Changes in the Number of Double-Strand DNA Breaks in Chinese Hamster V79 Cells Exposed to γ-Radiation with Different Dose Rates


{kind=link}
{kind=link}
Abstract
:1. Introduction
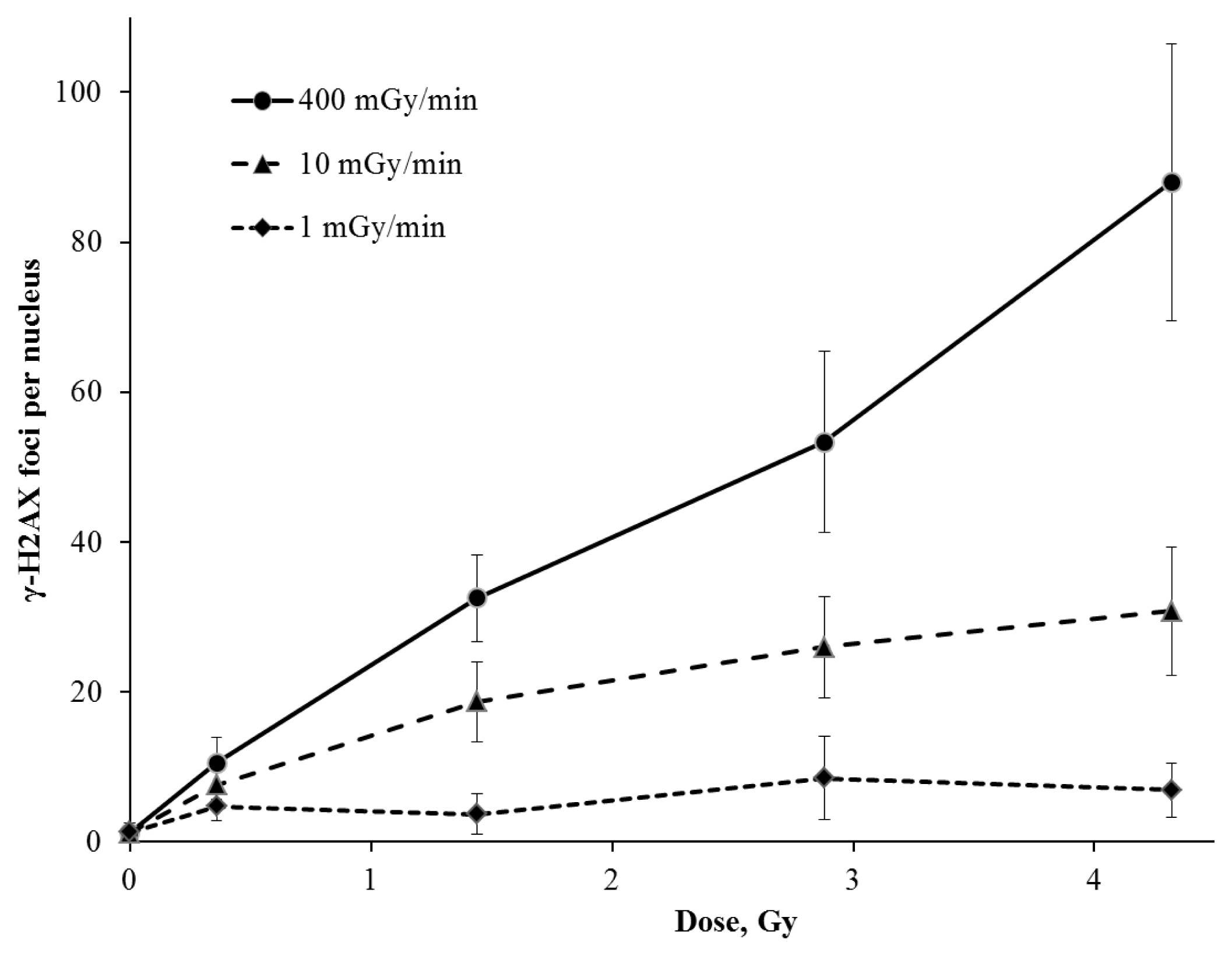
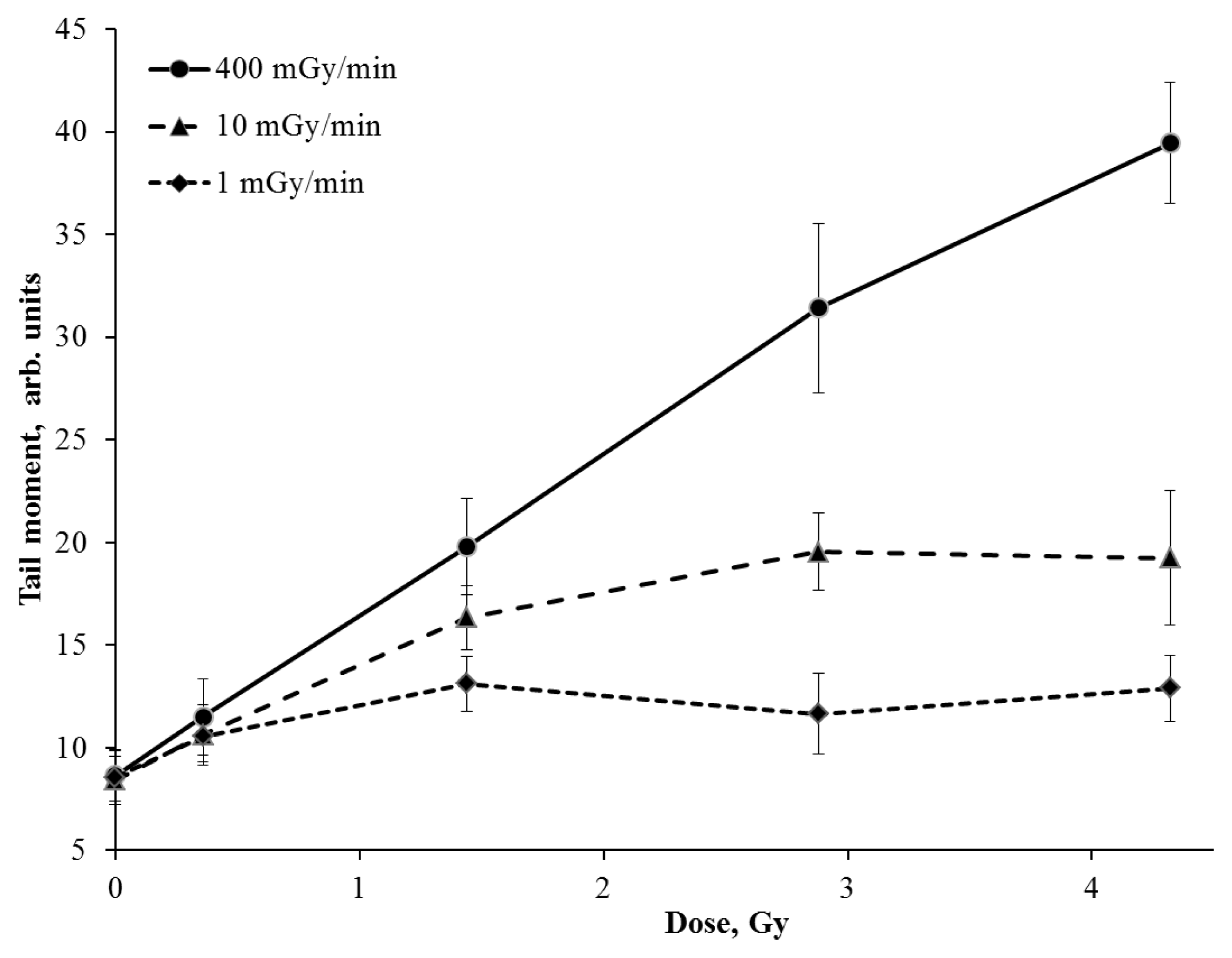
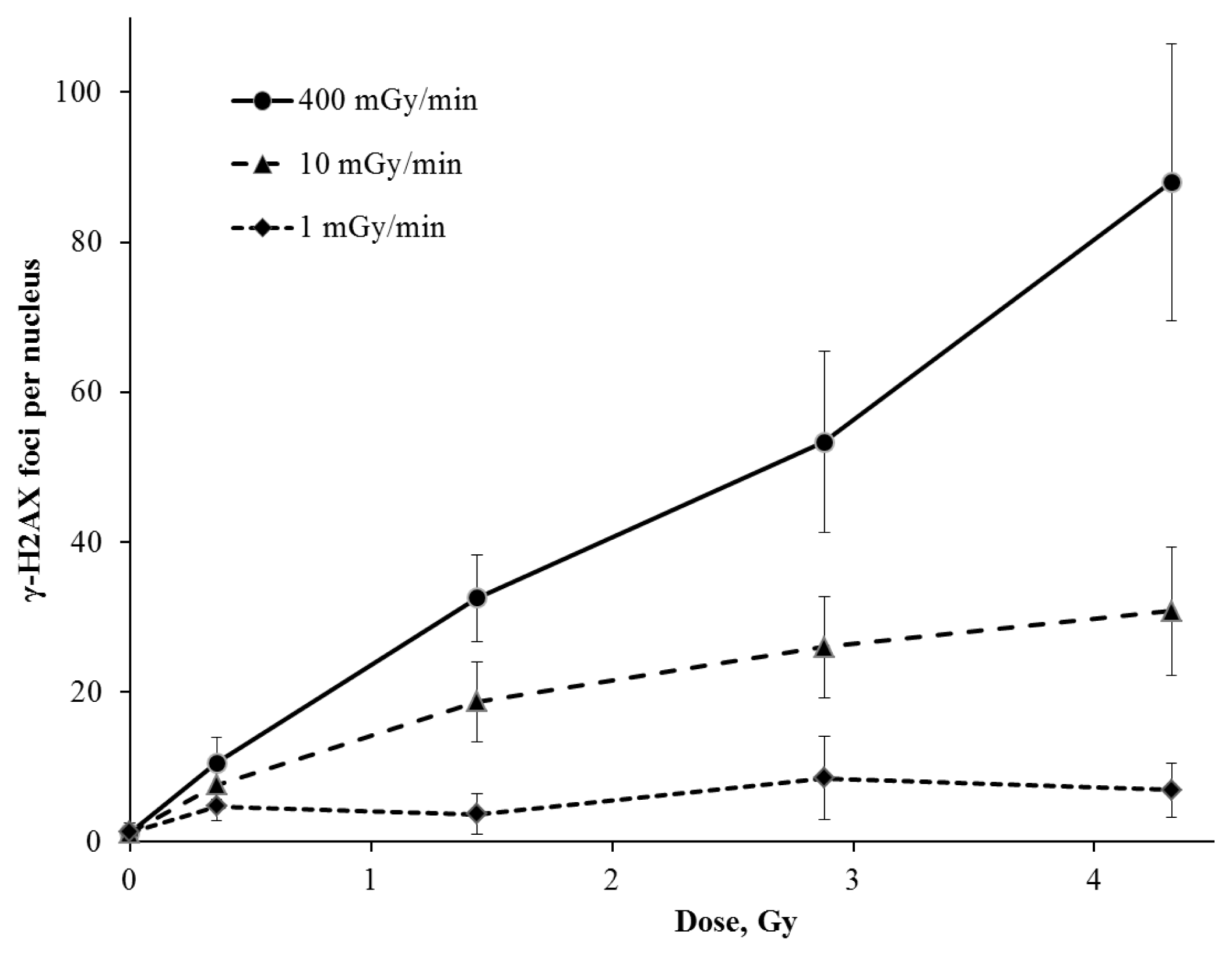
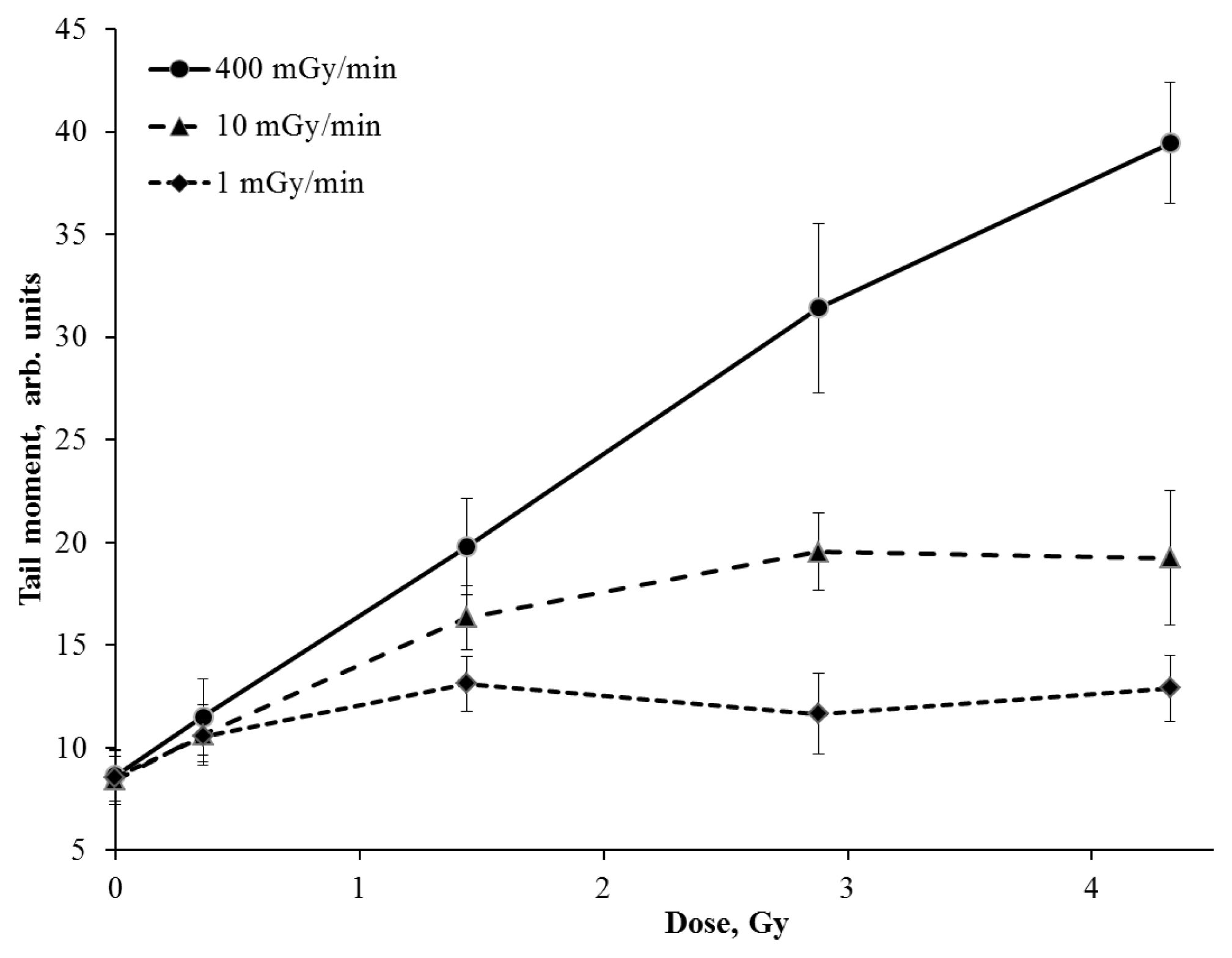
2. Results and Discussion
3. Experimental Section
3.1. Cell Line and Culture Technique
3.2. Cell Irradiation
3.3. Immunocytochemistry for γ-H2AX
3.4. DNA Comet Assay
3.5. Statistics
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Jeggo, P.A.; Lobrich, M. Contribution of DNA repair and cell cycle checkpoint arrest to the maintenance of genomic stability. DNA Repair (Amst. ) 2006, 5, 1192–1198. [Google Scholar]
- Sutherland, B.M.; Bennett, P.V.; Sidorkina, O.; Laval, J. Clustered damages and total lesions induced in DNA by ionizing radiation: Oxidized bases and strand breaks. Biochemistry 2000, 39, 8026–8031. [Google Scholar]
- Goodarzi, A.A.; Jeggo, P.; Lobrich, M. The influence of heterochromatin on DNA double strand break repair: Getting the strong, silent type to relax. DNA Repair (Amst. ) 2010, 9, 1273–1282. [Google Scholar]
