Lactoferrin Directly Scavenges Hydroxyl Radicals and Undergoes Oxidative Self-Degradation: A Possible Role in Protection against Oxidative DNA Damage

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
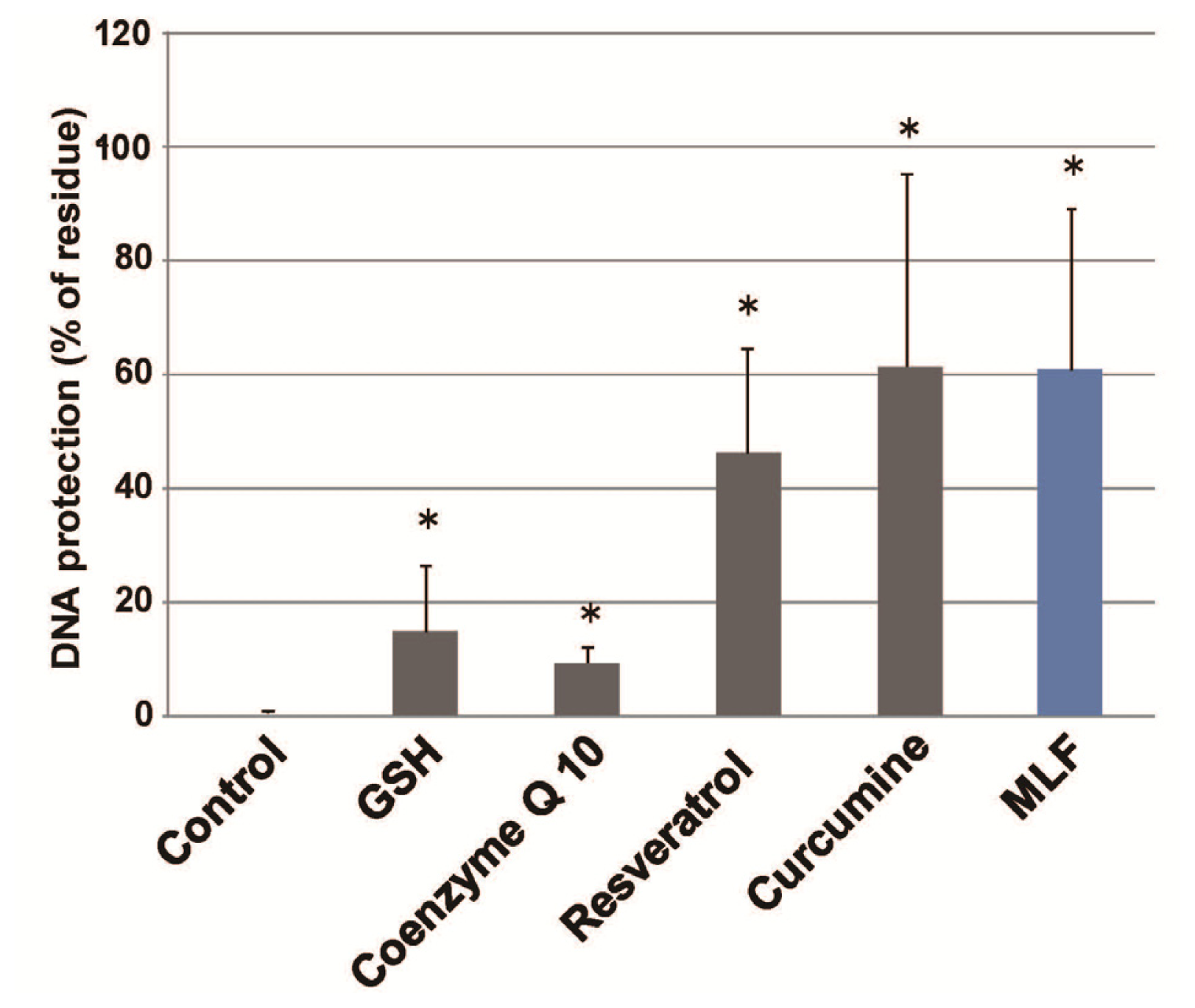
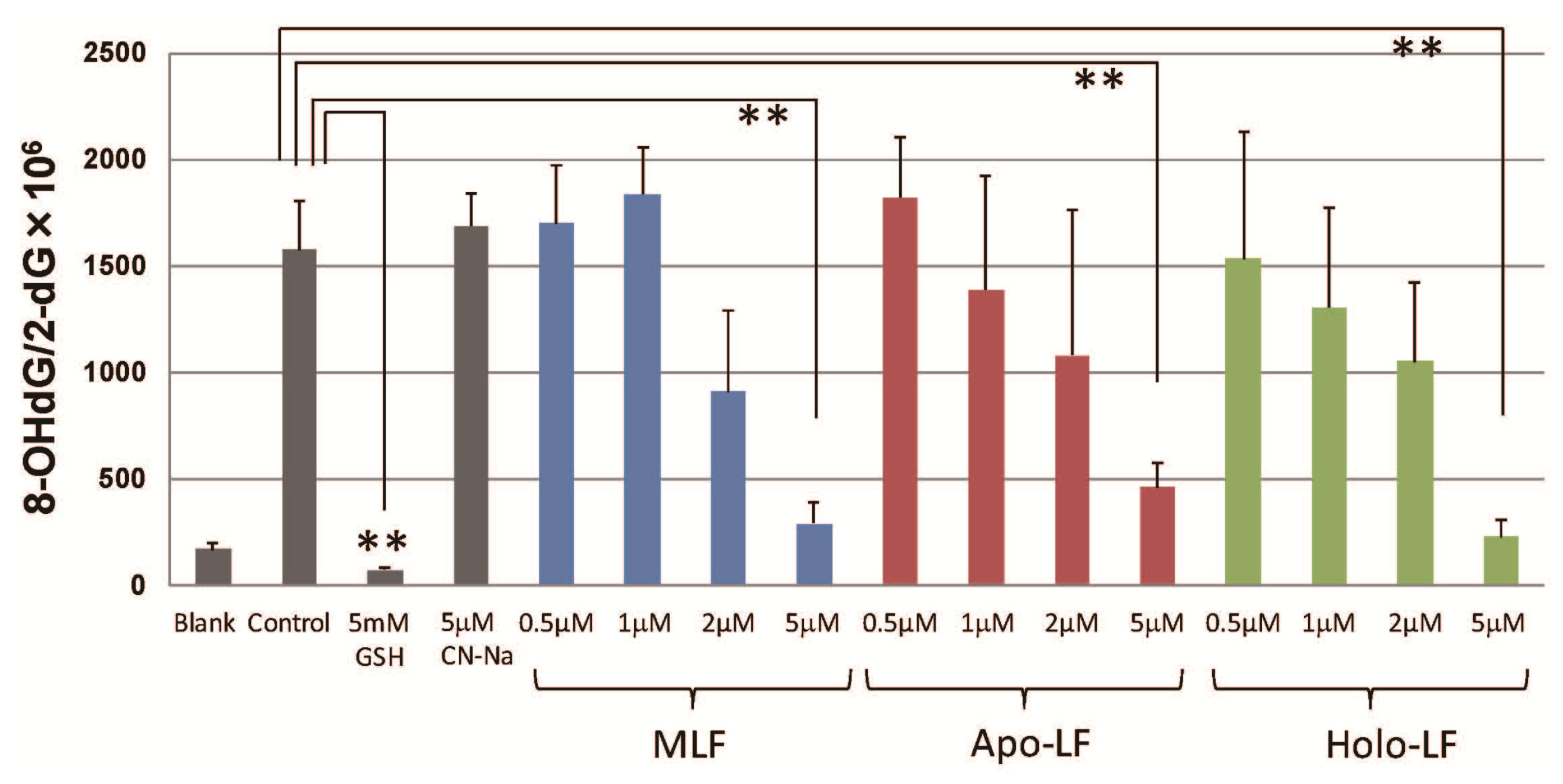
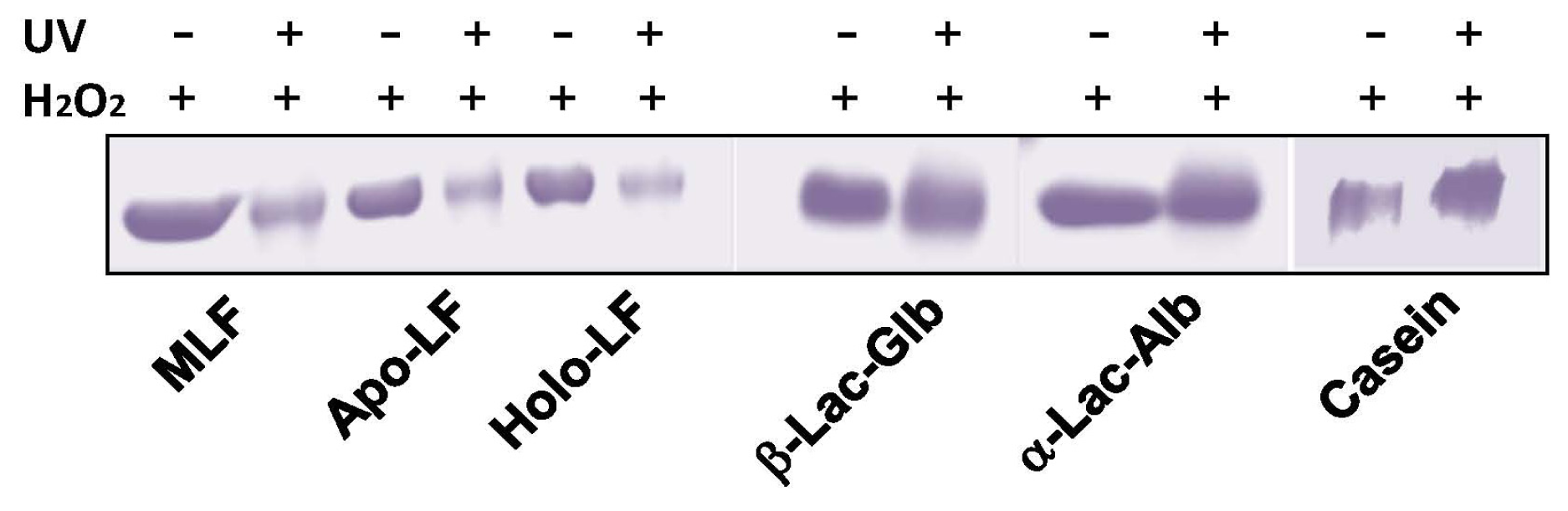
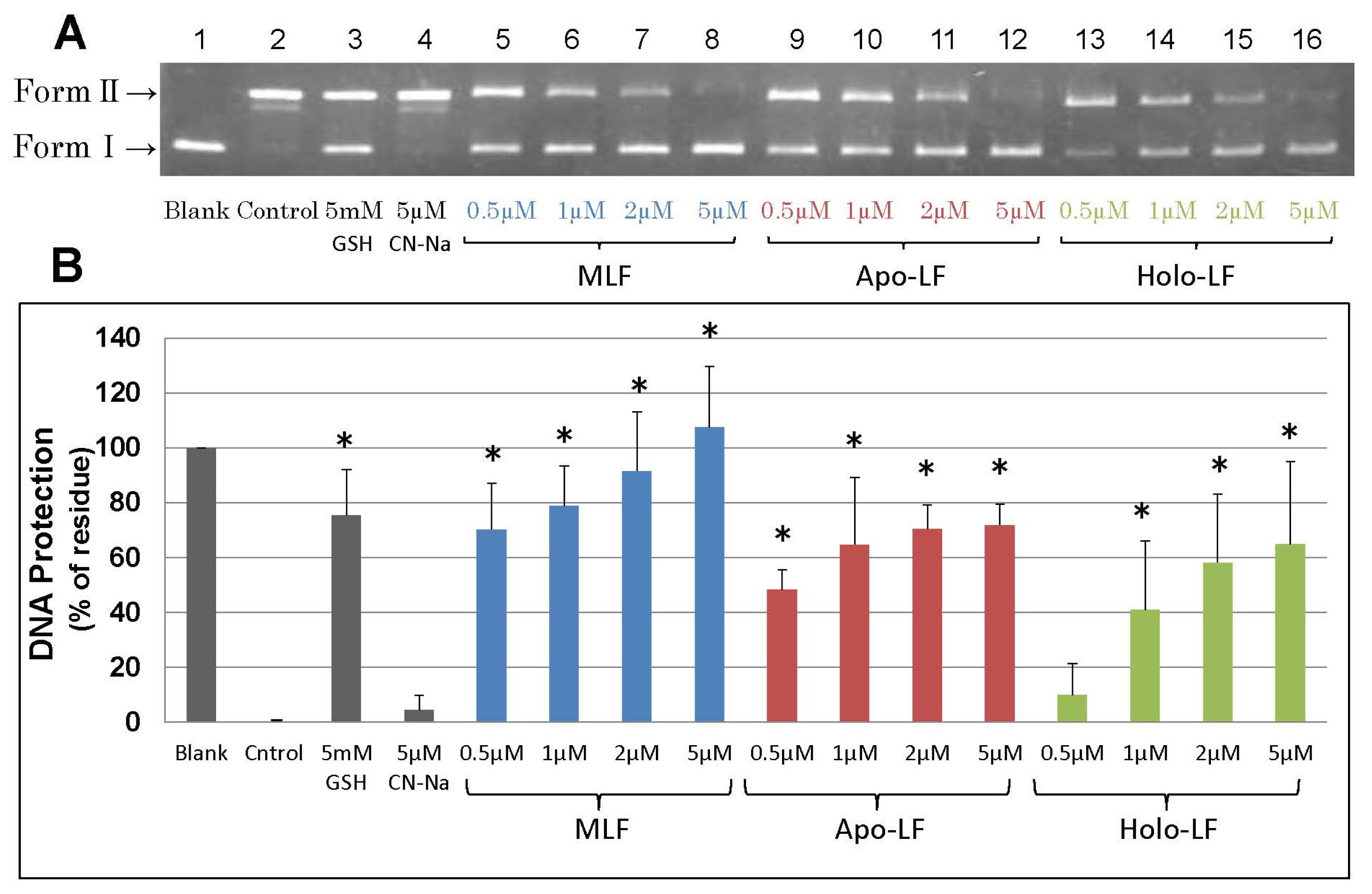
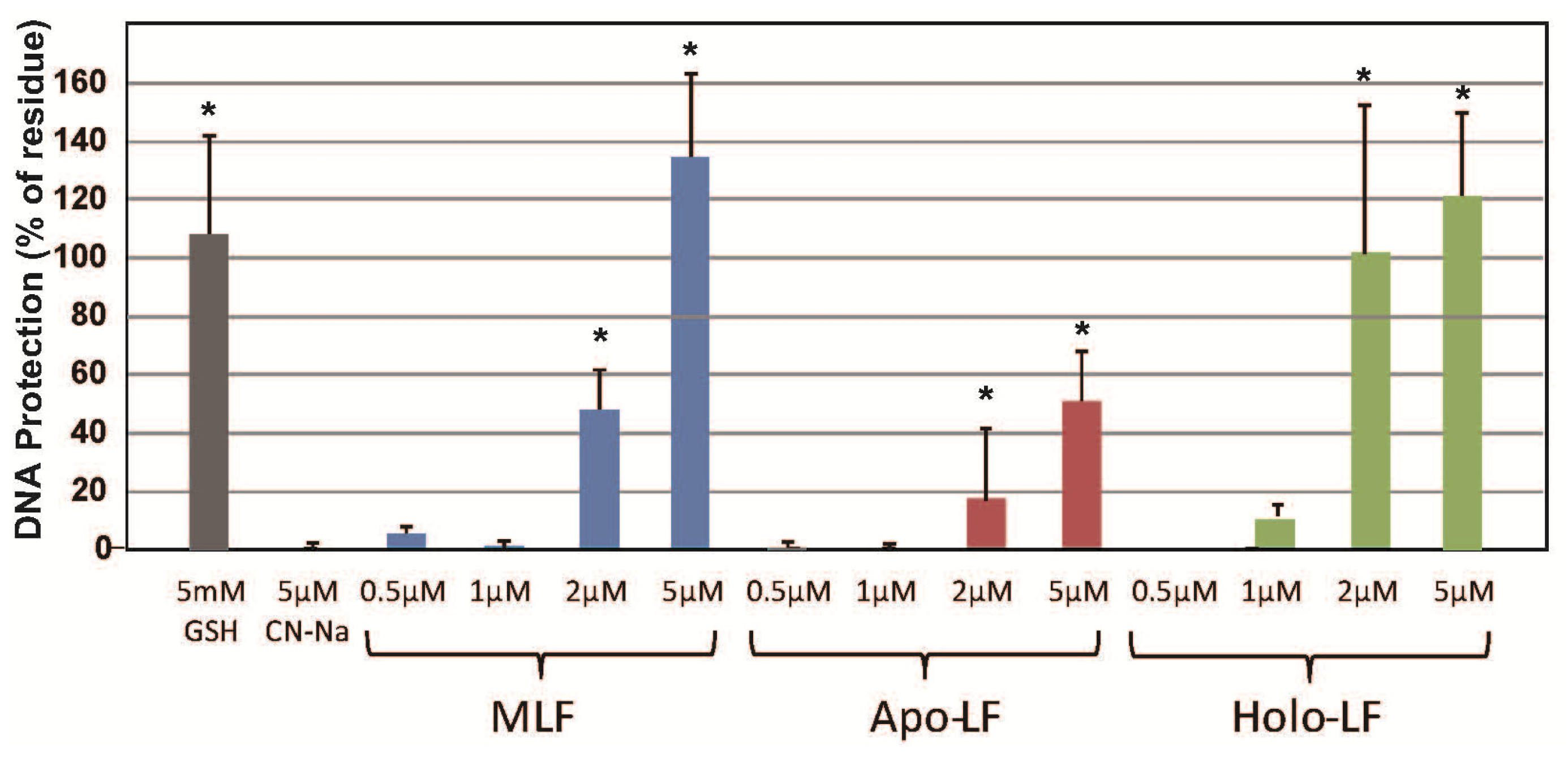
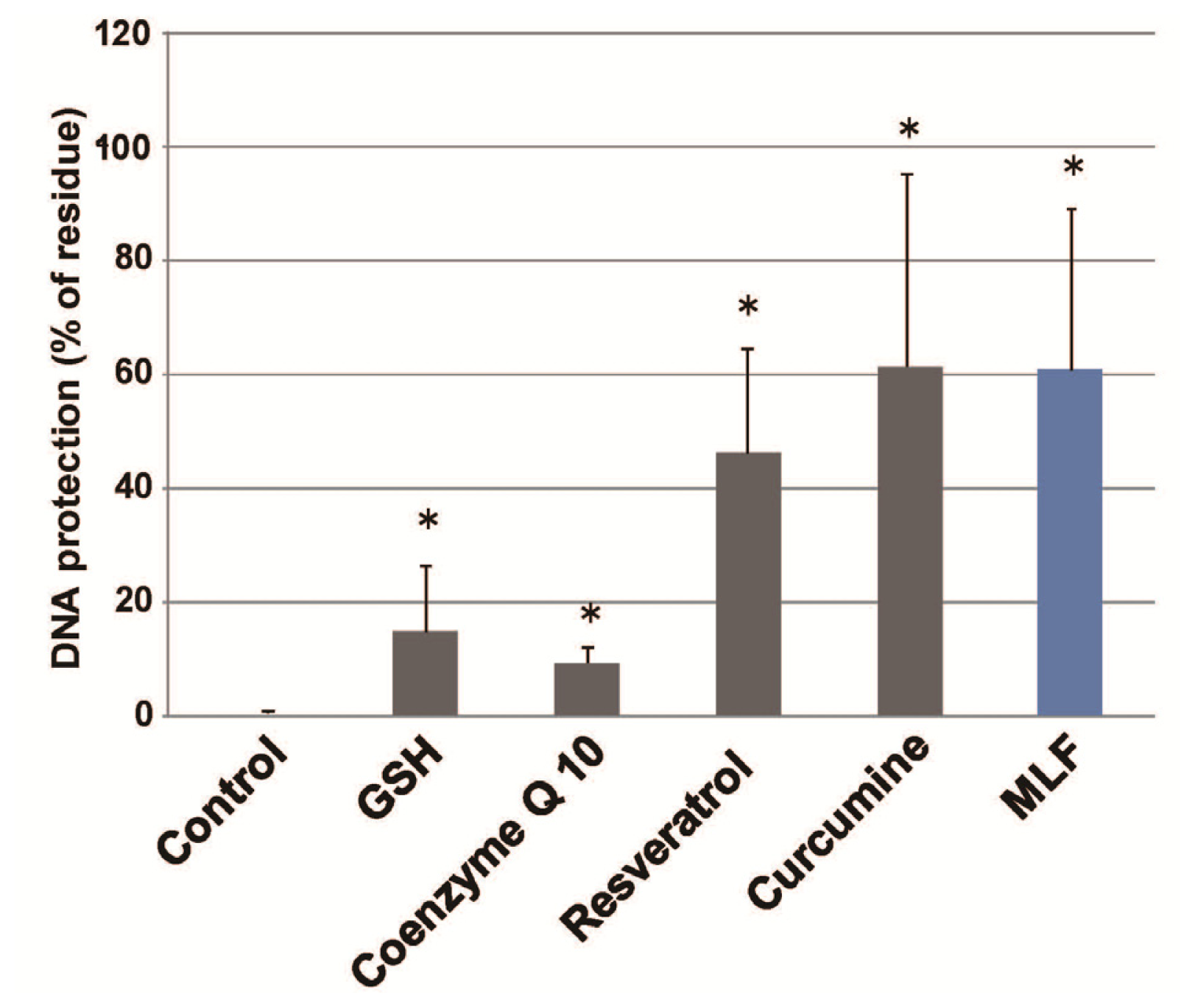
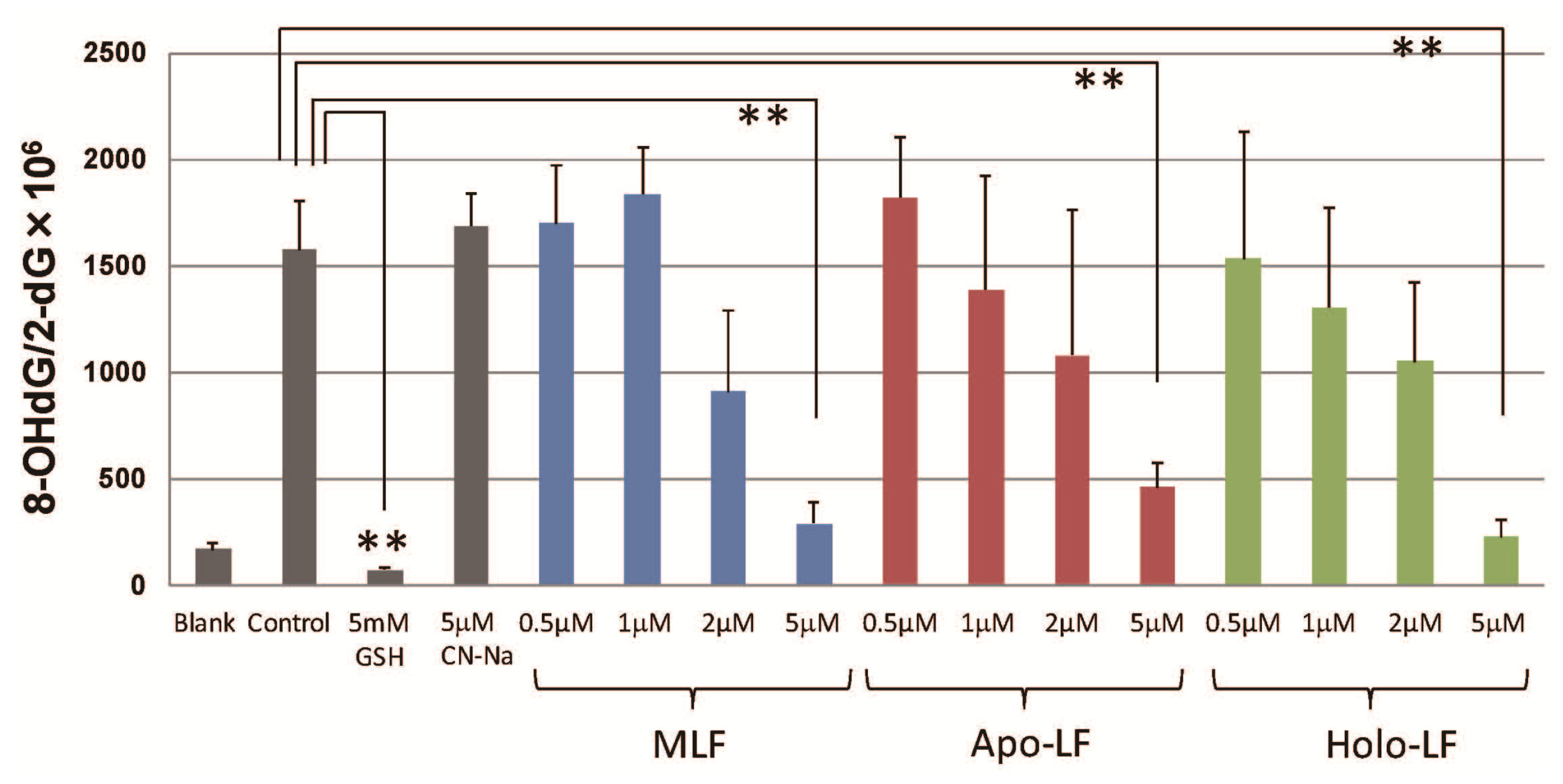
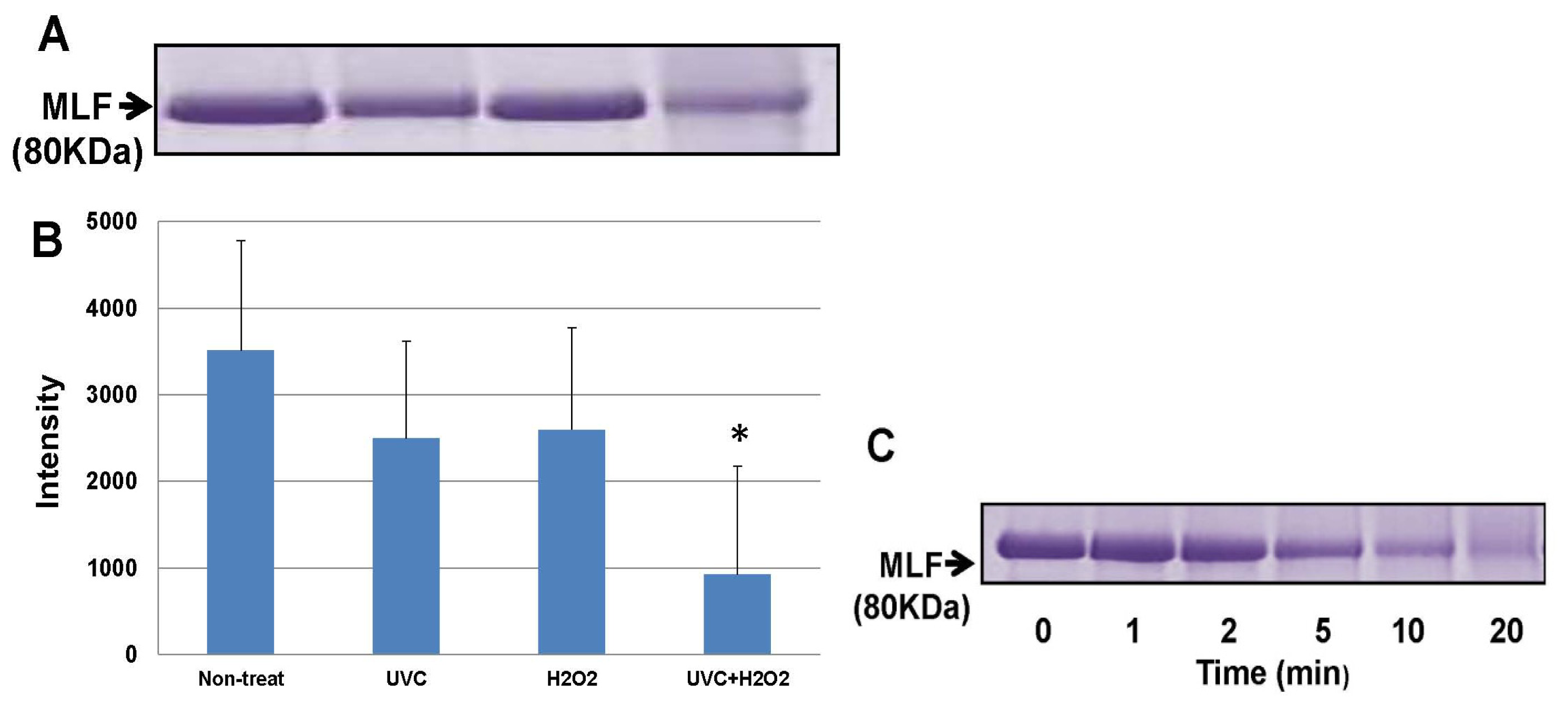
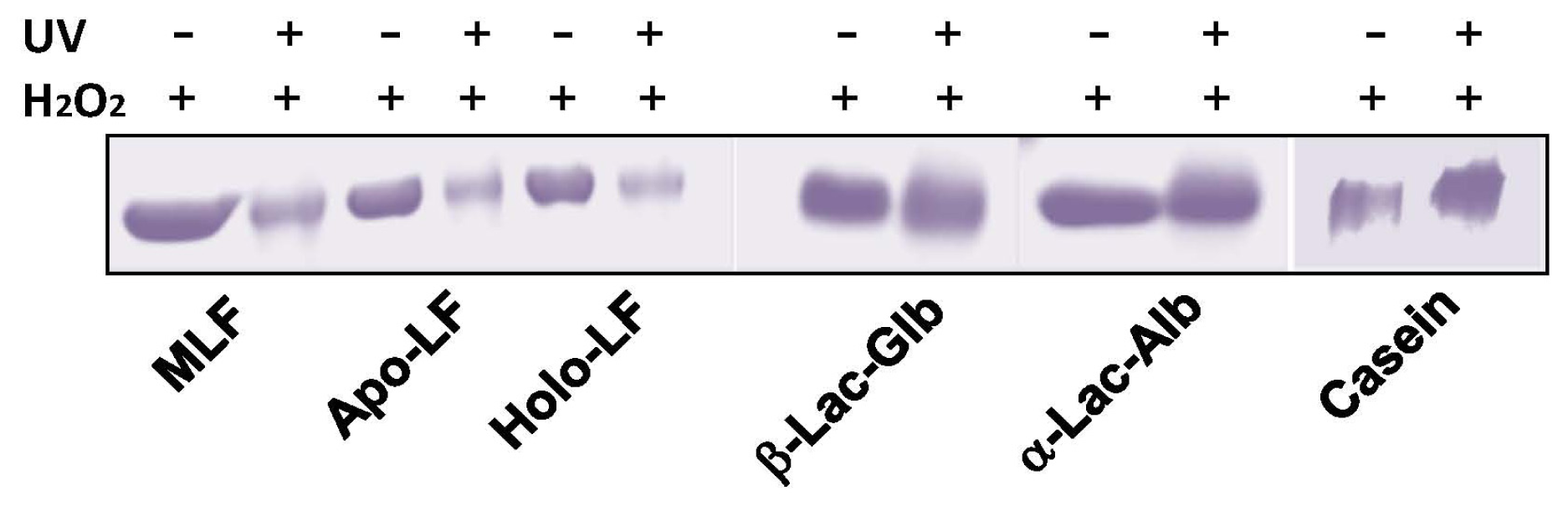
2. Results
3. Discussion
4. Experimental Section
4.1. Materials
4.2. DNA Double Strand Breaks
4.3. UV Irradiation of Plasmid DNA and Calf Thymus DNA
4.4. HPLC-EC Analysis of 8-OHdG within DNA
4.5. Oxidative Alteration of LF by Exposure to Hydroxyl Radicals
4.6. Statistical Analysis
5. Conclusions
Conflicts of Interest
Abbreviations
| LF | lactoferrin |
