Long Non-Coding RNA in Cancer


Department of Molecular Genetics, Institute of Pathology, University of Ljubljana, SI-1000 Ljubljana, Slovenia
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2013, 14(3), 4655-4669; https://doi.org/10.3390/ijms14034655
Submission received: 1 November 2012
/
Revised: 3 January 2013
/
Accepted: 31 January 2013
/
Published: 26 February 2013
(This article belongs to the Special Issue Regulation by non-coding RNAs 2013)
Abstract
:Long non-coding RNAs (lncRNAs) are pervasively transcribed in the genome and are emerging as new players in tumorigenesis due to their various functions in transcriptional, posttranscriptional and epigenetic mechanisms of gene regulation. LncRNAs are deregulated in a number of cancers, demonstrating both oncogenic and tumor suppressive roles, thus suggesting their aberrant expression may be a substantial contributor in cancer development. In this review, we will summarize their emerging role in human cancer and discuss their perspectives in diagnostics as potential biomarkers.

1. Introduction
The central dogma of molecular biology postulates gene-coding through storage of genetic information and proteins as the main molecules of cellular functions, while RNA has the role of an intermediary between DNA sequence and encoded protein. The findings of the human genome project thus came as a surprise, since only 1.5% of the human genome encodes protein-coding genes [1–5]. Development of new techniques revolutionized the molecular world with evidence that at least 90% of the human genome is actively transcribed [6,7]. The human transcriptome has shown more complexity than previously assumed since the protein-coding transcripts are being a minority, compared to a more complex group of non-coding RNAs (ncRNAs), such as microRNAs (miRNAs), long non-coding RNAs (lncRNAs), small nucleolar RNAs (snoRNAs), small interfering RNAS (siRNAs), small nuclear (snRNAs), and piwi-interacting RNAs (piRNAs) [8–15]. Although initially thought to be transcriptional noise, ncRNA may play a crucial role in cellular development, physiology and pathologies.
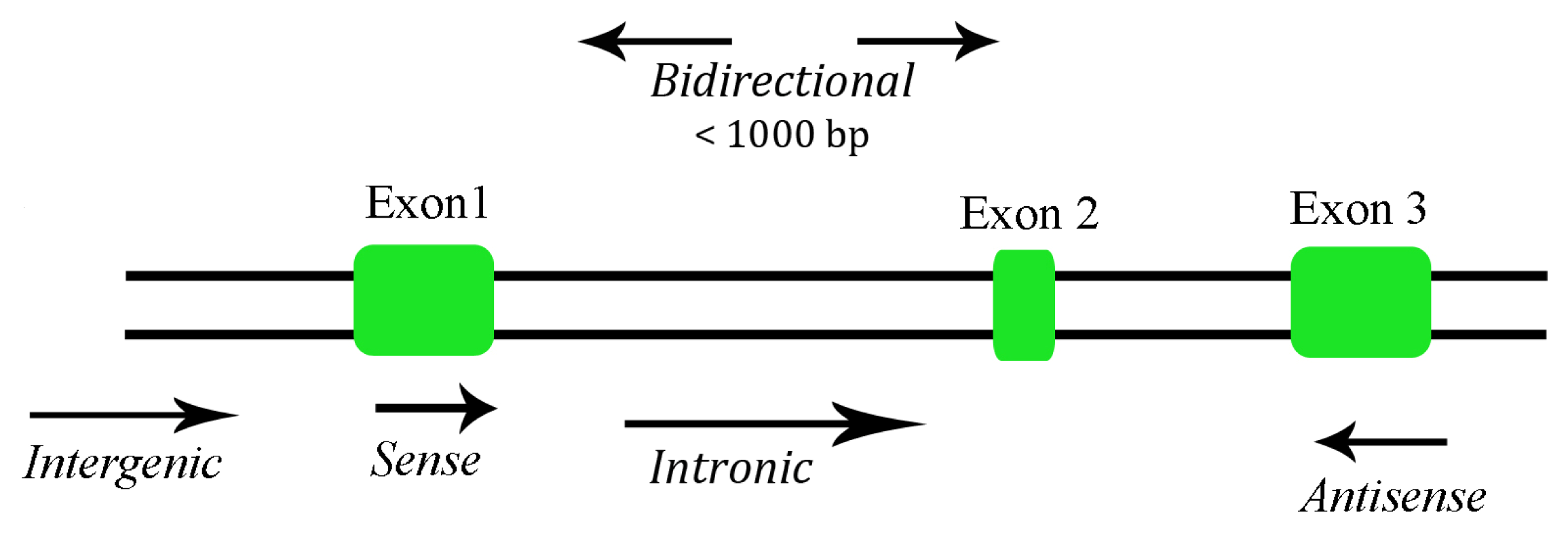
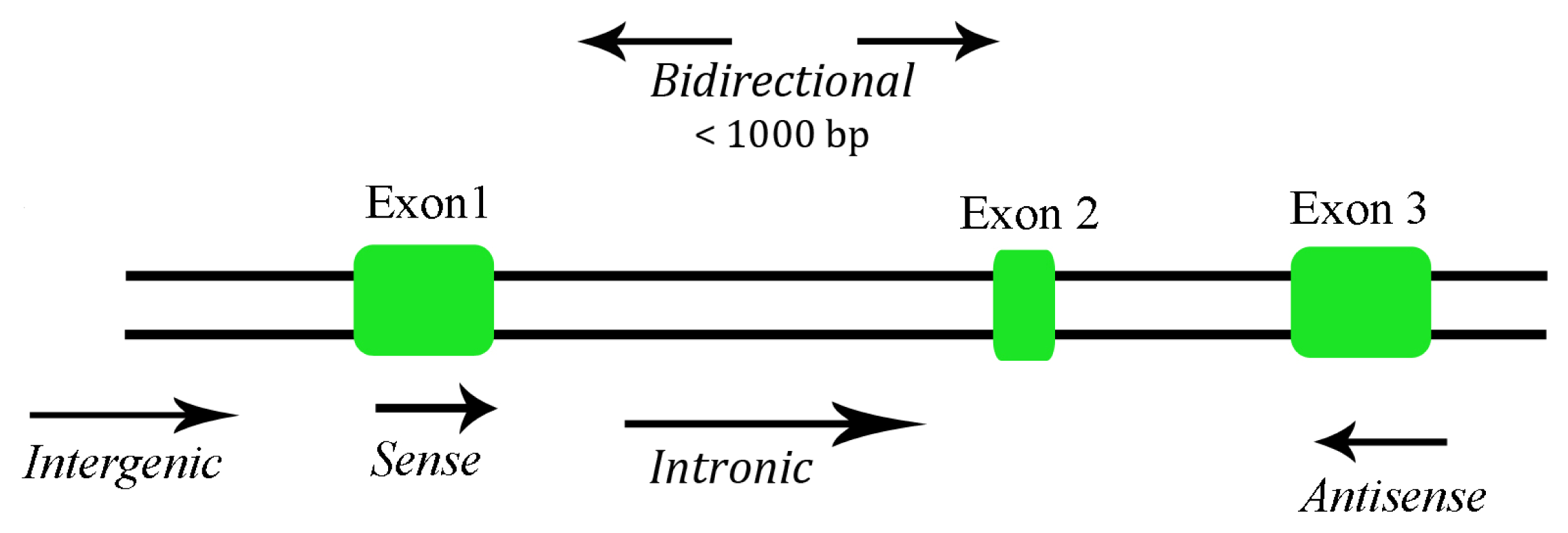
Depending on their size, ncRNAs are divided into two major groups. Transcripts shorter than 200 nucleotides are referred to as small ncRNAs, which include miRNA, siRNA, piRNA, etc. The other group is composed of lncRNA, where the transcripts lack a significant open reading frame, and have length of 200 nt up to 100 kilobases. A lncRNA can be placed into one or more of five broad categories: (1) sense, or (2) antisense, when overlapping one or more exons of another transcript on the same, or opposite, strand, respectively; (3) bidirectional, when the sequence is located on the opposite strand from a neighboring coding transcript whose transcription is initiated less than 1000 base pairs away, (4) intronic, when it is derived wholly from within an intron of a second transcript, or (5) intergenic, when it lies within the genomic interval between two genes [16] (Figure 1). There are some lncRNAs that are transcribed by RNA polymerase III while the majority of lncRNAs are transcribed by RNA polymerase II, spliced and polyadenylated [17]. Most of the lncRNAs are located in the cytoplasm, although there are some found in both cytoplasm and nucleus [18].
