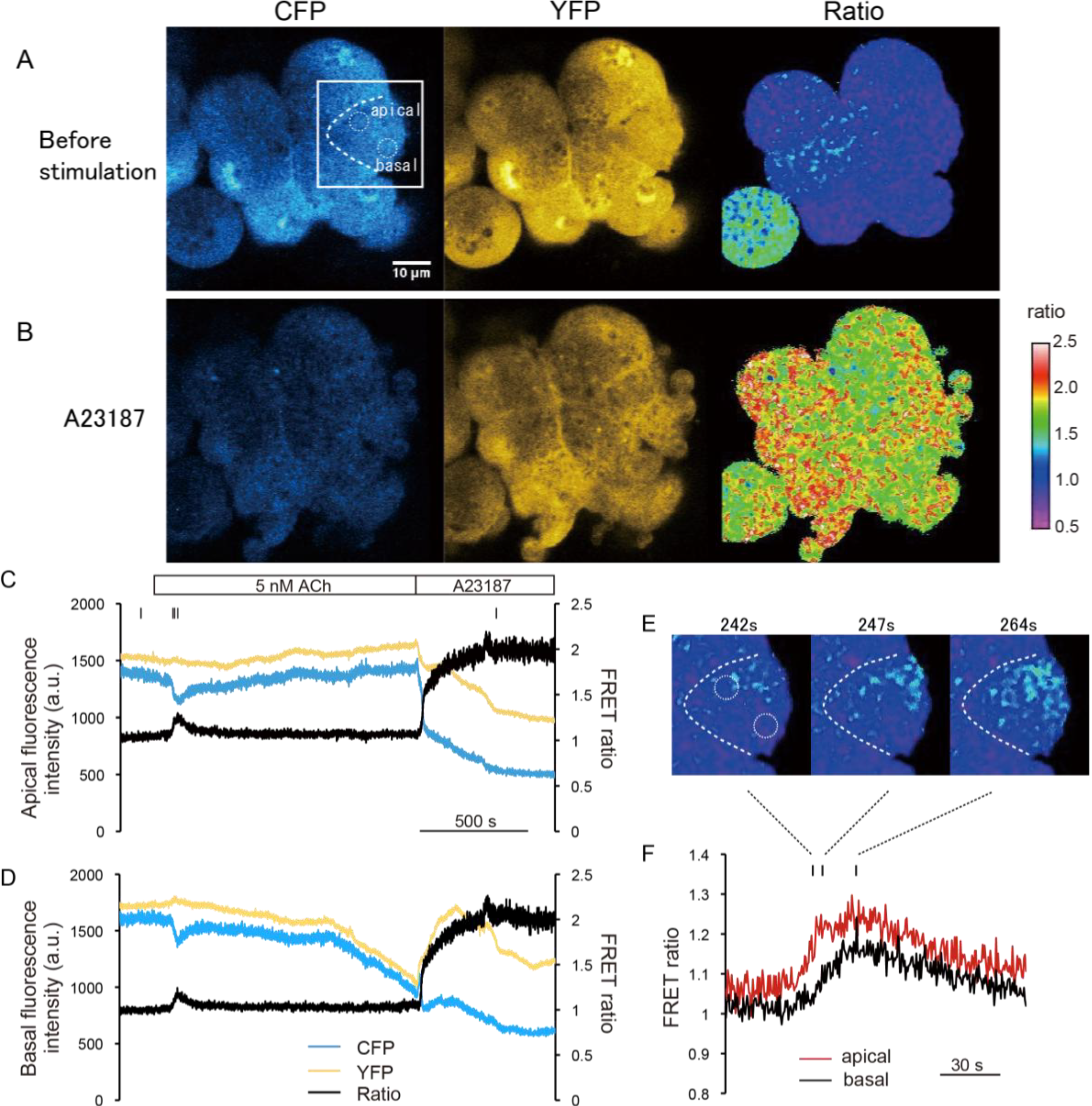
Next, we carried out a time-course analysis in the apical and basal regions of cells (
Figure 2C–F). In this experiment, acinar cells were exposed to agonist solution containing 5 nM ACh, followed by an ionophore solution containing 1 mM [Ca
2+]. The FRET ratio transiently increased just after the ACh stimulus (initial peak), and settled to a slightly higher level than before the stimulus (sustained plateau) in both apical and the basal regions (
Figure 2C,D). In
Figure 2C,D, YFP signal slightly changed reciprocally to the donor signal (CFP). Theoretically, FRET can be quantified solely from either donor or acceptor signals, but practically retiometric analysis contrasting changes in both donor and acceptor signals are useful to exclude the possibility that the signal changes are caused by changes in concentration of the FRET probe or by drift in the focal plane. In this case, since the resting [Ca
2+] estimated to be 100 nM is relatively higher than
Kd value of YC-Nano (15 nM), the dynamic range of CFP and/or YFP signal intensity is thought to be small in compared to
in vitro condition [
3]. The onset of the initial peak was slightly faster in the apical region,
i.e., the increase of [Ca
2+] originated in the apical region and propagated to the basal region (
Figure 2E,F). This result is consistent with a previous report that ACh stimulation induces the formation of apicobasal Ca
2+ waves in acini of pancreatic acinar cells [
17] and of guinea pig nasal glands [
18]. Subsequently, the cells were exposed to Ca
2+/A23187 solution, and the FRET ratio became homogenously saturate (
Figure 2C,D). The value of the saturated FRET ratio (
Rmax) was estimated as 2.01 ± 0.08 (mean ± SE;
n = 6 cells).
Figure 2.
Dynamic range of the fluorescence ratio of cyan and yellow fluorescent proteins (CFP and YFP) (Förster resonance energy transfer ratio, FRET ratio) in pancreas acinar cells of YC-Nano15-205 mice. (A,B) Change in FRET signal following ionophore application. Two-photon excited fluorescence images of CFP and YFP, and fluorescence resonance energy transfer (FRET) ratio (YFP/CFP) image (right) before (A, −57 