Effect of a Static Magnetic Fields and Fluoride Ions on the Antioxidant Defense System of Mice Fibroblasts

Abstract
:1. Introduction
2. Results and Discussion
2.1. Effects of Fluoride on Mice Fibroblast Cultures
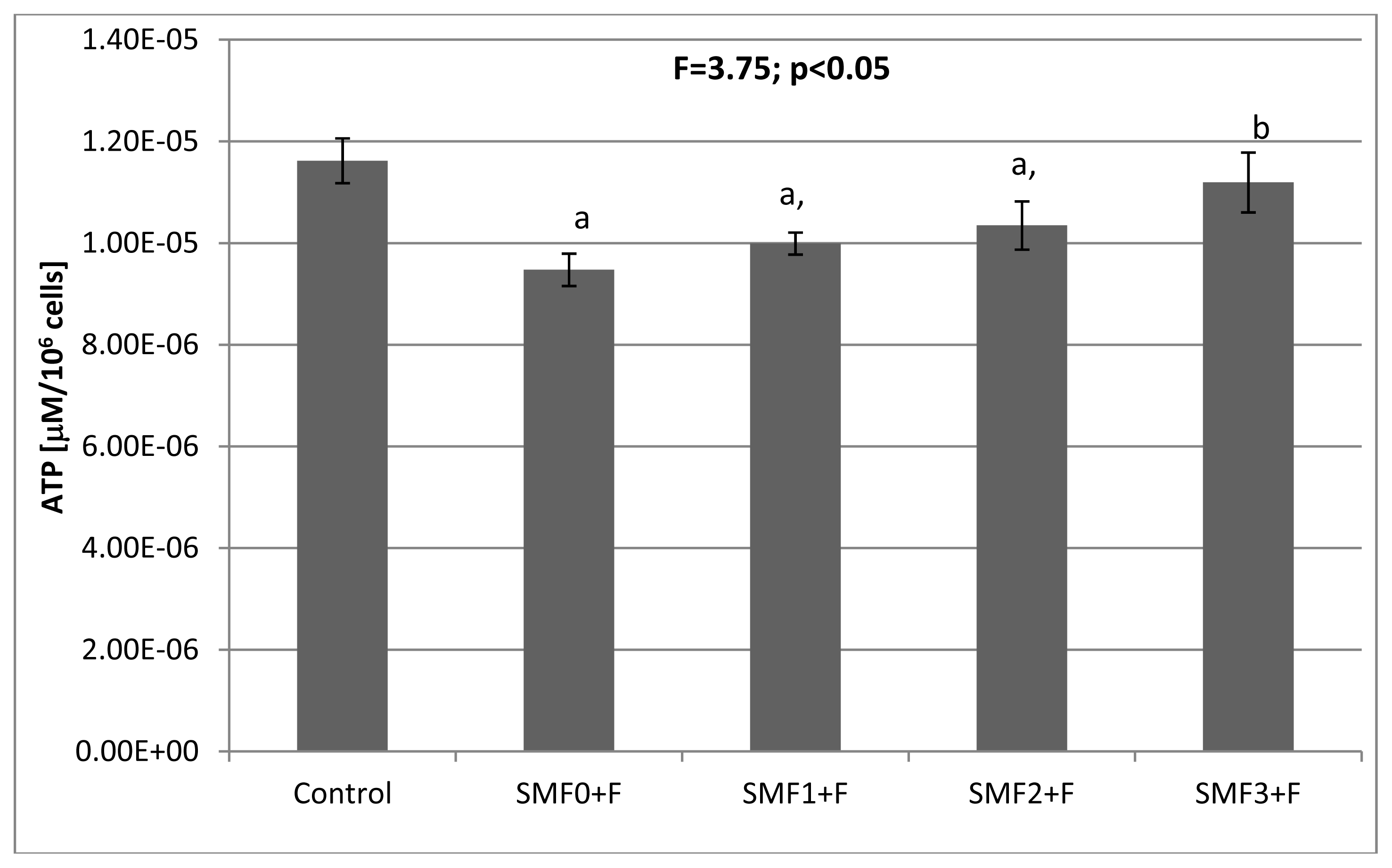
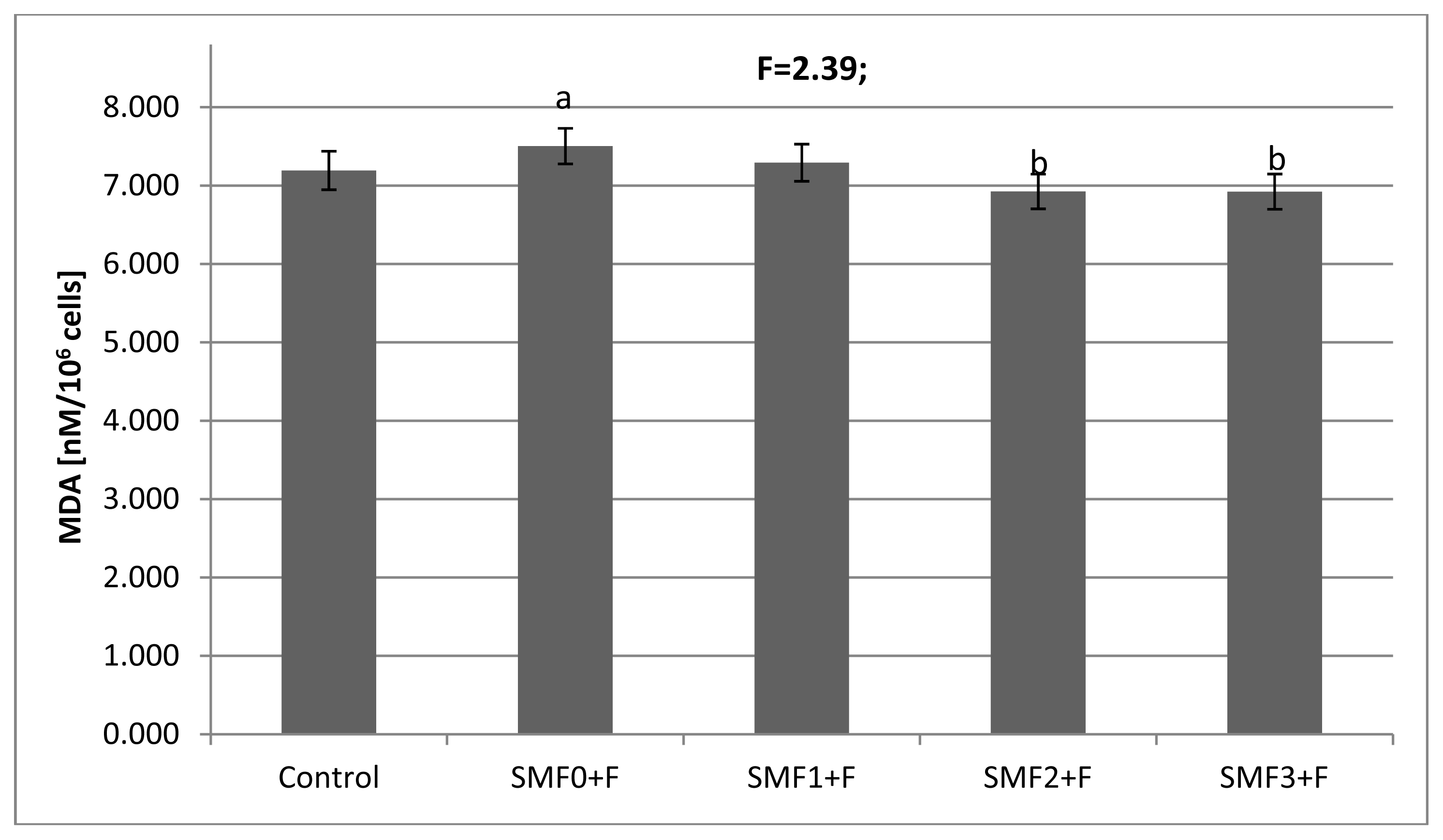
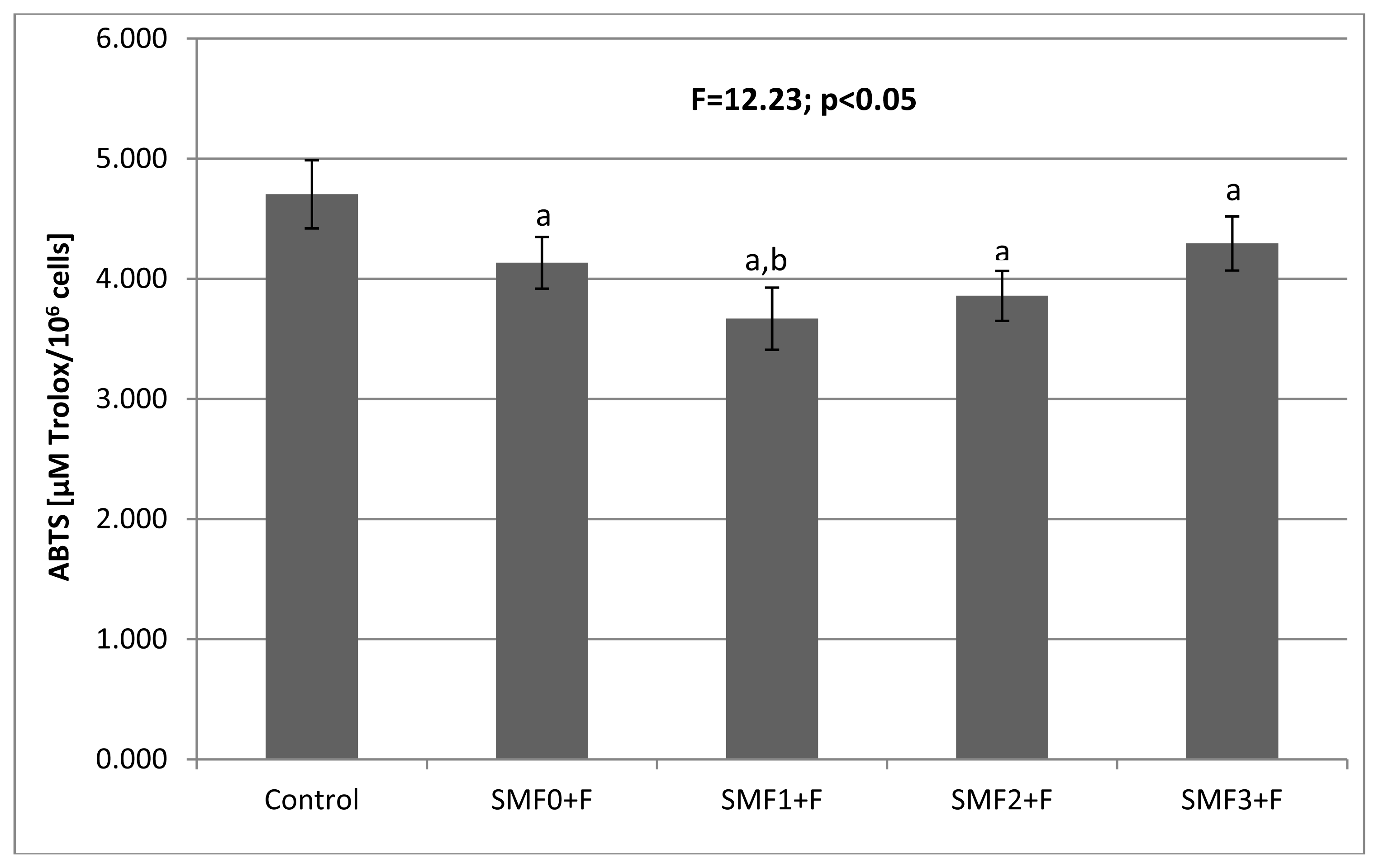
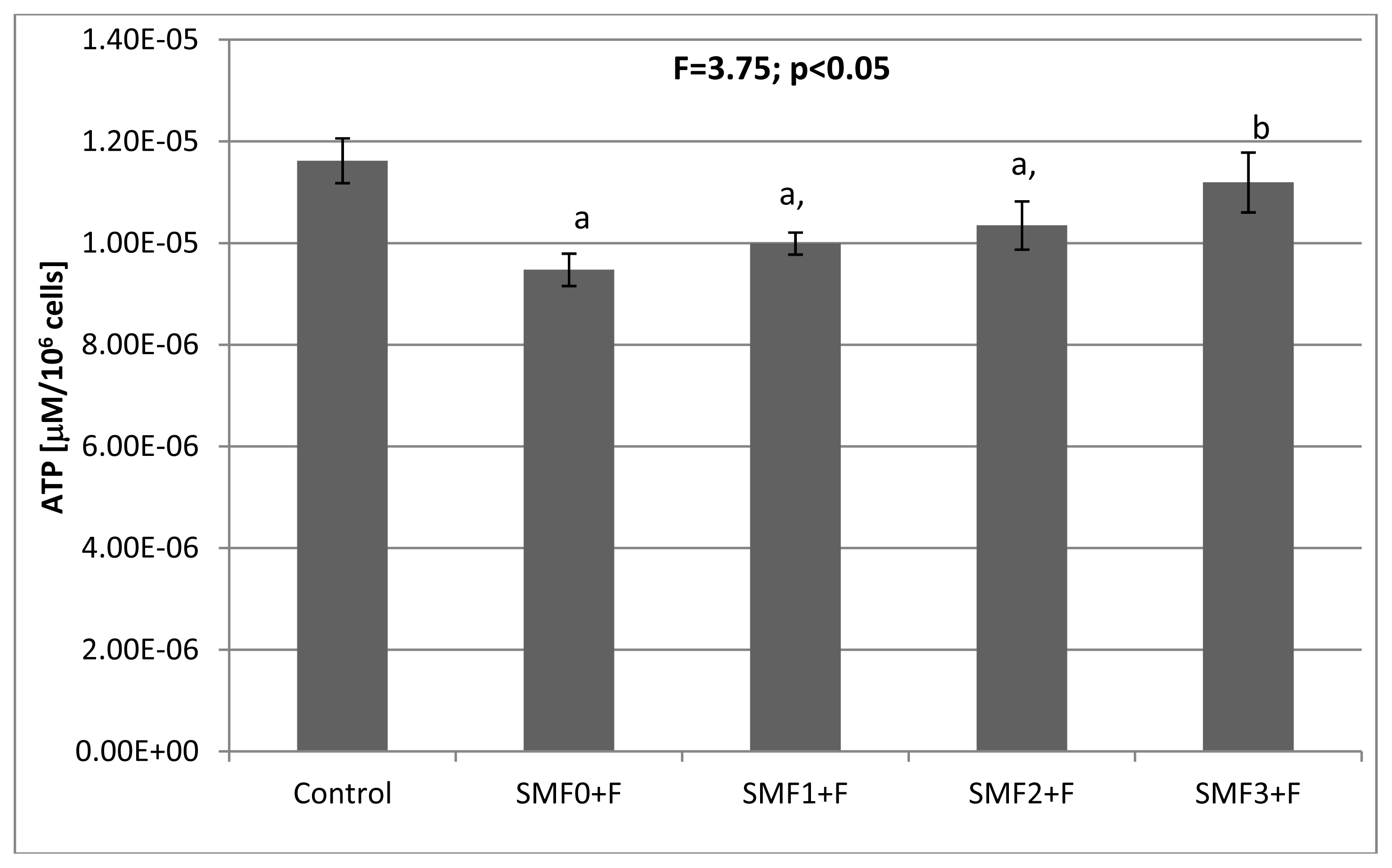
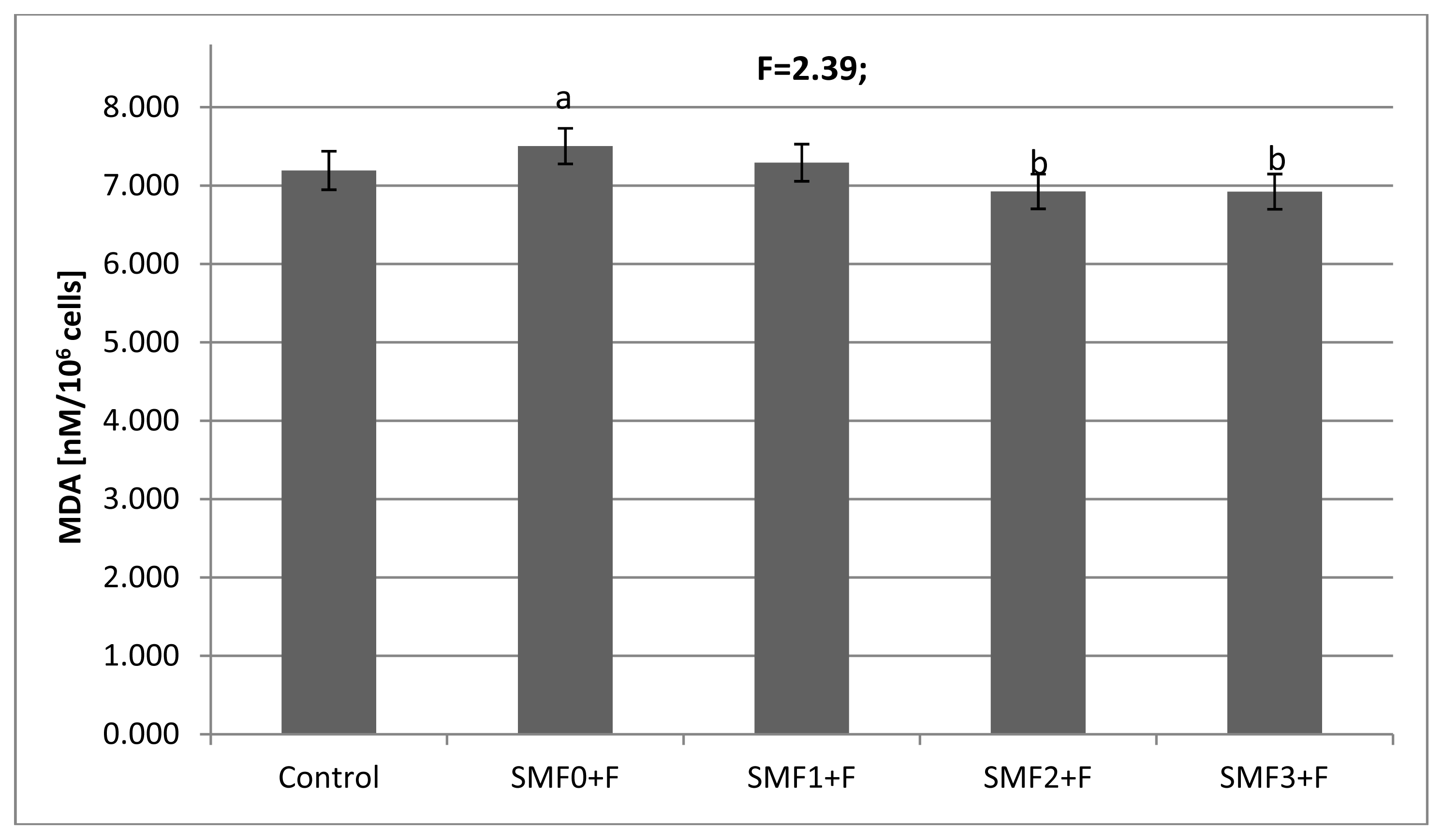
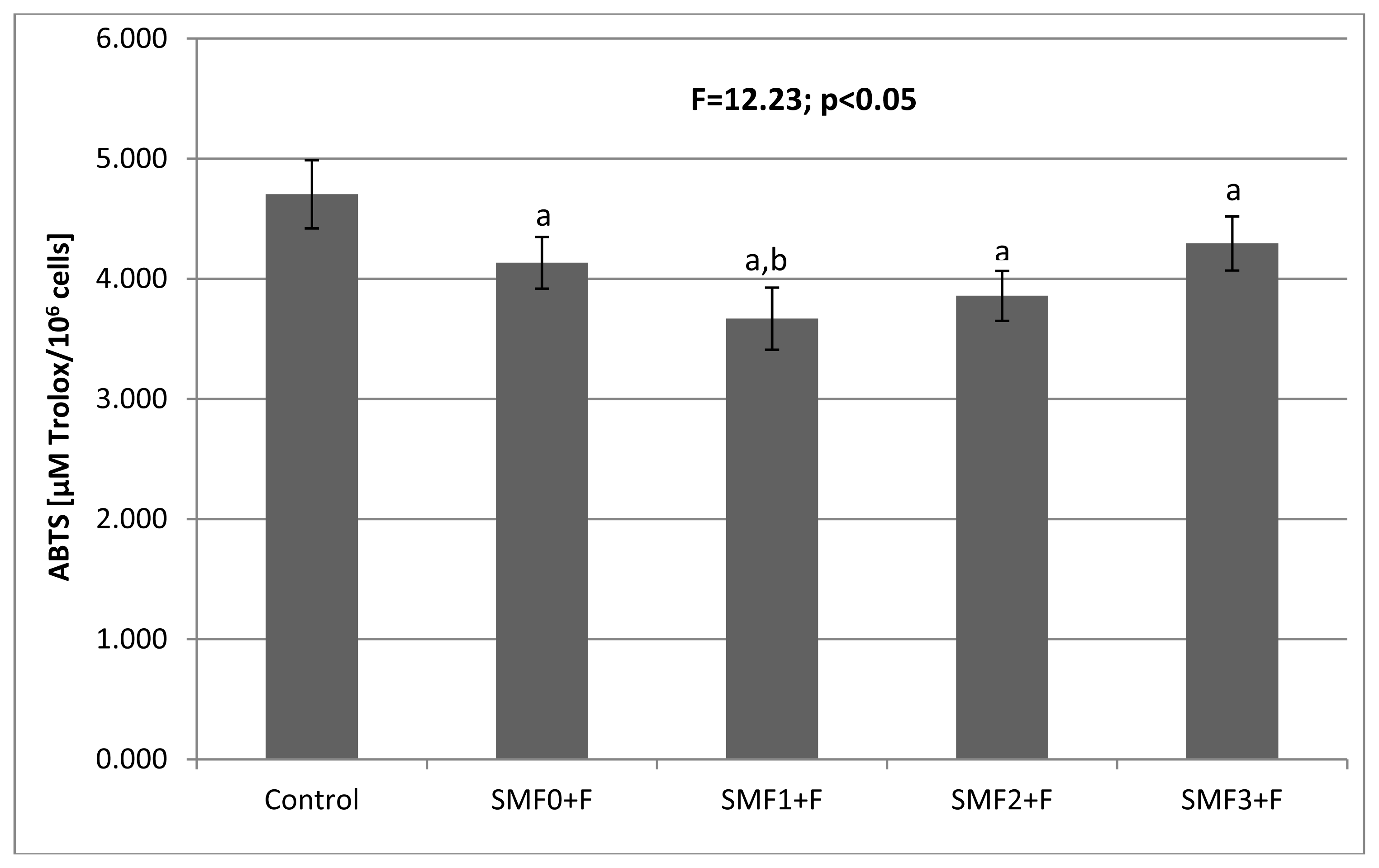
2.2. Effects of Static Magnetic Fields and Fluoride on Mice Fibroblast Cultures
3. Experimental Section
3.1. Chemicals
3.2. Magnetic Test Chamber Features
3.3. Fibroblast Isolation
3.4. Cell Culture Conditions
3.5. Cell Viability Assay
3.6. Lactate Dehydrogenase (LDH) Release Assay
3.7. Preparation of Cell Homogenate
3.8. Biochemical Analysis
3.8.1. Superoxide Dismutase (SOD) Activity Assay
3.8.2. Glutathione Peroxidase (GPx) Activity Assay
3.8.3. Catalase (CAT) Activity Assay
3.8.4. Transferase (GST) Activity Assay
3.8.5. Determination of the Total Antioxidant Status (ABTS)
3.8.6. Lipid Peroxidation Assay
3.8.7. Determination of Adenosine Triphosphate (ATP) Concentration
3.9. Statistical Analysis
4. Conclusions
Acknowledgements
