Combination of Oxyanion Gln114 Mutation and Medium Engineering to Influence the Enantioselectivity of Thermophilic Lipase from Geobacillus zalihae


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
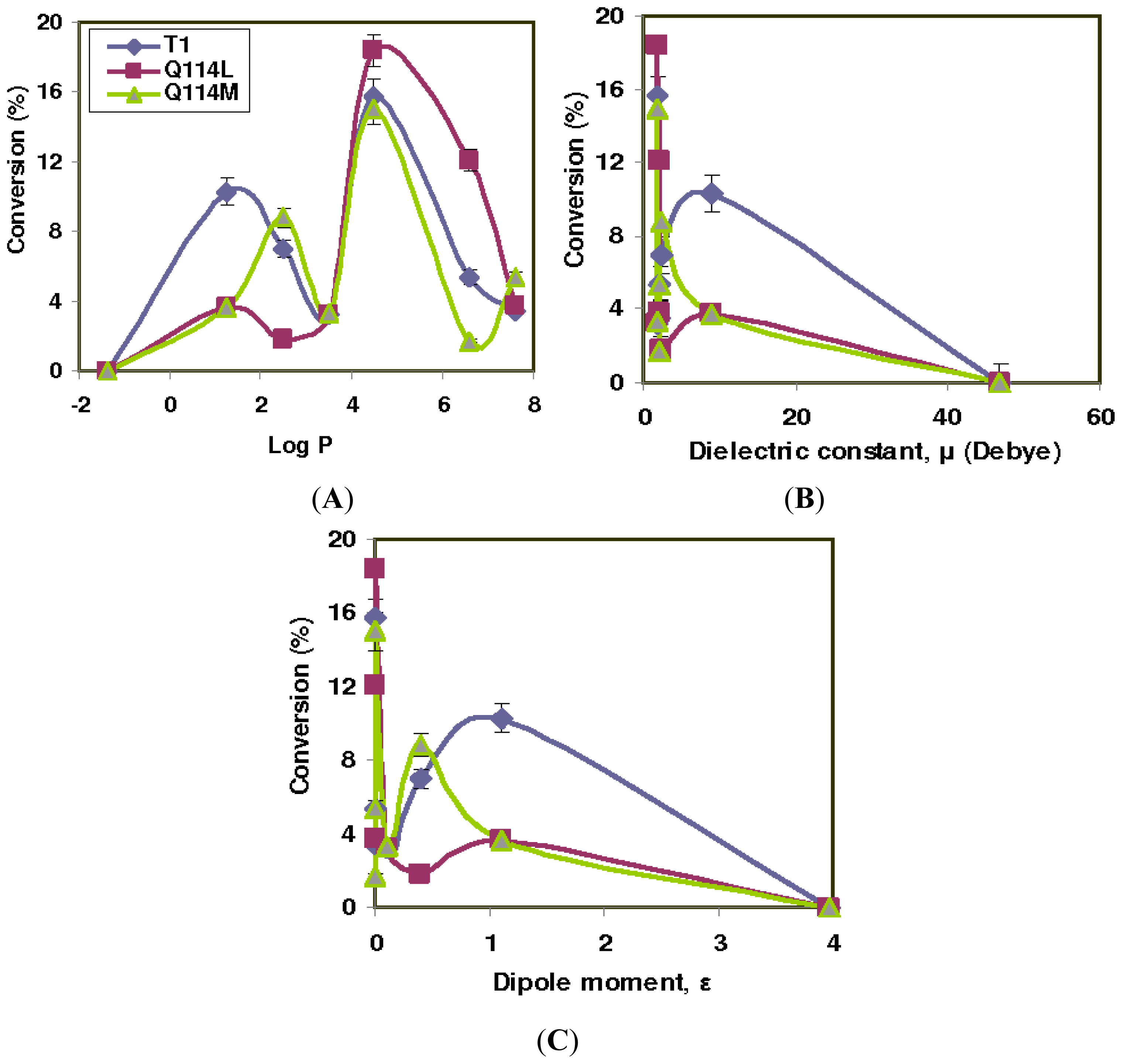
