HuR-Regulated mRNAs Associated with Nuclear hnRNP A1-RNP Complexes

 ,
,
Abstract
:
1. Introduction
2. Results
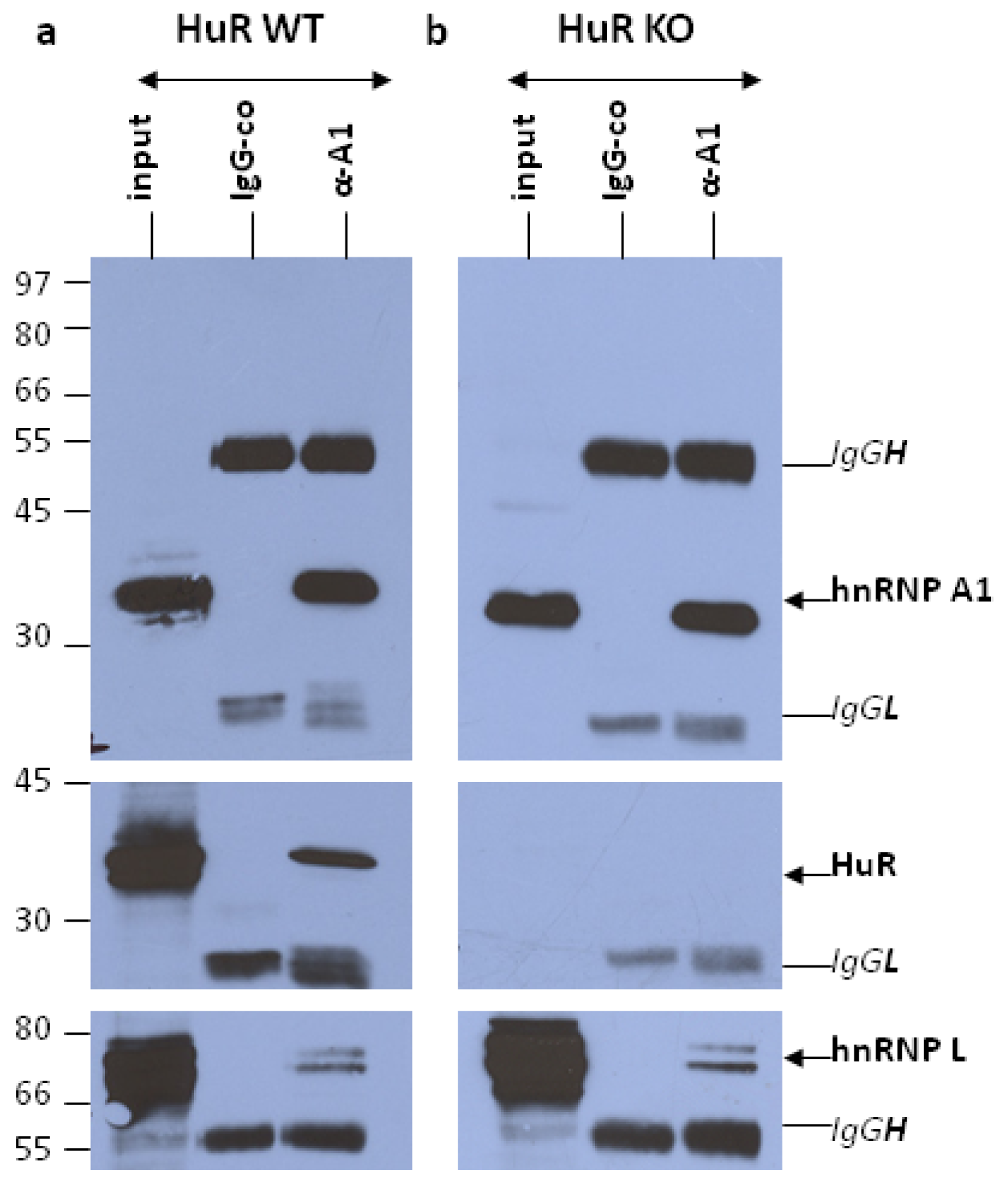
2.1. Ribonomic Profiling of hnRNP A1-RNP Complexes Isolated from Nuclear Extracts of HuR-Containing and HuR-Lacking MEF Cells
2.1.1. RNA Immunoprecipitation (RIP) Assay
2.1.2. Microarray (Chip) Analysis
2.2. Differential Association of Target RNA Transcripts with hnRNP A1-RNP Complexes in Nuclear Extracts of HuR-Containing and HuR-Lacking MEFs
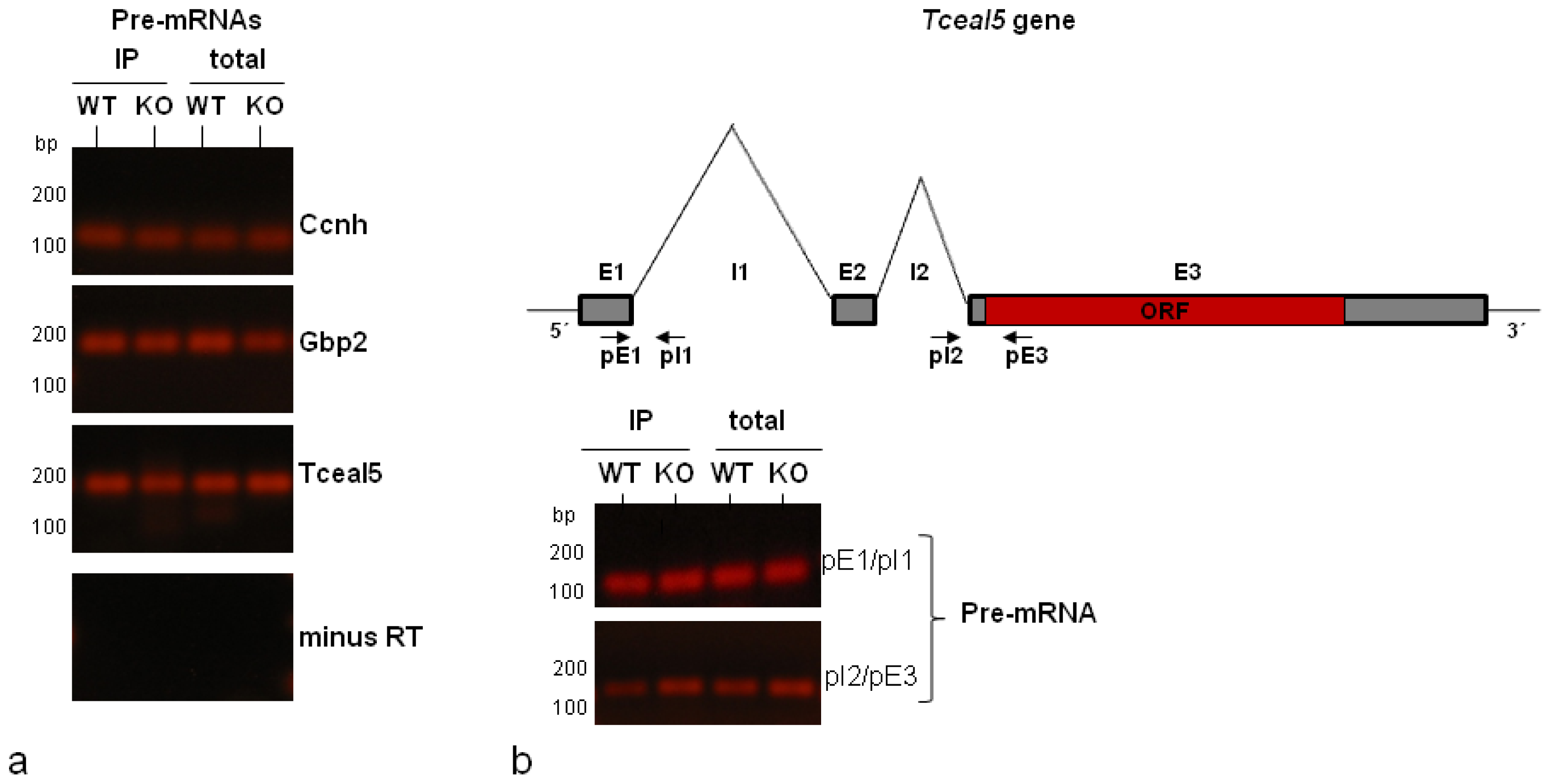
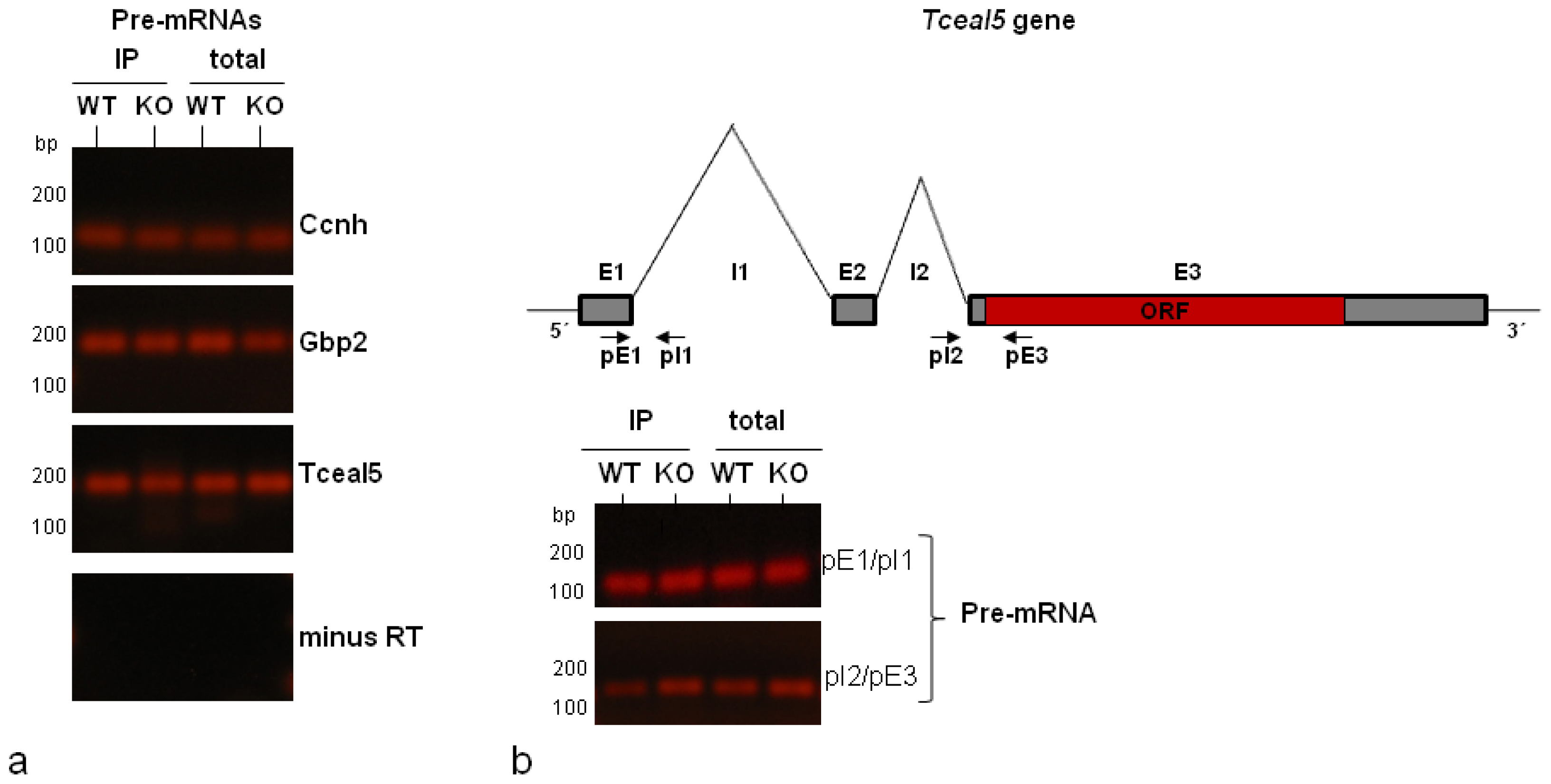
2.3. Validation by RT-qPCR of Selected mRNA Targets: Presence of Spliced and Pre-Spliced mRNAs in Isolated hnRNP A1-RNP Complexes
3. Discussion
4. Experimental Section
4.1. Experimental Cellular System
4.2. Nuclear Extract Preparation
4.3. RNA Immunoprecipitation Coupled with Microarray Analysis—RIP-Chip Assay
4.3.1. RNA Immunoprecipitation (RIP) Step
4.3.2. RNA Extraction Step
4.3.3. Microarray Hybridization and Data Analysis
4.4. Immunoblotting
4.5. Real-Time PCR Analysis (RT-qPCR)
5. Conclusions



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Definition | p-value | Fold increase/decrease | |
|---|---|---|---|---|
| Up-regulated genes | Pitx2 | paired-like homeodomain transcription factor 2, transcript variant 2 | 0.00503 | 90 |
| Psmd8 | proteasome (prosome, macropain) 26S subunit, non-ATPase, 8 | 0.00007 | 53 | |
| Gm15698 | predicted gene, transcription elongation factor B (SIII), polypeptide 2, non-coding RNA. | 0.00363 | 23 | |
| Glrx1 | Glutaredoxin | 0.00080 | 21 | |
| Parm1 | prostate androgen-regulated mucin-like protein 1 | 0.00021 | 21 | |
| Gfpt1 | glutamine fructose-6-phosphate transaminase 1 | 0.00050 | 17 | |
| Csprs | cytokine receptor-like factor 1 | 0.00041 | 15 | |
| Thumpd1 | THUMP domain containing 1 | 0.00002 | 13 | |
| Sorbs2 | sorbin and SH3 domain containing 2 | 0.00298 | 13 | |
| Mef2c | myocyte enhancer factor 2C | 0.00551 | 12 | |
| Down-regulated genes | Xlr4a | X-linked lymphocyte-regulated 4A | 0.00200 | 81 |
| Mmp3 | Matrix metallopeptidase 3 | 0.00006 | 71 | |
| Cyp7b1 | cytochrome P450, family 7, subfamily b, polypeptide 1 | 0.00143 | 69 | |
| Rsad2 | radical S-adenosyl methionine domain containing 2 | 0.00161 | 49 | |
| Osr2 | odd-skipped related 2 (Drosophila) | 0.00032 | 48 | |
| Tomm22 | translocase of outer mitochondrial membrane 22 homolog (yeast), nuclear gene encoding mitochondrial protein | 0.00012 | 42 | |
| Atf4 | Activating transcription factor 4 | 0.00098 | 32 | |
| Mmp13 | matrix metallopeptidase 13 | 0.00397 | 31 | |
| Usp18 | ubiquitin specific peptidase 18 | 0.00562 | 24 | |
