Thrombospondin-1 Is a Putative Target Gene of Runx2 and Runx3

Abstract
:1. Introduction
2. Results and Discussion
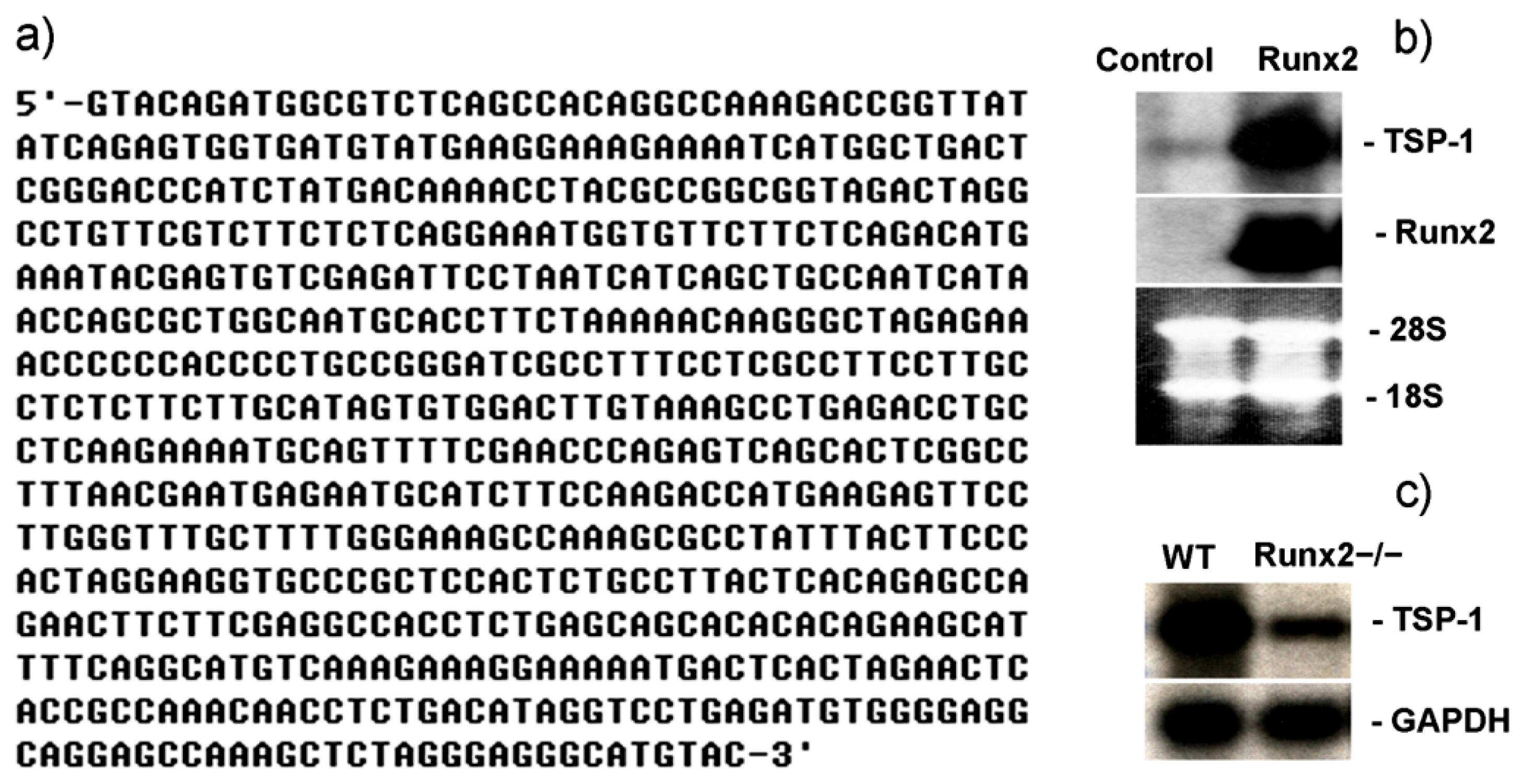
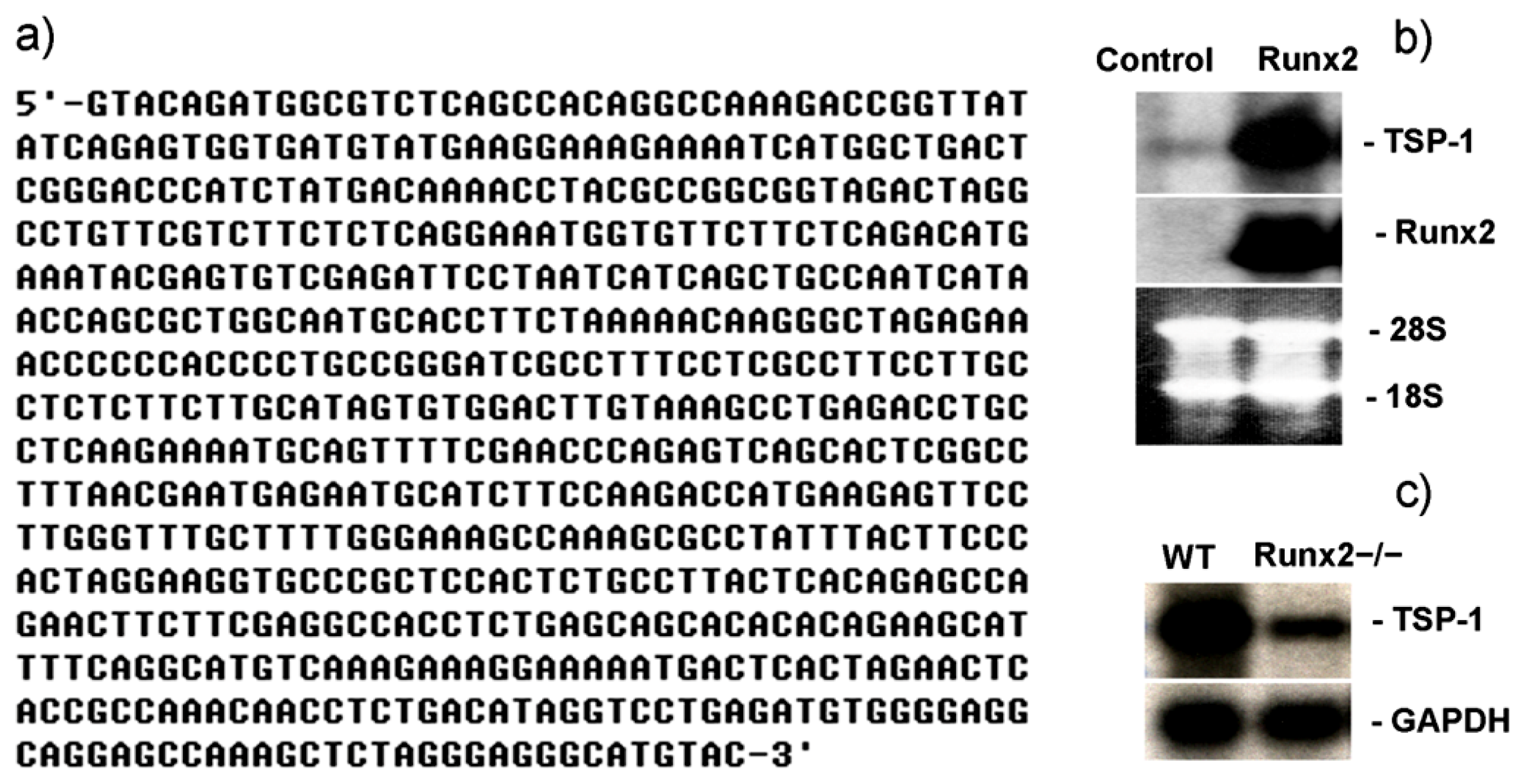
2.1. Runx2 Mediated Regulation of TSP-1 Gene Transcription in Mesenchymal Stem Cells
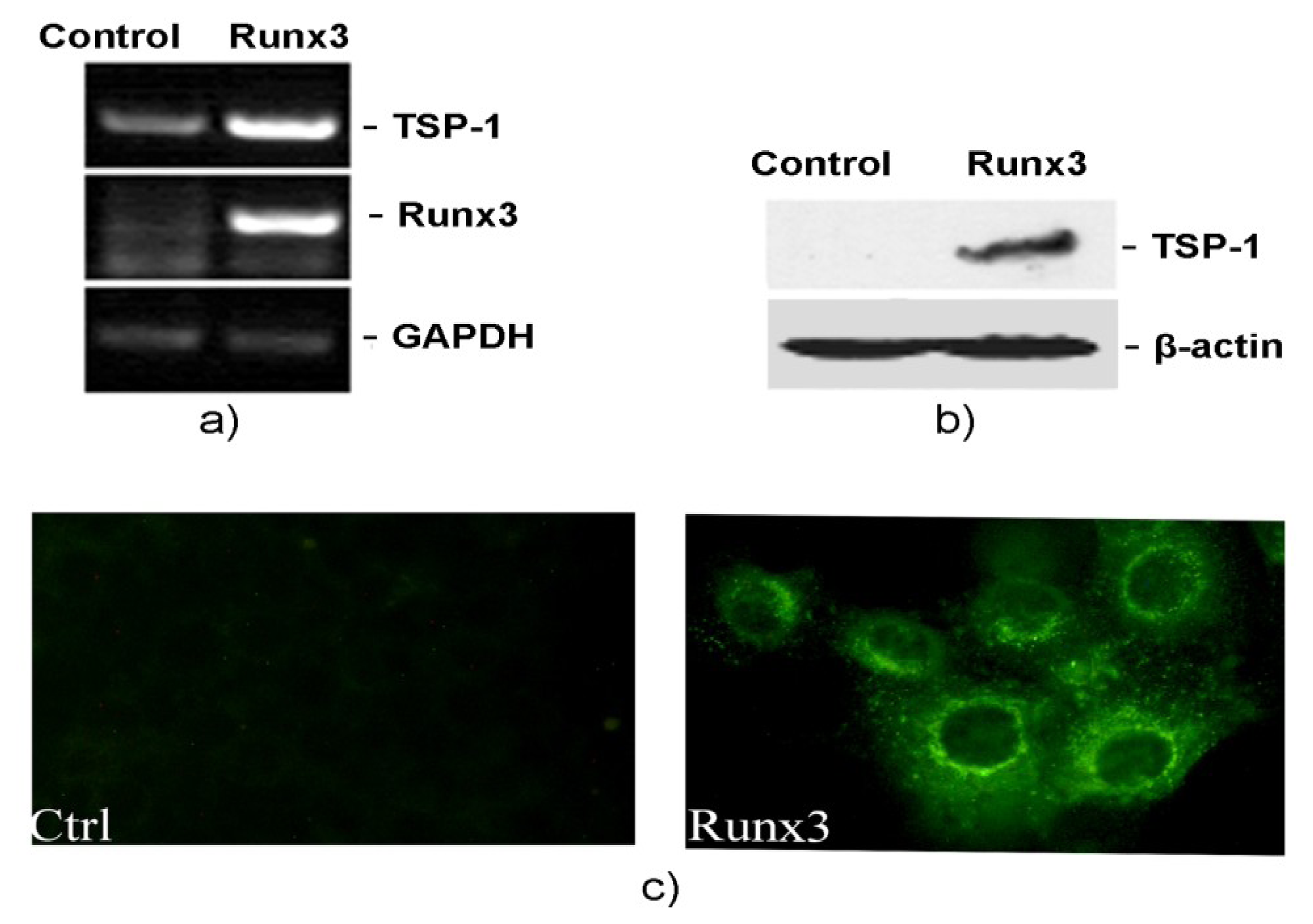
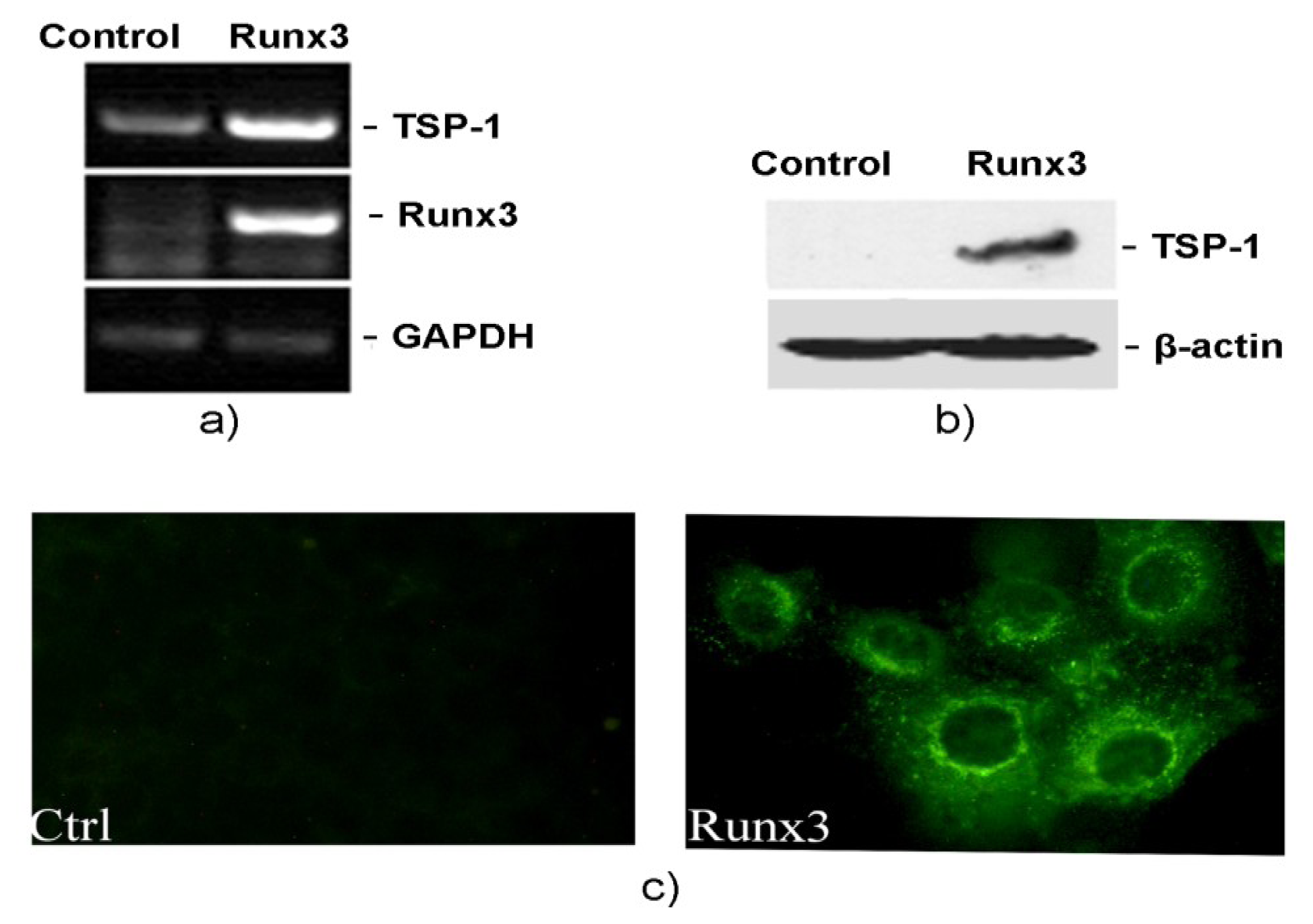
2.2. Runx3 Mediated Regulation of TSP-1 Gene Expression in B16-F10 Melanoma Cells
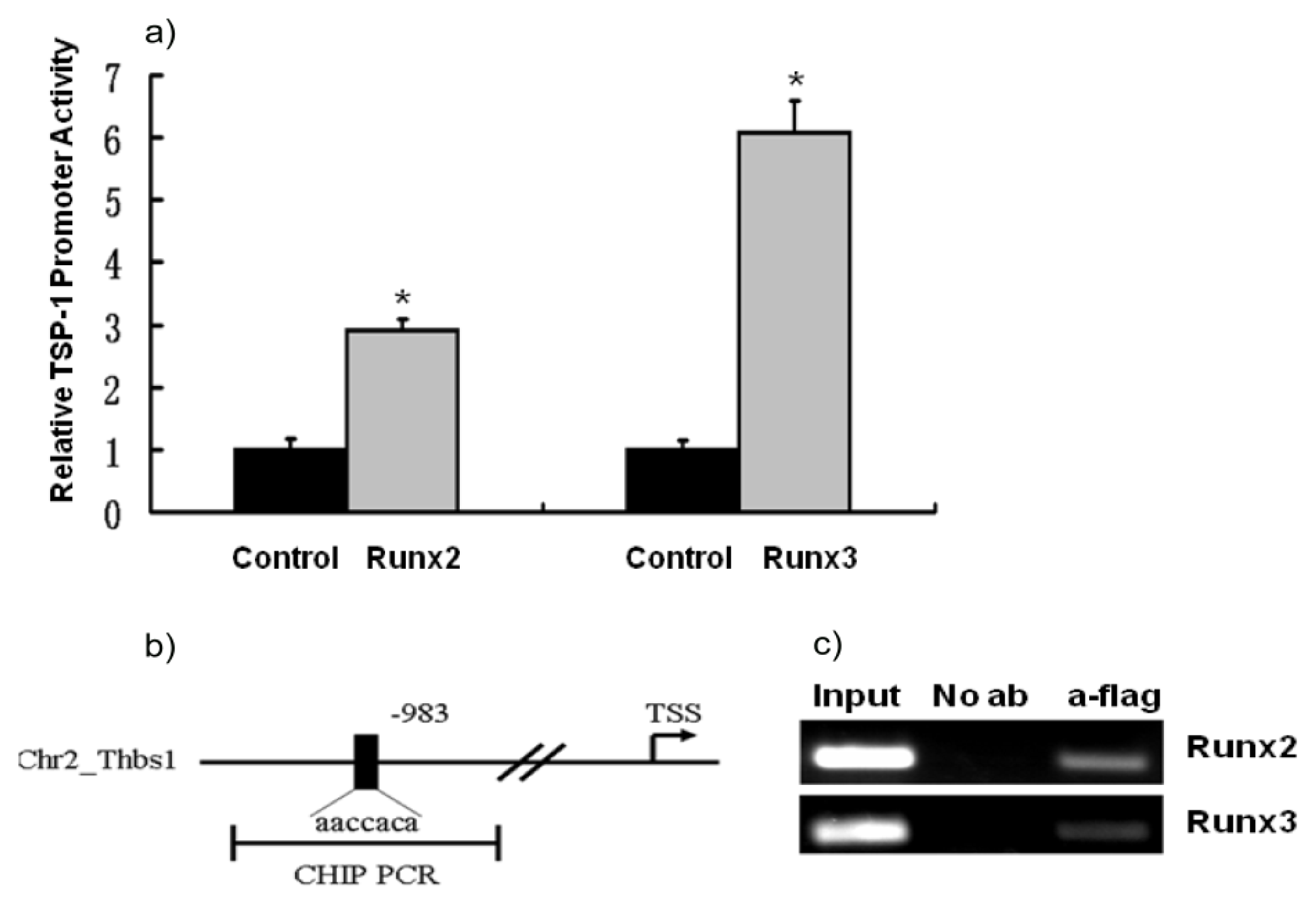
2.3. TSP-1 Gene Promoter Is Regulated by Runx2 and Runx3
3. Experimental Section
3.1. Cell Culture
3.2. cDNA Subtraction
3.3. RNA Preparation, Northern Blot and RT-PCR
3.4. Transient Transfection and Luciferase Assay
3.5. Chromatin Immunoprecipitation (CHIP)
3.6. Western Blot and Immunofluorescence
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Murphy-Ullrich, J.E.; Iozzo, R.V. Thrombospondins in physiology and disease: New tricks for old dogs. Matrix Biol. J. Int. Soc 2012, 31, 152–154. [Google Scholar]
- Sherbina, N.V.; Bornstein, P. Modulation of thrombospondin gene expression during osteoblast differentiation in MC3T3-E1 cells. Bone 1992, 13, 197–201. [Google Scholar]
- Varani, J.; Riser, B.L.; Hughes, L.A.; Carey, T.E.; Fligiel, S.E.; Dixit, V.M. Characterization of thrombospondin synthesis, secretion and cell surface expression by human tumor cells. Clin. Exp. Metastasis 1989, 7, 265–276. [Google Scholar]