| EDTA | ethylenediaminetetraacetic acid |
| ROS | reactive oxygen species |
| 8-OHdG | 8-hydroxydeoxyguanosine |
| holo-LF | iron-saturated lactoferrin |
| apo-LF | apolactoferrin |
| MLF | native milk lactoferrin |
References
- Schanbacher, F.L.; Goodman, R.E.; Talhouk, R.S. Bovine mammary lactoferrin: Implications from messenger ribonucleic acid (mRNA) sequence and regulation contrary to other milk proteins. J. Dairy. Sci 1993, 76, 3812–3831. [Google Scholar]
- van der Strate, B.W.; Beljaars, L.; Molema, G.; Harmsen, M.C.; Meijer, D.K. Antiviral activities of lactoferrin. Antiviral Res 2001, 52, 225–239. [Google Scholar]
- Bennett, R.M.; Kokocinski, T. Lactoferrin content of peripheral blood cells. Br. J. Haematol 1978, 39, 509–521. [Google Scholar]
- Caccavo, D.; Pellegrino, N.M.; Altamura, M.; Rigon, A.; Amati, L.; Amoroso, A.; Jirillo, E. Antimicrobial and immunoregulatory functions of lactoferrin and its potential therapeutic application. J. Endotoxin Res 2002, 8, 403–417. [Google Scholar]