| Mmp10 | matrix metallopeptidase 10 | 0.00190 | 23 | |
| Gene Symbol | Definition | p-value | Enrichment | |
|---|---|---|---|---|
| Top Targets in HuR WT | Ppp1r11 | protein phosphatase 1, regulatory (inhibitor) subunit 11 | 0.00041 | 4.90 |
| Pdcd5 | programmed cell death 5 | 0.00412 | 4.78 | |
| Auh | AU RNA binding protein/enoyl-coenzyme A hydratase | 0.00822 | 4.26 | |
| Top Targets in HuR KO | Serf1 | small EDRK-rich factor 1 | 0.00029 | 5.52 |
| Gm4832 | predicted gene Gm4832 | 0.00018 | 5.38 | |
| Dynlt3 | dynein light chain Tctex-type 3 | 0.00361 | 5.18 | |
| Ccnh | cyclin H | 0.00072 | 5.03 | |
| Spp1 | secreted phosphoprotein 1 | 0.00006 | 5.01 | |
| Uchl5 | ubiquitin carboxyl-terminal esterase L5 | 0.00004 | 4.97 | |
| Mrps18c | mitochondrial ribosomal protein S18C | 0.00002 | 4.84 | |
| Hspe1 | heat shock protein 1 (chaperonin 10) | 0.00008 | 4.83 | |
| Ndufa2 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, assembly factor 2 | 0.00047 | 4.81 | |
| Tnfaip8 | tumor necrosis factor, alpha-induced protein 8 | 0.00781 | 4.81 | |
| Etfa | electron transferring flavoprotein, alpha polypeptide | 0.00035 | 4.76 | |
| Rps3a | ribosomal protein S3a | 0.00271 | 4.66 | |
| Dazap2 | DAZ associated protein 2 | 0.00008 | 4.53 | |
| Cfdp1 | craniofacial development protein 1 | 0.00169 | 4.52 | |
| Rexo2 | REX2, RNA exonuclease 2 homolog (S. cerevisiae) | 0.00016 | 4.45 | |
| Naca | nascent polypeptide-associated complex alpha polypeptide | 0.00021 | 4.41 | |
| Gnpda2 | Glucosamine-6-phosphate deaminase 2 | 0.00008 | 4.34 | |
| 1810022K09Rik | RIKEN cDNA 1810022K09 gene | 0.00000 | 4.22 | |
| Auh | AU RNA binding protein/enoyl-coenzyme A hydratase | 0.00258 | 4.19 | |
| Hspa8 | Heat shock protein 8 | 0.00126 | 4.16 | |
| Cox20 | COX20 Cox2 chaperone | 0.00121 | 4.04 | |
| Ppp1r11 | protein phosphatase 1, regulatory (inhibitor) subunit 11 | 0.00287 | 4.01 | |
| GO Annotation | p-value | Enrichment | |
|---|---|---|---|
| Targets in HuR WT | Translation | 0.00000000001 | 32/228 |
| protein transport | 0.00000000039 | 26/435 | |
| protein folding | 0.00000000234 | 12/99 | |
| intracellular protein transmembrane transport | 0.00000000797 | 9/56 | |
| RNA splicing | 0.00000002600 | 14/166 | |
| transport | 0.00000007535 | 49/1500 | |
| metabolic process | 0.00000014388 | 26/574 | |
| mRNA processing | 0.00000014604 | 15/216 | |
| transcription | 0.00000115190 | 44/1412 | |
| response to oxidative stress | 0.00000167931 | 6/40 | |
| oxidation reduction | 0.00000416171 | 22/527 | |
| protein catabolic process | 0.00000510429 | 5/31 | |