- Halazonetis, T.D.; Gorgoulis, V.G.; Bartek, J. An oncogene-induced DNA damage model for cancer development. Science 2008, 319, 1352–1355. [Google Scholar]
- Mladenov, E.; Magin, S.; Soni, A.; Iliakis, G. DNA double-strand break repair as determinant of cellular radiosensitivity to killing and target in radiation therapy. Front. Oncol 2013, 3, 113. [Google Scholar]
- Grudzenski, S.; Raths, A.; Conrad, S.; Rube, C.E.; Lobrich, M. Inducible response required for repair of low-dose radiation damage in human fibroblasts. Proc. Natl. Acad. Sci. USA 2010, 107, 14205–14210. [Google Scholar]
- Mosconi, M.; Giesen, U.; Langner, F.; Mielke, C.; Dalla Rosa, I.; Dirks, W.G. 53BP1 and MDC1 foci formation in HT-1080 cells for low- and high-LET microbeam irradiations. Radiat. Environ. Biophys 2011, 50, 345–352. [Google Scholar]
- Mladenov, E.; Iliakis, G. Induction and repair of DNA double strand breaks: The increasing spectrum of non-homologous end joining pathways. Mutat. Res 2011, 711, 61–72. [Google Scholar]
- Alloni, D.; Campa, A.; Friedland, W.; Mariotti, L.; Ottolenghi, A. Track structure, radiation quality and initial radiobiological events: Considerations based on the PARTRAC code experience. Int. J. Radiat. Biol 2012, 88, 77–86. [Google Scholar]
