Could Radiotherapy Effectiveness Be Enhanced by Electromagnetic Field Treatment?

 ,
,
Abstract
:1. Introduction to Electromagnetic Fields: What They Are and How They Act
2. Biological Effects of EMFs
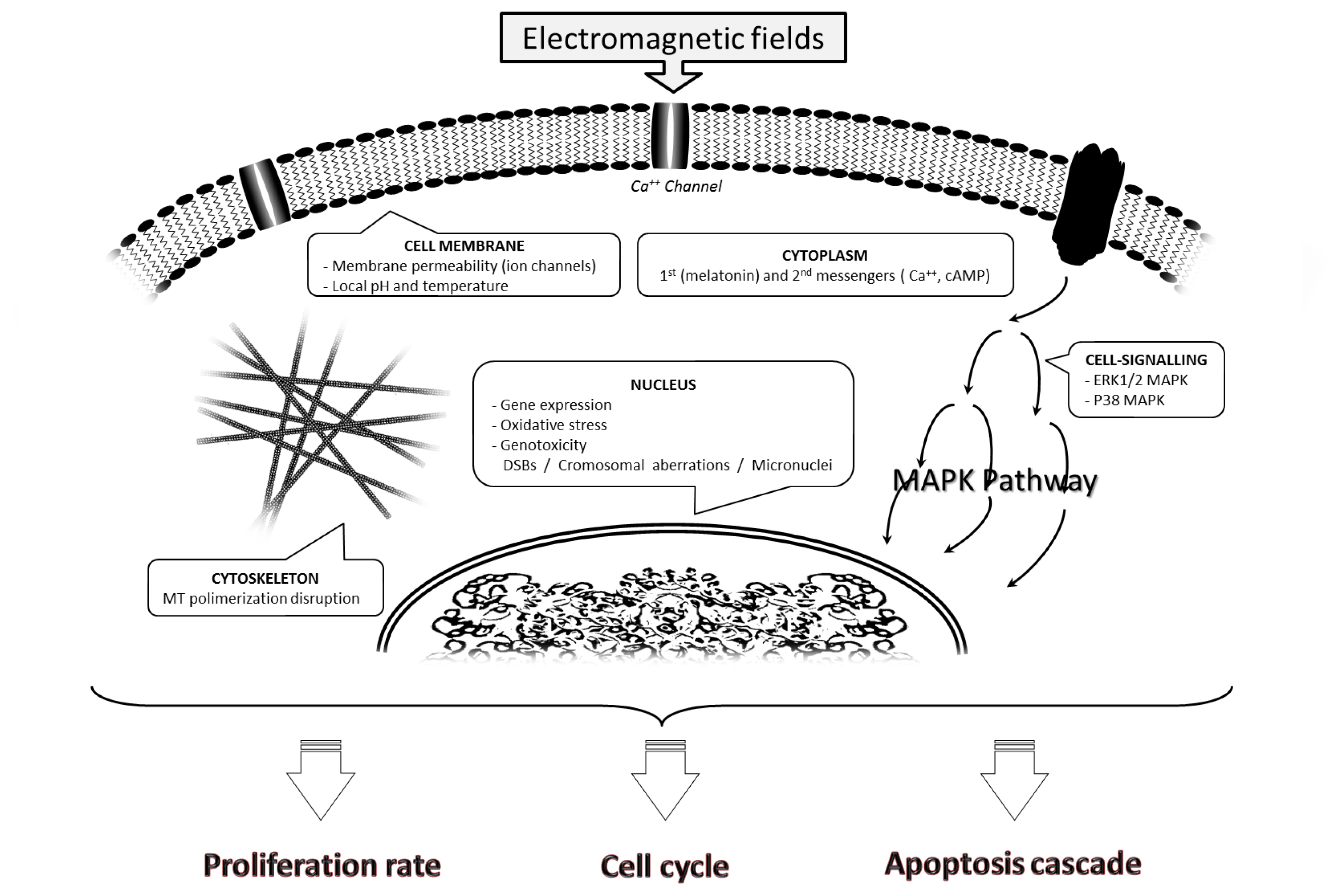
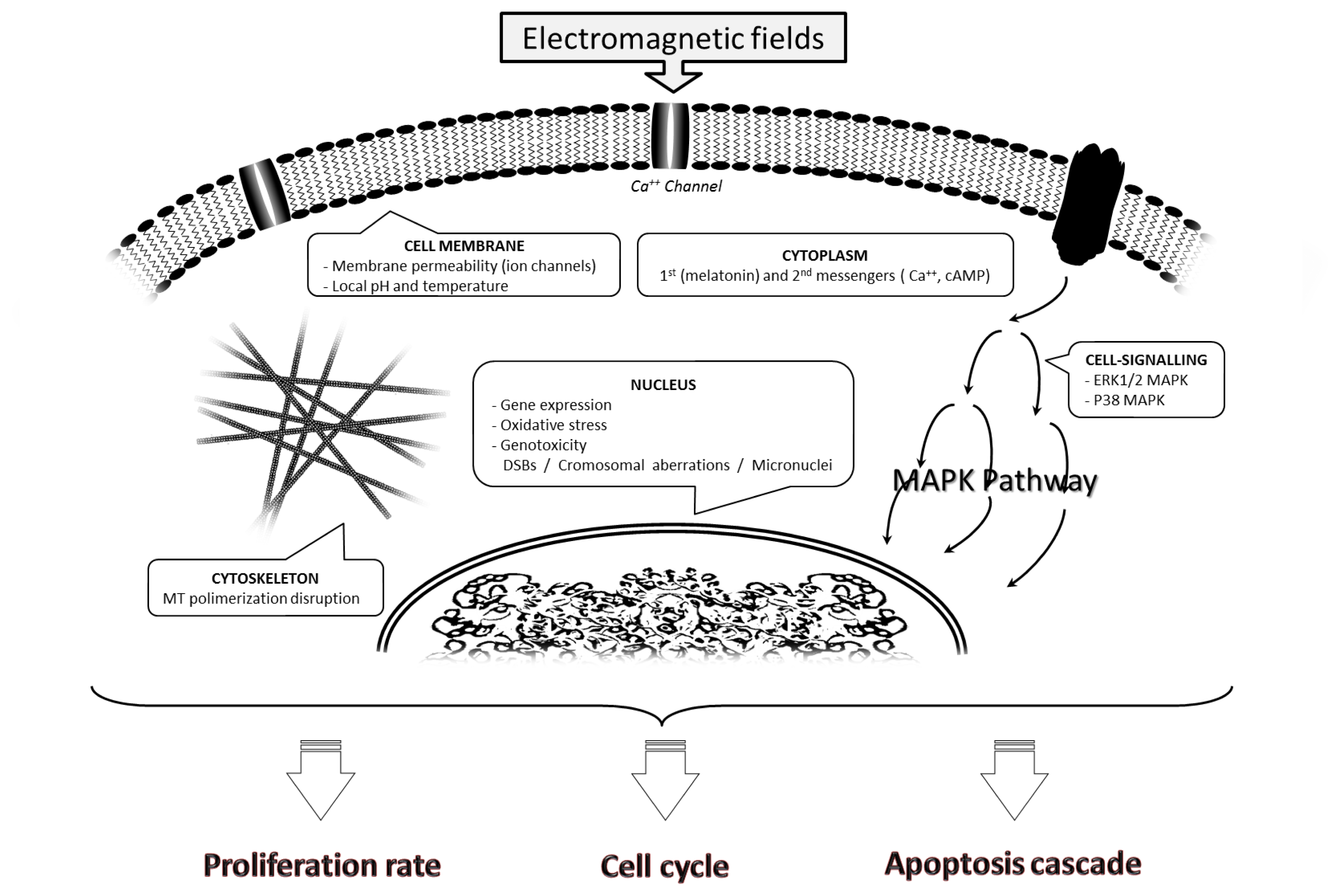
2.1. Effects on the Cell Membrane
2.2. Effects on the Cell Cytoskeleton
2.3. Effects on Cytoplasm
2.3.1. Disruption of First and Second Messengers
2.3.2. Alterations in the MAPK Pathway
2.4. Nuclear Effects
2.4.1. Gene Expression
2.4.2. DNA Damage
2.4.2.1. Oxidative Stress
2.4.2.2. Double Strand Breaks, Chromosomal Aberrations and Micronucleus Induction
2.5. Modulation of Proliferation Rate and Cell Cycle Progression
2.6. Induced Apoptosis Cascade
3. EMFs and Magnetic Fields as Useful Adjuvants during Radiotherapy
3.1. Genotoxic Effect of Ionizing Radiation
3.2. In Vitro Effects of EMF and Ionizing Radiation
3.3. In Vivo Effects of EMF and Ionizing Radiation
4. Conclusions and Future Directions
Acknowledgments
Conflict of Interest
References
- Malmivuo, J.; Plonsey, R. Bioelectromagnetism: Principles and Applications of Bioelectric and Biomagnetic Fields; Oxford University Press: New York, NY, USA, 1995. [Google Scholar]
- IARC Working Group on the Evalutation of Carcinogenic Risks to Humans. Non-ionizing radiation, part 1: Static and extremely low-frequency (ELF) electric and magnetic fields. IARC Monogr. Eval. Carcinog. Risks to Hum. 2002, 80, 1–395.
- Schreier, N.; Huss, A.; Röösli, M. The prevalence of symptoms attributed to electromagnetic field exposure: A cross-sectional representative survey in Switzerland. Soc. Prev. Med 2006, 51, 202–209. [Google Scholar]
- Rubin, G.J.; Nieto-Hernandez, R.; Wessely, S. Idiopathic environmental intolerance attributed to electromagnetic fields (formerly “electromagnetic hypersensitivity”): An updated systematic review of provocation studies. Bioelectromagnetics 2010, 31, 1–11. [Google Scholar]
- Fröhlich, H. Bose condensation of strongly excited longitudinal electric modes. Phys. Lett. A 1968, 26, 402–403. [Google Scholar]
- Fröhlich, H. Quantum mechanical concepts in biology. Theoret. Phys. Biol 1969, 1, 13–22. [Google Scholar]
- Fröhlich, H. Long-range coherence and energy storage in biological systems. Int. J. Quantum Chem 1968, 2, 641–649. [Google Scholar]
- Fröhlich, H. Collective behaviour of non-linearly coupled oscillating fields (with applications to biological systems). J. Collect. Phenom 1973, 1, 101. [Google Scholar]
- Frohlich, H. Coherent electric vibrations in biological systems and the cancer problem. IEEE Trans. Microw. Theory Tech 1978, 26, 613–618. [Google Scholar]
- Funk, R.H.; Monsees, T.; Özkucur, N. Electromagnetic effects—From cell biology to medicine. Prog. Histochem. Cytochem 2009, 43, 177–264. [Google Scholar]
- Galvanovskis, J.; Sandblom, J. Amplification of electromagnetic signals by ion channels. Biophys. J 1997, 73, 3056–3065. [Google Scholar]
- Cagni, E.; Remondini, D.; Mesirca, P.; Castellani, G.C.; Verondini, E.; Bersani, F. Effects of exogenous electromagnetic fields on a simplified ion channel model. J. Biol. Phys 2007, 33, 183–194. [Google Scholar]
- Pokorny, J. Physical aspects of biological activity and cancer. AIP Adv. 2012, 2. [Google Scholar] [CrossRef]
- Kirson, E.D.; Gurvich, Z.; Schneiderman, R.; Dekel, E.; Itzhaki, A.; Wasserman, Y.; Schatzberger, R.; Palti, Y. Disruption of cancer cell replication by alternating electric fields. Cancer Res 2004, 64, 3288–3295. [Google Scholar]