- Clezardin, P.; Jouishomme, H.; Chavassieux, P.; Marie, P.J. Thrombospondin is synthesized and secreted by human osteoblasts and osteosarcoma cells: A model to study the different effects of thrombospondin in cell adhesion. Eur. J. Biochem 1989, 181, 721–726. [Google Scholar]
- Varma, V.; Yao-Borengasser, A.; Bodles, A.M.; Rasouli, N.; Phanavanh, B.; Nolen, G.T.; Kern, E.M.; Nagarajan, R.; Spencer, H.J., III; Lee, M.J.; et al. Thrombospondin-1 is an adipokine associated with obesity, adipose inflammation, and insulin resistance. Diabetes 2008, 57, 432–439. [Google Scholar]
- Adams, J.C.; Lawler, J. The thrombospondins. Int. J. Biochem. Cell. Biol 2004, 36, 961–968. [Google Scholar]
- Lopez-Dee, Z.; Pidcock, K.; Gutierrez, L.S. Thrombospondin-1: Multiple paths to inflammation. Mediat. Inflamm 2011, 2011, 296069, :1–296069:10.. [Google Scholar]
- Silverstein, R.L.; Febbraio, M. CD36-TSP-HRGP interactions in the regulation of angiogenesis. Curr. Pharm. Des 2007, 13, 3559–3567. [Google Scholar]
- Kazerounian, S.; Yee, K.O.; Lawler, J. Thrombospondins in cancer. Cell. Mol. Life Sci 2008, 65, 700–712. [Google Scholar]
- Rusnati, M.; Urbinati, C.; Bonifacio, S.; Presta, M.; Taraboletti, G. Thrombospondin-1 as a paradigm for the development of antiangiogenic agents endowed with multiple mechanisms of action. Pharmaceuticals 2010, 3, 1241–1278. [Google Scholar]