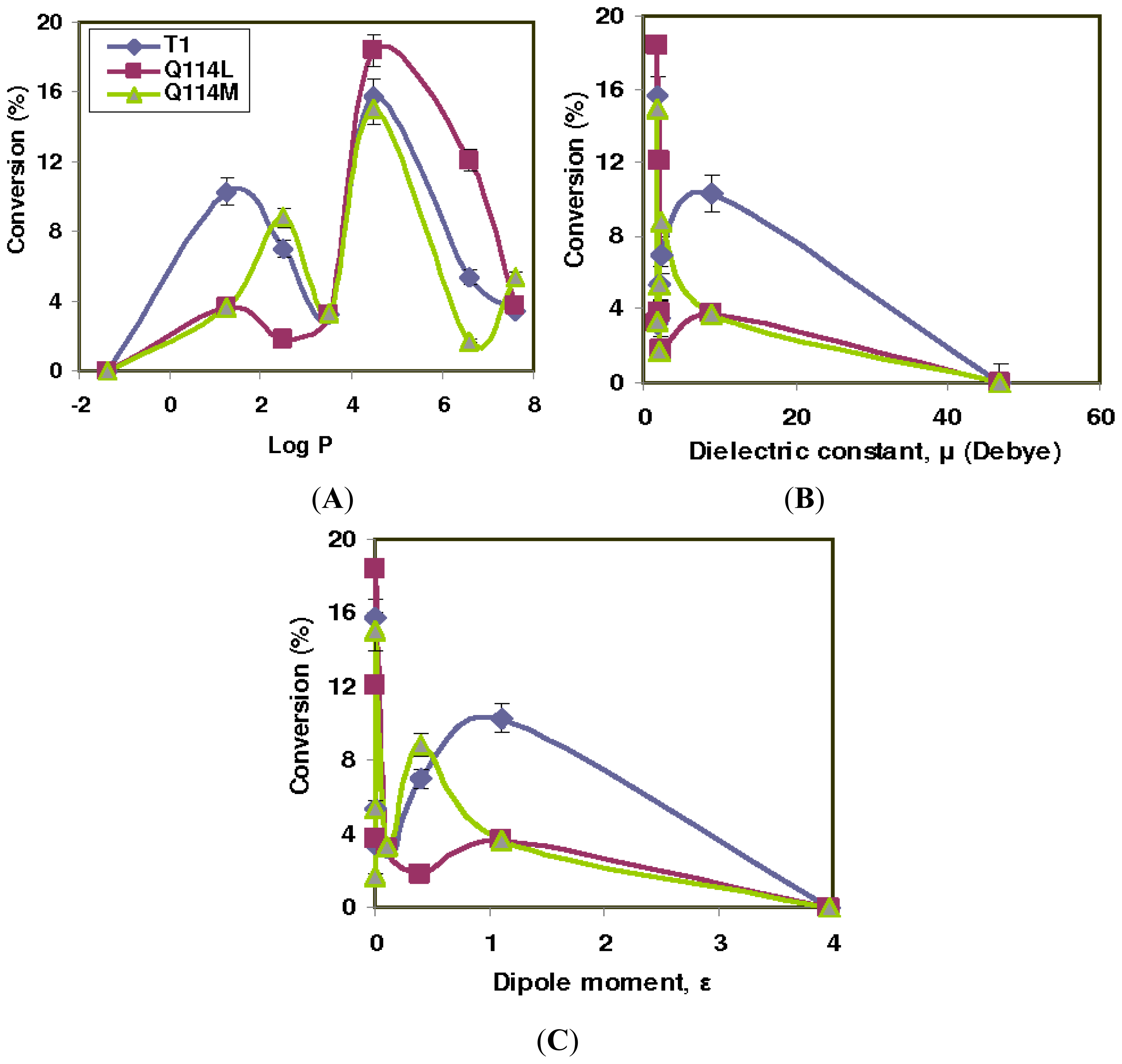
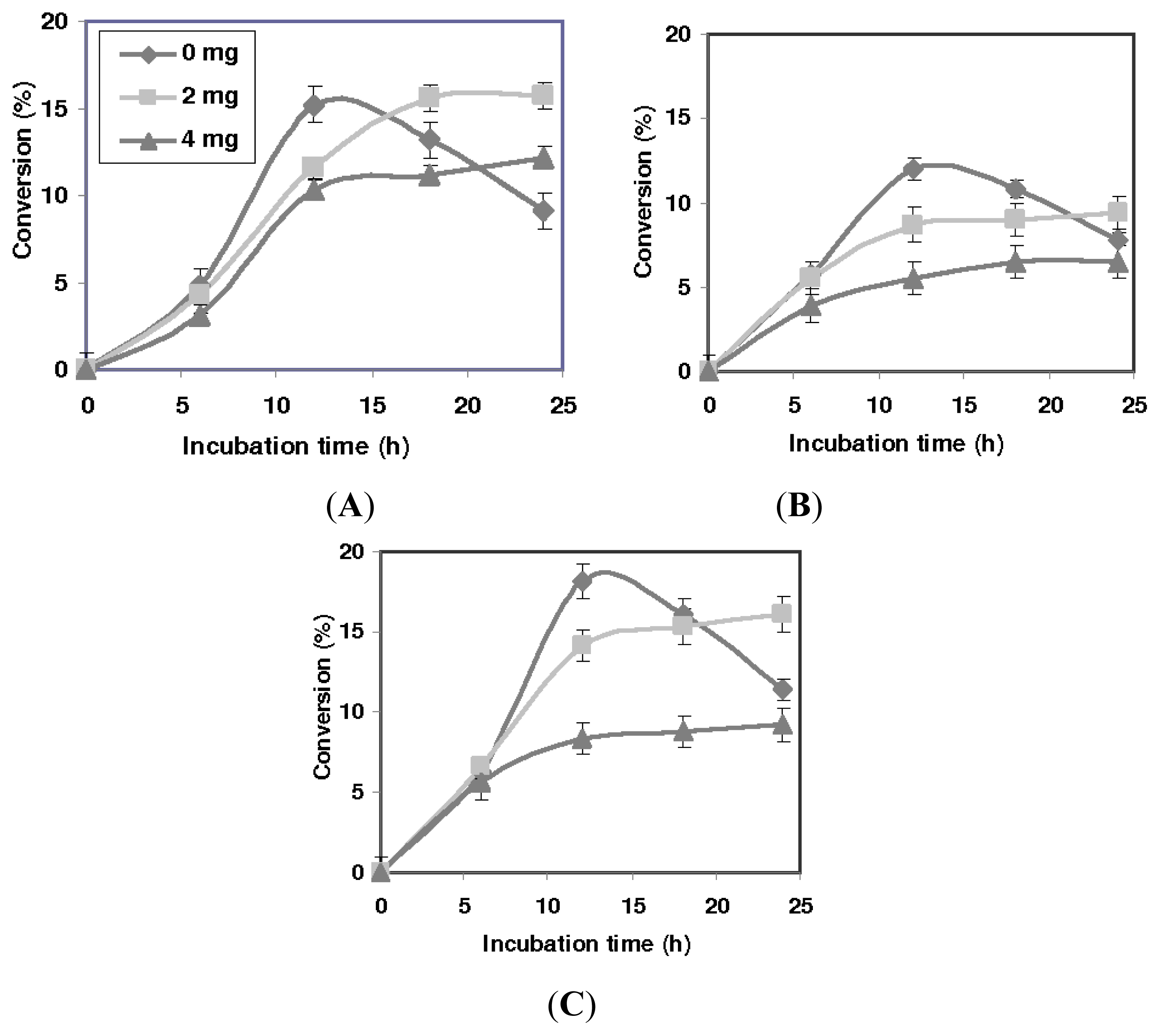
2.1. Effect of Solvents