- Touitou, Y.; Bogdan, A.; Lambrozo, J.; Selmaoui, B. Is melatonin the hormonal missing link between magnetic field effects and human diseases? Cancer Causes Control 2006, 17, 547–552. [Google Scholar]
- Strauch, B.; Herman, C.; Dabb, R.; Ignarro, L.J.; Pilla, A.A. Evidence-based use of pulsed electromagnetic field therapy in clinical plastic surgery. Aesthet. Surg. J 2009, 29, 135–143. [Google Scholar]
- Manikonda, P.K.; Rajendra, P.; Devendranath, D.; Gunasekaran, B.; Channakeshava Aradhya, R.S.S.; Sashidhar, R.B.; Subramanyam, C. Influence of extremely low frequency magnetic fields on Ca2+ signaling and nmda receptor functions in rat hippocampus. Neurosci. Lett. 2007, 413, 145–149. [Google Scholar]
- Jin, M.; Blank, M.; Goodman, R. Erk1/2 phosphorylation, induced by electromagnetic fields, diminishes during neoplastic transformation. J. Cell. Biochem 2000, 78, 371–379. [Google Scholar]
- Buttiglione, M.; Roca, L.; Montemurno, E.; Vitiello, F.; Capozzi, V.; Cibelli, G. Radiofrequency radiation (900 MHz) induces Egr-1 gene expression and affects cell-cycle control in human neuroblastoma cells. J. Cell. Physiol 2007, 213, 759–767. [Google Scholar]
- Kim, J.; Ha, C.S.; Lee, H.J.; Song, K. Repetitive exposure to a 60-Hz time-varying magnetic field induces DNA double-strand breaks and apoptosis in human cells. Biochem. Biophys. Res. Commun 2010, 400, 739–744. [Google Scholar]
- Busca, R.; Abbe, P.; Mantoux, F.; Aberdam, E.; Peyssonnaux, C.; Eychene, A.; Ortonne, J.-P.; Ballotti, R. Ras mediates the cAMP-dependent activation of extracellular signal-regulated kinases (ERKs) in melanocytes. EMBO J 2000, 19, 2900–2910. [Google Scholar]
- Yamaguchi, T.; Wallace, D.P.; Magenheimer, B.S.; Hempson, S.J.; Grantham, J.J.; Calvet, J.P. Calcium restriction allows camp activation of the b-raf/erk pathway, switching cells to a cAMP-dependent growth-stimulated phenotype. J. Biol. Chem 2004, 279, 40419–40430. [Google Scholar]
- Ruiz-Gómez, M.J.; Martínez-Morillo, M. Electromagnetic fields and the induction of DNA strand breaks. Electromagn. Biol. Med 2009, 28, 201–214. [Google Scholar]
- Anastassopoulou, J.; Theophanides, T. Magnesium-DNA interactions and the possible relation of magnesium to carcinogenesis. Irradiation and free radicals. Crit. Rev. Oncol. Hematol 2002, 42, 79–91. [Google Scholar]
- Lee, H.J.; Jin, Y.B.; Lee, J.S.; Choi, J.I.; Lee, J.W.; Myung, S.H.; Lee, Y.S. Combined effects of 60 Hz electromagnetic field exposure with various stress factors on cellular transformation in NIH3T3 cells. Bioelectromagnetics 2011, 33, 207–214. [Google Scholar]
- Jin, Y.B.; Kang, G.-Y.; Lee, J.S.; Choi, J.-I.; Lee, J.-W.; Hong, S.-C.; Myung, S.H.; Lee, Y.-S. Effects on micronuclei formation of 60-Hz electromagnetic field exposure with ionizing radiation, hydrogen peroxide, or c-Myc overexpression. Int. J. Radiat. Biol 2012, 88, 374–380. [Google Scholar]
- Goraca, A.; Ciejka, E.; Piechota, A. Effects of extremely low frequency magnetic field on the parameters of oxidative stress in heart. J. Physiol. Pharmacol 2010, 61, 333–338. [Google Scholar]
- Falone, S.; Grossi, M.R.; Cinque, B.; D’Angelo, B.; Tettamanti, E.; Cimini, A.; di Ilio, C.; Amicarelli, F. Fifty hertz extremely low-frequency electromagnetic field causes changes in redox and differentiative status in neuroblastoma cells. Int. J. Biochem. Cell Biol 2007, 39, 2093–2106. [Google Scholar]
- Focke, F.; Schuermann, D.; Kuster, N.; Schär, P. DNA fragmentation in human fibroblasts under extremely low frequency electromagnetic field exposure. Mutat. Res. Fundam. Mol. Mech. Mutagenes 2010, 683, 74–83. [Google Scholar]