- Sargiannidou, I.; Zhou, J.; Tuszynski, G.P. The role of thrombospondin-1 in tumor progression. Exp. Biol. Med. (Maywood) 2001, 226, 726–733. [Google Scholar]
- Streit, M.; Velasco, P.; Brown, L.F.; Skobe, M.; Richard, L.; Riccardi, L.; Lawler, J.; Detmar, M. Overexpression of thrombospondin-1 decreases angiogenesis and inhibits the growth of human cutaneous squamous cell carcinomas. Am. J. Pathol 1999, 155, 441–452. [Google Scholar]
- Rodriguez-Manzaneque, J.C.; Lane, T.F.; Ortega, M.A.; Hynes, R.O.; Lawler, J.; Iruela-Arispe, M.L. Thrombospondin-1 suppresses spontaneous tumor growth and inhibits activation of matrix metalloproteinase-9 and mobilization of vascular endothelial growth factor. Proc. Natl. Acad. Sci. USA 2001, 98, 12485–12490. [Google Scholar]
- Volpert, O.V.; Alani, R.M. Wiring the angiogenic switch: Ras, myc, and thrombospondin-1. Cancer Cell 2003, 3, 199–200. [Google Scholar]
- Zhang, X.; Lawler, J. Thrombospondin-based antiangiogenic therapy. Microvasc. Res 2007, 74, 90–99. [Google Scholar]
- Martin-Manso, G.; Galli, S.; Ridnour, L.A.; Tsokos, M.; Wink, D.A.; Roberts, D.D. Thrombospondin 1 promotes tumor macrophage recruitment and enhances tumor cell cytotoxicity of differentiated U937 cells. Cancer Res 2008, 68, 7090–7099. [Google Scholar]