s) and after (B, 1557 s) stimulation with Ca2+/A23187 solution are shown; these timings are also indicated as small bars at the top of (C). In both (A) and (B), the FRET ratio is distributed homogenously in the cell; (C,D) Time course of CFP and YFP signals and FRET ratio in the “apical” and “basal” circles in the cell shown inside withe box in (A). After stimulation with a minimum concentration of 5 nM acetylcholine (ACh), the FRET ratio increased slightly; (E,F) Detailed view of FRET signal changes upon stimulation of 5 nM ACh in the same experimental trial shown in (A–D). Timings of the images in (E) correspond to the small bars at the top of (F). Unidirectional propagation of the [Ca2+] increase from the apical to the basal region of the cell is shown in (E); and (G) Time course of changes in the FRET ratio in the apical and basal regions of the cell shown in (A).
Figure 2.
Dynamic range of the fluorescence ratio of cyan and yellow fluorescent proteins (CFP and YFP) (Förster resonance energy transfer ratio, FRET ratio) in pancreas acinar cells of YC-Nano15-205 mice. (A,B) Change in FRET signal following ionophore application. Two-photon excited fluorescence images of CFP and YFP, and fluorescence resonance energy transfer (FRET) ratio (YFP/CFP) image (right) before (A, −57 s) and after (B, 1557 s) stimulation with Ca2+/A23187 solution are shown; these timings are also indicated as small bars at the top of (C). In both (A) and (B), the FRET ratio is distributed homogenously in the cell; (C,D) Time course of CFP and YFP signals and FRET ratio in the “apical” and “basal” circles in the cell shown inside withe box in (A). After stimulation with a minimum concentration of 5 nM acetylcholine (ACh), the FRET ratio increased slightly; (E,F) Detailed view of FRET signal changes upon stimulation of 5 nM ACh in the same experimental trial shown in (A–D). Timings of the images in (E) correspond to the small bars at the top of (F). Unidirectional propagation of the [Ca2+] increase from the apical to the basal region of the cell is shown in (E); and (G) Time course of changes in the FRET ratio in the apical and basal regions of the cell shown in (A).