- Hynes, J.; Floyd, S.; Soini, A.E.; O’Connor, R.; Papkovsky, D.B. Fluorescence-based cell viability screening assays using water-soluble oxygen probes. J. Biomol. Screen 2003, 8, 264–272. [Google Scholar]
- Wolf, F.I.; Torsello, A.; Tedesco, B.; Fasanella, S.; Boninsegna, A.; D’Ascenzo, M.; Grassi, C.; Azzena, G.B.; Cittadini, A. 50-Hz extremely low frequency electromagnetic fields enhance cell proliferation and DNA damage: Possible involvement of a redox mechanism. BBA Mol. Cell Res 2005, 1743, 120–129. [Google Scholar]
- Simkó, M.; Droste, S.; Kriehuber, R.; Weiss, D.G. Stimulation of phagocytosis and free radical production in murine macrophages by 50 Hz electromagnetic fields. Eur. J. Cell Biol 2001, 80, 562–566. [Google Scholar]
- Santoro, N.; Lisi, A.; Pozzi, D.; Pasquali, E.; Serafino, A.; Grimaldi, S. Effect of extremely low frequency (ELF) magnetic field exposure on morphological and biophysical properties of human lymphoid cell line (RAJI). BBA Mol. Cell Res 1997, 1357, 281–290. [Google Scholar]
- Pilla, A.; Fitzsimmons, R.; Muehsam, D.; Wu, J.; Rohde, C.; Casper, D. Electromagnetic fields as first messenger in biological signaling: Application to calmodulin-dependent signaling in tissue repair. BBA Gen. Subj 2011, 1810, 1236–1245. [Google Scholar]
- Löscher, W.; Mevissen, M. Animal studies on the role of 50/60-Hertz magnetic fields in carcinogenesis. Life Sci 1994, 54, 1531–1543. [Google Scholar]
- Touitou, Y.; Lambrozo, J.; Camus, F.; Charbuy, H. Magnetic fields and the melatonin hypothesis: A study of workers chronically exposed to 50-Hz magnetic fields. Am. J. Physiol. Regul. Integr. Comp. Physiol 2003, 284, R1529–R1535. [Google Scholar]
- Luchetti, F.; Canonico, B.; Betti, M.; Arcangeletti, M.; Pilolli, F.; Piroddi, M.; Canesi, L.; Papa, S.; Galli, F. Melatonin signaling and cell protection function. FASEB J 2010, 24, 3603–3624. [Google Scholar]
- Mediavilla, M.D.; Sanchez-Barcelo, E.J.; Tan, D.X.; Manchester, L.; Reiter, R.J. Basic mechanisms involved in the anti-cancer effects of melatonin. Curr. Med. Chem 2010, 17, 4462–4481. [Google Scholar]
- Liburdy, R.P.; Sloma, T.R.; Sokolic, R.; Yaswen, P. ELF magnetic fields, breast cancer, and melatonin: 60 Hz fields block melatonin’s oncostatic action on ER+ breast cancer cell proliferation. J. Pineal Res 1993, 14, 89–97. [Google Scholar]
- Aldinucci, C.; Palmi, M.; Sgaragli, G.; Benocci, A.; Meini, A.; Pessina, F.; Pessina, G.P. The effect of pulsed electromagnetic fields on the physiologic behaviour of a human astrocytoma cell line. BBA Mol. Cell Res 2000, 1499, 101–108. [Google Scholar]
- Sergeev, I.; Rhoten, W. Regulation of intracellular calcium in human breast cancer cells. Endocrine 1998, 9, 321–327. [Google Scholar]
- Follin-Arbelet, V.; Hofgaard, P.; Hauglin, H.; Naderi, S.; Sundan, A.; Blomhoff, R.; Bogen, B.; Blomhoff, H. Cyclic AMP induces apoptosis in multiple myeloma cells and inhibits tumor development in a mouse myeloma model. BMC Cancer 2011, 11. [Google Scholar] [CrossRef]
- Rocha, A.S.; Paternot, S.; Coulonval, K.; Dumont, J.E.; Soares, P.; Roger, P.P. Cyclic AMP inhibits the proliferation of thyroid carcinoma cell lines through regulation of cdk4 phosphorylation. Mol. Biol. Cell 2008, 19, 4814–4825. [Google Scholar]
- Dumaz, N.; Light, Y.; Marais, R. Cyclic AMP blocks cell growth through Raf-1-dependent and Raf-1-independent mechanisms. Mol. Cell. Biol 2002, 22, 3717–3728. [Google Scholar]