2. Long Non-Coding RNA Functions
LncRNAs are involved in almost every step of a life cycle of genes and regulate diverse functions. Several lncRNAs can regulate gene expression at various levels, including chromatin modification, transcription, and posttranscriptional processing [19].
So far, their role was extensively studied in epigenetic regulation, such as imprinting. Diploid organisms carry two alleles of each of the parents’ autosomal genes. In most cases, both of the alleles are expressed equally, except when a subset of genes shows imprinting in which expression is restricted by epigenetic mechanism to either maternal or paternal allele [17]. X-inactivation (XCI) is a process that equalizes gene expression between males and females by inactivating one X in female cells [17]. Some lncRNAs participate in global cellular behavior by controlling apoptosis, cell death and cell growth [15,20]. LncRNA can also mediate epigenetic modification by recruiting chromatin remodeling complex to specific chromatin loci, e.g., HOTAIR by polycomb repression complex 2 (PCR2) and/or lysine-specific demethylase 1 (LSD1), CCND1 by protein termed translocated in liposarcoma (TLS), and ANRIL by polycomb repression complex 1 and 2 (PCR1 and PCR2) [5,21–25]. The mode of action of some lncRNAs is interaction with their intracellular steroid receptors. Other lncRNAs function by regulating transcription through a variety of mechanisms that include interacting with RNA-binding proteins, acting as a coactivator of transcription factors, or repressing a major promoter of their target gene [22]. In addition to chromatin modification and transcriptional regulation, lncRNAs can regulate gene expression at the posttranscriptional level.
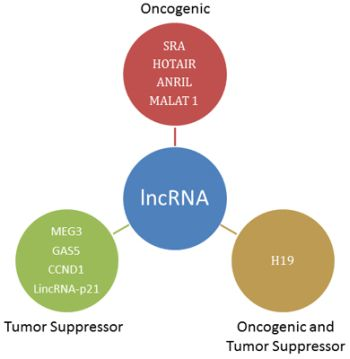
3. Oncogenic lncRNA
SRA—Steroid Receptor RNA Activator is a coactivator for steroid receptors and acts as an ncRNA found in the nucleus and cytoplasm. SRA regulates gene expression mediated by steroid receptors through complexing with proteins also containing steroid receptor coactivator 1 (SRC-1) [26]. The SRA1 gene can also encode a protein that acts as a coactivator and corepressor [27]. SRA levels have been found to be upregulated in breast tumors where it is assumed that increased SRA levels change the steroid receptors’ actions, contributing to breast tumorigenesis. While the expression of SRA in normal tissues is low, it is highly up-regulated in various tumors of the human breast, uterus and ovary. This evidence supports that SRA is a potential biomarker of steroid-dependent tumors [26].
HOTAIR—HOX Antisense Intergenic RNA with a length of 2.2 kb was found in the HOXC locus and is transcribed in antisense manner [28]. It is the first lncRNA discovered to be involved in tumorigenesis. In breast cancer, both primary and metastatic, the expression is up regulated; in the latter case up to 2000-fold increase was shown [23]. The high expression level of HOTAIR in primary breast cancer is also correlated to metastasis, and poor survival rate [23]. The level of HOTAIR expression is higher in patients with lymph node metastasis in hepatocellular cancer [29].
Polycomb group proteins mediate repression of transcription of thousands of genes that control differentiation pathways during development, and have roles in stem cell pluripotency and human cancer [23,30–34]. The target of PRC2 is the HOXD locus on chromosome 2 where the PRC2 in association with HOTAIR causes the transcriptional silencing of several metastasis suppressor genes resulting in breast epithelial cells having the expression of embryonic fibroblast. Alternating the level of HOTAIR results in enhanced PRC2 repressive activity [23]. HOTAIR acts as a molecular scaffold having two known chromatin modification complexes. The 5′ region of lncRNA binds to the PRC2 complex responsible for H3K27 methylation and the 3′ region binds to LSD1, which mediates enzymatic demethylation of H3K4 [24,30,35]. This result suggests the possible function of HOTAIR as a scaffold binding to selected histone modification enzymes and therefore causing histone modification on target genes [30]. Although the precise mechanism is still not known, it is clear that HOTAIR remodels chromatin to promote cancer invasiveness.
HOTAIR as an epigenetic regulator in gene expression is deregulated in different cancers [23,36–38]. In hepatocellular carcinoma (HCC) and HCC patients with liver transplantation, the levels of HOTAIR compared with normal liver tissue are elevated. Expression levels of HOTAIR can also be used as an independent prognostic marker for HCC recurrence and lower survival rate [31]. HOTAIR can be a potential biomarker for the existence of lymph node metastasis in HCC [29].
ANRIL—Antisense ncRNA in the INK4 locus
Many transcripts coding for proteins have anti-sense partners, whose perturbation can alter the expression of the sense transcripts [39]. Some of these genes are tumor suppressors, which can be epigenetically silenced by antisense ncRNA [40].
ANRIL activates two polycomb repressor complexes, PRC1 and PRC2 [21,25], resulting in chromatin reorganization which silences the INK4b-ARF-INK4a locus encoding tumor suppressors p15INK4b, p14ARF, p16INK4a, which are active in cell cycle inhibition, senescence and stress-induced apoptosis. Overexpression of ANRIL in prostate cancer has shown silencing of INK4b-ARF-INK4a and p15/CDKN2B by heterochromatin reformation [25,41]. The repression is mediated by direct binding to combox 7 (CBX 7) and SUZ12, members of PRC1 and PRC2, respectively [21,25].
