The Role of Lipid Domains in Bacterial Cell Processes

Abstract
:1. Introduction
2. Bacterial Membranes
3. Formation of Lipid Domains
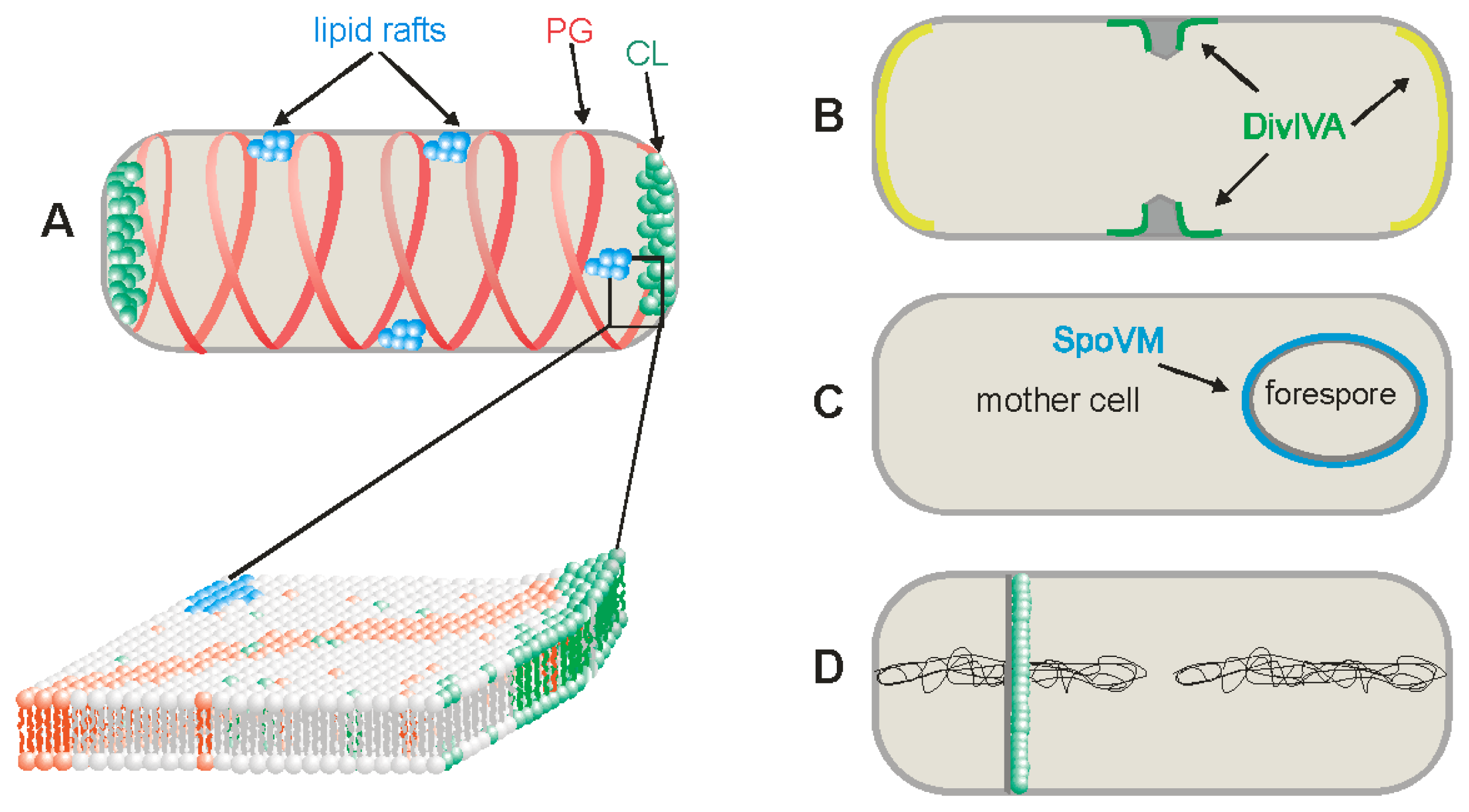
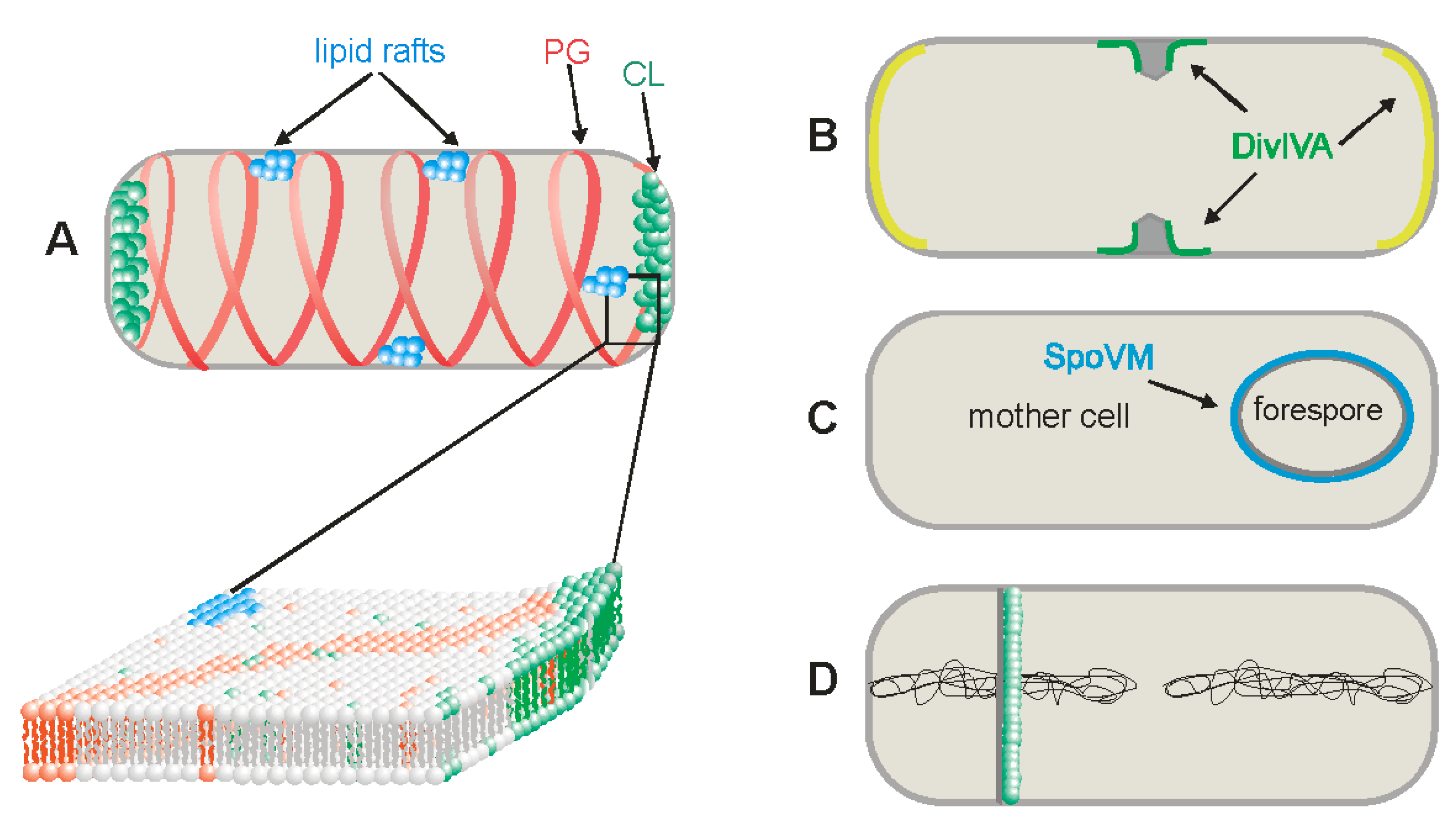
3.1. Cardiolipin Domains
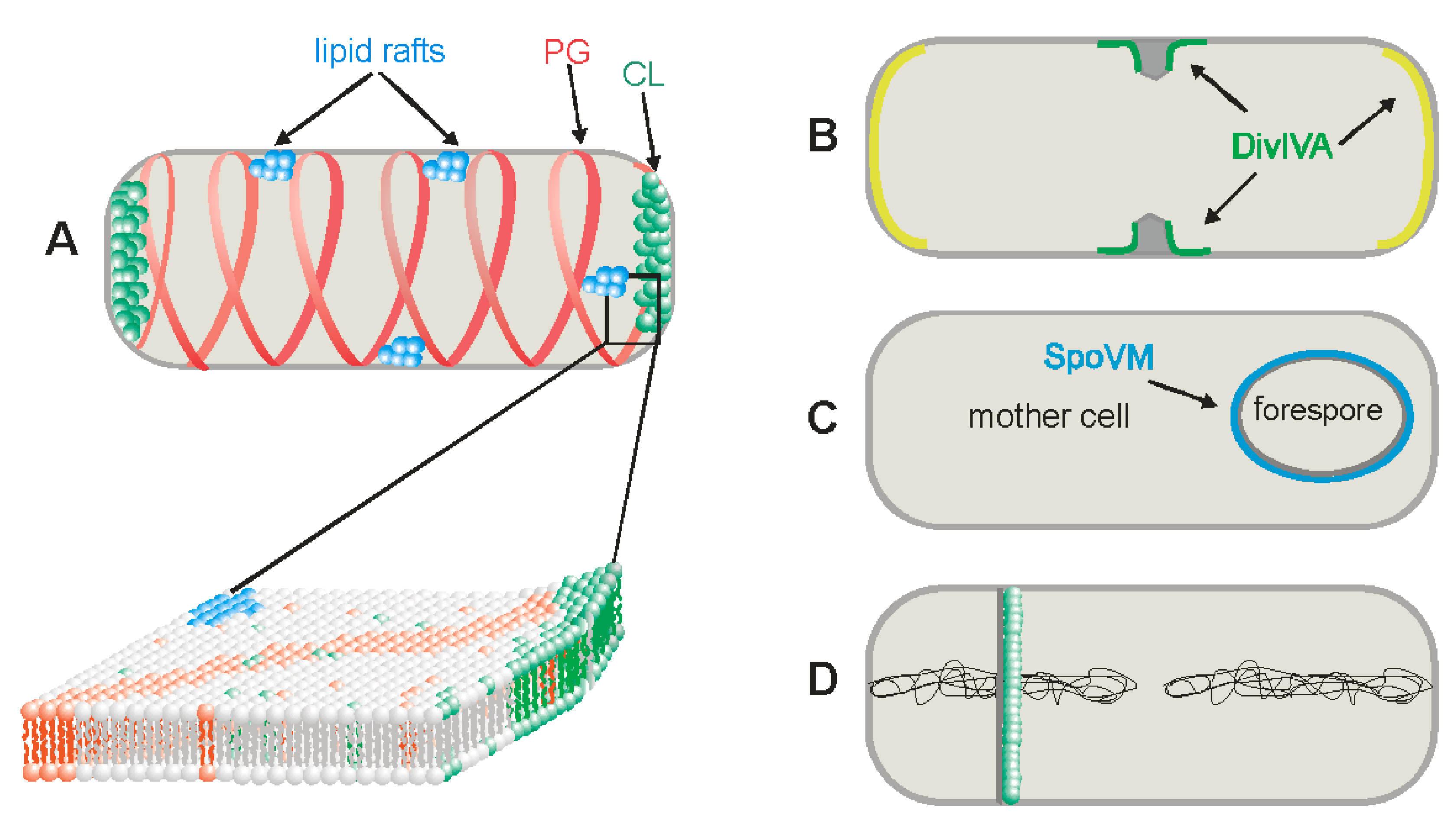
3.2. Phosphatidylglycerol Domains

{kind=link}
{kind=link}
| Lipid domain/organization | Proteins | References |
|---|---|---|
| Cardiolipin—CL | ProP | [22,23] |
| Phosphatidylglycerol—PG | MinD, FtsA, SecA | [5,44] |
| Polyisoprenoids—lipid rafts | FloT, FloA, KinC, YqfA, NfeD2 | [6,46] |
| negative curvature membranes | DivIVA | [7,47] |
| positive curvature membranes | SpoVM | [8] |
| rearagement of lipid membrane composition | SpoIIR | [14] |
| membrane potential | MinD, FtsA, MreB, MreBH, Mbl | [9] |
3.3. Lipid Rafts
4. Negative and Positive Curvature Membranes
5. Role of Membrane Potential for Protein Localization
6. Changes in Lipid Composition of the Membranes during Sporulation
7. Conclusion
Acknowledgments
Conflict of Interest
References
- Govindarajan, S.; Nevo-Dinur, K.; Amster-Choder, O. Compartmentalization and spatiotemporal organization of macromolecules in bacteria. FEMS Microbiol. Rev. 2012, 36, 1005–1022. [Google Scholar]
- Matsumoto, K.; Kusaka, J.; Nishibori, A.; Hara, H. Lipid domains in bacterial membranes. Mol. Microbiol. 2006, 61, 1110–1117. [Google Scholar] [CrossRef]
- Singer, S.J.; Nicolson, G.L. The fluid mosaic model of the structure of cell membranes. Science 1972, 175, 720–731. [Google Scholar]
- Mileykovskaya, E.; Dowhan, W. Cardiolipin membrane domains in prokaryotes and eukaryotes. Biochim. Biophys. Acta 2009, 1788, 2084–2091. [Google Scholar] [CrossRef]
- Barák, I.; Muchová, K.; Wilkinson, A.J.; O’Toole, P.J.; Pavlendová, N. Lipid spirals in Bacillus subtilis and their role in cell division. Mol. Microbiol. 2008, 68, 1315–1327. [Google Scholar] [CrossRef]
- Lopez, D.; Kolter, R. Functional microdomains in bacterial membranes. Genes Dev. 2010, 24, 1893–1902. [Google Scholar] [CrossRef]
- Lenarcic, R.; Halbedel, S.; Visser, L.; Shaw, M.; Wu, L.J.; Errington, J.; Marenduzzo, D.; Hamoen, L.W. Localisation of DivIVA by targeting to negatively curved membranes. EMBO J. 2009, 28, 2272–2282. [Google Scholar] [CrossRef]
