Biofilms: The Stronghold of Legionella pneumophila

{kind=link}
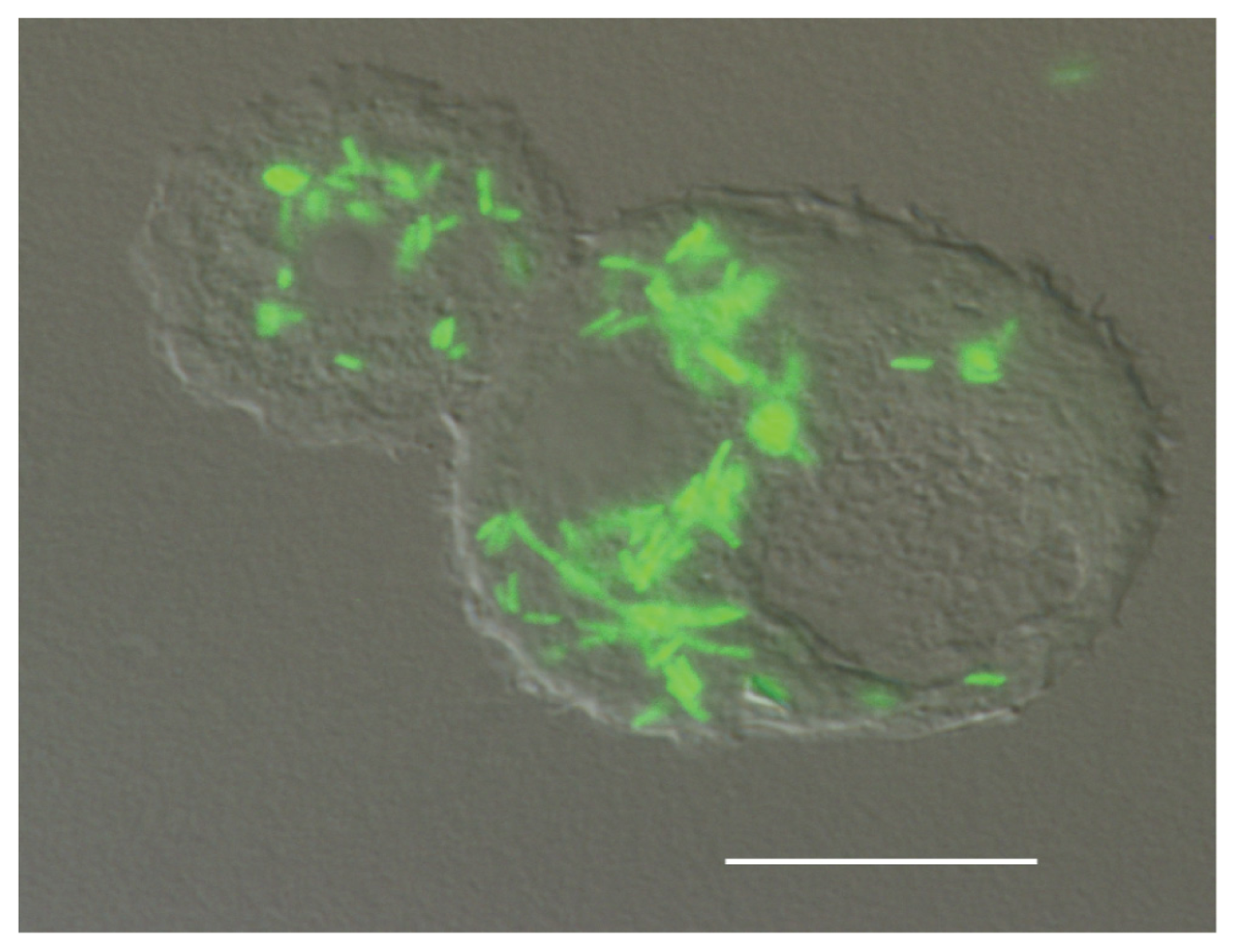{kind=link}
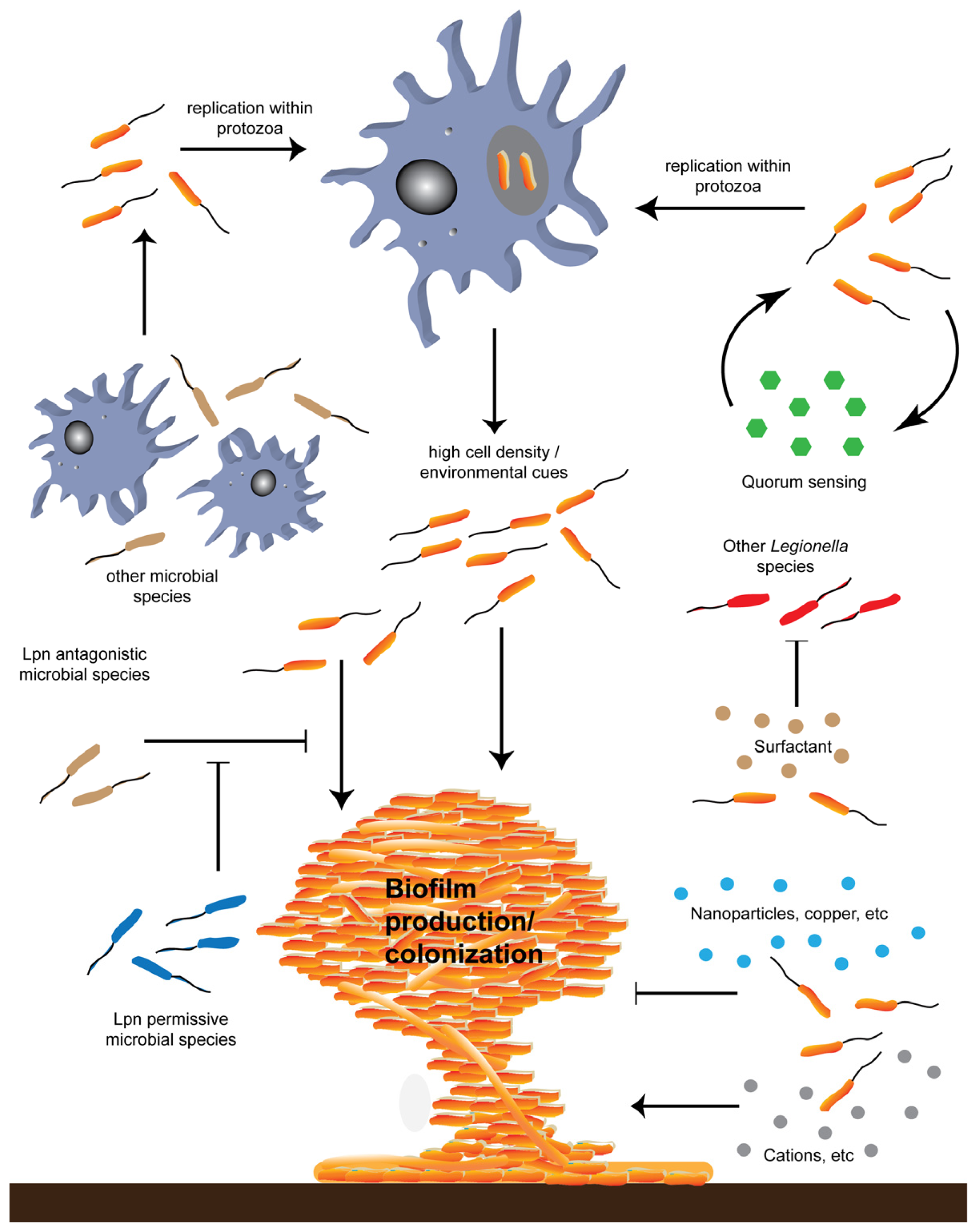
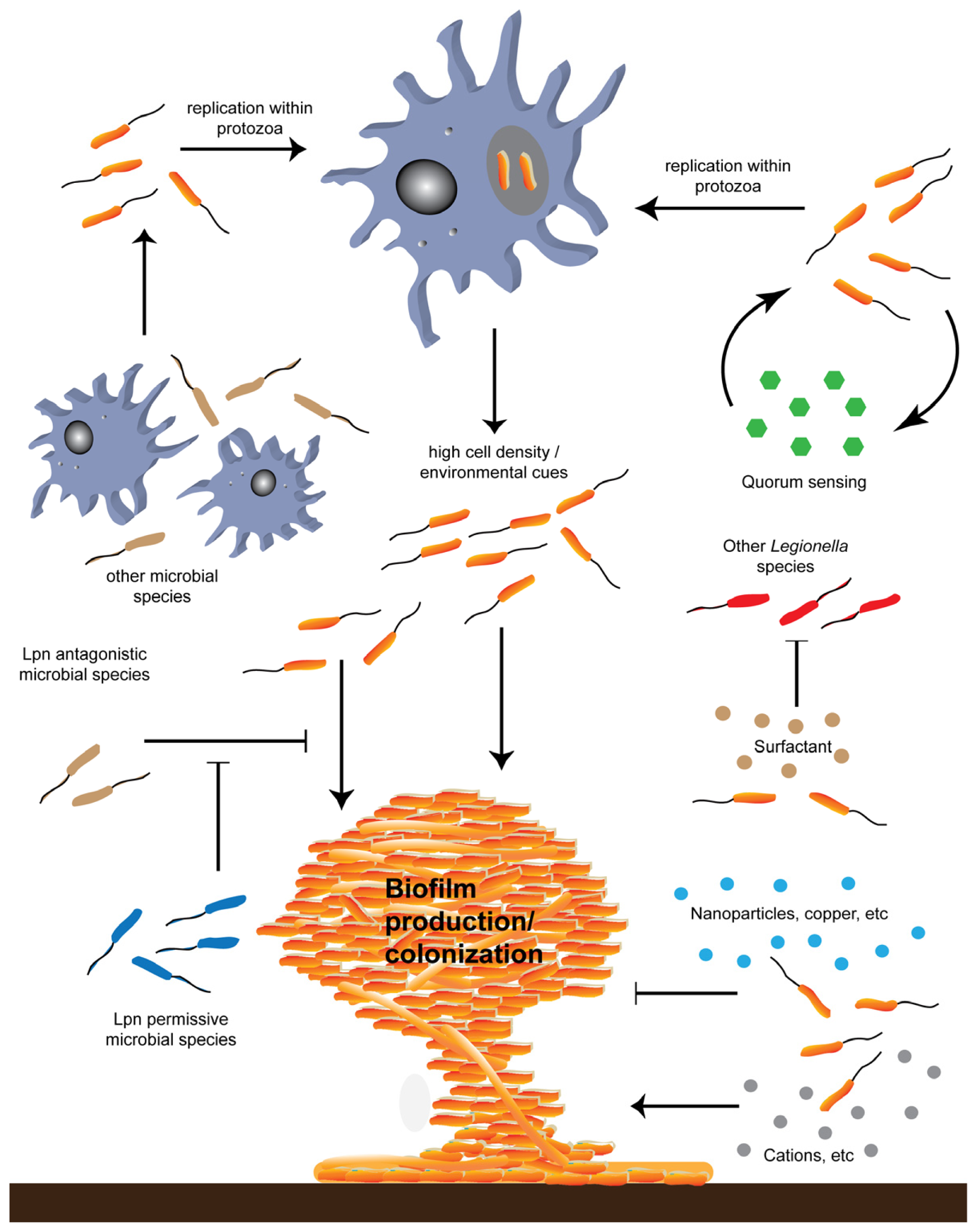{kind=link}
Abstract
:1. Introduction
2. Protozoa and L. pneumophila Biofilm Formation
3. Physiochemical Determinants in L. pneumophila Biofilm Formation and Colonization
4. Regulation of L. pneumophila Endogenous Factors that May Influence Biofilm Colonization
5. The Role of Non-Protozoa Microbial Species in L. pneumophila Biofilm Colonization
6. The Resistance of L. pneumophila Containing Biofilms to Biocides
7. Conclusions


Acknowledgments
Conflicts of Interest
References
- Carratala, J.; Fernandez-Sabe, N.; Ortega, L.; Castellsague, X.; Roson, B.; Dorca, J.; Fernandez-Aguera, A.; Verdaguer, R.; Martinez, J.; Manresa, F.; et al. Outpatient care compared with hospitalization for community-acquired pneumonia: A randomized trial in low-risk patients. Ann. Intern. Med 2005, 142, 165–172. [Google Scholar]
- Yu, V.L.; Plouffe, J.F.; Pastoris, M.C.; Stout, J.E.; Schousboe, M.; Widmer, A.; Summersgill, J.; File, T.; Heath, C.M.; Paterson, D.L.; et al. Distribution of Legionella species and serogroups isolated by culture in patients with sporadic community-acquired Legionellosis: An international collaborative survey. J. Infect. Dis 2002, 186, 127–128. [Google Scholar]
- Bentham, R.H.; Broadbent, C.R. A model for autumn outbreaks of Legionnaires’ disease associated with cooling towers, linked to system operation and size. Epidemiol. Infect 1993, 111, 287–295. [Google Scholar]
- Fields, B.S.; Benson, R.F.; Besser, R.E. Legionella and Legionnaires’ disease: 25 years of investigation. Clin. Microbiol. Rev 2002, 15, 506–526. [Google Scholar]
- Feazel, L.M.; Baumgartner, L.K.; Peterson, K.L.; Frank, D.N.; Kirk Harris, J.; Pace, N.R. Opportunistic pathogens enriched in showerhead biofilms. Proc. Natl. Acad. Sci. USA 2009, 106, 16393–16398. [Google Scholar]
- McEvoy, M.; Batchelor, N.; Hamilton, G.; MacDonald, A.; Faiers, M.; Sills, A.; Lee, J.; Harrison, T. A cluster of cases of Legionnaires’ disease associated with exposure to a spa pool on display. Commun. Dis. Public Health 2000, 3, 43–45. [Google Scholar]
- Wadowsky, R.M.; Yee, R.B.; Mezmar, L.; Wing, E.J.; Dowling, J.N. Hot water systems as sources of Legionella pneumophila in hospital and nonhospital plumbing fixtures. Appl. Environ. Microbiol 1982, 43, 1104–1110. [Google Scholar]
- Hindre, T.; Bruggemann, H.; Buchrieser, C.; Hechard, Y. Transcriptional profiling of Legionella Pneumophila biofilm cells and the influence of iron on biofilm formation. Microbiology 2008, 154, 30–41. [Google Scholar]
- Pecastaings, S.; Berge, M.; Dubourg, K.M.; Roques, C. Sessile Legionella pneumophila is able to grow on surfaces and generate structured monospecies biofilms. Biofouling 2010, 26, 809–819. [Google Scholar]
- Kwon, S.; Moon, E.; Kim, T.; Hong, S.; Park, H. Pyrosequencing demonstrated complex microbial communities in a membrane filtration system for a drinking water treatment plant. Microbes Environ 2011, 26, 149–155. [Google Scholar]