- Carron, J.A.; Bowler, W.B.; Wagstaff, S.C.; Gallagher, J.A. Expression of members of the thrombospondin family by human skeletal tissues and cultured cells. Biochem. Biophys. Res. Commun 1999, 263, 389–391. [Google Scholar]
- Robey, P.G.; Young, M.F.; Fisher, L.W.; McClain, T.D. Thrombospondin is an osteoblast-derived component of mineralized extracellular matrix. J. Cell Biol 1989, 108, 719–727. [Google Scholar]
- Ueno, A.; Yamashita, K.; Nagata, T.; Tsurumi, C.; Miwa, Y.; Kitamura, S.; Inoue, H. cDNA cloning of bovine thrombospondin 1 and its expression in odontoblasts and predentin. Biochim. Biophys. Acta 1998, 1382, 17–22. [Google Scholar]
- Nishiwaki, T.; Yamaguchi, T.; Zhao, C.; Amano, H.; Hankenson, K.D.; Bornstein, P.; Toyama, Y.; Matsuo, K. Reduced expression of thrombospondins and craniofacial dysmorphism in mice overexpressing Fra1. J. Bone Miner. Res 2006, 21, 596–604. [Google Scholar]
- Ueno, A.; Miwa, Y.; Miyoshi, K.; Horiguchi, T.; Inoue, H.; Ruspita, I.; Abe, K.; Yamashita, K.; Hayashi, E.; Noma, T. Constitutive expression of thrombospondin 1 in MC3T3-E1 osteoblastic cells inhibits mineralization. J. Cell. Physiol 2006, 209, 322–332. [Google Scholar]
- Gelse, K.; Klinger, P.; Koch, M.; Surmann-Schmitt, C.; von der Mark, K.; Swoboda, B.; Hennig, F.F.; Gusinde, J. Thrombospondin-1 prevents excessive ossification in cartilage repair tissue induced by osteogenic protein-1. Tissue Eng. Part A 2011, 17, 2101–2112. [Google Scholar]