![Ijms 15 19971 g002]()
We performed another time-course analysis to determine the lower limit of the FRET ratio (
Rmin) of YC-Nano15 (
Figure 2G). In this experiment, the extracellular fluid was first replaced with a Ca
2+-free buffer that did not affect the FRET ratio of the cell. When this fluid was replaced with Ca
2+-free EGTA buffer containing CPA and FCCP (indicated as “EGTA”), the FRET ratio rapidly increased, and then gradually decrease to a final value of 1.63 ± 0.04 (mean ± SE;
n = 5 cells), probably reflecting release of Ca
2+ from intracellular storage to the cytoplasm, followed by its leakage to the outside. Next, the fluid was replaced with Ca
2+-free BAPTA-AM buffer (indicated as “BAPTA-AM”). In this case, the FRET ratio decreased below the initial value, indicating that the membrane-permeable chelator further depleted intracellular Ca
2+. After that, the fluid was replaced with a Ca
2+-containing buffer (indicated as “1 mM Ca
2+”), but the FRET ratio continued decreasing, probably because the effect of the accumulated intracellular chelator surpassed the influx of Ca
2+ from outside. Finally, the fluid was replaced with Ca
2+ ionophore in Ca
2+-free EGTA buffer (indicated as “A23187”). In this case, since 10 μM BAPTA-AM solution was superfused constantly, Ca
2+-free BAPTA-AM could be sufficiently maintained in the cytosol,
i.e., the intracellular concentration of BAPTA-AM must be enormously higher than that of YC-Nano 15, stoichemiometrically most of YC-Nano 15 could be shifted to the unbounded form. The FRET ratio then further decreased to a plateau of 0.76 ± 0.02 (mean ± SE;
n = 6 cells), reflecting complete depletion of intracellular Ca