MALAT 1—Metastasis-Associated Lung Adenocarcinoma Transcript 1
This lncRNA was first associated with high metastatic potential and poor patient prognosis during a comparative screen of non-small cell lung cancer patients with or without metastatic tumors [42]. MALAT1 is widely expressed in normal human tissues [42,43] and is found to be upregulated in a variety of human cancers of the breast, prostate, colon, liver and uterus [44–47]. The MALAT1 locus at 11q13.1 has been identified to harbor chromosomal translocation break points associated with cancer [48–50]. MALAT1 is localized in nuclear speckles and widely expressed in normal tissues [42,43], but was found to be upregulated in hepatocellular carcinoma, breast, pancreas, osteosarcoma, colon and prostate cancers [44–47,51]. It has been shown that increased expression of MALAT1 can be used as a prognostic marker for HCC patients following liver transplantation [52].
A number of studies have implicated MALAT1 in the regulation of cell mobility, due to its high levels of expression in cancers. For example, RNA interference-mediated silencing of MALAT1 reduced the in vitro migration of lung adenocarcinoma cells by influencing the expression of motility-related genes [53]. Recent studies on knockout MALAT1 mice have not displayed any cellular phenotype. Future studies will be needed where mice will be exposed to different stresses, such as induction of cancer, which will potentially unveil its function. It is known that MALAT1 as well as HOTAIR play vital roles in human cells but it is possible that they have no significant role in living animals under normal physiological conditions [54–56].
4. Oncogenic and Tumor Suppressor lncRNA
H19 is expressed from the maternal allele and has a pivotal role in genomic imprinting during cell growth and development [57]. The locus contains H19 and insulin-like growing factor 2 (IGF2), which are imprinted. This leads to differential expression of both genes, H19 from maternal and IGF2 from paternal allele [57,58]. The loss of imprinting results in misexpression of H19 and was observed in many tumors including hepatocellular and bladder cancer [59,60]. This lncRNA has been linked to oncogenic and tumor suppressor properties [57]. cMYC induces the expression of H19 in different cell types where H19 potentiates tumorigenesis [58]. In addition c-MYC also down-regulates expression of IGF2 imprinted gene. H19 transcripts are precursors for miR-675 which functionally down-regulates the tumor suppressor gene for retinoblastoma in human colorectal cancer [61]. Data support H19 deregulation causing either oncogenic or tumor suppressor properties, although the exact mechanism is still elusive.
5. Tumor Suppressor lncRNA
MEG3—Maternally Expressed Gene 3
LncRNA MEG3 is a transcript of the maternally imprinted gene. In normal pituitary cells MEG3 is expressed, the loss of expression is observed in pituitary adenomas and the majority of meningiomas and meningioma cell lines [62,63]. MEG3 activates regulation of tumor suppressor protein p53. Normally, p53 protein levels are extremely low due to its rapid degradation via the ubiquitin-proteasome pathway. The ubiquitination of p53 is mainly mediated by MDM2, an E3 ubiquitin ligase. MEG3 down-regulates MDM2 expression, which suggests that MDM2 down-regulation is one of the mechanisms whereby MEG3 activates p53 [64]. MEG3 significantly increases p53 protein level and stimulates p53-dependent transcription [65]. MEG3 enhances p53 binding to target promoters such as GDF15 but not p21 and is also able to inhibit cell proliferation in the absence p53, suggesting that MEG3 is a p53 dependent and independent tumor suppressor [62–65].
GAS5—Growth Arrest-Specific 5 is widely expressed in embryonic and adult tissues. Expression is almost undetectable in growing leukemia cells and abundant in saturation density-arrested cells [66,67]. GAS5 functions as a starvation or growth arrest-linked riborepressor for the glucocorticoid receptors by binding to their DNA binding domain inhibiting the association of these receptors with their DNA recognition sequence. This suppresses the induction of several responsive genes including the gene encoding cellular inhibitor of apoptosis 2 (cIAP2), reducing cell metabolism and synthesizes cells to apoptosis [4,67]. GAS5 can induce apoptosis directly or indirectly in the prostate and breast cancer cell lines, where it was shown that GAS5 has a significantly lower expression in breast cancers compared to normal breast epithelial tissues [68].
CCND1/Cyclin D1 is a heterogenous lncRNA transcribed from the promoter region of the Cyclin D1 gene. Cyclin D1 is a cell cycle regulator that is frequently mutated, amplified and over expressed in a variety of cancers [69]. LncRNA recruits the RNA-binding proteinTLS, which is a key transcriptional regulatory sensor of DNA damage signals. Upon binding TLS undergoes allosteric modification, modulating activities of CREB-binding protein (CBP) and p300, resulting in inhibition of the cyclin D1 gene expression [22].
LincRNA-p21 expression is directly induced by the p53 signaling pathway. It is required for global repression of genes that interfere with p53 function to regulate cellular apoptosis. Lincrna-p21 mediated gene repression occurs through physical interaction with RNA-binding protein hnRNP K leading to the promoters of genes being repressed in a p53 dependent manner [70].
6. Diagnostic Benefits of lncRNA
So far, the majority of cancer biomarkers are protein-coding genes, their transcripts or the proteins. The non-coding regions are evolving as a biomarker hotspots only recently. By the advent of high-throughput sequencing, we are now able to identify deregulated expression of transcriptome at much higher resolution, what allow us to decipher smaller changes in the expression level. In the case of lncRNAs, where their main function is regulation of other genes expression, the importance of lncRNAs maintained expression is evident. Since cancer is a complicated disease, which involves many factors, molecular biomarkers are valuable diagnostic and prognostic tools that could ease the disease management. Compared to protein-coding RNAs, using lncRNA as markers is of advantage since their own expression is a better indicator of the tumor status. Many lncRNAs are now connected to cancer due to new technologies and are emerging into the field of molecular biology as new regulatory players. Several lncRNA were found to be deregulated in a wide variety of cancers (Table 1).
In breast cancer research higher expressions of SRA and SRAP, compared to normal tissue were observed. Possibly SRAP expression contributes to higher survival for patient undergoing Tamoxifen treatment [90].
The expression of MALAT 1 is elevated in osteosarcoma patients with poor response to chemotherapy, which suggests that this transcript plays a crucial role in the pathology of tumors [53].
Additionally MALAT 1 serves as an independent prognostic marker for patient survival in early stage non-small cell lung cancer [42].
In hepatocellular carcinoma, (HCC) definitive diagnosis of lymph node metastasis is difficult without histological evidence. It has been demonstrated that a significant correlation between HOTAIR gene expression and lymph node metastasis exists, suggesting that measuring HOTAIR lncRNA is a potential biomarker for predicting lymph node metastasis [29]. Upregulation of HOTAIR is closely associated with gastrointestinal stromal tumor (GIST) aggressiveness and metastasis and it can be used as a potential biomarker [38,91].
MALAT1 is a powerful biomarker for HCC recurrence prediction following liver transplantation. Moreover, silencing MALAT1 activity in HCC would be a potential anticancer therapy to prevent tumor recurrence after orthotopic liver transplantation [52].
SPRY4-IT1 expression is substantially increased in patient melanoma cell samples compared to melanocytes. The elevated expression of SPRY4-IT1 in melanoma cells, its accumulation in the cell cytoplasm, and its effects on cell dynamics suggest that the misexpression of SPRY4-IT1 may have an important role in melanoma development, and could be an early biomarker and a key regulator for melanoma pathogenesis in humans [85].