Conflicts of Interest
References
- Todorović, D.; Mirčić, D.; Ilijin, L.; Mrdaković, M.; Vlahović, M.; Prolić, Z.; Mataruga, V.P. Effect of magnetic fields on antioxidative defense and fitness-related traits of Baculum extradentatum (Insecta, Phasmatodea). Bioelectromagnetics 2012, 33, 265–273. [Google Scholar]
- Lee, B.C.; Johng, H.M.; Lim, J.K.; Jeong, J.H.; Baik, K.Y.; Nam, T.J.; Lee, J.H.; Kim, J.; Sohn, U.D.; Yoon, G.; et al. Effect of extremely low frequency magnetic field on the antioxidant defense system in mouse brain: a chemiluminescence study. J. Photochem. Photobiol. B 2004, 73, 43–48. [Google Scholar]
- Sahebjamei, H.; Abdolmaleki, P.; Ghanati, F. Effects of magnetic field on the antioxidant enzyme activities of suspension-cultured tobacco cells. Bioelectromagnetics 2007, 28, 42–47. [Google Scholar]
- Miyakoshi, J. Effects of a static magnetic field at the cellular level. Prog. Biophys. Mol. Biol 2005, 87, 213–223. [Google Scholar]
- Glinka, M.; Gawron, S.A.; Sieroń, A.; Pawłowska-Góral, K.; Cieślar, G.; Sieroń-Stoltny, K. Action of the static magnetic fields on the antioxidant activity in the fibroblasts’ culture. Prz. Elektrotech 2013, 1, 41–44. [Google Scholar]