- Schimmelpfeng, J.; Stein, J.-C.; Dertinger, H. Action of 50 Hz magnetic fields on cyclic AMP and intercellular communication in monolayers and spheroids of mammalian cells. Bioelectromagnetics 1995, 16, 381–386. [Google Scholar]
- Hogan, M.V.; Wieraszko, A. An increase in cAMP concentration in mouse hippocampal slices exposed to low-frequency and pulsed magnetic fields. Neurosci. Lett 2004, 366, 43–47. [Google Scholar]
- Friedman, J.; Kraus, S.; Hauptman, Y.; Schiff, Y.; Seger, R. Mechanism of short-term ERK activation by electromagnetic fields at mobile phone frequencies. Biochem. J. 2007, 405. [Google Scholar] [CrossRef]
- Dent, P.; Yacoub, A.; Fisher, P.B.; Hagan, M.P.; Grant, S. MAPK pathways in radiation responses. Oncogene 2003, 22, 5885–5896. [Google Scholar]
- Nie, K.; Henderson, A. MAP kinase activation in cells exposed to a 60 Hz electromagnetic field. J. Cell. Biochem 2003, 90, 1197–1206. [Google Scholar]
- Lin, H.; Han, L.; Blank, M.; Head, M.; Goodman, R. Magnetic field activation of protein-DNA binding. J. Cell. Biochem 1998, 70, 297–303. [Google Scholar]
- Li, H.; Zeng, Q.; Weng, Y.; Lu, D.; Jiang, H.; Xu, Z. Effects of elf magnetic fields on protein expression profile of human breast cancer cell MCF7. Sci. China Ser. C 2005, 48, 506–514. [Google Scholar]
- Marnett, L.J. Oxyradicals and DNA damage. Carcinogenesis 2000, 21, 361–370. [Google Scholar]
- Colotta, F.; Allavena, P.; Sica, A.; Garlanda, C.; Mantovani, A. Cancer-related inflammation, the seventh hallmark of cancer: Links to genetic instability. Carcinogenesis 2009, 30, 1073–1081. [Google Scholar]
- Desai, N.; Kesari, K.; Agarwal, A. Pathophysiology of cell phone radiation: Oxidative stress and carcinogenesis with focus on male reproductive system. Reprod. Biol. Endocrinol. 2009, 7. [Google Scholar] [CrossRef]
- Zmyślony, M.; Politanski, P.; Rajkowska, E.; Szymczak, W.; Jajte, J. Acute exposure to 930 MHz CW electromagnetic radiation in vitro affects reactive oxygen species level in rat lymphocytes treated by iron ions. Bioelectromagnetics 2004, 25, 324–328. [Google Scholar]
- Irmak, M.K.; Fadıllıoğlu, E.; Güleç, M.; Erdoğan, H.; Yağmurca, M.; Akyol, Ö. Effects of electromagnetic radiation from a cellular telephone on the oxidant and antioxidant levels in rabbits. Cell Biochem. Funct. 2002, 20, 279–283. [Google Scholar]
- Patruno, A.; Pesce, M.; Marrone, A.; Speranza, L.; Grilli, A.; de Lutiis, M.A.; Felaco, M.; Reale, M. Activity of matrix metallo proteinases (MMPs) and the tissue inhibitor of MMP (TIMP)-1 in electromagnetic field-exposed THP-1 cells. J. Cell. Physiol 2012, 227, 2767–2774. [Google Scholar]
- Hintzsche, H.; Jastrow, C.; Kleine-Ostmann, T.; Kärst, U.; Schrader, T.; Stopper, H. Terahertz electromagnetic fields (0.106 THz) do not induce manifest genomic damage in vitro. PloS One 2012, 7. [Google Scholar] [CrossRef]
- Winker, R.; Ivancsits, S.; Pilger, A.; Adlkofer, F.; Rüdiger, H. Chromosomal damage in human diploid fibroblasts by intermittent exposure to extremely low-frequency electromagnetic fields. Mutat. Res. Genet. Toxicol. Environ. Mutagenes 2005, 585, 43–49. [Google Scholar]
- Ivancsits, S.; Diem, E.; Pilger, A.; Rudiger, H.W.; Jahn, O. Induction of DNA strand breaks by intermittent exposure to extremely-low-frequency electromagnetic fields in human diploid fibroblasts. Mutat. Res. Genet. Toxicol. Environ. Mutagenes 2002, 519, 1–13. [Google Scholar]
- Ruiz-Gómez, M.J.; Sendra-Portero, F.; Martínez-Morillo, M. Effect of 2.45 mT sinusoidal 50 Hz magnetic field on Saccharomyces cerevisiae strains deficient in DNA strand breaks repair. Int. J. Radiat. Biol 2010, 86, 602–611. [Google Scholar]