- Murga, R.; Forster, T.S.; Brown, E.; Pruckler, J.M.; Fields, B.S.; Donlan, R.M. Role of biofilms in the survival of Legionella pneumophila in a model potable-water system. Microbiology 2001, 147, 3121–3126. [Google Scholar]
- Rowbotham, T.J. Pontiac fever, ameobae and Legionllae. Lancet 1981, 317, 40–41. [Google Scholar]
- Molmeret, M.; Horn, M.; Wagner, M.; Santic, M.; Abu Kwaik, Y. Amoebae as training grounds for intracellular bacterial pathogens. Appl. Environ. Microbiol 2005, 71, 20–28. [Google Scholar]
- Steinert, M.; Hentschel, U.; Hacker, J. Legionella pneumophila: An aquatic microbe goes astray. FEMS Microbiol. Rev 2002, 26, 149–162. [Google Scholar]
- Yan, L.; Cerny, R.L.; Cirillo, J.D. Evidence that hsp90 is involved in the altered interactions of Acanthamoeba castellanii variants with bacteria. Eukaryotic Cell 2004, 3, 567–578. [Google Scholar]
- Chaabna, Z.; Forey, F.; Reyrolle, M.; Jarraud, S.; Atlan, D.; Fontvieille, D.; Gilbert, C. Molecular diversity and high virulence of Legionella pneumophila strains isolated from biofilms developed within a warm spring of a thermal spa. BMC Microbiol 2013, 13, 17. [Google Scholar]
- Abu Khweek, A.; Fernandez Davila, N.S.; Caution, K.; Akhter, A.; Abdulrahman, B.A.; Tazi, M.; Hassan, H.; Novotny, L.A.; Bakaletz, L.O.; Amer, A.O. Biofilm-derived Legionella pneumophila evades the innate immune response in macrophages. Front. Cell Infect. Microbiol 2013, 3, 18. [Google Scholar]
- Barker, J.; Lambert, P.A.; Brown, M.R. Influence of intra-amoebic and other growth conditions on the surface properties of Legionella pneumophila. Infect. Immun 1993, 61, 3503–3510. [Google Scholar]
- Newsome, A.L.; Farone, M.B.; Berk, S.G.; Gunderson, J.H. Free living amoebae as opportunistic hosts for intracellular bacterial parasites. J. Eukaryot. Microbiol 2001, 48, 13S–14S. [Google Scholar]
- Rowbotham, T.J. Isolation of Legionella pneumophila from clinical specimens via amoebae, and the interaction of those and other isolates with amoebae. J. Clin. Pathol 1983, 36, 978–986. [Google Scholar]
- Skinner, A.R.; Anand, C.M.; Malic, A.; Kurtz, J.B. Acanthamoebae and environmental spread of Legionella pneumophila. Lancet 1983, 2, 289–290. [Google Scholar]
- Taylor, M.; Ross, K.; Bentham, R. Legionella, protozoa, and biofilms: Interactions within complex microbial systems. Microb. Ecol 2009, 58, 538–547. [Google Scholar]
- Valster, R.M.; Wullings, B.A.; van der Kooij, D. Detection of protozoan hosts for Legionella pneumophila in engineered water systems by using a biofilm batch test. Appl. Environ. Microbiol 2010, 76, 7144–7153. [Google Scholar]
- Valster, R.M.; Wullings, B.A.; van den Berg, R.; van der Kooij, D. Relationships between free-living protozoa, cultivable Legionella spp., and water quality characteristics in three drinking water supplies in the caribbean. Appl. Environ. Microbiol 2011, 77, 7321–7328. [Google Scholar]
- Liu, R.; Yu, Z.; Guo, H.; Liu, M.; Zhang, H.; Yang, M. Pyrosequencing analysis of eukaryotic and bacterial communities in faucet biofilms. Sci. Total Environ 2012, 435–436, 124–131. [Google Scholar]