- Chionna, A.; Dwikat, M.; Panzarini, E.; Tenuzzo, B.; Carlà, E.C.; Verri, T.; Pagliara, P.; Abbro, L.; Dini, L. Cell shape and plasma membrane alterations after static magnetic fields exposure. Eur. J. Histochem 2003, 47, 299–308. [Google Scholar]
- Chionna, A.; Tenuzzo, B.; Panzarini, E.; Dwikat, M.; Abbro, L.; Dini, L. Time-dependent modifications of Hep G2 cells during exposure to Static Magnetic Fields. Bioelectrmagnetics 2005, 26, 275–286. [Google Scholar]
- Štolfa, S.; Škorvánek, M.; Štolfa, P.; Rosocha, J.; Vaško, G.; Sabo, J. Effects of static magnetic field and pulsed electromagnetic field on viability of human chondrocytes in vitro. Physiol. Res. 2007, 56, 45–49. [Google Scholar]
- Gartzke, J.; Lange, K. Celluar target of weak magnetic fields: Ionic conduction along actin filaments of microvilli. Am. J. Physiol. 2002, 283, 1333–1346. [Google Scholar]
- Teodori, L.; Gohde, W.; Valente, M.G.; Tagliaferri, F.; Coletti, D.; Perniconi, B.; Bergamaschi, A.; Cerella, C.; Ghibelli, L. Static magnetic fields affect calcium fluxes and inhibit stress-induced apoptosis in human glioblastoma cells. Cytometry 2002, 49, 143–149. [Google Scholar]