- Ramamurthi, K.S.; Lecuyer, S.; Stone, H.A.; Losick, R. Geometric cue for protein localization in a bacterium. Science 2009, 323, 1354–1357. [Google Scholar]
- Strahl, H.; Hamoen, L.W. Membrane potential is important for bacterial cell division. Proc. Natl. Acad. Sci. USA 2010, 107, 12281–12286. [Google Scholar] [CrossRef]
- Vanounou, S.; Pines, D.; Pines, E.; Parola, A.H.; Fishov, I. Coexistence of domains with distinct order and polarity in fluid bacterial membranes. Photochem. Photobiol. 2002, 76, 1–11. [Google Scholar] [CrossRef]
- Vanounou, S.; Parola, A.H.; Fishov, I. Phosphatidylethanolamine and phosphatidylglycerol are segregated into different domains in bacterial membrane. A study with pyrene-labelled phospholipids. Mol. Microbiol. 2003, 49, 1067–1079. [Google Scholar] [CrossRef]
- Mileykovskaya, E.; Dowhan, W. Role of membrane lipids in bacterial division-site selection. Curr. Opin. Microbiol. 2005, 8, 135–142. [Google Scholar]
- Gidden, J.; Denson, J.; Liyanage, R.; Ivey, D.M.; Lay, J.O. Lipid compositions in Escherichia coli and Bacillus subtilis during growth as determined by MALDI-TOF and TOF/TOF mass spectrometry. Int. J. Mass Spectrom. 2009, 283, 178–184. [Google Scholar]
- Pedrido, M.E.; de Oña, P.; Ramirez, W.; Leñini, C.; Goñi, A.; Grau, R. Spo0A links de novo fatty acid synthesis to sporulation and biofilm development in Bacillus subtilis. Mol. Microbiol. 2012. [Google Scholar] [CrossRef]
- Mileykovskaya, E.; Dowhan, W. Visualization of phospholipid domains in Escherichia coli by using the cardiolipin-specific fluorescent dye 10-N-nonyl acridine orange. J. Bacteriol. 2000, 182, 1172–1175. [Google Scholar] [CrossRef]
- Dowhan, W.; Mileykovskaya, E.; Bogdanov, M. Diversity and versatility of lipid-protein interactions revealed by molecular genetic approaches. Biochim. Biophys. Acta 2004, 1666, 19–39. [Google Scholar] [CrossRef]
- Kawai, F.; Shoda, M.; Harashima, R.; Sadaie, Y.; Hara, H.; Matsumoto, K. Cardiolipin domains in Bacillus subtilis Marburg membranes. J. Bacteriol. 2004, 186, 1475–1483. [Google Scholar] [CrossRef]
- Septinus, M.; Berthold, T.; Naujok, A.; Zimmermann, H.W. Hydrophobic acridine dyes for fluorescent staining of mitochondria in living cells. 3. Specific accumulation of the fluorescent dye NAO on the mitochondrial membranes in HeLa cells by hydrophobic interaction. Depression of respiratory activity, changes in the ultrastructure of mitochondria due to NAO. Increase of fluorescence in vital stained mitochondria in situ by irradiation. Histochemistry 1985, 82, 51–66. [Google Scholar] [CrossRef]
