Evaluation of the Gamma-H2AX Assay for Radiation Biodosimetry in a Swine Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
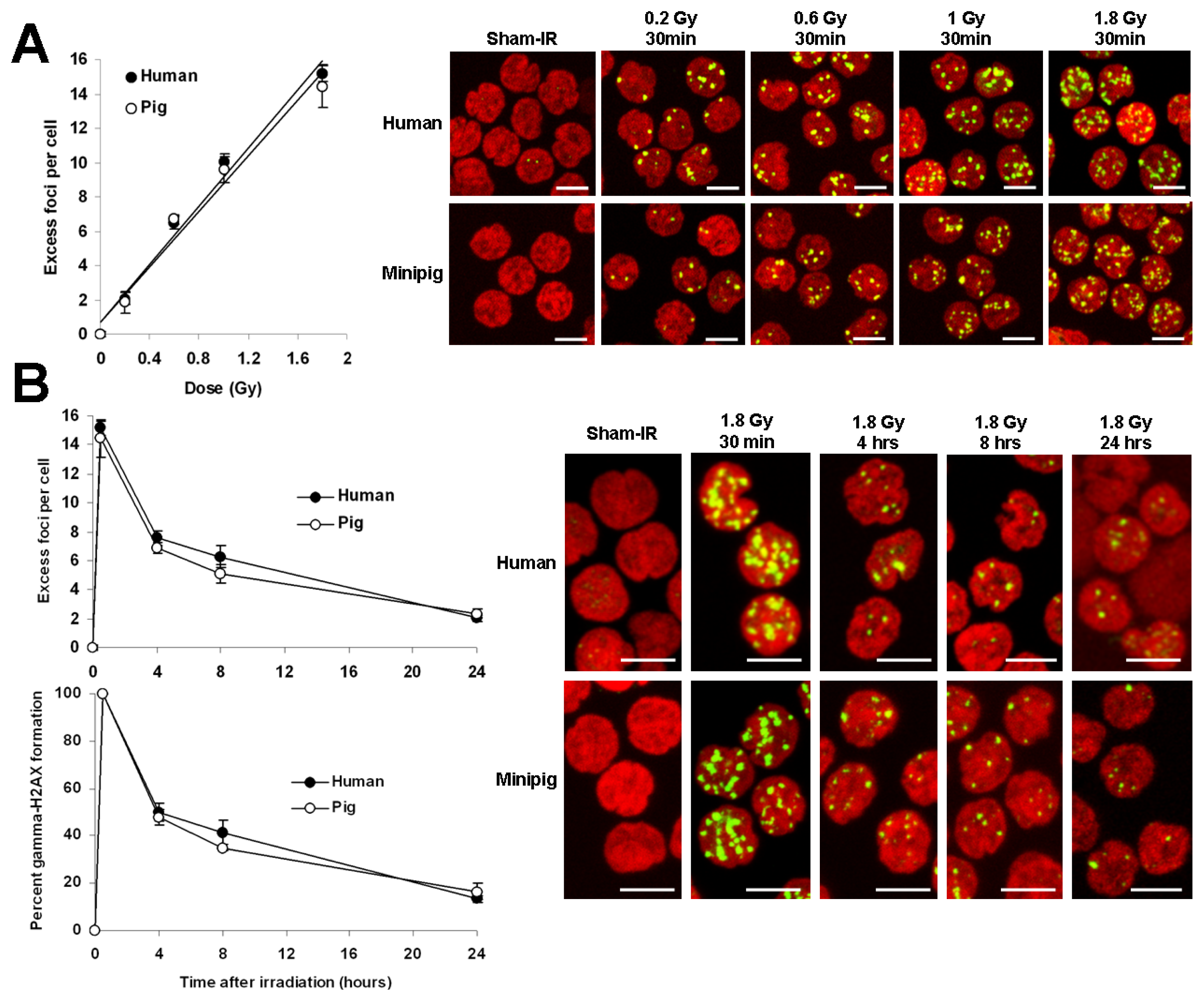
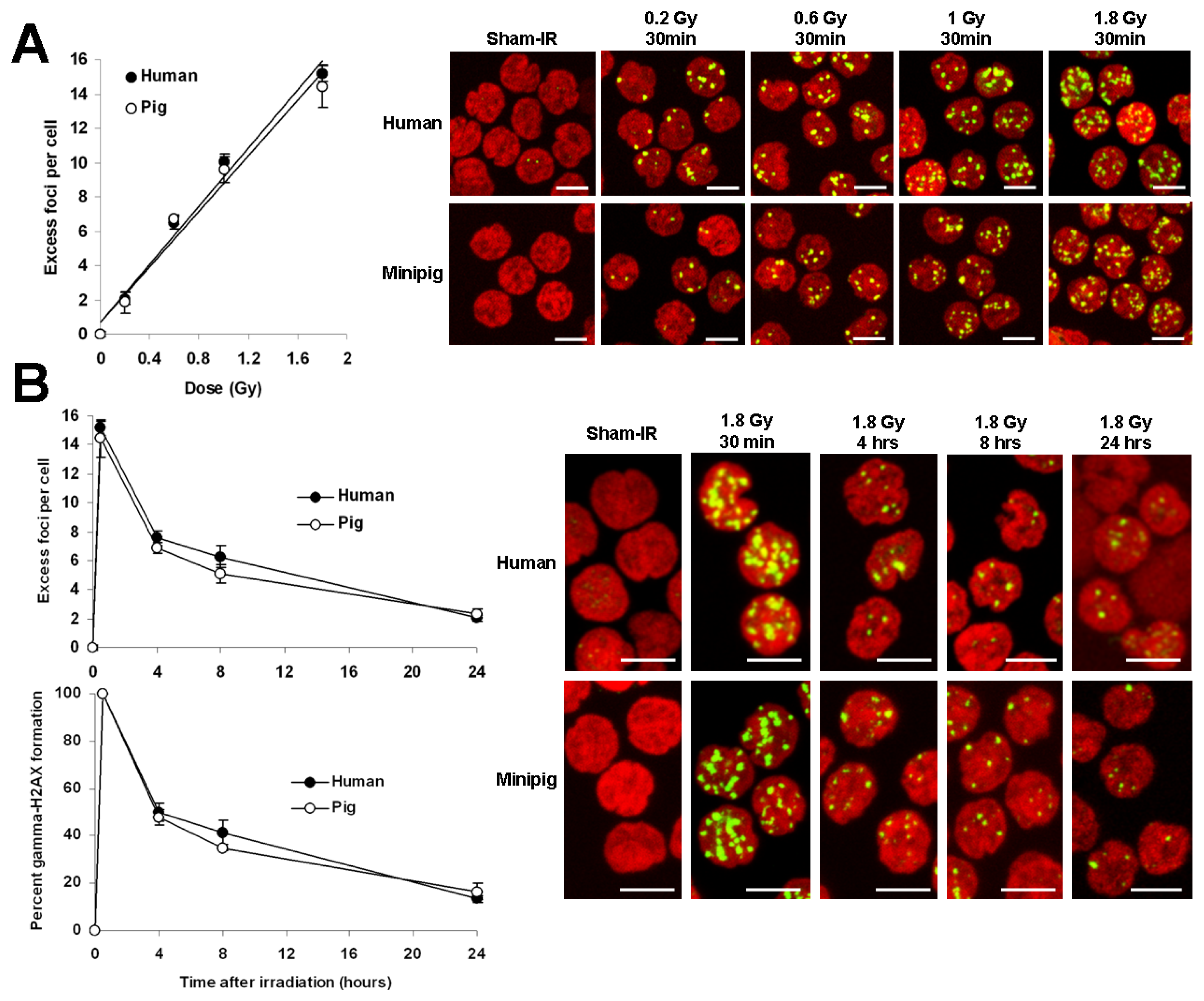
2.1. Comparative Analysis of γ-H2AX Kinetics in Human and Minipig Lymphocytes after ex Vivo Irradiation
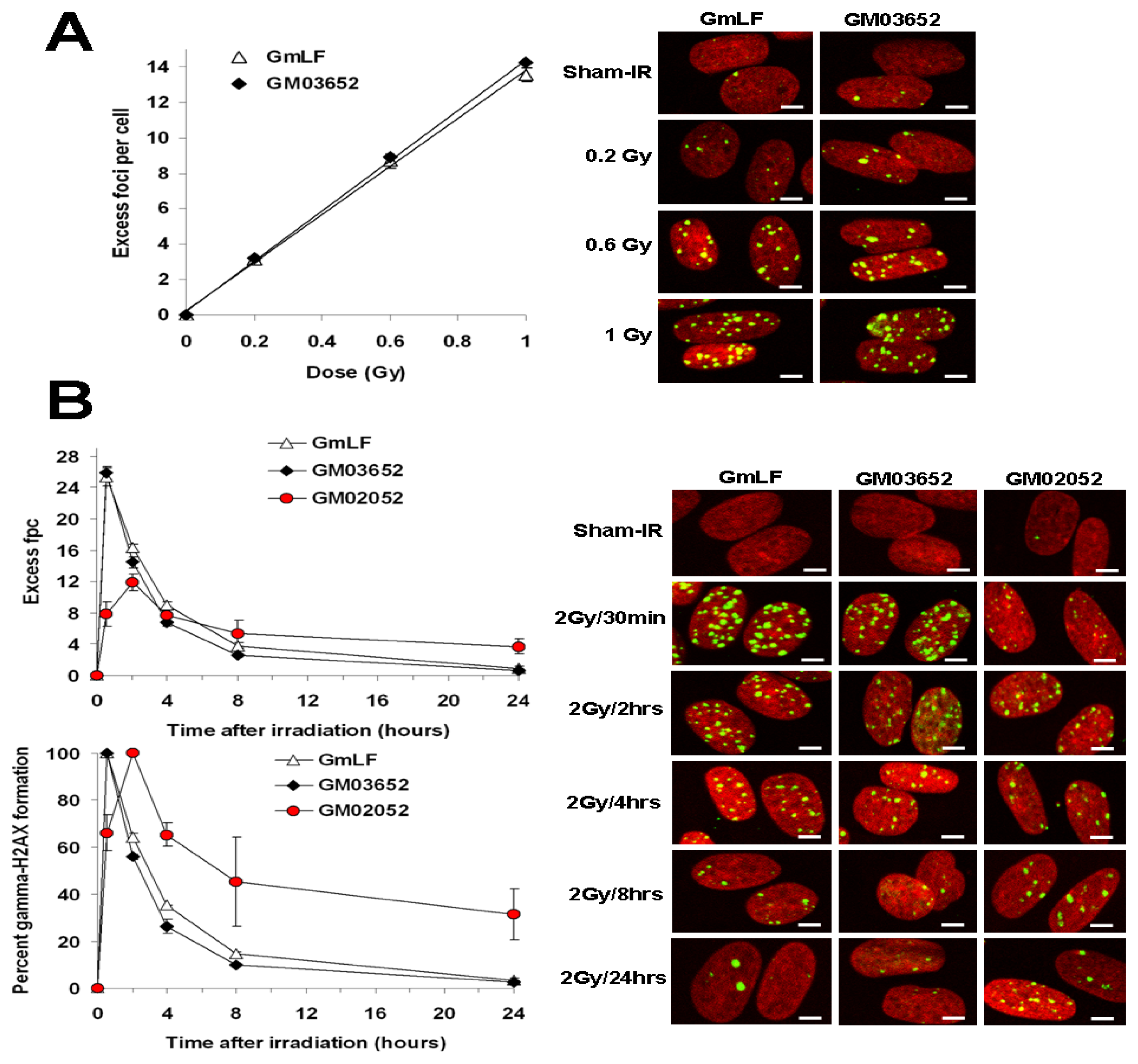
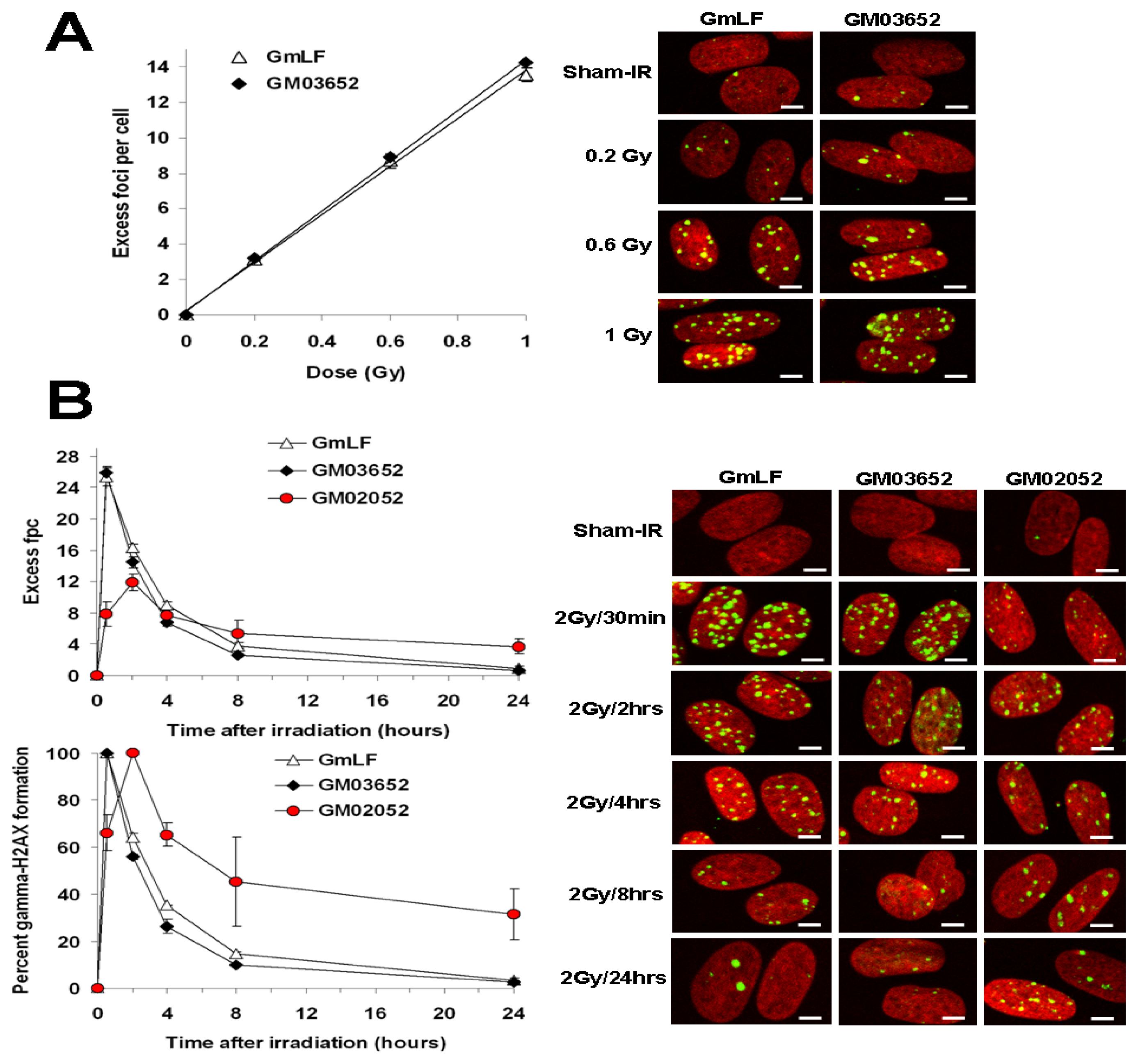
2.2. Comparative Analysis of γ-H2AX Kinetics in Human and Minipig Fibroblasts
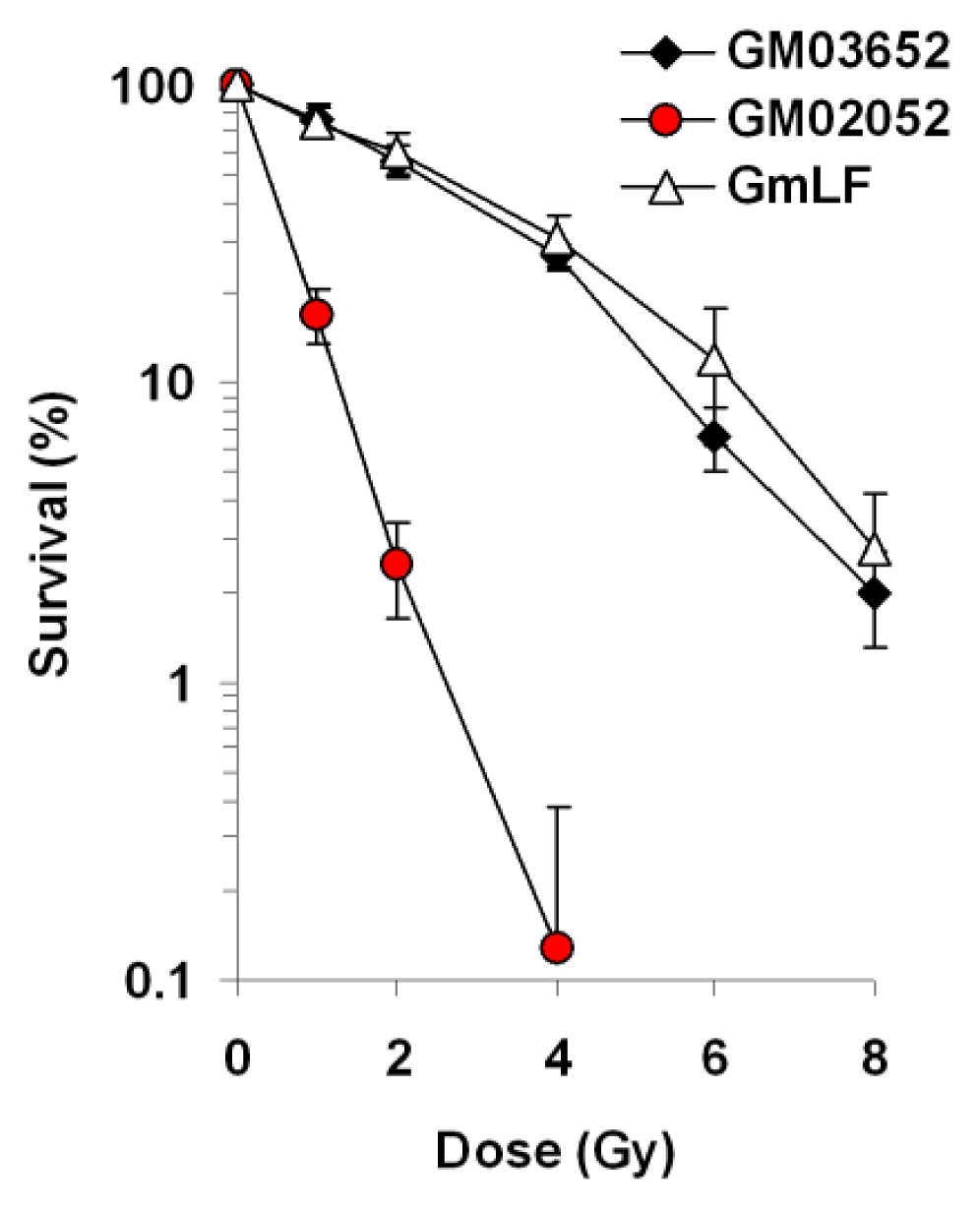
2.3. Comparative Analysis of Radiation Sensitivity in Human and Minipig Fibroblasts
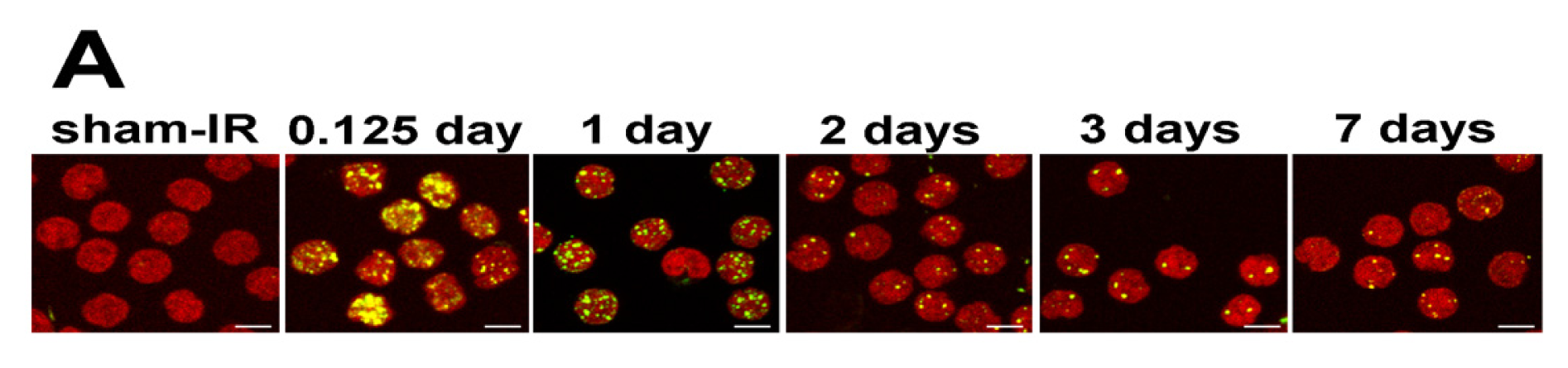
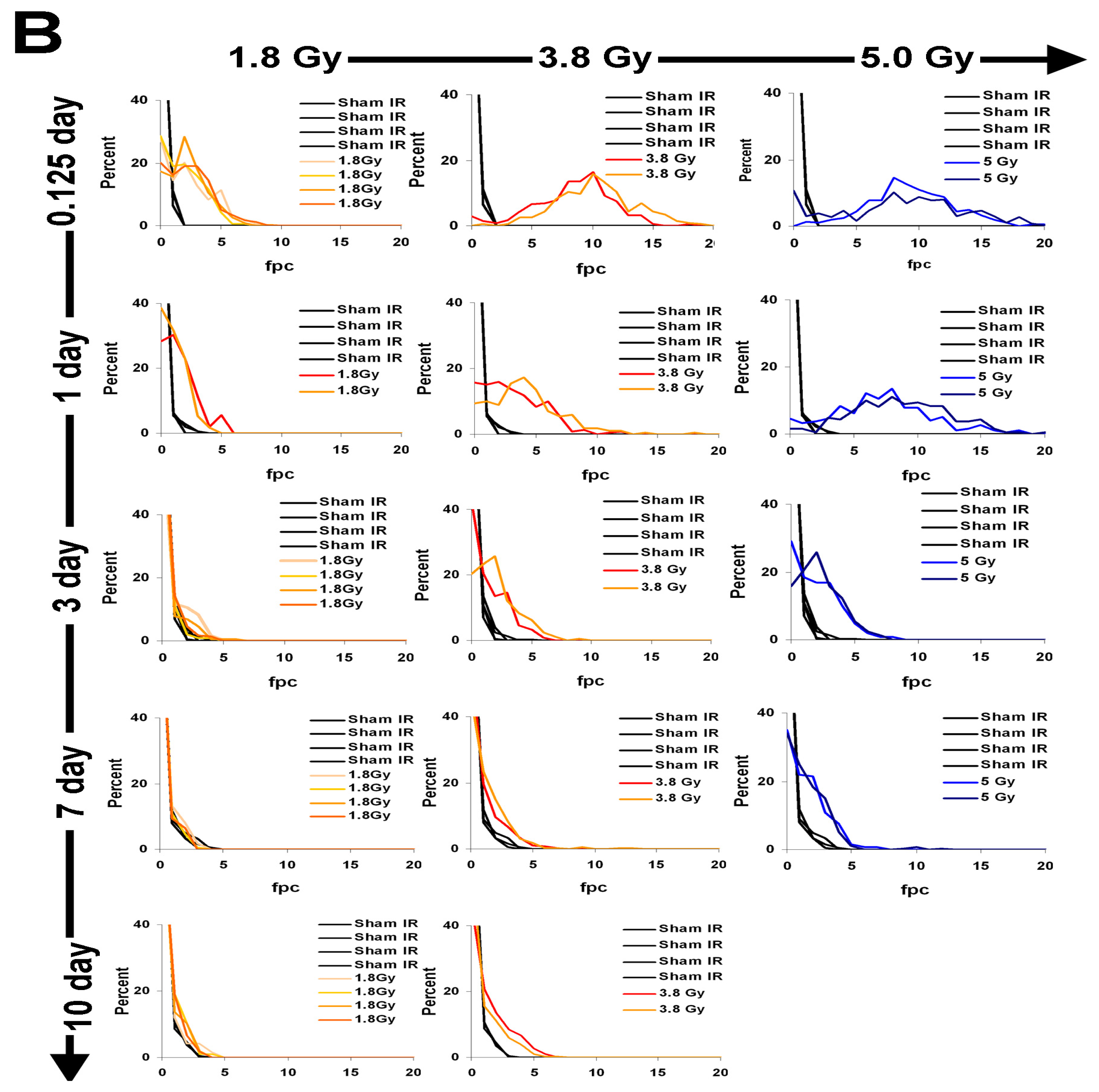
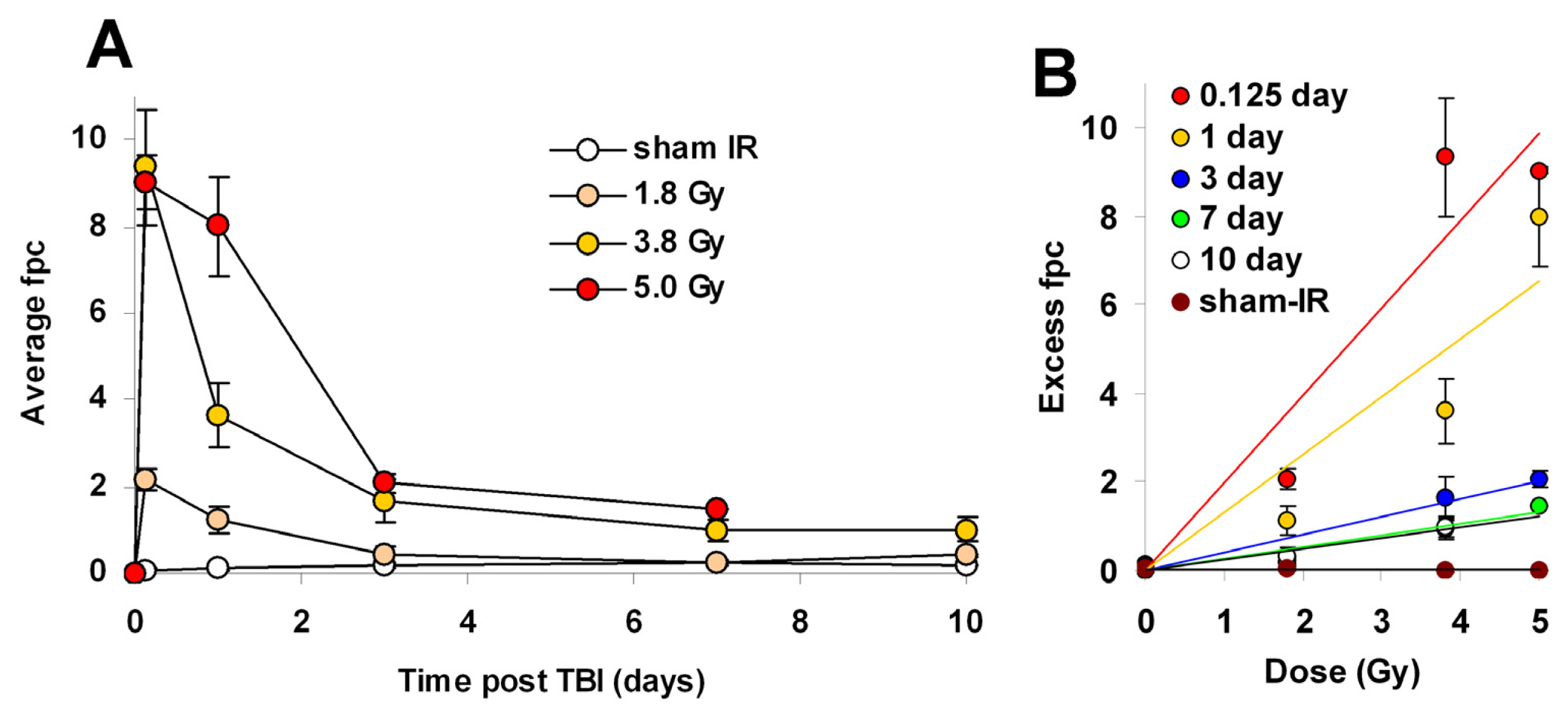
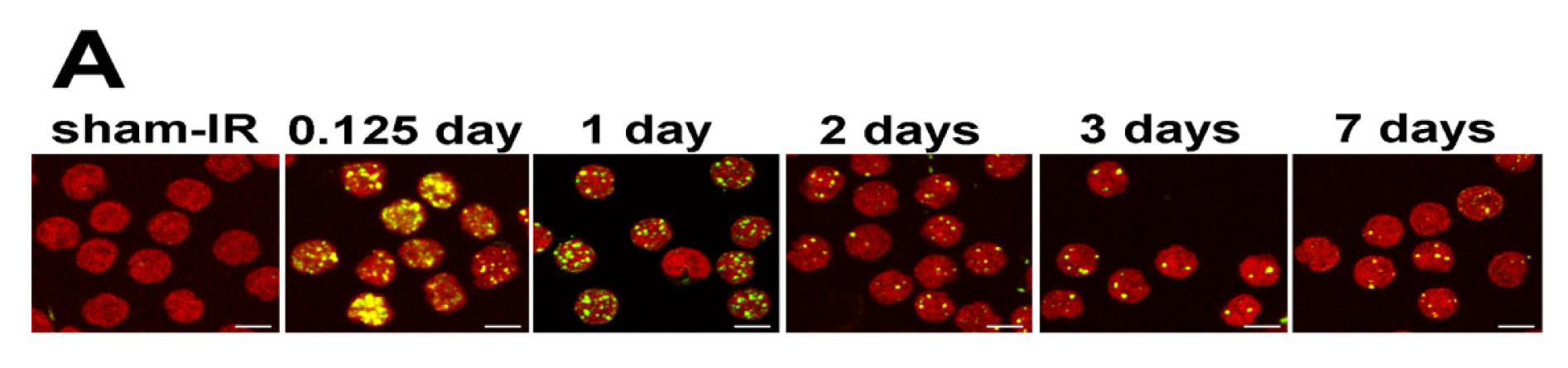
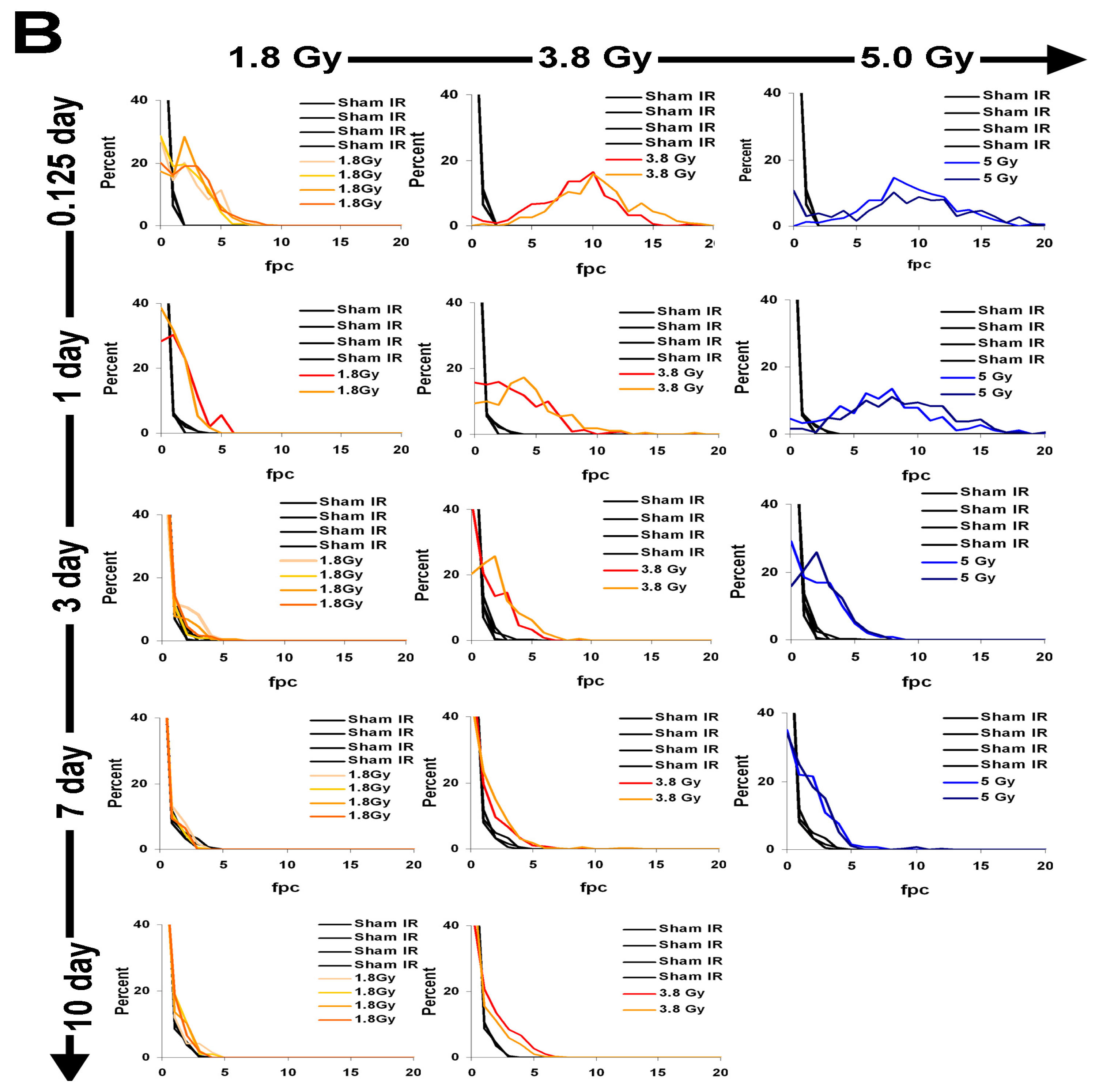
2.4. γ-H2AX Kinetics after Total-Body Irradiation of Minipigs
3. Experimental Section
3.1. Reagents
3.2. Cell Line and Cell Culture
3.3. Animal Housing and Care
3.4. Animal Irradiation Procedures
3.5. Blood Sampling
3.6. Sample Irradiation and Lymphocyte Isolation
3.7. γ-H2AX Detection
3.8. Clonogenic Survival Assay
3.9. Statistical Analysis
4. Conclusions
Acknowledgements
Conflict of Interest
References
- Brenner, D.J.; Hall, E.J. Computed tomography—An increasing source of radiation exposure. N. Engl. J. Med 2007, 357, 2277–2284. [Google Scholar]
- Saenko, V.; Ivanov, V.; Tsyb, A.; Bogdanova, T.; Tronko, M.; Demidchik, Y.; Yamashita, S. The Chernobyl accident and its consequences. Clin. Oncol 2011, 23, 234–243. [Google Scholar]
- Thielen, H. The Fukushima Daiichi nuclear accident—An overview. Health Phys 2012, 103, 169–174. [Google Scholar]
- Augustine, A.D.; Gondre-Lewis, T.; McBride, W.; Miller, L.; Pellmar, T.C.; Rockwell, S. Animal models for radiation injury, protection and therapy. Radiat. Res 2005, 164, 100–109. [Google Scholar]