- Mampel, J.; Spirig, T.; Weber, S.S.; Haagensen, J.A.; Molin, S.; Hilbi, H. Planktonic replication is essential for biofilm formation by Legionella pneumophila in a complex medium under static and dynamic flow conditions. Appl. Environ. Microbiol 2006, 72, 2885–2895. [Google Scholar]
- Temmerman, R.; Vervaeren, H.; Noseda, B.; Boon, N.; Verstraete, W. Necrotrophic growth of Legionella pneumophila. Appl. Environ. Microbiol 2006, 72, 4323–4328. [Google Scholar]
- Declerck, P.; Behets, J.; van Hoef, V.; Ollevier, F. Replication of Legionella pneumophila in floating biofilms. Curr. Microbiol 2007, 55, 435–440. [Google Scholar]
- Hsu, B.; Huang, C.; Chen, J.; Chen, N.; Huang, J. Comparison of potentially pathogenic free-living amoeba hosts by Legionella spp. in substrate-associated biofilms and floating biofilms from spring environments. Water Res 2011, 45, 5171–5183. [Google Scholar]
- Atlas, R.M. Legionella: From environmental habitats to disease pathology, detection and control. Environ. Microbiol 1999, 1, 283–293. [Google Scholar]
- Hoffman, P.S. Invasion of eukaryotic cells by Legionella pneumophila: A common strategy for all hosts? Can. J. Infect. Dis 1997, 8, 139–146. [Google Scholar]
- Donlan, R.M.; Forster, T.; Murga, R.; Brown, E.; Lucas, C.; Carpenter, J.; Fields, B. Legionella pneumophila associated with the protozoan Hartmannella vermiformis in a model multi-species biofilm has reduced susceptibility to disinfectants. Biofouling 2005, 21, 1–7. [Google Scholar]
- Thomas, V.; Bouchez, T.; Nicolas, V.; Robert, S.; Loret, J.F.; Levi, Y. Amoebae in domestic water systems: Resistance to disinfection treatments and implication in Legionella persistence. J. Appl. Microbiol 2004, 97, 950–963. [Google Scholar]
- Storey, M.V.; Winiecka-Krusnell, J.; Ashbolt, N.J.; Stenstrom, T. The efficacy of heat and chlorine treatment against thermotolerant Acanthamoebae andLegionellae. Scand. J. Infect. Dis 2004, 36, 656–662. [Google Scholar]
- Wright, J.B.; Ruseska, I.; Athar, M.A.; Corbett, S.; Costerton, J.W. Legionella pneumophila grows adherent to surfaces in vitro and in situ. Infect. Control Hosp. Epidemiol. 1989, 10, 408–415. [Google Scholar]
- Rogers, J.; Dowsett, A.B.; Dennis, P.J.; Lee, J.V.; Keevil, C.W. Influence of plumbing materials on biofilm formation and growth of Legionella pneumophila in potable water systems. Appl. Environ. Microbiol 1994, 60, 1842–1851. [Google Scholar]
- Rogers, J.; Dowsett, A.B.; Dennis, P.J.; Lee, J.V.; Keevil, C.W. Influence of temperature and plumbing material selection on biofilm formation and growth of Legionella pneumophila in a model potable water system containing complex microbial flora. Appl. Environ. Microbiol 1994, 60, 1585–1592. [Google Scholar]
- Moritz, M.M.; Flemming, H.; Wingender, J. Integration of Pseudomonas aeruginosa and Legionella pneumophila in drinking water biofilms grown on domestic plumbing materials. Int. J. Hyg. Environ. Health 2010, 213, 190–197. [Google Scholar]
- Geesey, G.G.; Wigglesworth-Cooksey, B.; Cooksey, K.E. Influence of calcium and other cations on surface adhesion of bacteria and diatoms: A review. Biofouling 2000, 15, 195–205. [Google Scholar]
- Koubar, M.; Rodier, M.H.; Frere, J. Involvement of minerals in adherence of Legionella pneumophila to surfaces. Curr. Microbiol 2013, 66, 437–442. [Google Scholar]