- Petit, J.M.; Maftah, A.; Ratinaud, M.H.; Julien, R. 10-N-nonyl acridine orange interacts with cardiolipin and allows the quantification of this phospholipid in isolated mitochondria. Eur. J. Biochem. 1992, 209, 267–273. [Google Scholar] [CrossRef]
- Petit, J.M.; Huet, O.; Gallet, P.F.; Maftah, A.; Ratinaud, M.H.; Julien, R. Direct analysis and significance of cardiolipin transverse distribution in mitochondrial inner membranes. Eur. J. Biochem. 1994, 220, 871–879. [Google Scholar] [CrossRef]
- Mukhopadhyay, R.; Huang, K.C.; Wingreen, N.S. Lipid localization in bacterial cells through curvature-mediated microphase separation. Biophys. J. 2008, 95, 1034–1049. [Google Scholar] [CrossRef]
- Romantsov, T.; Helbig, S.; Culham, D.E.; Gill, C.; Stalker, L.; Wood, J.M. Cardiolipin promotes polar localization of osmosensory transporter ProP in Escherichia coli. Mol. Microbiol. 2007, 64, 1455–1465. [Google Scholar] [CrossRef]
- Mileykovskaya, E. Subcellular localization of Escherichia coli osmosensory transporter ProP: focus on cardiolipin membrane domains. Mol. Microbiol. 2007, 64, 1419–1422. [Google Scholar] [CrossRef]
- Koppelman, C.-M.; den Blaauwen, T.; Duursma, M.C.; Heeren, R.M.A.; Nanninga, N. Escherichia coli minicell membranes are enriched in cardiolipin. J. Bacteriol. 2001, 183, 6144–6147. [Google Scholar] [CrossRef]
- Kawai, F.; Hara, H.; Takamatsu, H.; Watabe, K.; Matsumoto, K. Cardiolipin enrichment in spore membranes and its involvement in germination of Bacillus subtilis Marburg. Genes Genet. Syst. 2006, 81, 69–76. [Google Scholar] [CrossRef]
- Fishov, I.; Woldringh, C.L. Visualization of membrane domains in Escherichia coli. Mol. Microbiol. 1999, 32, 1166–1172. [Google Scholar] [CrossRef]
- Brumback, A.C.; Lieber, J.L.; Angleson, J.K.; Betz, W.J. Using FM1-43 to study neuropeptide granule dynamics and exocytosis. Methods 2004, 33, 287–294. [Google Scholar] [CrossRef]
- Muchova, K.; Wilkinson, A.J.; Barak, I. Changes of lipid domains in Bacillus subtilis cells with disrupted cell wall peptidoglycan. FEMS Microbiol. Lett. 2011, 325, 92–98. [Google Scholar] [CrossRef]