- Waselenko, J.K.; MacVittie, T.J.; Blakely, W.F.; Pesik, N.; Wiley, A.L.; Dickerson, W.E.; Tsu, H.; Confer, D.L.; Coleman, C.N.; Seed, T.; et al. Medical management of the acute radiation syndrome: Recommendations of the Strategic National Stockpile Radiation Working Group. Ann. Intern. Med 2004, 140, 1037–1051. [Google Scholar]
- Stubblefield, M.D. Radiation fibrosis syndrome: Neuromuscular and musculoskeletal complications in cancer survivors. PM R 2011, 3, 1041–1054. [Google Scholar]
- Shah, D.J.; Sachs, R.K.; Wilson, D.J. Radiation-induced cancer: A modern view. Br. J. Radiol 2012, 85, e1166–e1173. [Google Scholar]
- Dorr, H.; Meineke, V. Acute radiation syndrome caused by accidental radiation exposure-therapeutic principles. BMC Med 2011, 9, 126. [Google Scholar]
- Jackson, I.L.; Xu, P.; Hadley, C.; Katz, B.P.; McGurk, R.; Down, J.D.; Vujaskovic, Z. A preclinical rodent model of radiation-induced lung injury for medical countermeasure screening in accordance with the FDA animal rule. Health Phys 2012, 103, 463–473. [Google Scholar]
- MacVittie, T.J.; Farese, A.M.; Bennett, A.; Gelfond, D.; Shea-Donohue, T.; Tudor, G.; Booth, C.; McFarland, E.; Jackson, W., 3rd. The acute gastrointestinal subsyndrome of the acute radiation syndrome: A rhesus macaque model. Health Phys. 2012, 103, 411–426. [Google Scholar]
- Hu, S.; Cucinotta, F.A. A cell kinetic model of granulopoiesis under radiation exposure: Extension from rodents to canines and humans. Radiat. Prot. Dosimetry 2011, 143, 207–213. [Google Scholar]
- Moroni, M.; Lombardini, E.; Salber, R.; Kazemzedeh, M.; Nagy, V.; Olsen, C.; Whitnall, M.H. Hematological changes as prognostic indicators of survival: Similarities between Gottingen minipigs, humans, and other large animal models. PLoS One 2011, 6, e25210. [Google Scholar]
- Redon, C.E.; Nakamura, A.J.; Gouliaeva, K.; Rahman, A.; Blakely, W.F.; Bonner, W.M. The use of gamma-H2AX as a biodosimeter for total-body radiation exposure in non-human primates. PLoS One 2010, 5, e15544. [Google Scholar]