- Bailey Dubose, K.; Zayzafoon, M.; Murphy-Ullrich, J.E. Thrombospondin-1 inhibits osteogenic differentiation of human mesenchymal stem cells through latent TGF-β activation. Biochem. Biophys. Res. Commun 2012, 422, 488–493. [Google Scholar]
- Nimmo, R.; Woollard, A. Worming out the biology of Runx. Dev. Biol 2008, 313, 492–500. [Google Scholar]
- Komori, T. Signaling networks in RUNX2-dependent bone development. J. Cell. Biochem 2011, 112, 750–755. [Google Scholar]
- Kitago, M.; Martinez, S.R.; Nakamura, T.; Sim, M.S.; Hoon, D.S. Regulation of RUNX3 tumor suppressor gene expression in cutaneous melanoma. Clin. Cancer Res 2009, 15, 2988–2994. [Google Scholar]
- Martinez, S.; Kuo, C.; Bilchik, A.; Kim, J.; Morton, D.; Hoon, D. Downregulation of tumor suppressor runx genes in malignant cutaneous melanoma. J. Clin. Oncol 2005, 23, S7531. [Google Scholar]
- Komori, T.; Yagi, H.; Nomura, S.; Yamaguchi, A.; Sasaki, K.; Deguchi, K.; Shimizu, Y.; Bronson, R.T.; Gao, Y.H.; Inada, M.; et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell 1997, 89, 755–764. [Google Scholar]
- Mundlos, S.; Otto, F.; Mundlos, C.; Mulliken, J.B.; Aylsworth, A.S.; Albright, S.; Lindhout, D.; Cole, W.G.; Henn, W.; Knoll, J.H.; et al. Mutations involving the transcription factor CBFA1 cause cleidocranial dysplasia. Cell 1997, 89, 773–779. [Google Scholar]