The novel potential biomarkers can be discovered through certain types of highly expressed cancer-associated lncRNAs [92]. Therapeutic benefit can be obtained through pathways mediating transcriptional gene silencing, especially those of tumor suppressors and oncogenes [93]. For patients’ comfort, biomarkers should be detected in samples obtained in a non-invasive way. Desirable samples are body fluids, such as serum or urine, where circulating nuclear acids (CNAs), both DNA and RNA species, are found. CNAs are found in plasma, cell-free serum, sputum and urine [29,94–97].
PRNCR1 (prostate cancer non-coding RNA1) expression was upregulated in some of the prostate cancer cells as well as precursor lesion prostatic intraepithelial neoplasia and considered as a tumor marker [75].
Suggestions that lncRNA can be used as biomarkers and/or drug targets have arisen from numerous studies observing the expression patterns of tumor tissues comparing to normal ones [14]. The possible therapies arising from this knowledge would be beneficial in cases where protein target drugs have not been effective. A recent study has shown that reduced expression of ncRAN enhanced the chemotherapeutic drug in vitro[98]. This opens another possibility of cancer treatment, where a combination of drugs would have much higher effect.
Often lncRNAs exhibit tissue specific patterns that distinguish them from miRNAs and protein-coding mRNAs that are expressed from multiple tissue types. Their specificity makes them precise biomarkers for cancer diagnostics [99]. PCA3 is a prostate-specific lncRNA overexpressed in prostate cancer. Although its functions are not understood, it was still utilized as a biomarker in a clinical test. Expression of the PCA3 transcript is determined from prostate cells in urine samples of patients [100,101]. Another lncRNA detected in body fluids is HULC, expression of which is disrupted in hepatocellular carcinomas and can be monitored in patients’ blood sera [102].
To understand the biology of cancer it will be essential to identify, annotate lncRNAs and study their expression profiles in human tissues and diseases [103,104]. With this, the potential of lncRNAs on biology and medicine will be revealed. Long non-coding RNAs have recently arisen as new discoveries in the field of molecular biology. Since only a few individual lncRNAs have been functionally studied, still a lot of questions remain to be addressed [4]. At the moment the full potential of cancer therapy is not yet developed. The future of it lies in specific targeting of cancer cells and specific delivery of the drugs. LncRNAs are a possible resource for developing diagnostics and therapies, although a better understanding of their function and precise mechanism through which they function are needed first [4]. Another possibility for cancer treatment lies in combination of drugs, where one would change the expression of lncRNA in a way for chemotherapeutic drug to have a higher effect. Since probably the lncRNA function through their secondary structure special molecules could be developed to disrupt their secondary structure or bind to them to form complexes through which an inactivation of lncRNA would occur. These molecules should be highly specific in order not to disrupt other molecules and mechanisms. To discover the right molecules more studies of the complex mechanisms involving lncRNA are needed.
7. Conclusions
RNA used to be just a messenger between coding genes and proteins encoded by them. However, “transcriptional noise” is turning out to be a very important part of regulation processes. With the discovery of LncRNA and their functions, the new world of molecular biology is emerging. There is much research still on the way towards a deeper understanding of regulation processes in which lncRNA is one of the important players. LncRNA deregulation in human disease is unveiling the complexity of cellular processes. Studying the mechanisms of lncRNA involvement in oncogenic and tumor suppressive pathways will lead to new cancer diagnostic markers and will pave the way to novel therapeutic targets.
Acknowledgments
This work was supported by program P3-0054 of the Slovenian Research Agency.
Conflict of Interest
The authors declare no conflict of interest.
References
- Stein, L.D. Human genome: End of the beginning. Nature 2004, 431, 915–916. [Google Scholar]
- Ponting, C.P.; Belgard, T.G. Transcribed dark matter: meaning or myth? Hum. Mol. Genet 2010, 19, R162–R168. [Google Scholar]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 49, 860–921. [Google Scholar]
- Gutschner, T.; Diederichs, S. The Hallmarks of Cancer: A long non-coding RNA point of view. RNA Biol 2012, 9, 703–719. [Google Scholar]
- Nie, L.; Wu, H.-J.; Hsu, J.-M.; Chang, S.-S.; LaBaff, A.; Li, C.-W.; Wang, Y.; Hsu, J.L.; Hung, M.-C. Long non-coding RNAs: Versatile master regulators of gene expression and crucial players in cancer. Am. J. Transl. Res 2012, 4, 127–150. [Google Scholar]
- Birney, E.; Stamatoyannopoulos, J.A.; Dutta, A.; Guigó, R.; Gingeras, T.R.; Margulies, E.H.; Weng, Z.; Snyder, M.; Dermitzakis, E.T.; Thurman, R.E.; et al. Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project. Bioessays 2007, 32, 599–608. [Google Scholar]
- Costa, F.F. Non-coding RNAs: Meet thy masters. Bioessays 2010, 32, 599–608. [Google Scholar]
- Kapranov, P.; Willingham, A.T.; Gingeras, T.R. Genome-wide transcription and the implications for genomic organization. Nat. Rev. Genet 2007, 8, 413–423. [Google Scholar]
- Frith, M.C.; Pheasant, M.; Mattick, J.S. The amazing complexity of the human transcriptome. Eur. J. Hum. Genet 2005, 13, 894–897. [Google Scholar]