| regulation of transcription, DNA-dependent | 0.00000672647 | 44/1512 | |
| electron transport chain | 0.00001006502 | 8/94 | |
| translational initiation | 0.00001094670 | 4/21 | |
| Targets in HuR KO | translation | 0.00000000006 | 32/228 |
| transcription | 0.00000000007 | 68/1412 | |
| protein transport | 0.00000000008 | 36/435 | |
| regulation of transcription, DNA-dependent | 0.00000000009 | 71/1512 | |
| Ubiquitin-dependent protein catabolic process | 0.00000000016 | 30/441 | |
| apoptosis | 0.00000003281 | 23/354 | |
| transport | 0.00000009805 | 56/1500 | |
| cell cycle | 0.00000053421 | 23/412 | |
| vesicle-mediated transport | 0.00000090952 | 12/134 | |
| response to DNA damage stimulus | 0.00000377796 | 13/176 | |
| DNA repair | 0.00000923405 | 12/165 | |
| small GTPase mediated signal transduction | 0.00004585070 | 12/192 | |
| oxidation reduction | 0.00009243521 | 22/527 | |
| cell division | 0.00040989564 | 11/209 | |
| Mitosis | 0.00095668335 | 8/140 | |
| intracellular protein transport | 0.00134751563 | 8/147 | |
| mRNA processing | 0.00184940082 | 10/216 | |
| RNA splicing | 0.00308169664 | 8/166 | |
| metabolic process | 0.00382078845 | 19/574 | |
| Gene Symbol | Definition | Features | Ratio of Enrichments KO/WT |
|---|---|---|---|
| Tceal5 | transcription elongation factor A (SII)-like 5 | Relief of transcription arrest by pol II; chromatin modification | 3.52 |
| Ccnh | cyclin H | Transcription-coupled repair; meiotic progression; embryonic development | 2.77 |
| Gbp2 | guanylate binding protein 2 | Member of GTPases; immune effector | 1.23 |
| Hoxc10 | homeo box C10 | Embryonic development; cellular transformation | 0.55 |
| Rad23a | RAD23a homolog (S. cerevisiae) | DNA damage response; proteasome degradation; oxidative stress | 0.57 |
| HnRNP A1 | heterogeneous nuclear ribonucleoprotein A1 | Prototype hnRNP protein; RNA processing (splicing and nuclear export) | 1.84 |
| Ccnc | cyclin c | Cell cycle progression and apoptosis | 2.38 |
| Mea1 | Male-enhanced antigen 1 | Mouse spermatogenesis | 0.38 |
Acknowledgments
Conflicts of Interest
References
- Moore, M.J. From birth to death: The complex life of eukaryotic mRNAs. Science 2005, 309, 1514–1518. [Google Scholar]
- Glisovic, T.; Bachorik, J.L.; Yong, J.; Dreyfuss, G. RNA-binding proteins and post-transcriptional gene regulation. FEBS Lett 2008, 58, 1977–1986. [Google Scholar]
- Dreyfuss, G.; Kim, V.N.; Kataoka, N. Messenger-RNA-binding proteins and the messages they carry. Nat. Rev. Mol. Cell Biol 2002, 3, 195–205. [Google Scholar]
- Han, S.P.; Tang, Y.H.; Smith, R. Functional diversity of the hnRNPs: Past, present and perspectives. Biochem. J 2010, 430, 379–392. [Google Scholar]