- Moroni, M.; Coolbaugh, T.V.; Lombardini, E.; Mitchell, J.M.; Moccia, K.D.; Shelton, L.J.; Nagy, V.; Whitnall, M.H. Hematopoietic radiation syndrome in the Gottingen minipig. Radiat. Res 2011, 176, 89–101. [Google Scholar]
- Jacobs, A. Use of nontraditional animals for evaluation of pharmaceutical products. Expert. Opin. Drug Metab. Toxicol 2006, 2, 345–349. [Google Scholar]
- Aigner, B.; Renner, S.; Kessler, B.; Klymiuk, N.; Kurome, M.; Wunsch, A.; Wolf, E. Transgenic pigs as models for translational biomedical research. J. Mol. Med 2010, 88, 653–664. [Google Scholar]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J., Jr; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Vet. Pathol. 2011, 49, 344–356. [Google Scholar]
- Bollen, P.; Ellegaard, L. The Gottingen minipig in pharmacology and toxicology. Pharmacol. Toxicol 1997, 80, 3–4. [Google Scholar]
- Ahmed, E.A.; Agay, D.; Schrock, G.; Drouet, M.; Meineke, V.; Scherthan, H. Persistent DNA damage after high dose in vivo gamma exposure of minipig skin. PLoS One 2012, 7, e39521. [Google Scholar]
- Hulet, S.W.; Moroni, M.; Whitnall, M.H.; Mioduszewski, R.J. The Minipig in Chemical, Biological and Radiological Research; CRC Press: Boca Raton, FL, USA, 2012; pp. 533–547. [Google Scholar]
- Bonner, W.M.; Redon, C.E.; Dickey, J.S.; Nakamura, A.J.; Sedelnikova, O.A.; Solier, S.; Pommier, Y. GammaH2AX and cancer. Nat. Rev. Cancer 2008, 8, 957–967. [Google Scholar]
- Nakamura, A.; Sedelnikova, O.A.; Redon, C.; Pilch, D.R.; Sinogeeva, N.I.; Shroff, R.; Lichten, M.; Bonner, W.M. Techniques for gamma-H2AX detection. Methods Enzymol 2006, 409, 236–250. [Google Scholar]
- Redon, C.E.; Nakamura, A.J.; Sordet, O.; Dickey, J.S.; Gouliaeva, K.; Tabb, B.; Lawrence, S.; Kinders, R.J.; Bonner, W.M.; Sedelnikova, O.A. gamma-H2AX detection in peripheral blood lymphocytes, splenocytes, bone marrow, xenografts, and skin. Methods Mol. Biol 2011, 682, 249–270. [Google Scholar]