- Lobrich, M.; Shibata, A.; Beucher, A.; Fisher, A.; Ensminger, M.; Goodarzi, A.A.; Barton, O.; Jeggo, P.A. Gammah2ax foci analysis for monitoring DNA double-strand break repair: Strengths, limitations and optimization. Cell Cycle 2010, 9, 662–669. [Google Scholar]
- Paull, T.T.; Rogakou, E.P.; Yamazaki, V.; Kirchgessner, C.U.; Gellert, M.; Bonner, W.M. A critical role for histone H2AX in recruitment of repair factors to nuclear foci after DNA damage. Curr. Biol 2000, 10, 886–895. [Google Scholar]
- Sharma, A.; Singh, K.; Almasan, A. Histone H2AX phosphorylation: A marker for DNA damage. Methods Mol. Biol 2012, 920, 613–626. [Google Scholar]
- Wojewodzka, M.; Buraczewska, I.; Kruszewski, M. A modified neutral comet assay: Elimination of lysis at high temperature and validation of the assay with anti-single-stranded DNA antibody. Mutat. Res 2002, 518, 9–20. [Google Scholar]
- Asaithamby, A.; Chen, D.J. Cellular responses to DNA double-strand breaks after low-dose gamma-irradiation. Nucleic Acids Res 2009, 37, 3912–3923. [Google Scholar]
- Goodarzi, A.A.; Jeggo, P.A. The repair and signaling responses to DNA double-strand breaks. Adv. Genet 2013, 82, 1–45. [Google Scholar]
