Neuroprotective Effects of Ultra-Low-Molecular-Weight Heparin on Cerebral Ischemia/Reperfusion Injury in Rats: Involvement of Apoptosis, Inflammatory Reaction and Energy Metabolism

Abstract
:1. Introduction
2. Results and Discussion
2.1. Effects of ULMWH on ATPase Activity and LD Content in Brain of Ischemic Cortex
2.2. Quantification of Apoptosis by Flow Cytometry
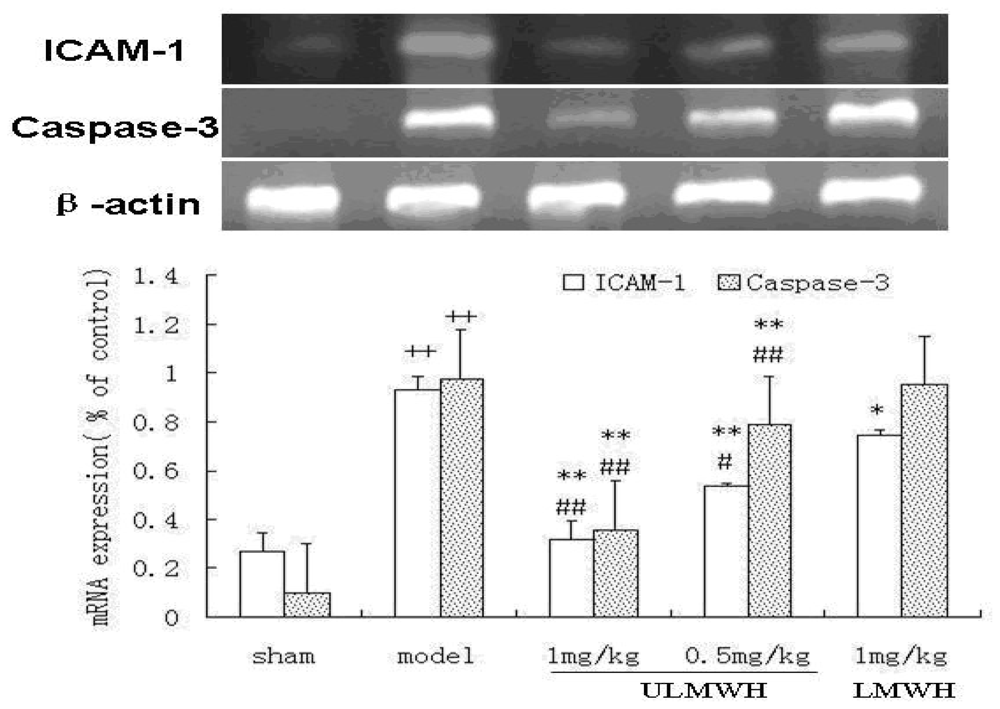
2.3. Semi-Quantitative RT-PCR of ICAM-1 and Caspase-3
3. Experimental Section
3.1. Chemicals
3.2. MCAO-Induced Transient Focal Cerebral Ischemia in Rats
3.3. Drug Treatment
3.4. The ATPase Activity and Lactic Acid Level in Brain Tissue after Injury
3.5. Quantification of Apoptosis by Flow Cytometry
3.6. Semi-Quantitative RT-PCR of ICAM-1 and Caspase-3
3.7. Statistical Analysis
4. Conclusions
- Conflict of InterestThe authors declare no conflict of interest.