- Tofani, S.; Barone, D.; Cintorino, M.; de Santi, M.M.; Ferrara, A.; Orlassino, R.; Ossola, P.; Peroglio, F.; Rolfo, K.; Ronchetto, F. Static and ELF magnetic fields induce tumor growth inhibition and apoptosis. Bioelectromagnetics 2001, 22, 419–428. [Google Scholar]
- Tatarov, I.; Panda, A.; Petkov, D.; Kolappaswamy, K.; Thompson, K.; Kavirayani, A.; Lipsky, M.M.; Elson, E.; Davis, C.C.; Martin, S.S. Effect of magnetic fields on tumor growth and viability. Comp. Med 2011, 61, 339–345. [Google Scholar]
- Yoshizawa, H.; Tsuchiya, T.; Mizoe, H.; Ozeki, H.; Kanao, S.; Yomori, H.; Sakane, C.; Hasebe, S.; Motomura, T.; Yamakawa, T. No effect of extremely low-frequency magnetic field observed on cell growth or initial response of cell proliferation in human cancer cell lines. Bioelectromagnetics 2002, 23, 355–368. [Google Scholar]
- Ruiz-Gómez, M.J.; Prieto-Barcia, M.I.; Ristori-Bogajo, E.; Martínez-Morillo, M. Static and 50 Hz magnetic fields of 0.35 and 2.45 mT have no effect on the growth of saccharomyces cerevisiae. Bioelectrochemistry 2004, 64, 151–155. [Google Scholar]
- Alexandre, B.; Frederico, C.; Brad, B.; Reginald, M.; Fin, B.; Niels, K.; Boris, P. Amplitude-modulated electromagnetic fields for the treatment of cancer: Discovery of tumor-specific frequencies and assessment of a novel therapeutic approach. J. Exp. Clin. Cancer Res. 2009, 28. [Google Scholar] [CrossRef]
- Costa, F.; de Oliveira, A.; Meirelles, R.; Machado, M.; Zanesco, T.; Surjan, R.; Chammas, M.; de Souza Rocha, M.; Morgan, D.; Cantor, A. Treatment of advanced hepatocellular carcinoma with very low levels of amplitude-modulated electromagnetic fields. Br. J. Cancer 2011, 105, 640–648. [Google Scholar]
- Zimmerman, J.; Pennison, M.; Brezovich, I.; Yi, N.; Yang, C.; Ramaker, R.; Absher, D.; Myers, R.; Kuster, N.; Costa, F. Cancer cell proliferation is inhibited by specific modulation frequencies. Br. J. Cancer 2011, 106, 307–313. [Google Scholar]
- Kirson, E.D.; Dbalý, V.; Tovaryš, F.; Vymazal, J.; Soustiel, J.F.; Itzhaki, A.; Mordechovich, D.; Steinberg-Shapira, S.; Gurvich, Z.; Schneiderman, R. Alternating electric fields arrest cell proliferation in animal tumor models and human brain tumors. Proc. Natl. Acad. Sci. USA 2007, 104, 10152–10157. [Google Scholar]
- Lange, S.; Richard, D.; Viergutz, T.; Kriehuber, R.; Weiss, D.G.; Simkó, M. Alterations in the cell cycle and in the protein level of cyclin D1, p21CIP1, and p16INK4a after exposure to 50 Hz MF in human cells. Radiat. Environ. Biophys 2002, 41, 131–137. [Google Scholar]
- Ruiz Gómez, M.J.; de la Peña, L.; Pastor, J.M.; Martínez Morillo, M.; Gil, L. 25 Hz electromagnetic field exposure has no effect on cell cycle distribution and apoptosis in U-937 and HCA-2/1cch cells. Bioelectrochemistry 2001, 53, 137–140. [Google Scholar]
- Gorczynska, E.; Wegrzynowicz, R. Structural and functional changes in organelles of liver cells in rats exposed to magnetic fields. Environ. Res 1991, 55, 188–198. [Google Scholar]
- Reubold, T.F.; Eschenburg, S. A molecular view on signal transduction by the apoptosome. Cell. Signal 2012, 24, 1420–1425. [Google Scholar]
- Caraglia, M.; Marra, M.; Mancinelli, F.; D’ambrosio, G.; Massa, R.; Giordano, A.; Budillon, A.; Abbruzzese, A.; Bismuto, E. Electromagnetic fields at mobile phone frequency induce apoptosis and inactivation of the multi-chaperone complex in human epidermoid cancer cells. J. Cell. Physiol 2005, 204, 539–548. [Google Scholar]