- Conneely, O.M. Antiinflammatory activities of lactoferrin. J. Am. Coll. Nutr 2001, 20, 389S–395S. [Google Scholar]
- Baveye, S.; Elass, E.; Mazurier, J.; Spik, G.; Legrand, D. Lactoferrin: A multifunctional glycoprotein involved in the modulation of the inflammatory process. Clin. Chem. Lab. Med 1999, 37, 281–286. [Google Scholar]
- Baynes, R.D.; Bezwoda, W.R. Lactoferrin and the inflammatory response. Adv. Exp. Med. Biol 1994, 357, 133–141. [Google Scholar]
- Kruzel, M.L.; Zimecki, M. Lactoferrin and immunologic dissonance: Clinical implications. Arch. Immunol. Ther. Exp 2002, 50, 399–410. [Google Scholar]
- Haber, F.; Weiss, J. On the catalysis of hydroperoxide. Naturwissenschaften 1932, 20, 948–950. [Google Scholar]
- Gutteridge, J.M. Hydroxyl radicals, iron, oxidative stress, and neurodegeneration. Ann. N. Y. Acad. Sci 1994, 738, 201–213. [Google Scholar]
- Pourzand, C.; Tyrrell, R.M. Apoptosis, the role of oxidative stress and the example of solar UV radiation. Photochem. Photobiol 1999, 70, 380–390. [Google Scholar]
- Genestra, M. Oxyl radicals, redox-sensitive signalling cascades and antioxidants. Cell Signal 2007, 19, 1807–1819. [Google Scholar]
- Cheng, K.C.; Cahill, D.S.; Kasai, H.; Nishimura, S.; Loeb, L.A. 8-Hydroxyguanine, an abundant form of oxidative DNA damage, causes G-T and A-C substitutions. J. Biol. Chem 1992, 267, 166–172. [Google Scholar]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-Hydroxy-2′-deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev 2009, 27, 120–139. [Google Scholar]
- Tsuda, H.; Kozu, T.; Iinuma, G.; Ohashi, Y.; Saito, Y.; Saito, D.; Akasu, T.; Alexander, D.B.; Futakuchi, M.; Fukamachi, K.; et al. Cancer prevention by bovine lactoferrin: From animal studies to human trial. Biometals 2010, 23, 399–409. [Google Scholar]