- Yaradou, D.F.; Raze, D.; Ginevra, C.; Ader, F.; Doleans-Jordheim, A.; Vandenesch, F.; Menozzi, F.D.; Etienne, J.; Jarraud, S. Zinc-dependent cytoadherence of Legionella pneumophila to human alveolar epithelial cells in vitro. Microb. Pathog. 2007, 43, 234–242. [Google Scholar]
- Bargellini, A.; Marchesi, I.; Righi, E.; Ferrari, A.; Cencetti, S.; Borella, P.; Rovesti, S. Parameters predictive of Legionella contamination in hot water systems: Association with trace elements and heterotrophic plate counts. Water Res 2011, 45, 2315–2321. [Google Scholar]
- Borella, P.; Montagna, M.T.; Romano-Spica, V.; Stampi, S.; Stancanelli, G.; Triassi, M.; Marchesi, I.; Bargellini, A.; Neglia, R.; Paglionico, N.; et al. Relationship between mineral content of domestic hot water and microbial contamination. J. Trace Elem. Med. Biol 2003, 17, 37–43. [Google Scholar]
- Boyd, C.D.; Chatterjee, D.; Sondermann, H.; O’Toole, G.A. LapG, required for modulating biofilm formation by Pseudomonas fluorescens Pf0-1, is a calcium-dependent protease. J. Bacteriol 2012, 194, 4406–4414. [Google Scholar]
- Chatterjee, D.; Boyd, C.D.; O’Toole, G.A.; Sondermann, H. Structural characterization of a conserved, calcium-dependent periplasmic protease from Legionella pneumophila. J. Bacteriol 2012, 194, 4415–4425. [Google Scholar]
- Giao, M.S.; Wilks, S.A.; Azevedo, N.F.; Vieira, M.J.; Keevil, C.W. Comparison between standard culture and peptide nucleic acid 16S rRNA hybridization quantification to study the influence of physico-chemical parameters on Legionella pneumophila survival in drinking water biofilms. Biofouling 2009, 25, 343–351. [Google Scholar]
- Pang, C.M.; Liu, W. Biological filtration limits carbon availability and affects downstream biofilm formation and community structure. Appl. Environ. Microbiol 2006, 72, 5702–5712. [Google Scholar]
- Van der Kooij, D.K.; Veenendaal, H.R.; Scheffer, W.J.H. Biofilm formation and multiplication of Legionella in a model warm water system with pipes of copper, stainless steel and cross-linked polyethylene. Water Res 2005, 39, 2789–2798. [Google Scholar]
- Martinelli, F.; Caruso, A.; Moschini, L.; Turano, A.; Scarcella, C.; Speziani, F. A comparison of Legionella pneumophila occurrence in hot water tanks and instantaneous devices in domestic, nosocomial, and community environments. Curr. Microbiol 2000, 41, 374–376. [Google Scholar]
- Ciesielski, C.A.; Blaser, M.J.; Wang, W.L. Role of stagnation and obstruction of water flow in isolation of Legionella pneumophila from hospital plumbing. Appl. Environ. Microbiol 1984, 48, 984–987. [Google Scholar]
- Fisher-Hoch, S.; Bartlett, C.L.; Tobin, J.O.; Gillett, M.B.; Nelson, A.M.; Pritchard, J.E.; Smith, M.G.; Swann, R.A.; Talbot, J.M.; Thomas, J.A. Investigation and control of an outbreaks of Legionnaires’ disease in a district general hospital. Lancet 1981, 1, 932–936. [Google Scholar]
- Makin, T.; Pride, M. Venturi system could be major breakthrough. Health Estate 2010, 64, 27–33. [Google Scholar]
- Storey, M.V.; Langmark, J.; Ashbolt, N.J.; Stenstrom, T.A. The fate of Legionella e within distribution pipe biofilms: Measurement of their persistence, inactivation and detachment. Water Sci. Technol 2004, 49, 269–275. [Google Scholar]