- Teodori, L.; Grabarek, J.; Smolewski, P.; Ghibelli, L.; Bergamaschi, A.; de Nicola, M.; Darzynkiewicz, Z. Exposure of cells to static magnetic fields accelerates loss of integrity of plasma membrane during apoptosis. Cytometry 2002, 49, 113–118. [Google Scholar]
- Fawell, J.; Bailey, K.; Chilton, J.; Dahi, E.; Fewtrell, L.; Magara, Y. Fluoride in Drinking-Water; World Health Organization (WHO) Press: Geneva, Switzerland, 2006. [Google Scholar]
- Narayanaswamy, M.; Piler, M.B. Effect of maternal exposure of fluoride on biometals and oxidative stress parameters in developing CNS of rat. Biol. Trace Elem. Res 2010, 133, 71–82. [Google Scholar]
- Cicek, E.; Aydin, G.; Akdogan, M.; Okutan, H. Effects of chronic ingestion of sodium fluoride on myocardium in a second generation of rats. Hum. Exp. Toxicol 2005, 24, 79–87. [Google Scholar]
- Pawłowska-Góral, K.; Pilawa, B. Detection of free radicals formed by in vitro metabolism of fluoride using EPR spectroscopy. Toxicol. In Vitro 2011, 25, 1269–1273. [Google Scholar]
- Dini, L.; Abbro, L. Bioeffects of moderate-intensity static magnetic fields on cell cultures. Micron 2005, 36, 195–217. [Google Scholar]