- Khachane, A.N.; Harrison, P.M. Mining mammalian transcript data for functional long non-coding RNAs. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Mattick, J.S.; Makunin, I.V. Non-coding RNA. Hum. Mol. Genet 2006, 15, R17–R29. [Google Scholar]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar]
- Washietl, S.; Hofacker, I.L.; Lukasser, M.; Huttenhofer, A.; Stadler, P.F. Mapping of conserved RNA secondary structures predicts thousands of functional noncoding RNAs in the human genome. Nat. Biotechnol 2005, 23, 1383–1390. [Google Scholar]
- Taft, R.J.; Pang, K.C.; Mercer, T.R.; Dinger, M.; Mattick, J.S. Non-coding RNAs: Regulators of disease. J. Pathol 2010, 220, 126–139. [Google Scholar]
- Sana, J.; Faltejskova, P.; Svoboda, M.; Slaby, O. Novel classes of non-coding RNAs and cancer. J. Transl. Med. 2012, 10. [Google Scholar] [CrossRef]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar]
- Banfai, B.; Jia, H.; Khatun, J.; Wood, E.; Risk, B.; Gundling, W.E.; Kundaje, A.; Gunawardena, H.P.; Yu, Y.; Xie, L.; et al. Long noncoding RNAs are rarely translated in two human cell lines. Genome Res 2012, 22, 1646–1657. [Google Scholar]
- Wilusz, J.E.; Sunwoo, H.; Spector, D.L. Long noncoding RNAs: Functional surprises from the RNA world. Genes Dev 2009, 23, 1494–1504. [Google Scholar]
- Wapinski, O.; Chang, H.Y. Long noncoding RNAs and human disease. Trends. Cell Biol 2011, 21, 354–361. [Google Scholar]
- Kotake, Y.; Nakagawa, T.; Kitagawa, K.; Suzuki, S.; Liu, N.; Kitagawa, M.; Xiong, Y. Long non-coding RNA ANRIL is required for the PRC2 recruitment to and silencing of p15(INK4B) tumor suppressor gene. Oncogene 2011, 30, 1956–1962. [Google Scholar]
- Wang, X.; Arai, S.; Song, X.; Reichart, D.; Du, K.; Pascual, G.; Tempst, P.; Rosenfeld, M.G.; Glass, C.K.; Kurokawa, R. Induced ncRNAs allosterically modify RNA-binding proteins in cis to inhibit transcription. Nature 2008, 454, 126–130. [Google Scholar]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar]
- Hayami, S.; Kelly, J.D.; Cho, H.S.; Yoshimatsu, M.; Unoki, M.; Tsunoda, T.; Field, H.I.; Neal, D.E.; Yamaue, H.; Ponder, B.A.; et al. Overexpression of LSD1 contributes to human carcinogenesis through chromatin regulation in various cancers. Int. J. Cancer 2011, 128, 574–586. [Google Scholar]
- Yap, K.L.; Li, S.; Munoz-Cabello, A.M.; Raguz, S.; Zeng, L.; Mujtaba, S.; Gil, J.; Walsh, M.J.; Zhou, M.M. Molecular interplay of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by polycomb CBX7 in transcriptional silencing of INK4a. Mol. Cell 2010, 38, 662–674. [Google Scholar]
- Lanz, R.B.; Chua, S.S.; Barron, N.; Soder, B.M.; DeMayo, F.; O’Malley, B.W. Steroid receptor RNA activator stimulates proliferation as well as apoptosis in vivo. Mol. Cell. Biol 2003, 23, 7163–7176. [Google Scholar]
- Chooniedass-Kothari, S.; Vincett, D.; Yan, Y.; Cooper, C.; Hamedani, M.K.; Myal, Y.; Leygue, E. The protein encoded by the functional steroid receptor RNA activator is a new modulator of ER alpha transcriptional activity. FEBS Lett 2010, 584, 1174–1180. [Google Scholar]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Functional Demarcation of Active and Silent Chromatin Domains in Human HOX Loci by Noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar]
- Geng, Y.J.; Xie, S.L.; Li, Q.; Ma, J.; Wang, G.Y. Large intervening non-coding RNA HOTAIR is associated with hepatocellular carcinoma progression. J. Int. Med. Res 2011, 39, 2119–2128. [Google Scholar]
- Tsai, M.C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long noncoding RNA as modular scaffold of histone modification complexes. Science 2010, 329, 689–693. [Google Scholar]
- Morey, L.; Helin, K. Polycomb group protein-mediated repression of transcription. Trends. Biochem. Sci 2010, 35, 323–332. [Google Scholar]
- Zhao, J.; Ohsumi, T.K.; Kung, J.T.; Ogawa, Y.; Grau, D.J.; Sarma, K.; Song, J.J.; Kingston, R.E.; Borowsky, M.; Lee, J.T. Genome-wide identification of polycomb-associated RNAs by RIP-seq. Mol. Cell 2010, 40, 939–953. [Google Scholar]
- Zhang, Z.; Jones, A.; Sun, C.W.; Li, C.; Chang, C.W.; Joo, H.Y.; Dai, Q.; Mysliwiec, M.R.; Wu, L.C.; Guo, Y.; et al. PRC2 complexes with JARID2, MTF2, and esPRC2p48 in ES cells to modulate ES cell pluripotency and somatic cell reprogramming. Stem Cells 2011, 29, 229–240. [Google Scholar]
- Simon, J.A.; Lange, C.A. Roles of the EZH2 histone methyltransferase in cancer epigenetics. Mutat. Res 2008, 647, 21–29. [Google Scholar]
- Sirchia, S.M.; Tabano, S.; Monti, L.; Recalcati, M.P.; Gariboldi, M.; Grati, F.R.; Porta, G.; Finelli, P.; Radice, P.; Miozzo, M. Misbehaviour of XIST RNA in breast cancer cells. PLoS One 2009, 4. [Google Scholar] [CrossRef]
- Yang, Z.; Zhou, L.; Wu, L.M.; Lai, M.C.; Xie, H.Y.; Zhang, F.; Zheng, S.S. Overexpression of long non-coding RNA HOTAIR predicts tumor recurrence in hepatocellular carcinoma patients following liver transplantation. Ann. Surg. Oncol 2011, 18, 1243–1250. [Google Scholar]
- Kogo, R.; Shimamura, T.; Mimori, K.; Kawahara, K.; Imoto, S.; Sudo, T.; Tanaka, F.; Shibata, K.; Suzuki, A.; Komune, S.; et al. Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin modification and is associated with poor prognosis in colorectal cancers. Cancer Res 2011, 71, 6320–6326. [Google Scholar]
- Niinuma, T.; Suzuki, H.; Nojima, M.; Nosho, K.; Yamamoto, H.; Takamaru, H.; Yamamoto, E.; Maruyama, R.; Nobuoka, T.; Miyazaki, Y.; et al. Upregulation of miR-196a and HOTAIR drive malignant character in gastrointestinal stromal tumors. Cancer Res 2012, 72, 1126–1136. [Google Scholar]