- De Mendoza, D.; Schujman, G.E.; Aguilar, P.S. Bacillus Subtilisand Its Closest Relatives: From Genes to Cells; Sonenshein, A.L., Hoch, J.A., Losick, R., Eds.; ASM Press: Washington, DC, USA, 2002; pp. 43–55. [Google Scholar]
- Harry, E.J. Bacterial cell division: regulating Z-ring formation. Mol. Microbiol. 2001, 40, 795–803. [Google Scholar] [CrossRef]
- Barák, I.; Wilkinson, A.J. Division site recognition in Escherichia coli and Bacillus subtilis. FEMS Microbiol. Rev. 2007, 31, 311–326. [Google Scholar]
- Hu, Z.; Lutkenhaus, J. Topological regulation of cell division in Escherichia coli involves rapid pole to pole oscillation of the division inhibitor MinC under the control of MinD and MinE. Mol. Microbiol. 1999, 34, 82–90. [Google Scholar] [CrossRef]
- Raskin, D.M.; de Boer, P.A. Rapid pole-to-pole oscillation of a protein required for directing division to the middle of Escherichia coli. Proc. Natl. Acad. Sci. USA 1999, 96, 4971–4976. [Google Scholar] [CrossRef]
- Shih, Y.L.; Le, T.; Rothfield, L. Division site selection in Escherichia coli involves dynamic redistribution of Min proteins within coiled structures that extend between the two cell poles. Proc. Natl. Acad. Sci. USA 2003, 100, 7865–7870. [Google Scholar] [CrossRef]
- Edwards, D.H.; Errington, J. The Bacillus subtilis DivIVA protein targets to the division septum and controls site specificity of cell division. Mol. Microbiol. 1997, 24, 905–915. [Google Scholar]
- Bramkamp, M.; Emmins, R.; Weston, L.; Donovan, C.; Daniel, R.A.; Errington, J. A novel component of the division-site selection system of Bacillus subtilis and a new mode of action for the division inhibitor MinCD. Mol. Microbiol. 2008, 70, 1556–1569. [Google Scholar] [CrossRef]
- Szeto, T.H.; Rowland, S.L.; Rothfield, L.I.; King, G.F. Membrane localization of MinD is mediated by a C-terminal motif that is conserved across eubacteria, archaea, and chloroplasts. Proc. Natl. Acad. Sci. USA 2002, 99, 15693–15698. [Google Scholar] [CrossRef]
- Hu, Z.L.; Lutkenhaus, J. A conserved sequence at the C-terminus of MinD is required for binding to the membrane and targeting MinC to the septum. Mol. Microbiol. 2003, 47, 345–355. [Google Scholar] [CrossRef]
- Mileykovskaya, E.; Fishov, I.; Fu, X.; Corbin, B.; Margolin, W.; Dowhan, W. Effects of phospholipid composition on MinD-membrane interactions in vitro and in vivo. J. Biol. Chem. 2003, 278, 22193–22198. [Google Scholar]