- Agalakova, N.I.; Gusev, G.P. Fluoride induces oxidative stress and ATP depletion in the rat erythrocytes in vitro. Environ. Toxicol. Pharmacol 2012, 34, 334–337. [Google Scholar]
- Altıntaş, L.; Eşsiz, D.; Eraslan, G.; İnce, S.; Arslanbaş, E. Prophylactic effect of N-acetylcysteine against sodium fluoride-induced blood oxidative stress in mice. Food Chem. Toxicol 2010, 48, 2838–2841. [Google Scholar]
- Nabavi, S.M.; Nabavi, S.F.; Eslami, S.; Moghaddam, A.H. In vivo protective effects of quercetin against sodium fluoride-induced oxidative stress in the hepatic tissue. Food Chem 2012, 132, 931–935. [Google Scholar]
- Lévêque, V.J.; Vance, C.K.; Nick, H.S.; Silverman, D.N. Redox properties of human manganese superoxide dismutase and active-site mutants. Biochemistry 2001, 40, 10586–10591. [Google Scholar]
- Kula, B.; Dróżdż, M. A study on magnetic field effects on fibroblast cultures. Part 2. The evaluation of the effects of static and extremely low frequency ELF magnetic fields on free-radical processes in fibroblast cultures. Bioelectrochem. Bioenerg 1996, 39, 27–30. [Google Scholar]
- Ghodbane, S.; Amara, S.; Garrel, C.; Arnaud, J.; Ducros, V.; Favier, A.; Sakly, M.; Abdelmelek, H. Selenium supplementation ameliorates static magnetic field-induced disorders in antioxidant status in rat tissues. Environ. Toxicol. Pharmacol 2011, 31, 100–106. [Google Scholar]