- Katayama, S.; Tomaru, Y.; Kasukawa, T.; Waki, K.; Nakanishi, M.; Nakamura, M.; Nishida, H.; Yap, C.C.; Suzuki, M.; Kawai, J.; et al. Antisense transcription in the mammalian transcriptome. Science 2005, 309, 1564–1566. [Google Scholar]
- Kim, W.Y.; Sharpless, N.E. The regulation of INK4/ARF in cancer and aging. Cell 2006, 127, 265–275. [Google Scholar]
- Yu, W.; Gius, D.; Onyango, P.; Muldoon-Jacobs, K.; Karp, J.; Feinberg, A.P.; Cui, H. Epigenetic silencing of tumour suppressor gene p15 by its antisense RNA. Nature 2008, 451, 202–206. [Google Scholar]
- Ji, P.; Diederichs, S.; Wang, W.; Boing, S.; Metzger, R.; Schneider, P.M.; Tidow, N.; Brandt, B.; Buerger, H.; Bulk, E.; et al. MALAT-1, a novel noncoding RNA, and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene 2003, 22, 8031–8041. [Google Scholar]
- Hutchinson, J.N.; Ensminger, A.W.; Clemson, C.M.; Lynch, C.R.; Lawrence, J.B.; Chess, A. A screen for nuclear transcripts identifies two linked noncoding RNAs associated with SC35 splicing domains. BMC Genomics 2007, 8. [Google Scholar] [CrossRef]
- Guffanti, A.; Iacono, M.; Pelucchi, P.; Kim, N.; Solda, G.; Croft, L.J.; Taft, R.J.; Rizzi, E.; Askarian-Amiri, M.; Bonnal, R.J.; et al. A transcriptional sketch of a primary human breast cancer by 454 deep sequencing. BMC Genomics 2009, 10. [Google Scholar] [CrossRef]
- Yamada, K.; Kano, J.; Tsunoda, H.; Yoshikawa, H.; Okubo, C.; Ishiyama, T.; Noguchi, M. Phenotypic characterization of endometrial stromal sarcoma of the uterus. Cancer Sci 2006, 97, 106–112. [Google Scholar]
- Lin, R.; Maeda, S.; Liu, C.; Karin, M.; Edgington, T.S. A large noncoding RNA is a marker for murine hepatocellular carcinomas and a spectrum of human carcinomas. Oncogene 2007, 26, 851–858. [Google Scholar]
- Luo, J.H.; Ren, B.; Keryanov, S.; Tsang, G.C.; Reo, U.N.M.; Monga, S.P.; Storm, A.; Demetris, A.J.; Nalesnik, M.; Yu, Y.P.; et al. Transcriptomic and genomic analysis of human hepatocellular carcinomas and hepatoblastomas. Hepatology 2006, 44, 1012–1024. [Google Scholar]
- Davis, I.J.; Hsi, B.L.; Arroyo, J.D.; Vargas, S.O.; Yeh, Y.A.; Motyckova, G.; Valencia, P.; Perez-Atayde, A.R.; Argani, P.; Ladanyi, M.; et al. Cloning of an Alpha-TFEB fusion in renal tumors harboring the t(6;11)(p21;q13) chromosome translocation. Proc. Natl. Acad Sci USA 2003, 100, 6051–6056. [Google Scholar]
- Kuiper, R.P.; Schepens, M.; Thijssen, J.; van Asseldonk, M.; van den Berg, E.; Bridge, J.; Schuuring, E.; Schoenmakers, E.F.; van Kessel, A.G. Upregulation of the transcription factor TFEB in t(6;11)(p21;q13)-positive renal cell carcinomas due to promoter substitution. Hum. Mol. Genet 2003, 12, 1661–1669. [Google Scholar]
- Rajaram, V.; Knezevich, S.; Bove, K.E.; Perry, A.; Pfeifer, J.D. DNA sequence of the translocation breakpoints in undifferentiated embryonal sarcoma arising in mesenchymal hamartoma of the liver harboring the t(11;19)(q11;q13.4) translocation. Genes Chromosomes Cancer 2007, 46, 508–513. [Google Scholar]
- Fellenberg, J.; Bernd, L.; Delling, G.; Witte, D.; Zahlten-Hinguranage, A. Prognostic significance of drug-regulated genes in high-grade osteosarcoma. Mod. Pathol 2007, 20, 1085–1094. [Google Scholar]
- Lai, M.C.; Yang, Z.; Zhou, L.; Zhu, Q.Q.; Xie, H.Y.; Zhang, F.; Wu, L.M.; Chen, L.M.; Zheng, S.S. Long non-coding RNA MALAT-1 overexpression predicts tumor recurrence of hepatocellular carcinoma after liver transplantation. Med. Oncol 2012, 29, 1810–1816. [Google Scholar]
- Tano, K.; Mizuno, R.; Okada, T.; Rakwal, R.; Shibato, J.; Masuo, Y.; Ijiri, K.; Akimitsu, N. MALAT-1 enhances cell motility of lung adenocarcinoma cells by influencing the expression of motility-related genes. FEBS Lett 2010, 584, 4575–4580. [Google Scholar]
- Nakagawa, S.; Ip, J.Y.; Shioi, G.; Tripathi, V.; Zong, X.; Hirose, T.; Prasanth, K.V. Malat1 is not an essential component of nuclear speckles in mice. RNA 2012, 18, 1487–1499. [Google Scholar]
- Bickmore, W.A.; Schorderet, P.; Duboule, D. Structural and Functional Differences in the Long Non-Coding RNA Hotair in Mouse and Human. PLoS Genet. 2011, 7. [Google Scholar] [CrossRef]
- Eiβmann, M.; Gutschner, T.; Hämmerle, M.; Günther, S.; Caudron-Herger, M.; Groß, M.; Schirmacher, P.; Rippe, K.; Braun, T.; Zörnig, M.; Diederichs, S. Loss of the abundant nuclear non-coding RNA MALAT1 is compatible with life and development. RNA Biol 2012, 9, 1076–1087. [Google Scholar]
- Gabory, A.; Jammes, H.; Dandolo, L. The H19 locus: role of an imprinted non-coding RNA in growth and development. Bioessays 2010, 32, 473–480. [Google Scholar]
- Barsyte-Lovejoy, D.; Lau, S.K.; Boutros, P.C.; Khosravi, F.; Jurisica, I.; Andrulis, I.L.; Tsao, M.S.; Penn, L.Z. The c-Myc oncogene directly induces the H19 noncoding RNA by allele-specific binding to potentiate tumorigenesis. Cancer Res 2006, 66, 5330–5337. [Google Scholar]
- van Bakel, H.; Nislow, C.; Blencowe, B.J.; Hughes, T.R. Most “dark matter” transcripts are associated with known genes. PLoS Biol. 2010, 8. [Google Scholar] [CrossRef]
- Oosumi, T.; Belknap, W.R.; Garlick, B. Mariner transposons in humans. Nature 1995, 378, 672–672. [Google Scholar]
- Tsang, W.P.; Ng, E.K.; Ng, S.S.; Jin, H.; Yu, J.; Sung, J.J.; Kwok, T.T. Oncofetal H19-derived miR-675 regulates tumor suppressor RB in human colorectal cancer. Carcinogenesis 2010, 31, 350–358. [Google Scholar]
- Gejman, R.; Batista, D.L.; Zhong, Y.; Zhou, Y.; Zhang, X.; Swearingen, B.; Stratakis, C.A.; Hedley-Whyte, E.T.; Klibanski, A. Selective loss of MEG3 expression and intergenic differentially methylated region hypermethylation in the MEG3/DLK1 locus in human clinically nonfunctioning pituitary adenomas. J. Clin. Endocrinol. Metab 2008, 93, 4119–4125. [Google Scholar]