2+,
i.e.,
Rmin under
in vivo conditions. This result indicates that YC-Nano15 can sense extremely low levels of intracellular Ca
2+ in mouse tissue. Nevertheless, because the transgenic mice are healthy and the homeostatic mechanism is maintained, the expression level of YC-Nano 15 is estimated to be moderate in the acinar cells. The expression level of YC-Nano 15 is critical to discuss the capability to sense the low [Ca
2+]
in vivo, in addition to that, the issue of the affinity
in vivo is also related to the discussion. We then experimentally estimated the
Kd value of YC-Nano
in vivo, for instance, considering the ratio change by the following calibration Equation (1) [
5].
Note that
Sf2/Sb2 is the fluorescence signal ratio of the donor channel (CFP) in free and bounded form. In this case,
Sf2/Sb2 is not exactly-determined, but is assumed to be around 3 and over (
Figure 2C,D). The
Kd value of YC-Nano
in vivo is assumed to be to be slightly more than the resting [Ca
2+]. On the other hand, the result also suggests that the expression level of YC-Nano is sufficient for the ratiometric analysis of physiological response to ACh stimulation in the acinar cells, but the affinity of YC-Nano 15
in vivo can vary tremendously according to the cell type and/or where it is expressed, probably reflecting the different degree of interaction with endogenous Ca
2+ binding proteins in different cell types [
13].
2.3. Spatiotemporal Analysis of Intracellular Ca2+ Oscillation and Exocytosis Induced by Stimulation with Agonist (ACh) in YC-Nano15-205 Mice
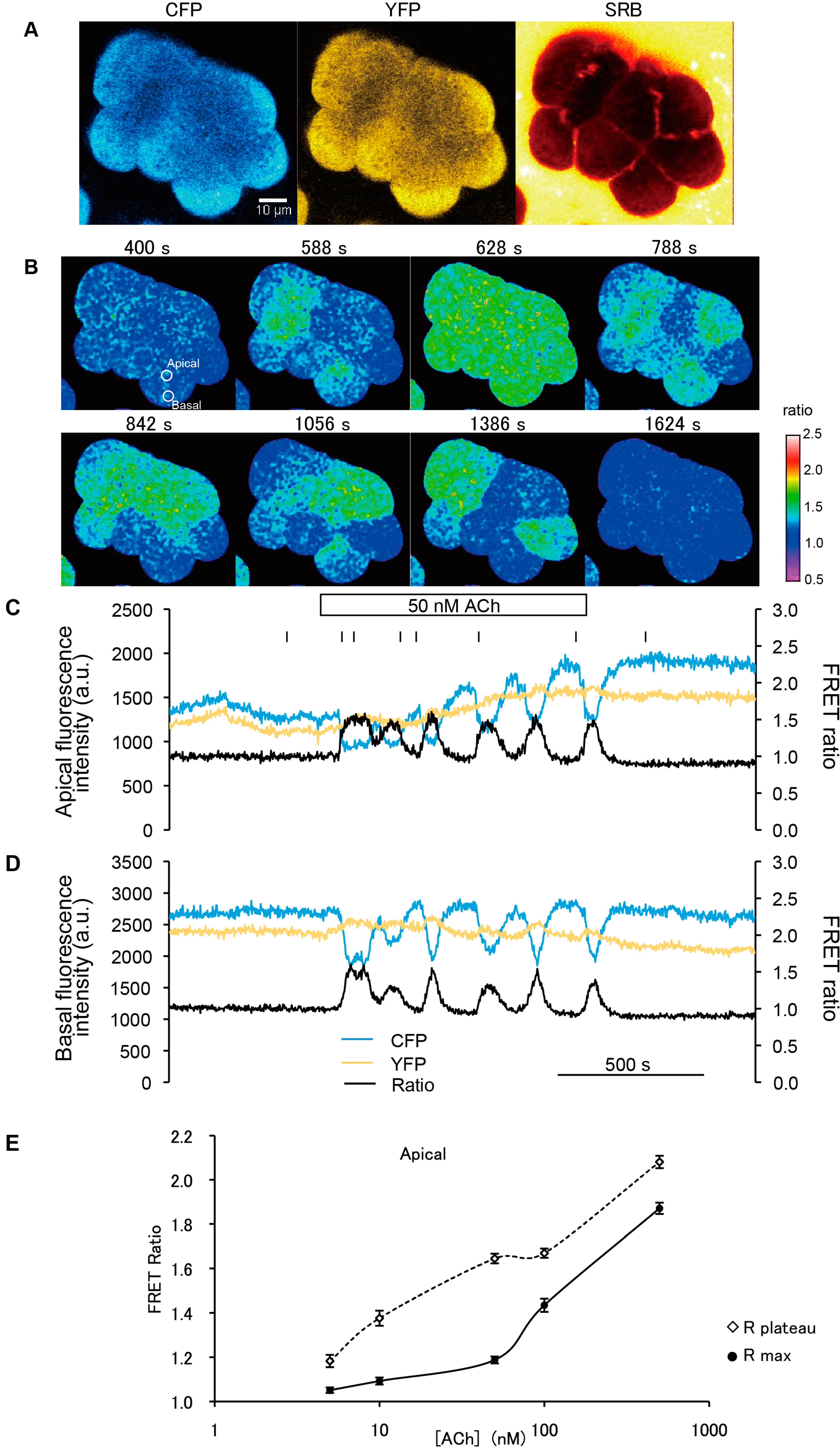
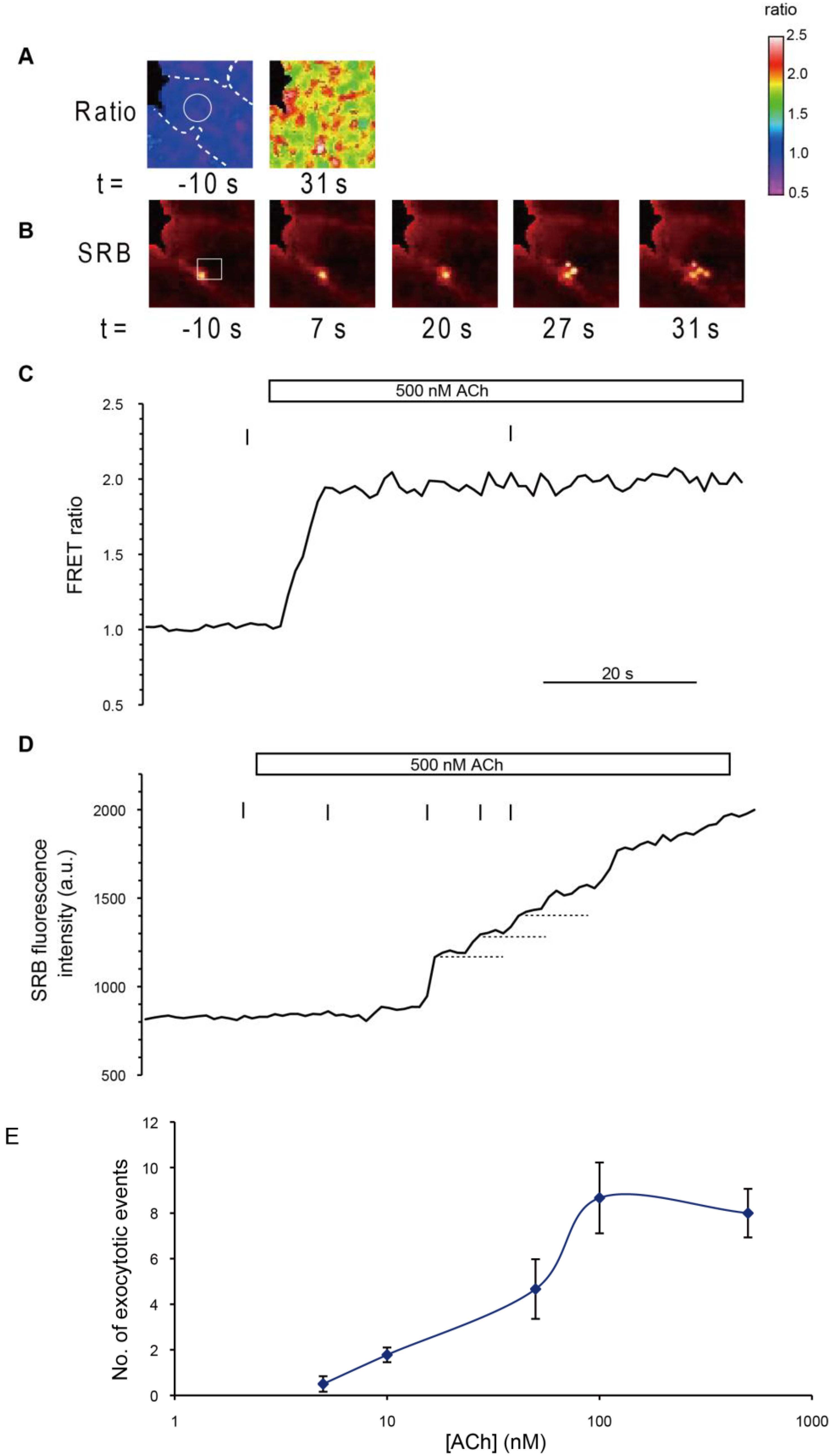
To examine both spatiotemporal Ca