- Cornell, R.B.; Taneva, S.G. Amphipathic helices as mediators of the membrane interaction of amphitropic proteins, and as modulators of bilayer physical properties. Curr. Protein Pept. Sci. 2006, 7, 539–552. [Google Scholar] [CrossRef]
- Peters, P.C.; Migocki, M.D.; Thoni, C.; Harry, E.J. A new assembly pathway for the cytokinetic Z ring from a dynamic helical structure in vegetatively growing cells of Bacillus subtilis. Mol. Microbiol. 2007, 64, 487–499. [Google Scholar] [CrossRef]
- Ben-Yehuda, S.; Losick, R. Asymmetric cell division in B. subtilis involves a spiral-like intermediate of the cytokinetic protein FtsZ. Cell 2002, 109, 257–266. [Google Scholar] [CrossRef]
- Pichoff, S.; Lutkenhaus, J. Tethering the Z ring to the membrane through a conserved membrane targeting sequence in FtsA. Mol. Microbiol. 2005, 55, 1722–1734. [Google Scholar] [CrossRef]
- Campo, N.; Tjalsma, H.; Buist, G.; Stepniak, D.; Meijer, M.; Veenhuis, M.; Westermann, M.; Müller, J.P.; Bron, S.; Kok, J.; et al. Subcellular sites for bacterial protein export. Mol. Microbiol. 2004, 53, 1583–1599. [Google Scholar] [CrossRef]
- Gold, V.A.; Robson, A.; Bao, H.; Romantsov, T.; Duong, F.; Collinson, I. The action of cardiolipin on the bacterial translocon. Proc. Natl. Acad. Sci. USA 2010, 107, 10044–10049. [Google Scholar]
- Dempwolff, F.; Moller, H.M.; Graumann, P. Motility and cell shape defects associated with deletions of Flotillin/Reggie paralogs in Bacillus subtilis and interplay of these proteins with NfeD proteins. J. Bacteriol. 2012, 194, 4652–4661. [Google Scholar] [CrossRef]
- Ramamurthi, K.S.; Losick, R. Negative membrane curvature as a cue for subcellular localization of a bacterial protein. Proc. Natl. Acad. Sci. USA 2009, 106, 13541–13545. [Google Scholar] [CrossRef]
- Langhorst, M.F.; Reuter, A.; Stuermer, C.A. Scaffolding microdomains and beyond: The function of reggie/flotillin proteins. Cell Mol. Life Sci. 2005, 62, 2228–2240. [Google Scholar]
- Meile, J.C.; Wu, L.J.; Ehrlich, S.D.; Errington, J.; Noirot, P. Systematic localisation of proteins fused to the green fluorescent protein in Bacillus subtilis: Identification of new proteins at the DNA replication factory. Proteomics 2006, 6, 2135–2146. [Google Scholar] [CrossRef]