- Zhang, X.; Gejman, R.; Mahta, A.; Zhong, Y.; Rice, K.A.; Zhou, Y.; Cheunsuchon, P.; Louis, D.N.; Klibanski, A. Maternally expressed gene 3, an imprinted noncoding RNA gene, is associated with meningioma pathogenesis and progression. Cancer Res 2010, 70, 2350–2358. [Google Scholar]
- Zhou, Y.; Zhang, X.; Klibanski, A. MEG3 noncoding RNA: A tumor suppressor. J. Mol. Endocrinol 2012, 48, R45–R53. [Google Scholar]
- Benetatos, L.; Vartholomatos, G.; Hatzimichael, E. MEG3 imprinted gene contribution in tumorigenesis. Int. J. Cancer 2011, 129, 773–779. [Google Scholar]
- Coccia, E.M.; Cicala, C.; Charlesworth, A.; Ciccarelli, C.; Rossi, G.B.; Philipson, L.; Sorrentino, V. Regulation and expression of a growth arrest-specific gene (gas5) during growth, differentiation, and development. Mol. Cell. Biol 1992, 12, 3514–3521. [Google Scholar]
- Kino, T.; Hurt, D.E.; Ichijo, T.; Nader, N.; Chrousos, G.P. Noncoding RNA gas5 is a growth arrest- and starvation-associated repressor of the glucocorticoid receptor. Sci Signal 2010, 3, ra8. [Google Scholar]
- Mourtada-Maarabouni, M.; Pickard, M.R.; Hedge, V.L.; Farzaneh, F.; Williams, G.T. GAS5, a non-protein-coding RNA, controls apoptosis and is downregulated in breast cancer. Oncogene 2009, 28, 195–208. [Google Scholar]
- Diehl, J.A. Cycling to Cancer with Cyclin D1. Cancer Biol. Ther 2002, 1, 226–231. [Google Scholar]
- Huarte, M.; Guttman, M.; Feldser, D.; Garber, M.; Koziol, M.J.; Kenzelmann-Broz, D.; Khalil, A.M.; Zuk, O.; Amit, I.; Rabani, M.; et al. A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response. Cell 2010, 142, 409–419. [Google Scholar]
- Amaral, P.P.; Clark, M.B.; Gascoigne, D.K.; Dinger, M.E.; Mattick, J.S. lncRNAdb: A reference database for long noncoding RNAs. Nucleic Acids Res 2011, 39, D146–D151. [Google Scholar]
- He, H.; Nagy, R.; Liyanarachchi, S.; Jiao, H.; Li, W.; Suster, S.; Kere, J.; de la Chapelle, A. A susceptibility locus for papillary thyroid carcinoma on chromosome 8q24. Cancer Res 2009, 69, 625–631. [Google Scholar]
- Chen, W.; Böcker, W.; Brosius, J.; Tiedge, H. Expression of neural BC200 RNA in human tumours. J. Pathol 1997, 183, 345–351. [Google Scholar]
- Iacoangeli, A.; Lin, Y.; Morley, E.J.; Muslimov, I.A.; Bianchi, R.; Reilly, J.; Weedon, J.; Diallo, R.; Bocker, W.; Tiedge, H. BC200 RNA in invasive and preinvasive breast cancer. Carcinogenesis 2004, 25, 2125–2133. [Google Scholar]
- Chung, S.; Nakagawa, H.; Uemura, M.; Piao, L.; Ashikawa, K.; Hosono, N.; Takata, R.; Akamatsu, S.; Kawaguchi, T.; Morizono, T.; et al. Association of a novel long non-coding RNA in 8q24 with prostate cancer susceptibility. Cancer Sci 2011, 102, 245–252. [Google Scholar]
- Hibi, K.; Nakamura, H.; Hirai, A.; Fujikake, Y.; Kasai, Y.; Akiyama, S.; Ito, K.; Takagi, H. Loss of H19 imprinting in esophageal cancer. Cancer Res 1996, 56, 480–482. [Google Scholar]
- Fellig, Y.; Ariel, I.; Ohana, P.; Schachter, P.; Sinelnikov, I.; Birman, T.; Ayesh, S.; Schneider, T.; de Groot, N.; Czerniak, A.; et al. H19 expression in hepatic metastases from a range of human carcinomas. J. Clin. Pathol 2005, 58, 1064–1068. [Google Scholar]
- Matouk, I.J.; de Groot, N.; Mezan, S.; Ayesh, S.; Abu-lail, R.; Hochberg, A.; Galun, E. The H19 non-coding RNA is essential for human tumor growth. PLoS One 2007, 2. [Google Scholar] [CrossRef]
- Arima, T.; Matsuda, T.; Takagi, N.; Wake, N. Association of IGF2 and H19 imprinting with choriocarcinoma development. Cancer Genet. Cytogenet 1997, 93, 39–47. [Google Scholar]
- Berteaux, N.; Lottin, S.; Monte, D.; Pinte, S.; Quatannens, B.; Coll, J.; Hondermarck, H.; Curgy, J.J.; Dugimont, T.; Adriaenssens, E. H19 mRNA-like noncoding RNA promotes breast cancer cell proliferation through positive control by E2F1. J. Biol. Chem 2005, 280, 29625–29636. [Google Scholar]
- Matouk, I.J.; Abbasi, I.; Hochberg, A.; Galun, E.; Dweik, H.; Akkawi, M. Highly upregulated in liver cancer noncoding RNA is overexpressed in hepatic colorectal metastasis. Eur. J. Gastroenterol. Hepatol 2009, 21, 688–692. [Google Scholar]
- Panzitt, K.; Tschernatsch, M.M.; Guelly, C.; Moustafa, T.; Stradner, M.; Strohmaier, H.M.; Buck, C.R.; Denk, H.; Schroeder, R.; Trauner, M.; et al. Characterization of HULC, a novel gene with striking up-regulation in hepatocellular carcinoma, as noncoding RNA. Gastroenterology 2007, 132, 330–342. [Google Scholar]
- Pasic, I.; Shlien, A.; Durbin, A.D.; Stavropoulos, D.J.; Baskin, B.; Ray, P.N.; Novokmet, A.; Malkin, D. Recurrent focal copy-number changes and loss of heterozygosity implicate two noncoding RNAs and one tumor suppressor gene at chromosome 3q13.31 in osteosarcoma. Cancer Res 2010, 70, 160–171. [Google Scholar]
- Poliseno, L.; Salmena, L.; Zhang, J.; Carver, B.; Haveman, W.J.; Pandolfi, P.P. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature 2010, 465, 1033–1038. [Google Scholar]
- Khaitan, D.; Dinger, M.E.; Mazar, J.; Crawford, J.; Smith, M.A.; Mattick, J.S.; Perera, R.J. The melanoma-upregulated long noncoding RNA SPRY4-IT1 modulates apoptosis and invasion. Cancer Res 2011, 71, 3852–3862. [Google Scholar]
- Wang, F.; Li, X.; Xie, X.; Zhao, L.; Chen, W. UCA1, a non-protein-coding RNA up-regulated in bladder carcinoma and embryo, influencing cell growth and promoting invasion. FEBS Lett 2008, 582, 1919–1927. [Google Scholar]
- Wang, X.S.; Zhang, Z.; Wang, H.C.; Cai, J.L.; Xu, Q.W.; Li, M.Q.; Chen, Y.C.; Qian, X.P.; Lu, T.J.; Yu, L.Z.; et al. Rapid identification of UCA1 as a very sensitive and specific unique marker for human bladder carcinoma. Clin. Cancer Res 2006, 12, 4851–4858. [Google Scholar]