- Mao, Z.; Bozzella, M.; Seluanov, A.; Gorbunova, V. DNA repair by nonhomologous end joining and homologous recombination during cell cycle in human cells. Cell Cycle 2008, 7, 2902–2906. [Google Scholar]
- Taleei, R.; Nikjoo, H. The Non-homologous end-joining (NHEJ) pathway for the repair of DNA double-strand breaks: I. A mathematical model. Radiat. Res 2013, 179, 530–539. [Google Scholar]
- Ishizaki, K.; Hayashi, Y.; Nakamura, H.; Yasui, Y.; Komatsu, K.; Tachibana, A. No induction of p53 phosphorylation and few focus formation of phosphorylated H2AX suggest efficient repair of DNA damage during chronic low-dose-rate irradiation in human cells. J. Radiat. Res 2004, 45, 521–525. [Google Scholar]
- Collis, S.J.; Schwaninger, J.M.; Ntambi, A.J.; Keller, T.W.; Nelson, W.G.; Dillehay, L.E.; Deweese, T.L. Evasion of early cellular response mechanisms following low level radiation-induced DNA damage. J. Biol. Chem 2004, 279, 49624–49632. [Google Scholar]
- Sugihara, T.; Murano, H.; Tanaka, K. Increased gamma-H2A.X intensity in response to chronic medium-dose-rate gamma-ray irradiation. PLoS One 2012, 7, e45320. [Google Scholar]
- Nakamura, H.; Fukami, H.; Hayashi, Y.; Tachibana, A.; Nakatsugawa, S.; Hamaguchi, M.; Ishizaki, K. Cytotoxic and mutagenic effects of chronic low-dose-rate irradiation on TERT-immortalized human cells. Radiat. Res 2005, 163, 283–288. [Google Scholar]