2+ dynamics and typical zymogen granule (ZG) exocytosis in pancreatic acini of YC-Nano15-205 mice, we stimulated isolated acinar cells with agonist (50 nM ACh) in sulforhodamine B (SRB) solution and observed them by two-photon excitation microscopy. Fluorescence signals of CFP, YFP, and SRB were detected simultaneously at an excitation wavelength of 850 nm (
Figure 3A). An increase in intracellular [Ca
2+] was initiated in the apical regions of acini, and [Ca
2+] oscillated during stimulation with agonist (ACh) (
Figure 3B and
Supplementary Movie S1). In this analysis, the FRET ratio shuttled between the baseline and the peak values (
Figure 3C,D). Exocytosis of individual secretory granules in acinar cells was evoked by this stimulation (
Supplementary Movie S2). In acinar cells stimulated with 1–100 µM ACh, Ca
2+ has been reported to transiently increase and reach a plateau [
18]. However, the change in [Ca
2+] induced by stimulation with ACh at concentrations less than 1 µM is difficult to characterize, because of the sensitivity limits of conventional synthetic Ca
2+ indicators. In the previous report [
14], both Ca
2+ transients and exocytosis evoked by less than 50 nM ACh were observed in pancreatic acinar cells by using Fura-2FF (
Kd = 40 µM) and Fura-2 (
Kd = 200 nM) [
14], but conventional synthetic Ca
2+ indicators could not capture detailed behavior of intracellular [Ca
2+] such as Ca
2+ oscillation due to the limitation of the sensitivity. In this study, for the first time, we were able to successfully visualize Ca
2+ oscillation under stimulation with agonist (ACh) at nanomolar levels in pancreatic acini of YC-Nano15 mice.