- Amara, S.; Abdelmelek, H.; Garrel, C.; Guiraud, P.; Douki, T.; Ravanat, J-L.; Favier, A.; Sakly, M.; Rhouma, B. Zinc supplementation ameliorates static magnetic field-induced oxidative stress in rat tissues. Environ. Toxicol. Pharmacol. 2007, 23, 193–197. [Google Scholar]
- Filipič, J.; Kraigher, B.; Tepuš, B.; Kokol, V.; Mandic-Mulec, I. Effects of low-density static magnetic fields on the growth and activities of wastewater bacteria Escherichiacoli and Pseudomonas putida. Bioresour. Technol 2012, 120, 225–232. [Google Scholar]
- Kabuto, H.; Yokoi, I.; Ogawa, N.; Mori, A.; Liburdy, R.P. Effects of magnetic fields on the accumulation of thiobarbituric acid reactive substances induced by iron salt and H2O2 in mouse brain homogenates or phosphotidylcholine. Pathophysiology 2001, 7, 283–288. [Google Scholar]
- Ishisaka, R.; Kanno, T.; Inai, Y.; Nakahara, H.; Akiyama, J.; Yoshioka, T.; Utsumi, K. Effects of a magnetic fields on the various functions of subcellular organelles and cells. Pathophysiology 2000, 7, 149–152. [Google Scholar]