- Bailey-Serres, J.; Sorenson, R.; Juntawong, P. Getting the message across: Cytoplasmic ribonucleoprotein complexes. Trends Plant Sci 2009, 14, 443–453. [Google Scholar]
- Wahl, M.C.; Will, C.I.; Luhrmann, R. The spliceosome: Design principles of a dynamic RNP machine. Cell 2009, 136, 701–718. [Google Scholar]
- Yates, L.A.; Norbury, C.J.; Gilbert, R.J. The long and short of microRNA. Cell 2013, 153, 516–519. [Google Scholar]
- Dreyfuss, G.; Matunis, J.M.; Pinol-Roma, S.; Burd, G.C. hnRNP proteins and the biogenesis of mRNA. Annu. Rev. Biochem 1993, 62, 289–321. [Google Scholar]
- Krecic, A.M.; Swanson, M.S. hnRNP complexes: Composition, structure, and function. Curr. Opin. Cell Biol 1999, 11, 363–371. [Google Scholar]
- He, Y.; Smith, R. Nuclear functions of heterogeneous nuclear ribonucleoproteins A/B. Cell. Mol. Life Sci 2009, 66, 1239–1256. [Google Scholar]
- Pinol-Roma, S.; Dreyfuss, G. Shuttling of pre-mRNA binding proteins between nucleus and cytoplasm. Nature 1992, 355, 730–732. [Google Scholar]
- Cammas, A.; Pileur, F.; Bonnal, S.; Lewis, S.M.; Leveque, N.; Holcik, M.; Vagner, S. Cytoplasmic relocalization of heterogeneous nuclear ribonucleoprotein A1 controls translation initiation of specific mRNAs. Mol. Biol. Cell 2007, 18, 5048–5059. [Google Scholar]
- Hinman, M.N.; Lou, H. Diverse molecular functions of the Hu proteins. Cell. Mol. Life Sci 2008, 64, 3168–3181. [Google Scholar]
- Fan, X.C.; Steitz, J.A. Overexpression of HuR, a nuclear-cytoplasmic shuttling protein, increases the in vivo stability of ARE-containing mRNAs. EMBO J 1998, 17, 3448–3460. [Google Scholar]
- Shim, J.; Karin, M. The control of mRNA stability in response to extracellular stimuli. Mol. Cell 2002, 14, 323–331. [Google Scholar]
- Barreau, C.; Paillard, L.; Osborne, H.B. AU-rich elements and associated factors: Are there unifying principles? Nucleic Acids Res 2005, 33, 7138–7150. [Google Scholar]
- Pullmann, R.; Kim, H.H.; Abdelmohsen, K.; Lal, A.; Martindale, J.I.; Yang, X.; Gorospe, M. Analysis of turnover and translation regulatory RNA-binding protein expression through binding to cognate mRNAs. Mol. Cell. Biol 2007, 27, 6265–6278. [Google Scholar]
- Zhao, T.T.; Graber, T.E.; Jordan, L.E.; Cloutier, M.; Lewis, S.M.; Goulet, I.; Cote, L.; Holcik, M. hnRNP A1 regulates UV-induced NF-kappaB signalling through destabilisation of cIAP1 mRNA. Cell Death Differ 2009, 16, 244–252. [Google Scholar]
- Keene, J.D. Why is Hu where? Shuttling of early-response-gene messenger RNA subsets. Proc. Natl. Acad. Sci. USA 1999, 96, 5–7. [Google Scholar]