- Rothkamm, K.; Lobrich, M. Evidence for a lack of DNA double-strand break repair in human cells exposed to very low X-ray doses. Proc. Natl. Acad. Sci. USA 2003, 100, 5057–5062. [Google Scholar]
- Sedelnikova, O.A.; Rogakou, E.P.; Panyutin, I.G.; Bonner, W.M. Quantitative detection of (125)IdU-induced DNA double-strand breaks with gamma-H2AX antibody. Radiat. Res 2002, 158, 486–492. [Google Scholar]
- Marchese, M.J.; Zaider, M.; Hall, E.J. Potentially lethal damage repair in human cells. Radiother. Oncol 1987, 9, 57–65. [Google Scholar]
- Dileto, C.L.; Travis, E.L. Fibroblast radiosensitivity in vitro and lung fibrosis in vivo: Comparison between a fibrosis-prone and fibrosis-resistant mouse strain. Radiat. Res 1996, 146, 61–67. [Google Scholar]
- Bertrand, O.F.; Mongrain, R.; Thorin, E.; Lehnert, S. In vitro response of human and porcine vascular cells exposed to high dose-rate gamma-irradiation. Int. J. Radiat. Biol 2000, 76, 999–1007. [Google Scholar]
- Redon, C.E.; Weyemi, U.; Parekh, P.R.; Huang, D.; Burrell, A.S.; Bonner, W.M. Gamma-H2AX and other histone post-translational modifications in the clinic. Biochim. Biophys. Acta 2012, 1819, 743–756. [Google Scholar]
- Markova, E.; Schultz, N.; Belyaev, I.Y. Kinetics and dose-response of residual 53BP1/gamma-H2AX foci: Co-localization, relationship with DSB repair and clonogenic survival. Int. J. Radiat. Biol 2007, 83, 319–329. [Google Scholar]
- Redon, C.; Dickey, J.S.; Bonner, W.M.; Sedelnikova, O. Gamma-H2AX as a biomarker of DNA damage induced by ionizing radiation in human peripheral blood lymphocytes and artificial skin. Adv. Space Res 2009, 43, 1171–1178. [Google Scholar]
- Markova, E.; Torudd, J.; Belyaev, I. Long time persistence of residual 53BP1/gamma-H2AX foci in human lymphocytes in relationship to apoptosis, chromatin condensation and biological dosimetry. Int. J. Radiat. Biol 2011, 87, 736–745. [Google Scholar]