- Lehtola, M.J.; Torvinen, E.; Kusnetsov, J.; Pitkanen, T.; Maunula, L.; von Bonsdorff, C.; Martikainen, P.J.; Wilks, S.A.; Keevil, C.W.; Miettinen, I.T. Survival of Mycobacterium avium, Legionella pneumophila, Escherichia coli, and caliciviruses in drinking water-associated biofilms grown under high-shear turbulent flow. Appl. Environ. Microbiol 2007, 73, 2854–2859. [Google Scholar]
- Liu, Z.; Lin, Y.E.; Stout, J.E.; Hwang, C.C.; Vidic, R.D.; Yu, V.L. Effect of flow regimes on the presence of Legionella within the biofilm of a model plumbing system. J. Appl. Microbiol 2006, 101, 437–442. [Google Scholar]
- Fliermans, C.B.; Cherry, W.B.; Orrison, L.H.; Smith, S.J.; Tison, D.L.; Pope, D.H. Ecological distribution of Legionella pneumophila. Appl. Environ. Microbiol 1981, 41, 9–16. [Google Scholar]
- Stout, J.E.; Yu, V.L.; Best, M.G. Ecology of Legionella pneumophila within water distribution systems. Appl. Environ. Microbiol 1985, 49, 221–228. [Google Scholar]
- Tiaden, A.; Spirig, T.; Sahr, T.; Walti, M.A.; Boucke, K.; Buchrieser, C.; Hilbi, H. The autoinducer synthase LqsA and putative sensor kinase lqss regulate phagocyte interactions, extracellular filaments and a genomic island of Legionella pneumophila. Environ. Microbiol 2010, 12, 1243–1259. [Google Scholar]
- Kessler, A.; Schell, U.; Sahr, T.; Tiaden, A.; Harrison, C.; Buchrieser, C.; Hilbi, H. The Legionella pneumophila orphan sensor kinase LqsT regulates competence and pathogen-host interactions as a component of the LAI-1 circuit. Environ. Microbiol 2013, 15, 646–662. [Google Scholar]
- Vandersmissen, L.; de Buck, E.; Saels, V.; Coil, D.A.; Anne, J. A Legionella pneumophila collagen-like protein encoded by a gene with a variable number of tandem repeats is involved in the adherence and invasion of host cells. FEMS Microbiol. Lett 2010, 306, 168–176. [Google Scholar]
- Duncan, C.; Prashar, A.; So, J.; Tang, P.; Low, D.E.; Terebiznik, M.; Guyard, C. Lcl of Legionella pneumophila is an immunogenic GAG binding adhesin that promotes interactions with lung epithelial cells and plays a crucial role in biofilm formation. Infect. Immun 2011, 6, 2168–2181. [Google Scholar]
- Mallegol, J.; Duncan, C.; Prashar, A.; So, J.; Low, D.E.; Terebeznik, M.; Guyard, C. Essential roles and regulation of the Legionella pneumophila collagen-like adhesin during biofilm formation. PLoS One 2012, 7, e46462. [Google Scholar]
- Stone, B.J.; Kwaik, Y.A. Expression of multiple pili by Legionella pneumophila: Identification and characterization of a type IV pilin gene and its role in adherence to mammalian and protozoan cells. Infect. Immun 1998, 66, 1768–1775. [Google Scholar]
- Lucas, C.E.; Brown, E.; Fields, B.S. Type IV Pili and Type II secretion play a limited role in Legionella pneumophila biofilm colonization and retention. Microbiology 2006, 152, 3569–3573. [Google Scholar]
- De Buck, E.; Maes, L.; Meyen, E.; van Mellaert, L.; Geukens, N.; Anne, J.; Lammertyn, E. Legionella pneumophila Philadelphia-1 tatB and tatC affect intracellular replication and biofilm formation. Biochem. Biophys. Res. Commun 2005, 331, 1413–1420. [Google Scholar]