- Gallouzi, I.E.; Steitz, J.A. Delineation of mRNA export pathways by the use of cell-permeable peptides. Science 2001, 294, 1895–1901. [Google Scholar]
- Zhu, H.; Zhou, H.-I.; Hasman, R.A.; Lou, H. Hu proteins regulate polyadenylation by blocking sites containing U-rich sequences. J. Biol. Chem 2007, 282, 2203–2210. [Google Scholar]
- Mukherjee, N.; Corcoran, D.L.; Nusbaum, J.D.; Reid, D.W.; Georgiev, S.; Hafner, M.; Ascano, M.; Tuschl, T.; Ohler, U.J.D.; Keene, J.D. Integrative regulatory mapping indicates that the RNA-binding protein HuR couples pre-mRNA processing and mRNA stability. Mol. Cell 2011, 43, 327–339. [Google Scholar]
- Lopez de Silanes, I.; Lal, A.; Gorospe, M. HuR: Post-transcriptional paths to malignancy. RNA Biol 2005, 2, 11–13. [Google Scholar]
- Srikantan, S.; Gorospe, M. HuR function in disease. Front. Biosci 2012, 17, 189–205. [Google Scholar]
- Tenenbaum, S.A.; Carson, C.C.; Lager, P.J.; Keene, J.D. Identifying mRNA subsets in messenger ribonucleoprotein complexes by using cDNA arrays. Proc. Natl. Acad. Sci. USA 2000, 97, 14085–14090. [Google Scholar]
- Hieronymus, H.; Silver, P.A. A systems view of mRNA biology. Genes Dev 2004, 18, 2845–2860. [Google Scholar]
- König, J.; Zarnack, K.; Rot, G.; Curk, T.; Kayikci, M.; Zupan, B.; Turner, D.J.; Luscombe, N.M.; Ule, J. iCLIP reveals the function of hnRNP particles in splicing at individual nucleotide resolution. Nat. Struct. Mol. Biol 2010, 17, 909–915. [Google Scholar]
- Ule, J.; Jensen, K.B.; Ruggiu, M.; Mele, A.; Ule, A.; Darnell, R.B. CLIP identifies Nova-regulated RNA networks in the brain. Science 2003, 302, 1212–1215. [Google Scholar]
- Kishore, S.; Jaskiewicz, L.; Burger, L.; Hausser, J.; Khorshid, M.; Zavolan, M. A quantitative analysis of CLIP methods for identifying binding sites of RNA-binding proteins. Nat. Methods 2011, 8, 559–564. [Google Scholar]
- Hafner, M.; Landthaler, M.; Burger, L.; Khorshid, M.; Hausser, J. Transcriptome-wide identification of RNA-binding protein and micro RNA target sites by PAR-CLIP. Cell 2010, 141, 129–141. [Google Scholar]
- Papadopoulou, C.; Patrinou-Georgoula, M.; Guialis, A. Extensive association of HuR with hnRNP proteins within immunoselected hnRNP and mRNP complexes. Biochim. Biophys. Acta 2010, 1804, 692–703. [Google Scholar]
- Papadopoulou, C.; Ganou, V.; Patrinou-Georgoula, M.; Guialis, A. HuR-hnRNP interactions and the effect of cellular stress. Mol. Cell. Biochem 2013, 372, 137–147. [Google Scholar]
- Katsanou, V.; Milatos, S.; Yiakouvaki, A.; Sgantzis, N.; Kotsoni, A.; Alexiou, M.; Harokopos, V.; Aidinis, V.; Hemberger, M.; Kontoyiannis, D.L. The RNA-binding protein Elavl1/HuR is essential for placental branching morphogenesis and embryonic development. Mol. Cell. Biol 2009, 29, 2762–2776. [Google Scholar]