- Kato, T.A.; Okayasu, R.; Bedford, J.S. Comparison of the induction and disappearance of DNA double strand breaks and gamma-H2AX foci after irradiation of chromosomes in G1-phase or in condensed metaphase cells. Mutat. Res 2008, 639, 108–112. [Google Scholar]
- Beucher, A.; Birraux, J.; Tchouandong, L.; Barton, O.; Shibata, A.; Conrad, S.; Goodarzi, A.A.; Krempler, A.; Jeggo, P.A.; Lobrich, M. ATM and Artemis promote homologous recombination of radiation-induced DNA double-strand breaks in G2. EMBO J 2009, 28, 3413–3427. [Google Scholar]
- McPherson, J.D.; Marra, M.; Hillier, L.; Waterston, R.H.; Chinwalla, A.; Wallis, J.; Sekhon, M.; Wylie, K.; Mardis, E.R.; Wilson, R.K.; et al. A physical map of the human genome. Nature 2001, 409, 934–941. [Google Scholar]
- Schook, L.B.; Beever, J.E.; Rogers, J.; Humphray, S.; Archibald, A.; Chardon, P.; Milan, D.; Rohrer, G.; Eversole, K. Swine Genome Sequencing Consortium (SGSC): A strategic roadmap for sequencing the pig genome. Comp. Funct. Genomics 2005, 6, 251–255. [Google Scholar]
- Forster, R.; Ancian, P.; Fredholm, M.; Simianer, H.; Whitelaw, B. The minipig as a platform for new technologies in toxicology. J. Pharmacol. Toxicol. Methods 2010, 62, 227–235. [Google Scholar]
- Dawson, H.D. A Comparative Assessment of the Pig, Mouse and Human Genomes. In The Minipig in Biomedical Research; McAnulty, P.A., Dayan, A.D., Ganderup, N.C., Hastings, K.L., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 323–342. [Google Scholar]
- Shiloh, Y. ATM and related protein kinases: Safeguarding genome integrity. Nat. Rev. Cancer 2003, 3, 155–168. [Google Scholar]
- Stewart, G.S.; Maser, R.S.; Stankovic, T.; Bressan, D.A.; Kaplan, M.I.; Jaspers, N.G.; Raams, A.; Byrd, P.J.; Petrini, J.H.; Taylor, A.M. The DNA double-strand break repair gene hMRE11 is mutated in individuals with an ataxia-telangiectasia-like disorder. Cell 1999, 99, 577–587. [Google Scholar]