- O’Mullane, M.J.; Baker, M.S. Elevated plasminogen receptor expression occurs as a degradative phase event in cellular apoptosis. Immunol. Cell Biol 1999, 77, 249–255. [Google Scholar]
- Girgert, R.; Emons, G.; Hanf, V.; Gründker, C. Exposure of MCF-7 breast cancer cells to electromagnetic fields up-regulates the plasminogen activator system. Int. J. Gynecol. Cancer 2009, 19, 334–338. [Google Scholar]
- Kovacic, P.; Somanathan, R. Electromagnetic fields: Mechanism, cell signaling, other bioprocesses, toxicity, radicals, antioxidants and beneficial effects. J. Recept. Signal Transduct 2010, 30, 214–226. [Google Scholar]
- Cameron, I.L.; Sun, L.Z.; Short, N.; Hardman, W.E.; Williams, C.D. Therapeutic electromagnetic field (TEMF) and gamma irradiation on human breast cancer xenograft growth, angiogenesis and metastasis. Cancer Cell Int. 2005, 5. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.; Jiang, S.; Chen, B. The effect of 100 Hz magnetic field combined with X-ray on hepatoma-implanted mice. Bioelectromagnetics 2011, 32, 322–324. [Google Scholar]
- Jeggo, P.A. Risks from low dose/dose rate radiation: What an understanding of DNA damage response mechanisms can tell us. Health Phys 2009, 97, 416–425. [Google Scholar]
- Kempner, E.S. Molecular size determination of enzymes by radiation inactivation. Adv. Enzymol. Relat. Areas Mol. Biol. 1988, 107–147. [Google Scholar]
- Miyakoshi, J.; Koji, Y.; Wakasa, T.; Takebe, H. Long-term exposure to a magnetic field (5 mT at 60 Hz) increases X-ray-induced mutations. J. Radiat. Res 1999, 40, 13–21. [Google Scholar]
- Ding, G.-R.; Yaguchi, H.; Yoshida, M.; Miyakoshi, J. Increase in X-ray-induced mutations by exposure to magnetic field (60 hz, 5 mt) in nf-κb-inhibited cells. Biochem. Biophys. Res. Commun 2000, 276, 238–243. [Google Scholar]
- Mairs, R.J.; Hughes, K.; Fitzsimmons, S.; Prise, K.M.; Livingstone, A.; Wilson, L.; Baig, N.; Clark, A.M.; Timpson, A.; Patel, G.; et al. Microsatellite analysis for determination of the mutagenicity of extremely low-frequency electromagnetic fields and ionising radiation in vitro. Mutat. Res. Genet. Toxicol. Environ. Mutagenes 2007, 626, 34–41. [Google Scholar]
- Joiner, M.; van der Kogel, A. Basic Clinical Radiobiology, 4th ed; Hodder Arnold: London, UK, 2009; Volume 1, pp. 41–55. [Google Scholar]
- Ding, G.R.; Nakahara, T.; Miyakoshi, J. Induction of kinetochore-positive and kinetochore-negative micronuclei in CHO cells by ELF magnetic fields and/or X-rays. Mutagenesis 2003, 18, 439–443. [Google Scholar]
- Lagroye, I.; Poncy, J.L. The effect of 50 Hz electromagnetic fields on the formation of micronuclei in rodent cell lines exposed to gamma radiation. Int. J. Radiat. Biol 1997, 72, 249–254. [Google Scholar]
- Manti, L.; D’Arco, A. Cooperative biological effects between ionizing radiation and other physical and chemical agents. Mutat. Res. Rev. Mutat. Res 2010, 704, 115–122. [Google Scholar]
- Jian, W.; Wei, Z.; Zhiqiang, C.; Zheng, F. X-ray-induced apoptosis of BEL-7402 cell line enhanced by extremely low frequency electromagnetic field in vitro. Bioelectromagnetics 2009, 30, 163–165. [Google Scholar]
- Ahmed, M.M.; Sells, S.F.; Venkatasubbarao, K.; Fruitwala, S.M.; Muthukkumar, S.; Harp, C.; Mohiuddin, M.; Rangnekar, V.M. Ionizing radiation-inducible apoptosis in the absence of p53 linked to transcription factor EGR-1. J. Biol. Chem 1997, 272, 33056–33061. [Google Scholar]
- Ahmed, M.M. Regulation of radiation-induced apoptosis by early growth response-1 gene in solid tumors. Curr. Cancer Drug Targets 2004, 4, 43–52. [Google Scholar]