- Dreyfuss, G.; Choi, Y.D.; Adam, S.A. Characterization of heterogeneous nuclear RNA-protein complexes in vivo with monoclonal antibodies. Mol. Cell. Biol 1984, 4, 1104–1114. [Google Scholar]
- Mili, S.; Shu, H.J.; Zhao, Y.; Pinol-Roma, S. Distinct RNP complexes of shuttling hnRNP proteins with pre-mRNA and mRNA: Candidate intermediates in formation and export of mRNA. Mol. Cell. Biol 2001, 21, 7307–7319. [Google Scholar]
- Eiring, A.M.; Neviani, P.; Santhanam, R.; Oaks, J.J.; Chang, J.S.; Notari, M.; Willis, W.; Gambacorti-Passerini, C.; Volinia, S.; Marcucci, G.; et al. Identification of novel posttranscriptional targets of the BCR/ABL oncoprotein by ribonomics: Requirements of E2F3 for BCR/ABL leukemogenesis. Blood 2008, 111, 816–828. [Google Scholar]
- Lal, A.; Mazan-Mamczarz, K.; Kawai, T.; Yang, X.; Martindale, J.L.; Gorospe, M. Concurrent vs. individual binding of HuR and AUF1 to common labile target mRNAs. EMBO J 2004, 23, 3092–3102. [Google Scholar]
- Calaluce, R.; Gubin, M.M.; Davis, J.W.; Magee, J.D.; Chen, J.; Kuwano, Y.; Gorospe, M.; Atasoy, U. The RNA binding protein HuR differentially regulates unique subsets of mRNAs in estrogen receptor negative and estrogen receptor positive breast cancer. BMC Cancer 2010, 10, 126–140. [Google Scholar]
- Masuda, K.; Abdelmohsen, K.; Kim, M.M.; Srikantan, S. Global dissociation of HuR-mRNA complexes promotes cell survival after ionizing radiation. EMBO J 2011, 30, 1040–1053. [Google Scholar]
- Mazan-Mamczarz, K.; Hagner, P.R.; Dai, B.; Wood, W.H.; Zhang, Y.; Becker, K.G.; Liu, Z.; Gartenhaus, R.B. Identification of transformation-related pathways in a breast epithelial cell model using a ribonomics approach. Cancer Res 2008, 68, 7730–7735. [Google Scholar]
- Vo, D.T.; Korshunov, A.; Dubuc, A.M.; Northcott, P.A.; Smith, A.D.; Pfister, S.M.; Taylor, M.D.; Janga, S.C.; Anant, S.; Vogel, C.; et al. The RNA-binding protein Musashi1 affects medulloblastoma growth via a network of cancer-related genes and is an indicator of poor prognosis. Am. J. Pathol 2012, 181, 1762–1772. [Google Scholar]
- Lebedera, S.; Jens, M.; Theil, K.; Schwanhausser, B.; Selbach, M.; Landthaler, M.; Rajewsky, N. Transcriptome-wide analysis of regulatory interactions of the RNA-binding protein HuR. Mol. Cell 2011, 43, 340–352. [Google Scholar]
- St Laurent, G.; Shtokalo, D.; Heydarian, M.; Palyanov, A.; Babiy, D.; Zhou, J.; Kumar, A.; Urcuqui-Inchima, S. Insights from the HuR-interacting transcriptome: ncRNAs, ubiquitin pathways, and patterns of secondary structure dependent RNA interactions. Mol. Genet. Genomics 2012, 287, 867–879. [Google Scholar]