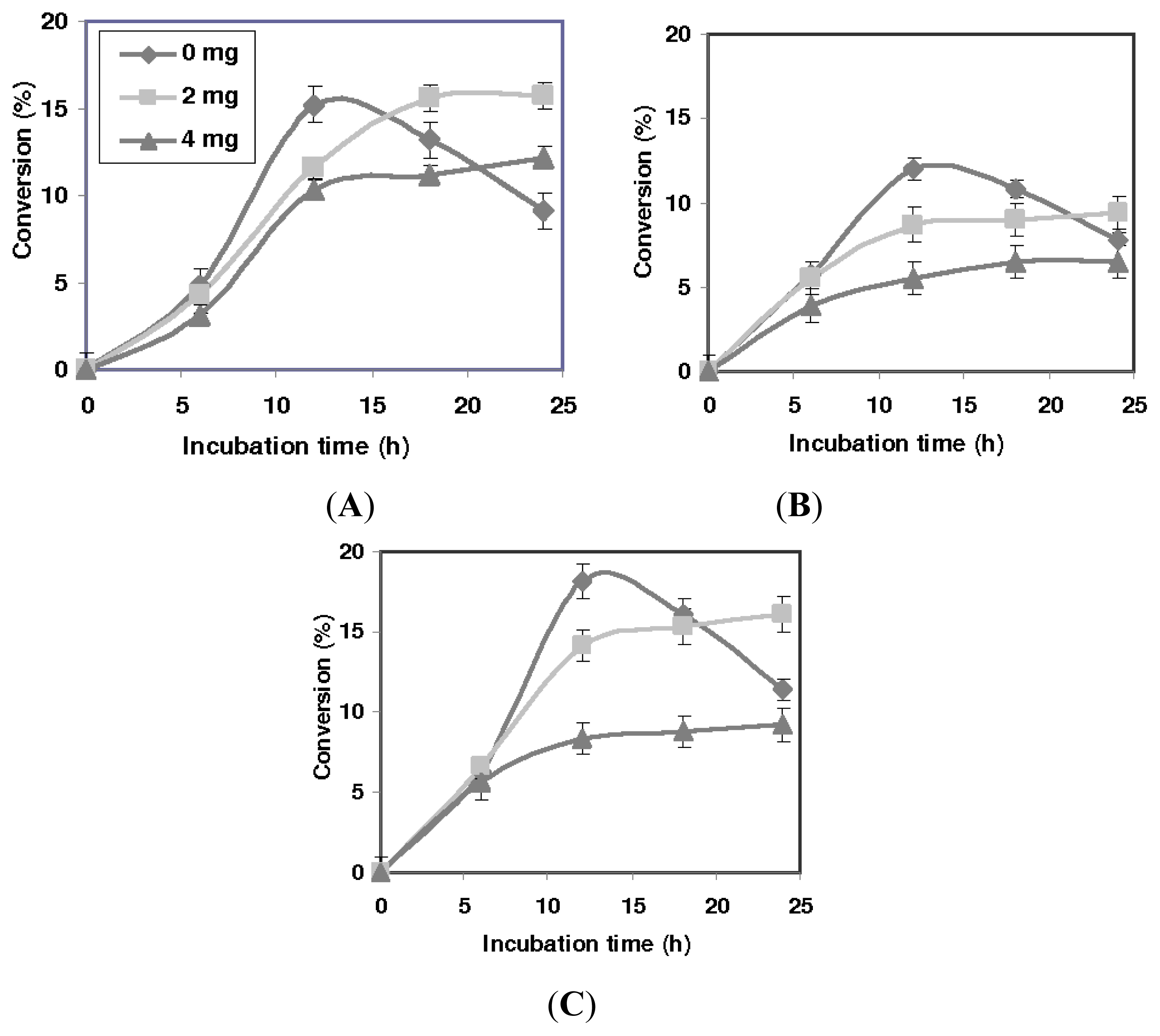
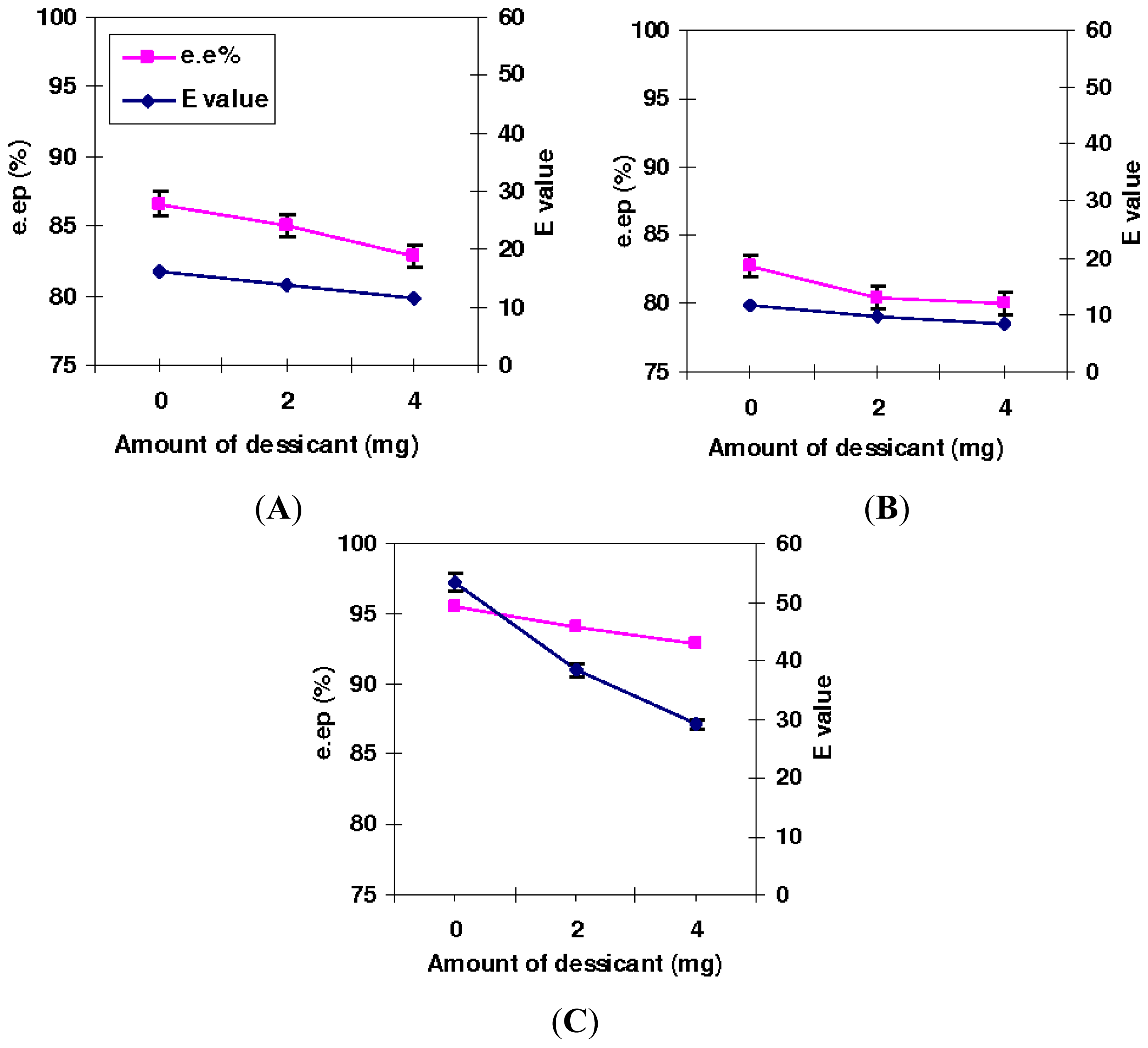