References
- Kao, T.K.; Qu, Y.C.; Kuo, J.S.; Chen, W.Y.; Liao, S.L.; Wu, C.W.; Chen, C.J.; Ling, N.N.; Zhang, Y.H.; Peng, W.H. Neuroprotection by tetramethylpyrazine against ischemic brain injury in rats. Neurochem. Int 2006, 48, 166–176. [Google Scholar]
- Hao, L.N.; Zhang, Q.Z.; Yu, T.G.; Cheng, Y.N.; Ji, S.L. Antagonistic effects of ultra-low-molecular-weight heparin on Aβ25–35-induced apoptosis in cultured rat cortical neurons. Brain Res 2011, 1368, 1–10. [Google Scholar]
- Yu, T.G.; Zhang, Q.Z.; Zhang, Z.G.; Wang, W.W.; Ji, S.L.; Du, G.H. Protective Effect of Ultra Low Molecular Weight Heparin on Glutamate-Induced Apoptosis in Cortical Cells. Yonsei Med. J 2008, 49, 486–495. [Google Scholar]
- Zhang, Z.G.; Zhang, Q.Z.; Cheng, Y.N.; Ji, S.L.; Du, G.H. Antagonistic effects of Ultra-Low-Molecular-Weight Heparin against cerebral ischemia/reperfusion injury in rats. Pharmacol. Res 2007, 56, 350–355. [Google Scholar]
- Tian, J.W.; Fu, F.H.; Geng, M.Y.; Jiang, Y.T.; Yang, J.X.; Jiang, W.; Wang, C.; Liu, K. Neuroprotective effect of 20(S)-ginsenoside Rg3 on cerebral ischemia in rats. Neurosci. Lett 2005, 374, 92–97. [Google Scholar]
- Yue, F.; Zhang, W.; Guo, J. The alterations of brain lactate, lactate dehydrogenase, creatine phosphokinase and its influence on these of peripheral blood or liver tissue and entero-barrier during brain hypoperfusion. Chin. J. Pathophysiol 1999, 15, 1106–1119. [Google Scholar]
- Hoxworth, J.M.; Xu, K.; Zhou, Y.; Lust, W.D.; LaManna, J.C. Cerebral metabolic profile, selective neuron loss, and survival of acute and chronic hyperglycemic rats following cardiac arrest and resuscitation. Brain Res 1999, 821, 467–479. [Google Scholar]
- Kirino, T. Delayed neuronal death. Neuropathology 2000, 20, S95–S97. [Google Scholar]
- Zhang, W.R.; Hayashi, T.; Sasaki, C.; Sato, K.; Nagano, I. Attenuation of oxidative DNA damage with a novel antioxidant EPC-K1 in rat brain neuronal cells after transient middle cerebral artery occlusion. Neurol. Res 2001, 23, 676–680. [Google Scholar]
- Khan, M.; Sekhon, B.; Jatana, M.; Giri, S.; Gilg, A.G. Administration of N-acetylcysteine after focal cerebral ischemia protects brain and reduces inflammation in a rat model of experimental stroke. J. Neurosci. Res 2004, 76, 519–527. [Google Scholar]
- Yamada, A.; Isono, M.; Hori, S.; Shimomura, T.; Nakano, T. Temporal and spatial profile of apoptotic cells after focal cerebral ischemia in rats. Neurol. Med. Chir 1999, 39, 575–583. [Google Scholar]
- Hu, X.; Johansson, I.M.; Brannstrom, T.; Olsson, T.; Wester, P. Long-lasting neuronal apoptotic cell death in regions with severe ischemia after photothrombotic ring stroke in rats. Acta Neuropathol 2002, 104, 462–470. [Google Scholar]
- Jean, W.C.; Spellman, S.R.; Nussbaum, E.S.; Low, W.C. Reperfusion injury after focal cerebral ischemia: role of inflammation and the therapeutic horizon. Neurosurgery 1998, 43, 1382–1397. [Google Scholar]
- Liu, S.J.; Zhou, S.W.; Xue, C.S. Effect of tetrandrine on neutrophilic recruitment response to brain ischemia/reperfusion. Acta Pharmacol. Sin 2001, 22, 971–975. [Google Scholar]
- Bowes, M.P.; Zivin, J.A.; Rothlein, R. Monoclonal antibody to the ICAM-1 adhesion site reduces neurological damage in a rabbit cerebral embolism stroke model. Exp. Neurol 1993, 119, 215–219. [Google Scholar]