- Jajte, J.; Grzegorczyk, J.; Zmyślony, M.; Rajkowska, E. Effect of 7 mT static magnetic field and iron ions on rat lymphocytes: apoptosis, necrosis and free radical processes. Bioelectrochemistry 2002, 57, 107–111. [Google Scholar]
- Philips, D.J. Dye Exclusion Test for Cell Viability. In Tissue, Culture, Methods and Application; Academic Press: New York, NY, USA, 1978; pp. 406–408. [Google Scholar]
- Legrand, C.; Bour, J.M.; Jacob, C.; Capiaumont, J.; Martial, A.; Marc, A.; Wudtke, M.; Kretzmer, G.; Demangel, C.; Duval, D. Lactate dehydrogenase (LDH) activity of the cultured eukaryotic cells as marker of the number of dead cells in the medium. J. Biotechnol 1992, 25, 231–243. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem 1971, 44, 276–287. [Google Scholar]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab Clin. Med 1967, 70, 158–169. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| SOD [U/106 cells] | GPx [U/106 cells] | CAT [nM/min/106 cells] | GST [nM/min/106 cells] | |
|---|---|---|---|---|
| Control | 0.78 ± 0.024 | 627.6 ± 2.07 | 10.36 ± 0.86 | 68.73 ± 3.07 |
| SMF0 + F | 0.56 ± 0.018 a | 503.6 ± 24.05 a | 8.66 ± 0.89 | 70.15 ± 2.67 |
| SMF1 + F | 0.56 ± 0.024 a | 541.7 ± 2.77 a,b | 8.72 ± 0.87 | 74.78 ± 1.06 a,b |