- Liu, W.; Toyosawa, S.; Furuichi, T.; Kanatani, N.; Yoshida, C.; Liu, Y.; Himeno, M.; Narai, S.; Yamaguchi, A.; Komori, T. Overexpression of Cbfa1 in osteoblasts inhibits osteoblast maturation and causes osteopenia with multiple fractures. J. Cell Biol 2001, 155, 157–166. [Google Scholar]
- Komori, T. Regulation of bone development and extracellular matrix protein genes by Runx2. Cell Tissue Res 2010, 339, 189–195. [Google Scholar]
- Deepak, V.; Zhang, Z.; Meng, L.; Zeng, X.; Liu, W. Reduced activity and cytoplasmic localization of Runx2 is observed in C3h10t1/2 cells overexpressing Tbx3. Cell Biochem. Funct 2011, 29, 348–350. [Google Scholar]
- Xiao, Z.S.; Simpson, L.G.; Quarles, L.D. IRES-dependent translational control of Cbfa1/Runx2 expression. J. Cell. Biochem 2003, 88, 493–505. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar]
- Zhang, Y.W.; Su, Y.; Volpert, O.V.; van de Woude, G.F. Hepatocyte growth factor/scatter factor mediates angiogenesis through positive VEGF and negative thrombospondin 1 regulation. Proc. Natl. Acad. Sci. USA 2003, 100, 12718–12723. [Google Scholar]
- Weinstat-Saslow, D.L.; Zabrenetzky, V.S.; VanHoutte, K.; Frazier, W.A.; Roberts, D.D.; Steeg, P.S. Transfection of thrombospondin 1 complementary DNA into a human breast carcinoma cell line reduces primary tumor growth, metastatic potential, and angiogenesis. Cancer Res 1994, 54, 6504–6511. [Google Scholar]