- Chopp, M.; Li, Y.; Jiang, N.; Zhang, R.L.; Prostak, J. Antibodies against adhesion molecules reduce apoptosis after transient middle cerebral artery occlusion in rat brain. J. Cereb. Blood Flow Metab 1996, 16, 578–584. [Google Scholar]
- Kanemoto, Y.; Nakase, H.; Akita, N.; Sakaki, T. Effects of antiintercellular adhesion molecule-1 antibody on reperfusion injury induced by late reperfusion in the rat middle cerebral artery occlusion model. Neurosurgery 2002, 51, 1034–1041. [Google Scholar]
- Vemuganti, R.; Dempsey, R.J.; Bowen, K.K. Inhibition of intercellular adhesion molecule-1 protein expression by antisense oligonucleotides is neuroprotective after transient middle cerebral artery occlusion in rat. Stroke 2004, 35, 179–184. [Google Scholar]
- Connolly, E.S., Jr; Winfree, C.J.; Springer, T.A.; Naka, Y.; Liao, H. Cerebral protection in homozygous null ICAM-1 mice after middle cerebral artery occlusion. Role of neutrophil adhesion in the pathogenesis of stroke. J. Clin. Invest. 1996, 97, 209–216. [Google Scholar]
- Kitagawa, K.; Matsumoto, M.; Mabuchi, T.; Yagita, Y.; Ohtsuki, T. Deficiency of intercellular adhesion molecule 1 attenuates microcirculatory disturbance and infarction size in focal cerebral ischemia. J. Cereb. Blood Flow Metab 1998, 18, 1336–1345. [Google Scholar]
- Wu, G.D.; Zhou, H.J.; Wu, X.H. Apoptosis of human umbilical vein endothelial cells induced by artesunate. Vascul. Pharmacol 2004, 41, 205–212. [Google Scholar]
- Chen, P.; Scicli, G.M.; Guo, M.; Fenstermacher, J.D.; Dahl, D.; Edwards, P.A.; Scicli, A.G. Role of angiotensin II in retinal leukostasis in the diabetic rat. Exp. Eye Res 2006, 83, 1041–1051. [Google Scholar]
- Shimamura, M.; Garcia, J.M.; Prough, D.S.; Dewitt, D.S.; Uchida, T.; Shah, S.A.; Avila, M.A.; Hellmich, H.L. Analysis of long-term gene expression in neurons of the hippocampal subfields following traumatic brain injury in rats. Neuroscience 2005, 131, 87–97. [Google Scholar]

{kind=link}
| Group | Dose (mg/kg) | Lactic acid (mmol/g) | ATPase (μmolPi/mg·h) | Percentage of cell apoptosis (%) |
|---|---|---|---|---|
| Sham | Saline | 0.75 ± 0.19 ** | 3.97 ± 0.60 ** | 5.6 ± 2.10 ** |
| Model | Saline | 1.06 ± 0.18 | 2.42 ± 0.67 | 20.3 ± 7.95 |
| ULMWH | 0.5 | 0.89 ± 0.13 * | 3.33 ± 0.57 * | 11.4 ± 3.70 * |
| ULMWH | 1 | 0.83 ± 0.09 ** | 3.49 ± 0.35 ** | 6.9 ± 2.32 ** |
| LMWH | 1 | 0.85 ± 0.13 * | 3.56 ± 0.57 ** | 10.6 ± 1.77 * |
© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, Z.-G.; Sun, X.; Zhang, Q.-Z.; Yang, H. Neuroprotective Effects of Ultra-Low-Molecular-Weight Heparin on Cerebral Ischemia/Reperfusion Injury in Rats: Involvement of Apoptosis, Inflammatory Reaction and Energy Metabolism. Int. J. Mol. Sci. 2013, 14, 1932-1939. https://doi.org/10.3390/ijms14011932
Zhang Z-G, Sun X, Zhang Q-Z, Yang H. Neuroprotective Effects of Ultra-Low-Molecular-Weight Heparin on Cerebral Ischemia/Reperfusion Injury in Rats: Involvement of Apoptosis, Inflammatory Reaction and Energy Metabolism. International Journal of Molecular Sciences. 2013; 14(1):1932-1939. https://doi.org/10.3390/ijms14011932
Chicago/Turabian StyleZhang, Zhi-Guo, Xin Sun, Qing-Zhu Zhang, and Hua Yang. 2013. "Neuroprotective Effects of Ultra-Low-Molecular-Weight Heparin on Cerebral Ischemia/Reperfusion Injury in Rats: Involvement of Apoptosis, Inflammatory Reaction and Energy Metabolism" International Journal of Molecular Sciences 14, no. 1: 1932-1939. https://doi.org/10.3390/ijms14011932