- Van der Spek, P.J.; Smit, E.M.; Beverloo, H.B.; Sugasawa, K.; Masutani, C.; Hanaoka, F.; Hoeijmakers, J.H.; Hagemeijer, A. Chromosomal localization of three repair genes: The xeroderma pigmentosum group C gene and two human homologs of yeast RAD23. Genomics 1995, 23, 651–658. [Google Scholar]
- Patel, S.A.; Simon, M.C. Functional analysis of the Cdk7.cyclin H.Mat1 complex in mouse embryonic stem cells and embryos. J. Biol. Chem 2010, 285, 15587–15598. [Google Scholar]
- Boucas, J.; Riabinska, A.; Jokic, M.; Herter-Sprie, G.S.; Chen, S.; Hopker, K.; Reinhardt, H.C. Posttranscriptional regulation of gene expression-adding another layer of complexity to the DNA damage response. Front. Genet 2012, 3. [Google Scholar] [CrossRef]
- Iwanaga, K.; Sueoka, N.; Sato, A.; Hayashi, S.; Sueoka, E. Heterogeneous nuclear ribonucleoprotein B1 protein impairs DNA repair mediated through the inhibition of DNA-dependent protein kinase activity. Biochem. Biophys. Res. Commun 2005, 333, 888–895. [Google Scholar]
- Lee, S.Y.; Park, J.H.; Kim, S.; Park, E.J.; Yun, Y.; Kwon, J. A proteomics approach for the identification of nucleophosmin and heterogeneous nuclear ribonucleoprotein C1/C2 as chromatin binding proteins in response to DNA double-strand breaks. Biochem. J 2005, 388, 7–15. [Google Scholar]
- Haley, B.; Paunesku, T.; Protić, M.; Woloschak, G.E. Response of heterogeneous ribonuclear proteins (hnRNP) to ionising radiation and their involvement in DNA damage repair. Int. J. Radiat. Biol 2009, 85, 643–655. [Google Scholar]
- Cok, S.J.; Acton, S.J.; Sexton, A.E.; Morrison, A.R. Identification of RNA-binding proteins in RAW 264.7 cells that recognize a lipopolysaccharide-responsive element in the 3-untranslated region of the murine cyclooxygenase-2 mRNA. J. Biol. Chem 2004, 279, 8196–8205. [Google Scholar]
- Chabot, B.; Blanchette, M.; Lapierre, I.; La Branche, H. An intron element modulating 5′ splice site selection in the hnRNP A1 pre-mRNA interacts with hnRNP A1. Mol. Cell. Biol 1997, 17, 1776–1786. [Google Scholar]
- Mazan-Mamczarz, K.; Kuwano, Y.; Zhan, M.; White, E.J.; Martindale, J.L.; Lal, A.; Gorospe, M. Identification of a signature motif in target mRNAs of RNA-binding protein AUF1. Nucleic Acids Res 2009, 37, 204–214. [Google Scholar]
- Farny, N.G.; Hurt, J.A.; Silver, P.A. Definition of global and transcript-specific mRNA export pathways in metazoans. Genes Dev 2008, 22, 66–78. [Google Scholar]