- Ahmed, M.M.; Venkatasubbarao, K.; Fruitwala, S.M.; Muthukkumar, S.; Wood, D.P., Jr; Sells, S.F.; Mohiuddin, M.; Rangnekar, V.M. EGR-1 induction is required for maximal radiosensitivity in A375-C6 melanoma cells. J. Biol. Chem. 1996, 271, 29231–29237. [Google Scholar]
- Wang, Y.; Liu, L.; Zhou, D. Inhibition of p38 MAPK attenuates ionizing radiation-induced hematopoietic cell senescence and residual bone marrow injury. Radiat. Res 2011, 176, 743–752. [Google Scholar]
- De la Cruz-Morcillo, M.; Valero, M.; Callejas-Valera, J.; Arias-Gonzalez, L.; Melgar-Rojas, P.; Galán-Moya, E.; García-Gil, E.; García-Cano, J.; Sánchez-Prieto, R. P38MAPK is a major determinant of the balance between apoptosis and autophagy triggered by 5-fluorouracil: Implication in resistance. Oncogene 2011, 31, 1073–1085. [Google Scholar]
- Dickinson, S.E.; Olson, E.R.; Zhang, J.; Cooper, S.J.; Melton, T.; Criswell, P.J.; Casanova, A.; Dong, Z.; Hu, C.; Saboda, K. P38 MAP kinase plays a functional role in UVB-induced mouse skin carcinogenesis. Mol. Carcinogenes 2011, 50, 469–478. [Google Scholar]
- Wood, C.D.; Thornton, T.M.; Sabio, G.; Davis, R.A.; Rincon, M. Nuclear localization of p38 MAPK in response to DNA damage. Int. J. Biol. Sci 2009, 5, 428–437. [Google Scholar]
- De la Cruz-Morcillo, M.A.; García-Cano, J.; Arias-González, L.; García-Gil, E.; Artacho-Cordón, F.; Ríos-Arrabal, S.; Valero, M.L.; Cimas, F.J.; Serrano-Oviedo, L.; Villas, M.V. Abrogation of the p38 MAPKα signaling pathway does not promote radioresistance but its activity is required for 5-fluorouracil-associated radiosensitivity. Cancer Lett 2013, 335, 66–74. [Google Scholar]
- Wang, X.; McGowan, C.H.; Zhao, M.; He, L.; Downey, J.S.; Fearns, C.; Wang, Y.; Huang, S.; Han, J. Involvement of the MKK6-p38γ cascade in γ-radiation-induced cell cycle arrest. Mol. Cell. Biol 2000, 20, 4543–4552. [Google Scholar]
- Cao, Y.; Zhang, W.; Lu, M.X.; Xu, Q.; Meng, Q.Q.; Nie, J.H.; Tong, J. 900-MHz microwave radiation enhances γ-ray adverse effects on SHG44 cells. J. Toxicol. Environ. Health A 2009, 72, 727–732. [Google Scholar]
- Baan, R.; Grosse, Y.; Lauby-Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Islami, F.; Galichet, L.; Straif, K. Carcinogenicity of radiofrequency electromagnetic fields. Lancet Oncol 2011, 12, 624–626. [Google Scholar]
- Ahlbom, I.; Cardis, E.; Green, A.; Linet, M.; Savitz, D.; Swerdlow, A. Review of the epidemiologic literature on EMF and health. Environ. Health Perspect 2001, 109, 911–933. [Google Scholar]
- Calvente, I.; Fernandez, M.; Villalba, J.; Olea, N.; Nuñez, M. Exposure to electromagnetic fields (non-ionizing radiation) and its relationship with childhood leukemia: A systematic review. Sci. Total Environ 2010, 408, 3062–3069. [Google Scholar]
- Linet, M.S.; Hatch, E.E.; Kleinerman, R.A.; Robison, L.L.; Kaune, W.T.; Friedman, D.R.; Severson, R.K.; Haines, C.M.; Hartsock, C.T.; Niwa, S. Residential exposure to magnetic fields and acute lymphoblastic leukemia in children. N. Engl. J. Med 1997, 337, 1–8. [Google Scholar]

{kind=link}
| Cell compartment | Frequency | Intensity | Time | EMF type | Biological effect | Reference |
|---|---|---|---|---|---|---|
| Cell membrane | 25 pulses/s | Peak: 0.25 T; Average: 0.5 T | 238 μs 1000 pulses/day | AC MF | Changes in the local pH and/or temperature | [10] |
| <100 Hz | <100 μT | 0–8 ns | MF | Formation of ion channels | [11] | |
| MW (≈1 GHz) | >105 V/m | Several periods of oscillation | EF | [12] | ||