- Cianciotto, N.P. Iron acquisition by Legionella pneumophila. Biometals 2007, 20, 323–331. [Google Scholar]
- Orsi, N. The antimicrobial activity of lactoferrin: Current status and perspectives. Biometals 2004, 17, 189–196. [Google Scholar]
- Robey, M.; Cianciotto, N.P. Legionella pneumophila feoAB promotes ferrous iron uptake and intracellular infection. Infect. Immun 2002, 70, 5659–5669. [Google Scholar]
- Tiaden, A.; Hilbi, H. A-hydroxyketone synthesis and sensing by Legionella and Vibrio. Sensors 2012, 12, 2899–2919. [Google Scholar]
- Spirig, T.; Tiaden, A.; Kiefer, P.; Buchrieser, C.; Vorholt, J.A.; Hilbi, H. The Legionella autoinducer synthase LqsA produces an α-hydroxyketone signaling molecule. J. Biol. Chem 2008, 283, 18113–18123. [Google Scholar]
- Tiaden, A.; Spirig, T.; Hilbi, H. Bacterial gene regulation by α-hydroxyketone signaling. Trends Microbiol 2010, 18, 288–297. [Google Scholar]
- Tiaden, A.; Spirig, T.; Carranza, P.; Bruggemann, H.; Riedel, K.; Eberl, L.; Buchrieser, C.; Hilbi, H. Synergistic contribution of the Legionella pneumophila Lqs genes to pathogen-host interactions. J. Bacteriol 2008, 190, 7532–7547. [Google Scholar]
- McDonough, K.A.; Rodriguez, A. The myriad roles of cyclic AMP in microbial pathogens: From signal to sword. Nat. Rev. Microbiol 2011, 10, 27–38. [Google Scholar]
- Levi, A.; Folcher, M.; Jenal, U.; Shuman, H.A. Cyclic diguanylate signaling proteins control intracellular growth of Legionella pneumophila. mBio 2011, 2, e00316–10. [Google Scholar]
- Carlson, H.K.; Vance, R.E.; Marletta, M.A. H-NOX regulation of c-di-GMP metabolism and biofilm formation in Legionella pneumophila. Mol. Microbiol 2010, 77, 930–942. [Google Scholar]
- Hammer, B.K.; Swanson, M.S. Co-ordination of Legionella pneumophila virulence with entry into stationary phase by ppGpp. Mol. Microbiol 1999, 33, 721–731. [Google Scholar]
- Bachman, M.A.; Swanson, M.S. RpoS co-operates with other factors to induce Legionella pneumophila virulence in the stationary phase. Mol. Microbiol 2001, 40, 1201–1214. [Google Scholar]
- Tiaden, A.; Spirig, T.; Weber, S.S.; Bruggemann, H.; Bosshard, R.; Buchrieser, C.; Hilbi, H. The Legionella pneumophila response regulator LqsR promotes host cell interactions as an element of the virulence regulatory network controlled by RpoS and LetA. Cell. Microbiol 2007, 9, 2903–2920. [Google Scholar]
- Hammer, B.K.; Tateda, E.S.; Swanson, M.S. A two-component regulator induces the transmission phenotype of stationary-phase Legionella pneumophila. Mol. Microbiol 2002, 44, 107–118. [Google Scholar]
- Molofsky, A.B.; Swanson, M.S. Legionella pneumophila CsrA is a pivotal repressor of transmission traits and activator of replication. Mol. Microbiol 2003, 50, 445–461. [Google Scholar]
- Piao, Z.; Sze, C.C.; Barysheva, O.; Iida, K.; Yoshida, S. Temperature-regulated formation of mycelial mat-like biofilms by Legionella pneumophila. Appl. Environ. Microbiol 2006, 72, 1613–1622. [Google Scholar]
- Konishi, T.; Yamashiro, T.; Koide, M.; Nishizono, A. Influence of temperature on growth of Legionella pneumophila biofilm determined by precise temperature gradient incubator. J. Biosci. Bioeng 2006, 101, 478–484. [Google Scholar]