- Osipov, A.N.; Klokov, D.Y.; Elakov, A.L.; Rozanova, O.M.; Zaichkina, S.I.; Aptikaeva, G.F.; Akhmadieva, A. Comparison in vivo study of genotoxic action of high- versus very low dose-rate gamma-irradiation. Nonlinearity Biol. Toxicol. Med 2004, 2, 223–232. [Google Scholar]
- Tanaka, K.; Kohda, A.; Toyokawa, T.; Ichinohe, K.; Oghiso, Y. Chromosome aberration frequencies and chromosome instability in mice after long-term exposure to low-dose-rate gamma-irradiation. Mutat. Res 2008, 657, 19–25. [Google Scholar]
- Tanaka, K.; Kohda, A.; Satoh, K.; Toyokawa, T.; Ichinohe, K.; Ohtaki, M.; Oghiso, Y. Dose-rate effectiveness for unstable-type chromosome aberrations detected in mice after continuous irradiation with low-dose-rate gamma rays. Radiat. Res 2009, 171, 290–301. [Google Scholar]
- Vilenchik, M.M.; Knudson, A.G. Endogenous DNA double-strand breaks: Production, fidelity of repair, and induction of cancer. Proc. Natl. Acad. Sci. USA 2003, 100, 12871–12876. [Google Scholar]