- Hernandez, D.; McConville, C.M.; Stacey, M.; Woods, C.G.; Brown, M.M.; Shutt, P.; Rysiecki, G.; Taylor, A.M. A family showing no evidence of linkage between the ataxia telangiectasia gene and chromosome 11q22–23. J. Med. Genet 1993, 30, 135–140. [Google Scholar]
- Atsumi, Y.; Fujimori, H.; Fukuda, H.; Inase, A.; Shinohe, K.; Yoshioka, Y.; Shikanai, M.; Ichijima, Y.; Unno, J.; Mizutani, S.; et al. Onset of quiescence following p53 mediated down-regulation of H2AX in normal cells. PLoS One 2011, 6, e23432. [Google Scholar]
- Suzuki, M.; Suzuki, K.; Kodama, S.; Yamashita, S.; Watanabe, M. Persistent amplification of DNA damage signal involved in replicative senescence of normal human diploid fibroblasts. Oxid. Med. Cell Longev 2012, 2012, 310534. [Google Scholar]
- Bhogal, N.; Kaspler, P.; Jalali, F.; Hyrien, O.; Chen, R.; Hill, R.P.; Bristow, R.G. Late residual gamma-H2AX foci in murine skin are dose responsive and predict radiosensitivity in vivo. Radiat. Res 2010, 173, 1–9. [Google Scholar]
- Redon, C.E.; Nakamura, A.J.; Gouliaeva, K.; Rahman, A.; Blakely, W.F.; Bonner, W.M. Q(gamma-H2AX), an analysis method for partial-body radiation exposure using gamma-H2AX in nonhuman primate lymphocytes. Radiat. Meas 2011, 46, 877–881. [Google Scholar]
- Lobrich, M.; Rief, N.; Kuhne, M.; Heckmann, M.; Fleckenstein, J.; Rube, C.; Uder, M. In vivo formation and repair of DNA double-strand breaks after computed tomography examinations. Proc. Natl. Acad. Sci. USA 2005, 102, 8984–8989. [Google Scholar]
- Bourton, E.C.; Plowman, P.N.; Smith, D.; Arlett, C.F.; Parris, C.N. Prolonged expression of the gamma-H2AX DNA repair biomarker correlates with excess acute and chronic toxicity from radiotherapy treatment. Int. J. Cancer 2011, 129, 2928–2934. [Google Scholar]