| SMF2 + F | 0.58 ± 0.013 a | 615.4 ± 26.12 b | 10.06 ± 0.59 | 76.54 ± 1.69 a,b |
| SMF3 + F | 0.63 ± 0.010 a,b | 695.2 ± 4.28 a,b | 10.69 ± 0.85 b | 83.16 ± 1.12 a,b |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kurzeja, E.; Synowiec-Wojtarowicz, A.; Stec, M.; Glinka, M.; Gawron, S.; Pawłowska-Góral, K. Effect of a Static Magnetic Fields and Fluoride Ions on the Antioxidant Defense System of Mice Fibroblasts. Int. J. Mol. Sci. 2013, 14, 15017-15028. https://doi.org/10.3390/ijms140715017
Kurzeja E, Synowiec-Wojtarowicz A, Stec M, Glinka M, Gawron S, Pawłowska-Góral K. Effect of a Static Magnetic Fields and Fluoride Ions on the Antioxidant Defense System of Mice Fibroblasts. International Journal of Molecular Sciences. 2013; 14(7):15017-15028. https://doi.org/10.3390/ijms140715017
Chicago/Turabian StyleKurzeja, Ewa, Agnieszka Synowiec-Wojtarowicz, Małgorzata Stec, Marek Glinka, Stanisław Gawron, and Katarzyna Pawłowska-Góral. 2013. "Effect of a Static Magnetic Fields and Fluoride Ions on the Antioxidant Defense System of Mice Fibroblasts" International Journal of Molecular Sciences 14, no. 7: 15017-15028. https://doi.org/10.3390/ijms140715017