- Banath, J.P.; Klokov, D.; MacPhail, S.H.; Banuelos, C.A.; Olive, P.L. Residual gammaH2AX foci as an indication of lethal DNA lesions. BMC Cancer 2010, 10, 4. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kotenko, K.V.; Bushmanov, A.Y.; Ozerov, I.V.; Guryev, D.V.; Anchishkina, N.A.; Smetanina, N.M.; Arkhangelskaya, E.Y.; Vorobyeva, N.Y.; Osipov, A.N. Changes in the Number of Double-Strand DNA Breaks in Chinese Hamster V79 Cells Exposed to γ-Radiation with Different Dose Rates. Int. J. Mol. Sci. 2013, 14, 13719-13726. https://doi.org/10.3390/ijms140713719
Kotenko KV, Bushmanov AY, Ozerov IV, Guryev DV, Anchishkina NA, Smetanina NM, Arkhangelskaya EY, Vorobyeva NY, Osipov AN. Changes in the Number of Double-Strand DNA Breaks in Chinese Hamster V79 Cells Exposed to γ-Radiation with Different Dose Rates. International Journal of Molecular Sciences. 2013; 14(7):13719-13726. https://doi.org/10.3390/ijms140713719
Chicago/Turabian StyleKotenko, Konstantin V., Andrey Y. Bushmanov, Ivan V. Ozerov, Denis V. Guryev, Natalya A. Anchishkina, Nadezhda M. Smetanina, Ekaterina Y. Arkhangelskaya, Natalya Y. Vorobyeva, and Andreyan N. Osipov. 2013. "Changes in the Number of Double-Strand DNA Breaks in Chinese Hamster V79 Cells Exposed to γ-Radiation with Different Dose Rates" International Journal of Molecular Sciences 14, no. 7: 13719-13726. https://doi.org/10.3390/ijms140713719