- Kozu, T.; Iinuma, G.; Ohashi, Y.; Saito, Y.; Akasu, T.; Saito, D.; Alexander, D.B.; Iigo, M.; Kakizoe, T.; Tsuda, H. Effect of orally administered bovine lactoferrin on the growth of adenomatous colorectal polyps in a randomized, placebo-controlled clinical trial. Cancer Prev. Res 2009, 2, 975–983. [Google Scholar]
- Rodrigues, L.; Teixeira, J.; Schmitt, F.; Paulsson, M.; Månsson, H.L. Lactoferrin and cancer disease prevention. Crit. Rev. Food Sci. Nutr 2009, 2, 975–983. [Google Scholar]
- Wei, H.; Cai, Q.; Rahn, R.; Zhang, X. Singlet oxygen involvement in ultraviolet (254 nm) radiation-induced formation of 8-hydroxy-deoxyguanosine in DNA. Free Radic. Biol. Med 1997, 23, 148–154. [Google Scholar]
- Wakabayashi, H.; Matsumoto, H.; Hashimoto, K.; Teraguchi, S.; Takase, M.; Hayasawa, H. Inhibition of iron/ascorbate-induced lipid peroxidation by an N-terminal peptide of bovine lactoferrin and its acylated derivatives. Biosci. Biotechnol. Biochem 1999, 63, 955–957. [Google Scholar]