| Cell cytoskeleton | 50 Hz | 0.5–1.5 mT | 45 min | MF | No influence of EMF on cytoskeleton and/or motor proteins | [13] |
| 50 Hz | 2 mT (rms) | 72 h | MF | Changes in microtubule polymerization | [14] | |
| 100–300 kHz | 2 V/cm | 24 h | AC EF | [15] | ||
| Cytoplasm | ||||||
| 1st/2nd messengers | 60 Hz | 1.9–11.95 mG | – | MF | Melatonin involved in transmission of EMF into the cell | [16] |
| 50 Hz | Peak: 3 mT | 24 h | EMF | Influence the transport of Ca2+ and, hence, its homeostasis | [17] | |
| 50 Hz | 2 mT | 5 min | MF | Deregulation of the cAMP concentration | [18] | |
| MAPK pathway | 60Hz | 0.8–300 μT | 30 min | EMF | Induction of ERK1/2 phosphorylation | [19] |
| 875 MHz | 0.005, 0.03 and 0.11 mW/cm2 | 30 min | EMF (S) | [20] | ||
| 900 MHz | 1 W/kg | 24 h | EMF (SAR) | [21] | ||
| 875 MHz | 0.10 mW/cm2 | 30 min | EMF (S) | P38 MAPK activation | [20] | |
| 60 Hz | 6 mT | 30 min/day;3 days | AC MF | [22] | ||
| Nucleus | ||||||
| Gene expression | 60 Hz | 0.8–300 μT | 60 min | MF | Increase of AP-1 transcription factor | [19] |
| 60 Hz | 8 μT | 20 min field-on; 20 min field-off | [23] | |||
| 50 Hz | 0.4 mT | 20 min | MF | Increase RNA binding protein levels | [24] | |
| 900 MHz | 1 W/kg | – | EMF (SAR) | Increase in mRNA levels of Egr-1, Bcl-2 and survivin genes | [21] | |
| DNA damage | ||||||
| 1. Oxidative stress | ||||||
| 1.1. Antioxidant system | 50 Hz | 1 mT rms | 24 h | AC MF | ↓ SOD activity | [25] |
| 50 Hz | 1 mT | <96 h | MF | = SOD activity | [26] | |
| 50 Hz | 1 mT rms | 24 h | AC MF | ↓ Catalase activity | [25] | |
| 50 Hz | 1 mT | <96h | MF | Glutathione S-transferase | [26] | |
| 50 Hz | 1 mT | <96h | MF | Glutathione peroxidase | [26] | |
| 50 Hz | 1 mT | <96h | MF | ↑ Reduced/total GSH ratio | [26] | |
| 1.2. Pro-oxidant molecules | 50 Hz | 1 mT rms | 24 h | MF | ↑ iNOS | [25] |
| 930 MHz | 5 W/m2 | 5–15 min | CW EMF (S) | ↑ ROS | [27] | |
| 900 MHz | 2 W peak 0.02 mW/cm2 | 30 min/day; 7 days | EMF (S) | [28] | ||
| 50 Hz | 1 mT rms | 24 h | AC MF | [25] | ||
| 50 Hz | 1 mT | <96 h | MF | = ROS | [26] | |
| 2. Genotoxicity | 60 Hz | 6 mT | 30 min/day; 1–3 days | AC MF | Increased levels of γH2AX | [22] |
| 50 Hz | 1 mT | 15 h, 5 field-on/10 field-off | MF | Double Strand Breaks | [29] | |
| 50 Hz | 1 mT | 24 h | MF | [30] | ||
| 50 Hz | 1 mT | 15 h | EMF | [31] | ||
| 50 Hz | 1 mT | 15 h, 5′ field-on/10′ field-off | MF | Chromosome aberrations | [29] | |
| 50 Hz | 1 mT | 2–24 h, 5′ field-on/10′ field-off. | MF | Micronucleus induction | [29] | |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Francisco, A.-C.; Mar, S.-A.M.d.; Irene, C.; Sandra, R.-A.; Josefa, L.; Elisa, R.-M.; Nicolás, O.; Isabel, N.M. Could Radiotherapy Effectiveness Be Enhanced by Electromagnetic Field Treatment? Int. J. Mol. Sci. 2013, 14, 14974-14995. https://doi.org/10.3390/ijms140714974
Francisco A-C, Mar S-AMd, Irene C, Sandra R-A, Josefa L, Elisa R-M, Nicolás O, Isabel NM. Could Radiotherapy Effectiveness Be Enhanced by Electromagnetic Field Treatment? International Journal of Molecular Sciences. 2013; 14(7):14974-14995. https://doi.org/10.3390/ijms140714974
Chicago/Turabian StyleFrancisco, Artacho-Cordón, Salinas-Asensio María del Mar, Calvente Irene, Ríos-Arrabal Sandra, León Josefa, Román-Marinetto Elisa, Olea Nicolás, and Núñez María Isabel. 2013. "Could Radiotherapy Effectiveness Be Enhanced by Electromagnetic Field Treatment?" International Journal of Molecular Sciences 14, no. 7: 14974-14995. https://doi.org/10.3390/ijms140714974