- Justice, S.S.; Hunstad, D.A.; Cegelski, L.; Hultgren, S.J. Morphological plasticity as a bacterial survival strategy. Nat. Rev. Microbiol 2008, 6, 162–168. [Google Scholar]
- Prashar, A.; Bhatia, S.; Tabatabaeiyazdi, Z.; Duncan, C.; Garduno, R.A.; Tang, P.; Low, D.E.; Guyard, C.; Terebiznik, M.R. Mechanism of invasion of lung epithelial cells by filamentous Legionella pneumophila. Cell. Microbiol 2012, 14, 1632–1655. [Google Scholar]
- Liles, M.R.; Viswanathan, V.K.; Cianciotto, N.P. Identification and temperature regulation of Legionella pneumophila genes involved in type IV pilus biogenesis and type II protein secretion. Infect. Immun 1998, 66, 1776–1782. [Google Scholar]
- Messi, P.; Anacarso, I.; Bargellini, A.; Bondi, M.; Marchesi, I.; de Niederhausern, S.; Borella, P. Ecological behaviour of three serogroups of Legionella pneumophila within a model plumbing system. Biofouling 2011, 27, 165. [Google Scholar]
- Tison, D.L.; Pope, D.H.; Cherry, W.B.; Fliermans, C.B. Growth of Legionella pneumophila in association with blue-green algae (Cyanobacteria). Appl. Environ. Microbiol 1980, 39, 456–459. [Google Scholar]
- Wadowsky, R.M.; Yee, R.B. Satellite growth of Legionella pneumophila with an environmental isolate of Flavobacterium breve. Appl. Environ. Microbiol 1983, 46, 1447–1449. [Google Scholar]
- Wery, N.; Bru-Adan, V.; Minervini, C.; Delgenes, J.; Garrelly, L.; Godon, J. Dynamics of Legionella spp. and bacterial populations during the proliferation of L. pneumophila in a cooling tower facility. Appl. Environ. Microbiol 2008, 74, 3030–3037. [Google Scholar]
- Stewart, C.R.; Burnside, D.M.; Cianciotto, N.P. The surfactant of Legionella pneumophila is secreted in a TolC-dependent manner and is antagonistic toward other Legionella species. J. Bacteriol 2011, 193, 5971–5984. [Google Scholar]
- Stewart, C.R.; Muthye, V.; Cianciotto, N.P. Legionella pneumophila persists within biofilms formed by Klebsiella pneumoniae, Flavobacterium sp., and Pseudomonas fluorescens under dynamic flow conditions. PLoS One 2012, 7, e50560. [Google Scholar]
- Kimura, S.; Tateda, K.; Ishii, Y.; Horikawa, M.; Miyairi, S.; Gotoh, N.; Ishiguro, M.; Yamaguchi, K. Pseudomonas aeruginosa las quorum sensing autoinducer suppresses growth and biofilm production in Legionella species. Microbiology 2009, 155, 1934–1939. [Google Scholar]
- Declerck, P.; Behets, J.; Delaedt, Y.; Margineanu, A.; Lammertyn, E.; Ollevier, F. Impact of non-Legionella bacteria on the uptake and intracellular replication of Legionella pneumophila in Acanthamoeba castellanii and Naegleria lovaniensis. Microb. Ecol 2005, 50, 536–549. [Google Scholar]
- Emtiazi, F.; Schwartz, T.; Marten, S.M.; Krolla-Sidenstein, P.; Obst, U. Investigation of natural biofilms formed during the production of drinking water from surface water embankment filtration. Water Res 2004, 38, 1197–1206. [Google Scholar]
- Giao, M.S.; Wilks, S.; Azevedo, N.F.; Vieira, M.J.; Keevil, C.W. Incorporation of natural uncultivable Legionella pneumophila into potable water biofilms provides a protective niche against chlorination stress. Biofouling 2009, 25, 335–341. [Google Scholar]