To determine the dynamic range of YC-Nano15, we also monitored the dependency of FRET ratios on ACh concentration. Acinar cells were stimulated at ACh concentrations of 5, 10, 50, 100, and 500 nM, and FRET ratios in the initial peak and subsequent plateau were calculated and plotted (
Figure 3E). In the previous report by Horikawa
et al., the ratiometric FRET imaging of YC-Nano 15 was performed in
Dictyostlium [
11]
. The FRET signal change (Δ
R/
R= (9.5 − 5.0)/5.0 × 100% = 90%) was showed
in vivo condition. In the result, though we measured in pancreatic acinar cells of mice, the dynamic range (Δ
R/R = (2.01 − 0.76)/0.76 × 100% = 165%) of YC-nano15 was calculated from
Rmax and
Rmin values under
in vivo condition. The relationship between ACh concentration and FRET ratio was consistent with results described in previous reports [
3,
14]. Thus, the dynamic range of YC-Nano15 in mouse tissue is around 5–100 nM.
Figure 3.
Acetylcholine (Ach)-induced Ca2+ oscillations revealed by FRET signals in two-photon excitation fluorescence images. (A) Two-photon excitation fluorescence images of CFP (left) and YFP (middle), and cross-section image in SRB solution to monitor exocytotic events (right), before stimulation (−144 s; time origin is the time at which the agonist was applied); (B) FRET images showing repeated Ca2+ waves from the apical to basal regions during 50 nM ACh stimulation; (C,D) Time course of changes in CFP, YFP signals, and FRET ratio in apical (C) and basal (D) regions in a cell. Regions correspond to the “apical” and “basal” circles in (B). Stimulation with a physiological agonist concentration of 50 nM ACh induced repeated unidirectional Ca2+ oscillations. After the agonist was washed out, the value of the FRET ratio returned to the initial level; and (E) Dose dependency of the FRET ratio on ACh concentration. In the apical region of acinar cells, the maximum peak of the FRET ratio increases as a function of acetylcholine concentration (white diamonds). FRET ratio values in sustained plateau during stimulation were plotted against the ACh concentration (black dots), although in some cases no plateau was observed (e.g., C and D, stimulated with 50 nM ACh). In those cases, FRET ratio values at the bottom of the oscillation curve were measured and plotted instead. Error bars represent S.E. The number of cells measured (n) at each concentration was as follows: 6 (5 nM), 17 (10 nM), 7 (50 nM), 8 (100 nM), and 10 (500 nM).
Figure 3.
Acetylcholine (Ach)-induced Ca2+ oscillations revealed by FRET signals in two-photon excitation fluorescence images. (A) Two-photon excitation fluorescence images of CFP (left) and YFP (middle), and cross-section image in SRB solution to monitor exocytotic events (right), before stimulation (−144 s; time origin is the time at which the agonist was applied); (B) FRET images showing repeated Ca2+ waves from the apical to basal regions during 50 nM ACh stimulation; (C,D) Time course of changes in CFP, YFP signals, and FRET ratio in apical (C) and basal (D) regions in a cell. Regions correspond to the “apical” and “basal” circles in (B). Stimulation with a physiological agonist concentration of 50 nM ACh induced repeated unidirectional Ca2+ oscillations. After the agonist was washed out, the value of the FRET ratio returned to the initial level; and (E) Dose dependency of the FRET ratio on ACh concentration. In the apical region of acinar cells, the maximum peak of the FRET ratio increases as a function of acetylcholine concentration (white diamonds). FRET ratio values in sustained plateau during stimulation were plotted against the ACh concentration (black dots), although in some cases no plateau was observed (e.g., C and D, stimulated with 50 nM ACh). In those cases, FRET ratio values at the bottom of the oscillation curve were measured and plotted instead. Error bars represent S.E. The number of cells measured (n) at each concentration was as follows: 6 (5 nM), 17 (10 nM), 7 (50 nM), 8 (100 nM), and 10 (500 nM).
![Ijms 15 19971 g003]()

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}