- Zhang, H.M.; Li, Z.; Tsudome, M.; Ito, S.; Takami, H.; Horikoshi, K. An alkali-inducible flotillin-like protein from Bacillus halodurans C-125. Protein J. 2005, 24, 125–131. [Google Scholar] [CrossRef]
- Donovan, C.; Bramkamp, M. Characterization and subcellular localization of a bacterial flotillin homologue. Microbiology 2009, 155, 1786–1799. [Google Scholar]
- Liu, C.I.; Liu, G.Y.; Song, Y.; Yin, F.; Hensler, M.E.; Jeng, W.Y.; Nizet, V.; Wang, A.H.; Oldfield, E. A cholesterol biosynthesis inhibitor blocks Staphylococcus aureus virulence. Science 2008, 319, 1391–1394. [Google Scholar]
- Wu, L.J.; Errington, J. Bacillus subtilis SpoIIIE protein required for DNA segregation during asymmetric cell division. Science 1994, 264, 572–575. [Google Scholar]
- Webb, C.D.; Teleman, A.; Gordon, S.; Straight, A.; Belmont, A.; Lin, D.C.; Grossman, A.D.; Wright, A.; Losick, R. Bipolar localization of the replication origin regions of chromosomes in vegetative and sporulating cells of B. subtilis. Cell 1997, 88, 667–674. [Google Scholar] [CrossRef]
- Wu, L.J.; Errington, J. Use of asymmetric cell division and spoIIIE mutants to probe chromosome orientation and organization in Bacillus subtilis. Mol. Microbiol. 1998, 27, 777–786. [Google Scholar]
- Ben-Yehuda, S.; Rudner, D.Z.; Losick, R. RacA, a bacterial protein that anchors chromosomes to the cell poles. Science 2003, 299, 532–536. [Google Scholar] [CrossRef]
- Wu, L.J.; Errington, J. RacA and the Soj-Spo0J system combine to effect polar chromosome segregation in sporulating Bacillus subtilis. Mol. Microbiol. 2003, 49, 1463–1475. [Google Scholar] [CrossRef]
- Edwards, D.H.; Thomaides, H.B.; Errington, J. Promiscuous targeting of Bacillus subtilis cell division protein DivIVA to division sites in Escherichia coli and fission yeast. EMBO J. 2000, 19, 2719–2727. [Google Scholar] [CrossRef]
- Stahlberg, H.; Kutejová, E.; Muchová, K.; Gregorini, M.; Lustig, A.; Muller, S.A.; Olivieri, V.; Engel, A.; Wilkinson, A.J.; Barák, I. Oligomeric structure of the Bacillus subtilis cell division protein DivIVA determined by transmission electron microscopy. Mol. Microbiol. 2004, 52, 1281–1290. [Google Scholar] [CrossRef]
- Fadda, D.; Santona, A.; D’Ulisse, V.; Ghelardini, P.; Ennas, M.G.; Whalen, M.B.; Massidda, O. Streptococcus pneumoniae DivIVA: localization and interactions in a MinCD-free context. J. Bacteriol. 2007, 189, 1288–1298. [Google Scholar] [CrossRef]