- Kalas, W.; Yu, J.L.; Milsom, C.; Rosenfeld, J.; Benezra, R.; Bornstein, P.; Rak, J. Oncogenes and Angiogenesis: Down-regulation of thrombospondin-1 in normal fibroblasts exposed to factors from cancer cells harboring mutant ras. Cancer Res 2005, 65, 8878–8886. [Google Scholar]
- Chen, F.; Bai, J.; Li, W.; Mei, P.; Liu, H.; Li, L.; Pan, Z.; Wu, Y.; Zheng, J. Runx3 suppresses migration, invasion and angiogenesis of human renal cell carcinoma. PLoS One 2013, 8, e56241. [Google Scholar]
- Lee, J.M.; Lee, D.J.; Bae, S.C.; Jung, H.S. Abnormal liver differentiation and excessive angiogenesis in mice lacking Runx3. Histochem. Cell Biol 2013, 139, 751–758. [Google Scholar]
- Peng, Z.; Wei, D.; Wang, L.; Tang, H.; Zhang, J.; Le, X.; Jia, Z.; Li, Q.; Xie, K. Runx3 inhibits the expression of vascular endothelial growth factor and reduces the angiogenesis, growth, and metastasis of human gastric cancer. Clin. Cancer Res 2006, 12, 6386–6394. [Google Scholar]
- Subramaniam, M.M.; Chan, J.Y.; Yeoh, K.G.; Quek, T.; Ito, K.; Salto-Tellez, M. Molecular pathology of Runx3 in human carcinogenesis. Biochim. Biophys. Acta 2009, 1796, 315–331. [Google Scholar]
- Roberts, D.D. Regulation of tumor growth and metastasis by thrombospondin-1. FASEB J 1996, 10, 1183–1191. [Google Scholar]