- Bennett, R.M.; Davis, J. Lactoferrin interacts with deoxyribonucleic acid: A preferential reactivity with double-stranded DNA and dissociation of DNA-anti-DNA complexes. J. Lab. Clin. Med 1982, 99, 127–138. [Google Scholar]
- He, J.; Furmanski, P. Sequence specificity and transcriptional activation in the binding of lactoferrin to DNA. Nature 1995, 373, 721–724. [Google Scholar]
- Ivanov, A.A.; Ulanova, A.M.; Stavrakova, N.M.; Deshevoĭ, IuB.; Nasonova, T.A.; Koterov, A.N.; Gutsenko, K.K.; Mal’tsev, V.N. Antiradiation effects of lactoferrin. Radiats. Biol. Radioecol 2009, 49, 456–461. (in Russian). [Google Scholar]
- Akiyama, Y.; Oshima, K.; Shin, K.; Wakabayashi, H.; Abe, F.; Nadano, D.; Matsuda, T. Intracellular retention and subsequent release of bovine milk lactoferrin taken up by human enterocyte-like cell lines, Caco-2, C2BBe1 and HT-29. Biosci. Biotechnol. Biochem 2013, 77, 1023–1029. [Google Scholar]
- Wakabayashi, H.; Yamauchi, K.; Takase, M. Inhibitory effects of bovine lactoferrin and lactoferricin B on Enterobacter sakazakii. Biocontrol Sci 2008, 13, 29–32. [Google Scholar]
- Kukielka, E.; Cederbaum, A.I. DNA strand cleavage as a sensitive assay for the production of hydroxyl radicals by microsomes: Role of cytochrome P4502E1 in the increased activity after ethanol treatment. Biochem. J 1994, 302, 773–779. [Google Scholar]
- Kukielka, E.; Cederbaum, A.I. Stimulation of NADH-dependent microsomal DNA strand cleavage by rifamycin SV. Biochem. J 1995, 307, 361–367. [Google Scholar]
- Asami, S.; Manabe, H.; Miyake, J.; Tsurudome, Y.; Hirano, T.; Yamaguchi, R.; Itoh, H.; Kasai, H. Cigarette smoking induces an increase in oxidative DNA damage, 8-hydroxydeoxyguanosine, in a central site of the human lung. Carcinogenesis 1997, 18, 1763–1766. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ogasawara, Y.; Imase, M.; Oda, H.; Wakabayashi, H.; Ishii, K. Lactoferrin Directly Scavenges Hydroxyl Radicals and Undergoes Oxidative Self-Degradation: A Possible Role in Protection against Oxidative DNA Damage. Int. J. Mol. Sci. 2014, 15, 1003-1013. https://doi.org/10.3390/ijms15011003
Ogasawara Y, Imase M, Oda H, Wakabayashi H, Ishii K. Lactoferrin Directly Scavenges Hydroxyl Radicals and Undergoes Oxidative Self-Degradation: A Possible Role in Protection against Oxidative DNA Damage. International Journal of Molecular Sciences. 2014; 15(1):1003-1013. https://doi.org/10.3390/ijms15011003
Chicago/Turabian StyleOgasawara, Yuki, Megumi Imase, Hirotsugu Oda, Hiroyuki Wakabayashi, and Kazuyuki Ishii. 2014. "Lactoferrin Directly Scavenges Hydroxyl Radicals and Undergoes Oxidative Self-Degradation: A Possible Role in Protection against Oxidative DNA Damage" International Journal of Molecular Sciences 15, no. 1: 1003-1013. https://doi.org/10.3390/ijms15011003