- Flardh, K. Essential role of DivIVA in polar growth and morphogenesis in Streptomyces coelicolor A3(2). Mol. Microbiol. 2003, 49, 1523–1536. [Google Scholar] [CrossRef]
- Hempel, A.M.; Wang, S.B.; Letek, M.; Gil, J.A.; Flardh, K. Assemblies of DivIVA mark sites for hyphal branching and can establish new zones of cell wall growth in Streptomyces coelicolor. J. Bacteriol. 2008, 190, 7579–7583. [Google Scholar] [CrossRef]
- Xu, H.; Chater, K.F.; Deng, Z.; Tao, M. A cellulose synthase-like protein involved in hyphal tip growth and morphological differentiation in streptomyces. J. Bacteriol. 2008, 190, 4971–4978. [Google Scholar]
- Letek, M.; Ordonez, E.; Vaquera, J.; Margolin, W.; Flardh, K.; Mateos, L.M.; Gil, J.A. DivIVA is required for polar growth in the MreB lacking rod-shaped actinomycete Corynebacterium glutamicum. J. Bacteriol. 2008, 190, 3283–3292. [Google Scholar]
- Kang, C.M.; Nyayapathy, S.; Lee, J.Y.; Suh, J.W.; Husson, R.N. Wag31, a homologue of the cell division protein DivIVA, regulates growth, morphology and polar cell wall synthesis in mycobacteria. Microbiology 2008, 154, 725–735. [Google Scholar] [CrossRef]
- Moseley, J.B.; Nurse, P. Cell division intersects with cell geometry. Cell 2010, 142, 184–188. [Google Scholar]
- Ramamurthi, K.S.; Clapham, K.R.; Losick, R. Peptide anchoring spore coat assembly to the outer forespore membrane in Bacillus subtilis. Mol. Microbiol. 2006, 62, 1547–1557. [Google Scholar] [CrossRef]
- Salje, J.; van den Ent, F.; de Boer, P.; Lowe, J. Direct membrane binding by bacterial actin MreB. Mol. Cell 2011, 43, 478–487. [Google Scholar]
- Bogdanov, M.; Dowhan, W. Lipid-dependent generation of dual topology for a membrane protein. J. Biol. Chem. 2012, 287, 37939–37948. [Google Scholar]
- Lambert, C.; Mann, S.; Prange, R. Assessment of determinants affecting the dual topology of hepadnaviral large envelope proteins. J. Gen. Virol. 2004, 85, 1221–1225. [Google Scholar]
- Levy, D. Membrane proteins which exhibit multiple topological orientations. Essays Biochem. 1996, 31, 49–60. [Google Scholar]
- Von Heijne, G. Membrane-protein topology. Nat. Rev. Mol. Cell Biol. 2006, 7, 909–918. [Google Scholar]