- Pieper, D.; Schirmer, S.; Prechtel, A.T.; Kehlenbach, R.H.; Hauber, J.; Chemnitz, J. Functional characterization of the HuR:CD83 mRNA interaction. PLoS One 2011, 6, e23290. [Google Scholar]
- Zhang, J.; Modi, Y.; Yarovinsky, T.; Yu, J.; Colling, M.; Kyriakidis, T.; Zhu, Y.; Sessa, W.C.; Pardi, R.; Bender, J.R. Macrophage b2 integrin-mediated, HuR-dependent stabilization of angiogenic factor-encoding mRNAs in inflammatory angiogenesis. Am. J. Pathol 2012, 180, 1751–1780. [Google Scholar]
- Brodsky, A.S.; Silver, P.A. Pre-mRNA processing factors are required for nuclear export. RNA 2000, 6, 1737–1749. [Google Scholar]
- Izquierdo, J.M. Heterogeneous ribonucleoprotein C displays a repressor activity mediated by T-cell intracellular antigen-1-related/like protein to modulate Fas exon 6 splicing through a mechanism involving Hu antigen R. Nucleic Acids Res 2010, 38, 8001–8014. [Google Scholar]
- Wang, J.; Wang, B.; Bi, J.; Zhang, C. Cytoplasmic HuR expression correlates with angiogenesis, lymphangiogenesis, and poor outcome in lung cancer. Med. Oncol 2011, 28, S577–S585. [Google Scholar]
- Baroni, T.E.; Lastro, M.T.; Ranganathan, A.C.; Tenenbaum, S.A.; Conklin, D.S.; Aguirre-Ghiso, J.A. Ribonomic and short hairpin RNA gene silencing methods to explore functional gene programs associated with tumor growth arrest. Methods Mol. Biol 2007, 383, 227–244. [Google Scholar]
- Chatziioannou, A.; Moulos, P.; Kolisis, F.N. Gene ARMADA: An integrated multi-analysis platform for microarray data implemented in MATLAB. BMC Bioinforma 2009, 10. [Google Scholar] [CrossRef]
- Chatziioannou, A.A.; Moulos, P. Exploiting statistical methodologies and controlled vocabularies for prioritized functional analysis of genomic experiments: The StRAnGER web application. Front. Neurosci 2011, 5. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Papadodima, O.; Chatziioannou, A.; Patrinou-Georgoula, M.; Kolisis, F.N.; Pletsa, V.; Guialis, A. HuR-Regulated mRNAs Associated with Nuclear hnRNP A1-RNP Complexes. Int. J. Mol. Sci. 2013, 14, 20256-20281. https://doi.org/10.3390/ijms141020256
Papadodima O, Chatziioannou A, Patrinou-Georgoula M, Kolisis FN, Pletsa V, Guialis A. HuR-Regulated mRNAs Associated with Nuclear hnRNP A1-RNP Complexes. International Journal of Molecular Sciences. 2013; 14(10):20256-20281. https://doi.org/10.3390/ijms141020256
Chicago/Turabian StylePapadodima, Olga, Aristotelis Chatziioannou, Meropi Patrinou-Georgoula, Fragiskos N. Kolisis, Vasiliki Pletsa, and Apostolia Guialis. 2013. "HuR-Regulated mRNAs Associated with Nuclear hnRNP A1-RNP Complexes" International Journal of Molecular Sciences 14, no. 10: 20256-20281. https://doi.org/10.3390/ijms141020256