- Shingu, T.; Bornstein, P. Overlapping Egr-1 and Sp1 sites function in the regulation of transcription of the mouse thrombospondin 1 gene. J. Biol. Chem 1994, 269, 32551–32557. [Google Scholar]
- Zhang, Z.; Deepak, V.; Meng, L.; Wang, L.; Li, Y.; Jiang, Q.; Zeng, X.; Liu, W. Analysis of HDAC1-mediated regulation of Runx2-induced osteopontin gene expression in C3h10t1/2 cells. Biotechnol. Lett 2012, 34, 197–203. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| GAPDH | CTCATGACCACAGTCCATGC | CACATTGGGGGTAGGAACAC |
| Runx3 | CTCGATGGTGGACGTGCTGG | ACCTTGATGGCTCGGTGGTAGG |
| TSP-1 | GAGTGCAAAGAAGTGCCTGATG | GGAATGGACAGTTGTCCCTGTC |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shi, X.; Deepak, V.; Wang, L.; Ba, X.; Komori, T.; Zeng, X.; Liu, W. Thrombospondin-1 Is a Putative Target Gene of Runx2 and Runx3. Int. J. Mol. Sci. 2013, 14, 14321-14332. https://doi.org/10.3390/ijms140714321
Shi X, Deepak V, Wang L, Ba X, Komori T, Zeng X, Liu W. Thrombospondin-1 Is a Putative Target Gene of Runx2 and Runx3. International Journal of Molecular Sciences. 2013; 14(7):14321-14332. https://doi.org/10.3390/ijms140714321
Chicago/Turabian StyleShi, Xiuming, Vishwa Deepak, Linghui Wang, Xueqing Ba, Toshihisa Komori, Xianlu Zeng, and Wenguang Liu. 2013. "Thrombospondin-1 Is a Putative Target Gene of Runx2 and Runx3" International Journal of Molecular Sciences 14, no. 7: 14321-14332. https://doi.org/10.3390/ijms140714321