- Frandsen, N.; Barák, I.; Karmazyn-Campelli, C.; Stragier, P. Transient gene asymmetry during sporulation and establishment of cell specificity in Bacillus subtilis. Genes Dev. 1999, 13, 394–399. [Google Scholar] [CrossRef]
- Wu, L.J.; Errington, J. Septal localization of the SpoIIIE chromosome partitioning protein in Bacillus subtilis. EMBO J. 1997, 16, 2161–2169. [Google Scholar] [CrossRef]
- Fawcett, P.; Eichenberger, P.; Losick, R.; Youngman, P. The transcriptional profile of early to middle sporulation in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2000, 97, 8063–8068. [Google Scholar]
- Losick, R.; Stragier, P. Crisscross regulation of cell-type specific gene expression during development in Bacillus subtilis. Nature 1992, 355, 601–604. [Google Scholar] [CrossRef]
- Barák, I.; Youngman, P. SpoIIE mutants of Bacillus subtilis comprises two distinct phenotypic classes consistent with a dual functional role for the SpoIIE protein. J. Bacteriol. 1996, 178, 4984–4989. [Google Scholar]
- Feucht, A.; Magnin, T.; Yudkin, M.D.; Errington, J. Bifunctional protein required for asymmetric cell division and cell-specific transcription in Bacillus subtilis. Genes Dev. 1996, 10, 794–803. [Google Scholar] [CrossRef]
- Tocheva, E.I.; Matson, E.G.; Morris, D.M.; Moussavi, F.; Leadetter, J.R.; Jensen, G.J. Peptidoglycan remodeling and conversion of an inner membrane into an outer membrane during sporulation. Cell 2011, 146, 799–812. [Google Scholar]
- Landgraf, D.; Okumus, B.; Chien, P.; Baker, T.A.; Paulsson, J. Segregation of molecules at cell division reveals native protein localization. Nat. Methods 2012, 9, 480–482. [Google Scholar] [CrossRef]
- Margolin, W. The price of tags in protein localization studies. J. Bacteriol. 2012, 194, 6369–6371. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Barák, I.; Muchová, K. The Role of Lipid Domains in Bacterial Cell Processes. Int. J. Mol. Sci. 2013, 14, 4050-4065. https://doi.org/10.3390/ijms14024050
Barák I, Muchová K. The Role of Lipid Domains in Bacterial Cell Processes. International Journal of Molecular Sciences. 2013; 14(2):4050-4065. https://doi.org/10.3390/ijms14024050
Chicago/Turabian StyleBarák, Imrich, and Katarína Muchová. 2013. "The Role of Lipid Domains in Bacterial Cell Processes" International Journal of Molecular Sciences 14, no. 2: 4050-4065. https://doi.org/10.3390/ijms14024050