- Sak, A.; Grehl, S.; Erichsen, P.; Engelhard, M.; Grannass, A.; Levegrun, S.; Pottgen, C.; Groneberg, M.; Stuschke, M. gamma-H2AX foci formation in peripheral blood lymphocytes of tumor patients after local radiotherapy to different sites of the body: Dependence on the dose-distribution, irradiated site and time from start of treatment. Int. J. Radiat. Biol 2007, 83, 639–652. [Google Scholar]
- Beels, L.; Bacher, K.; de Wolf, D.; Werbrouck, J.; Thierens, H. Gamma-H2AX foci as a biomarker for patient X-ray exposure in pediatric cardiac catheterization: Are we underestimating radiation risks? Circulation 2009, 120, 1903–1909. [Google Scholar]
- Kuefner, M.A.; Grudzenski, S.; Schwab, S.A.; Wiederseiner, M.; Heckmann, M.; Bautz, W.; Lobrich, M.; Uder, M. DNA double-strand breaks and their repair in blood lymphocytes of patients undergoing angiographic procedures. Invest. Radiol 2009, 44, 440–446. [Google Scholar]
- Zwicker, F.; Swartman, B.; Sterzing, F.; Major, G.; Weber, K.J.; Huber, P.E.; Thieke, C.; Debus, J.; Herfarth, K. Biological in vivo measurement of dose distribution in patients’ lymphocytes by gamma-H2AX immunofluorescence staining: 3D conformal- vs. step-and-shoot IMRT of the prostate gland. Radiat. Oncol 2011, 6, 62. [Google Scholar]
- Testard, I.; Sabatier, L. Biological dosimetry for astronauts: A real challenge. Mutat. Res 1999, 430, 315–326. [Google Scholar]
- Fenech, M.; Bonassi, S.; Turner, J.; Lando, C.; Ceppi, M.; Chang, W.P.; Holland, N.; Kirsch-Volders, M.; Zeiger, E.; Bigatti, M.P.; et al. Intra- and inter-laboratory variation in the scoring of micronuclei and nucleoplasmic bridges in binucleated human lymphocytes. Results of an international slide-scoring exercise by the HUMN project. Mutat. Res 2003, 534, 45–64. [Google Scholar]
- Turner, H.C.; Brenner, D.J.; Chen, Y.; Bertucci, A.; Zhang, J.; Wang, H.; Lyulko, O.V.; Xu, Y.; Shuryak, I.; Schaefer, J.; et al. Adapting the gamma-H2AX assay for automated processing in human lymphocytes. 1. Technological aspects. Radiat. Res 2011, 175, 282–290. [Google Scholar]