- Dallosso, A.R.; Hancock, A.L.; Malik, S.; Salpekar, A.; King-Underwood, L.; Pritchard-Jones, K.; Peters, J.; Moorwood, K.; Ward, A.; Malik, K.T.; et al. Alternately spliced WT1 antisense transcripts interact with WT1 sense RNA and show epigenetic and splicing defects in cancer. RNA 2007, 13, 2287–2299. [Google Scholar]
- de Kok, J.B.; Verhaegh, G.W.; Roelofs, R.W.; Hessels, D.; Kiemeney, L.A.; Aalders, T.W.; Swinkels, D.W.; Schalken, J.A. DD3(PCA3), a very sensitive and specific marker to detect prostate tumors. Cancer Res 2002, 62, 2695–2689. [Google Scholar]
- Leygue, E. Steroid receptor RNA activator (SRA1): Unusual bifaceted gene products with suspected relevance to breast cancer. Nucl. Recept. Signal. 2007, 4. [Google Scholar] [CrossRef]
- Qi, P.; Du, X. The long non-coding RNAs, a new cancer diagnostic and therapeutic gold mine. Mod. Pathol. 2012. [Google Scholar] [CrossRef]
- Gibb, E.A.; Brown, C.J.; Lam, W.L. The functional role of long non-coding RNA in human carcinomas. Mol. Cancer 2011, 10. [Google Scholar] [CrossRef]
- Morris, K.V. RNA-directed transcriptional gene silencing and activation in human cells. Oligonucleotides 2009, 19, 299–306. [Google Scholar]
- Schöler, N.; Langer, C.; Döhner, H.; Buske, C.; Kuchenbauer, F. Serum microRNAs as a novel class of biomarkers: A comprehensive review of the literature. Exp. Hematol 2010, 38, 1126–1130. [Google Scholar]
- Xie, Y.; Todd, N.W.; Liu, Z.; Zhan, M.; Fang, H.; Peng, H.; Alattar, M.; Deepak, J.; Stass, S.A.; Jiang, F. Altered miRNA expression in sputum for diagnosis of non-small cell lung cancer. Lung Cancer 2010, 67, 170–176. [Google Scholar]
- Xing, L.; Todd, N.W.; Yu, L.; Fang, H.; Jiang, F. Early detection of squamous cell lung cancer in sputum by a panel of microRNA markers. Mod. Pathol 2010, 23, 1157–1164. [Google Scholar]
- Kosaka, N.; Iguchi, H.; Ochiya, T. Circulating microRNA in body fluid: a new potential biomarker for cancer diagnosis and prognosis. Cancer Sci 2010, 101, 2087–2092. [Google Scholar]
- Zhu, Y.; Yu, M.; Li, Z.; Kong, C.; Bi, J.; Li, J.; Gao, Z. ncRAN, a newly identified long noncoding RNA, enhances human bladder tumor growth, invasion, and survival. Urology 2011, 77, 510.e1–510.e5. [Google Scholar]
- Prensner, J.R.; Iyer, M.K.; Balbin, O.A.; Dhanasekaran, S.M.; Cao, Q.; Brenner, J.C.; Laxman, B.; Asangani, I.A.; Grasso, C.S.; Kominsky, H.D.; et al. Transcriptome sequencing across a prostate cancer cohort identifies PCAT-1, an unannotated lincRNA implicated in disease progression. Nat. Biotechnol 2011, 29, 742–749. [Google Scholar]
- Hessels, D.; Klein Gunnewiek, J.M.T.; van Oort, I.; Karthaus, H.F.M.; van Leenders, G.J.L.; van Balken, B.; Kiemeney, L.A.; Witjes, J.A.; Schalken, J.A. DD3PCA3-based Molecular Urine Analysis for the Diagnosis of Prostate Cancer. Eur. Urol 2003, 44, 8–16. [Google Scholar]
- Tinzl, M.; Marberger, M.; Horvath, S.; Chypre, C. DD3PCA3 RNA Analysis in Urine – A New Perspective for Detecting Prostate Cancer. Eur. Urol 2004, 46, 182–187. [Google Scholar]
- Muro, E.M.; Andrade-Navarro, M.A. Pseudogenes as an alternative source of natural antisense transcripts. BMC Evol. Biol. 2010, 10. [Google Scholar] [CrossRef]
- Morris, K.V.; Santoso, S.; Turner, A.M.; Pastori, C.; Hawkins, P.G. Bidirectional transcription directs both transcriptional gene activation and suppression in human cells. PLoS Genet. 2008, 4. [Google Scholar] [CrossRef]
- Lyle, R.; Watanabe, D.; te Vruchte, D.; Lerchner, W.; Smrzka, O.W.; Wutz, A.; Schageman, J.; Hahner, L.; Davies, C.; Barlow, D.P. The imprinted antisense RNA at the Igf2r locus overlaps but does not imprint Mas1. Nat. Genet 2000, 25, 19–21. [Google Scholar]
Figure 1.
Categories of long non-coding RNA (lncRNA).

{kind=link}
{kind=link}
| Name | Cytoband (Size) | Cancer Types | References |
|---|---|---|---|
| AK023948 | 8q24.22 (2807 nt) | Papillary thyroid carcinoma (down regulated) | [72] |
| ANRIL | 9p21.3 (~3.9kb) | Prostate, leukemia | [36,41] |
| BC200 | 2p21 (200 nt) | Breast, cervix, esophagus, lung, ovary, parotid, tongue | [73,74] |
| PRNCR1 | 8q24.2 (13 kb) | Prostate | [75] |
| H19 | 11p15.5 (2.3 kb) | Bladder, lung, liver, breast, esophagus, choriocarcinoma, colon | [57,58,76–80] |
| HOTAIR | 12q13.13 (2.2 kb) | Breast, hepatocellular | [23,29,30,36] |
| HULC | 6p24.3 (~500 nt) | Hepatocellular | [4,81,82] |
| LincRNA-p21 | ~3.1 kb | Represses p53 pathway; induces apoptosis | [70] |
| Loc285194 | 3q13.31 (2105 nt) | Osteosarcoma | [83] |
| Malat1 | 11q13.1 (7.5 kb) | breast, prostate, colon, liver, uterus | [44–47] |
| MEG3 | 14q32.2 (1.6 kb) | Brain (down-regulated) | [62,65] |
| PTNEP1 | 9p13.3 (3.9 kb) | Prostate | [84] |
| Spry4-it1 | 5q31.3 (~700 nt) | Melanoma | [85] |
| SRA | 5q31.3 (1965 nt) | Breast, uterus, ovary (down-regulated) | [26,27] |
| UCA1/CUDR | 19p13.12 (1.4, 2.2, 2.7 kb) | Bladder, colon, cervix, lung, thyroid, liver, breast, esophagus, stomach | [86,87] |
| Wt1-as | 11p13 (isoforms) | acute myeloid leukemia | [88] |
| PCA3 | 9q21.22 (0.6–4 kb) | Prostate | [89] |
| GAS5 | 1q25.1 (isoforms) | Breast (down-regulated) | [68] |
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Hauptman, N.; Glavač, D. Long Non-Coding RNA in Cancer. Int. J. Mol. Sci. 2013, 14, 4655-4669. https://doi.org/10.3390/ijms14034655
AMA Style
Hauptman N, Glavač D. Long Non-Coding RNA in Cancer. International Journal of Molecular Sciences. 2013; 14(3):4655-4669. https://doi.org/10.3390/ijms14034655
Chicago/Turabian StyleHauptman, Nina, and Damjan Glavač. 2013. "Long Non-Coding RNA in Cancer" International Journal of Molecular Sciences 14, no. 3: 4655-4669. https://doi.org/10.3390/ijms14034655