- Alleron, L.; Merlet, N.; Lacombe, C.; Frere, J. Long-term survival of Legionella pneumophila in the viable but nonculturable state after monochloramine treatment. Curr. Microbiol 2008, 57, 497–502. [Google Scholar]
- Raftery, T.D.; Lindler, H.; McNealy, T.L. Altered host cell–bacteria interaction due to nanoparticle interaction with a bacterial biofilm. Microb. Ecol 2013, 65, 496–503. [Google Scholar]
- Stojak, A.R.; Raftery, T.; Klaine, S.J.; McNealy, T.L. Morphological responses of Legionella pneumophila biofilm to nanoparticle exposure. Nanotoxicology 2011, 5, 730–742. [Google Scholar]
- Rogers, J.; Dowsett, A.B.; Keevil, C.W. A paint incorporating silver to control mixed biofilms containing Legionella pneumophila. J. Ind. Microbiol 1995, 15, 377–383. [Google Scholar]
- Liu, Z.; Stout, J.E.; Boldin, M.; Rugh, J.; Diven, W.F.; Yu, V.L. Intermittent use of copper-silver ionization for Legionella control in water distribution systems: A potential option in buildings housing individuals at low risk of infection. Clin. Infect Dis 1998, 26, 138–140. [Google Scholar]
- Schwartz, T.; Hoffmann, S.; Obst, U. Formation of natural biofilms during chlorine dioxide and u.v. disinfection in a public drinking water distribution system. J. Appl. Microbiol 2003, 95, 591–601. [Google Scholar]
- Walker, J.T.; Mackerness, C.W.; Mallon, D.; Makin, T.; Williets, T.; Keevil, C.W. Control of Legionella pneumophila in a hospital water system by chlorine dioxide. J. Ind. Microbiol 1995, 15, 384–390. [Google Scholar]
- Pryor, M.; Springthorpe, S.; Riffard, S.; Brooks, T.; Huo, Y.; Davis, G.; Sattar, S.A. Investigation of opportunistic pathogens in municipal drinking water under different supply and treatment regimes. Water Sci. Technol 2004, 50, 83–90. [Google Scholar]
- Williams, M.M.; Braun-Howland, E. Growth of Escherichia coli in model distribution system biofilms exposed to hypochlorous acid or monochloramine. Appl. Environ. Microbiol 2003, 69, 5463–5471. [Google Scholar]
- Kusnetsov, J.M.; Keskitalo, P.J.; Ahonen, H.E.; Tulkki, A.I.; Miettinen, I.T.; Martikainen, P.J. Growth of Legionella and other heterotrophic bacteria in a circulating cooling water system exposed to ultraviolet irradiation. J. Appl. Microbiol 1994, 77, 461–466. [Google Scholar]
- Cargill, K.L.; Pyle, B.H.; Sauer, R.L.; McFeters, G.A. Effects of culture conditions and biofilm formation on the iodine susceptibility of Legionella pneumophila. Can. J. Microbiol 1992, 38, 423–429. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Abdel-Nour, M.; Duncan, C.; Low, D.E.; Guyard, C. Biofilms: The Stronghold of Legionella pneumophila. Int. J. Mol. Sci. 2013, 14, 21660-21675. https://doi.org/10.3390/ijms141121660
Abdel-Nour M, Duncan C, Low DE, Guyard C. Biofilms: The Stronghold of Legionella pneumophila. International Journal of Molecular Sciences. 2013; 14(11):21660-21675. https://doi.org/10.3390/ijms141121660
Chicago/Turabian StyleAbdel-Nour, Mena, Carla Duncan, Donald E. Low, and Cyril Guyard. 2013. "Biofilms: The Stronghold of Legionella pneumophila" International Journal of Molecular Sciences 14, no. 11: 21660-21675. https://doi.org/10.3390/ijms141121660