- Rube, C.E.; Fricke, A.; Schneider, R.; Simon, K.; Kuhne, M.; Fleckenstein, J.; Graber, S.; Graf, N.; Rube, C. DNA repair alterations in children with pediatric malignancies: Novel opportunities to identify patients at risk for high-grade toxicities. Int. J. Radiat. Oncol. Biol. Phys 2010, 78, 359–369. [Google Scholar]
- Moroni, M.; Coolbaugh, T.V.; Mitchell, J.M.; Lombardini, E.; Moccia, K.D.; Shelton, L.J.; Nagy, V.; Whitnall, M.H. Vascular access port implantation and serial blood sampling in a Gottingen minipig (Sus scrofa domestica) model of acute radiation injury. J. Am. Assoc. Lab. Anim. Sci 2011, 50, 65–72. [Google Scholar]
- Adam, S.J.; Rund, L.A.; Kuzmuk, K.N.; Zachary, J.F.; Schook, L.B.; Counter, C.M. Genetic induction of tumorigenesis in swine. Oncogene 2007, 26, 1038–1045. [Google Scholar]
- Moraes, M.C.; Neto, J.B.; Menck, C.F. DNA repair mechanisms protect our genome from carcinogenesis. Front. Biosci 2012, 17, 1362–1388. [Google Scholar]


© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Moroni, M.; Maeda, D.; Whitnall, M.H.; Bonner, W.M.; Redon, C.E. Evaluation of the Gamma-H2AX Assay for Radiation Biodosimetry in a Swine Model. Int. J. Mol. Sci. 2013, 14, 14119-14135. https://doi.org/10.3390/ijms140714119
Moroni M, Maeda D, Whitnall MH, Bonner WM, Redon CE. Evaluation of the Gamma-H2AX Assay for Radiation Biodosimetry in a Swine Model. International Journal of Molecular Sciences. 2013; 14(7):14119-14135. https://doi.org/10.3390/ijms140714119
Chicago/Turabian StyleMoroni, Maria, Daisuke Maeda, Mark H. Whitnall, William M. Bonner, and Christophe E. Redon. 2013. "Evaluation of the Gamma-H2AX Assay for Radiation Biodosimetry in a Swine Model" International Journal of Molecular Sciences 14, no. 7: 14119-14135. https://doi.org/10.3390/ijms140714119