2.2. Effect of the Presence of Desiccant
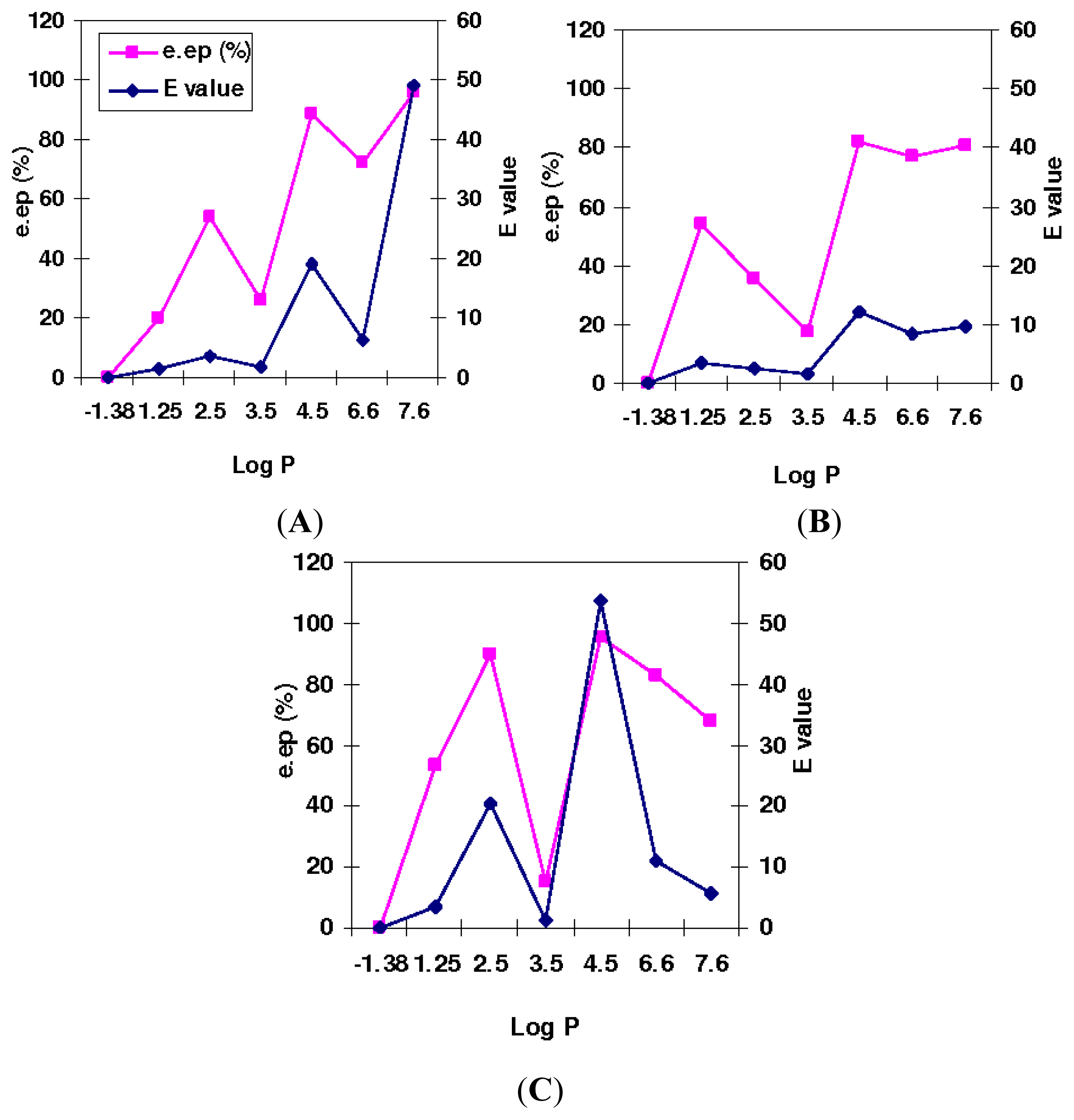
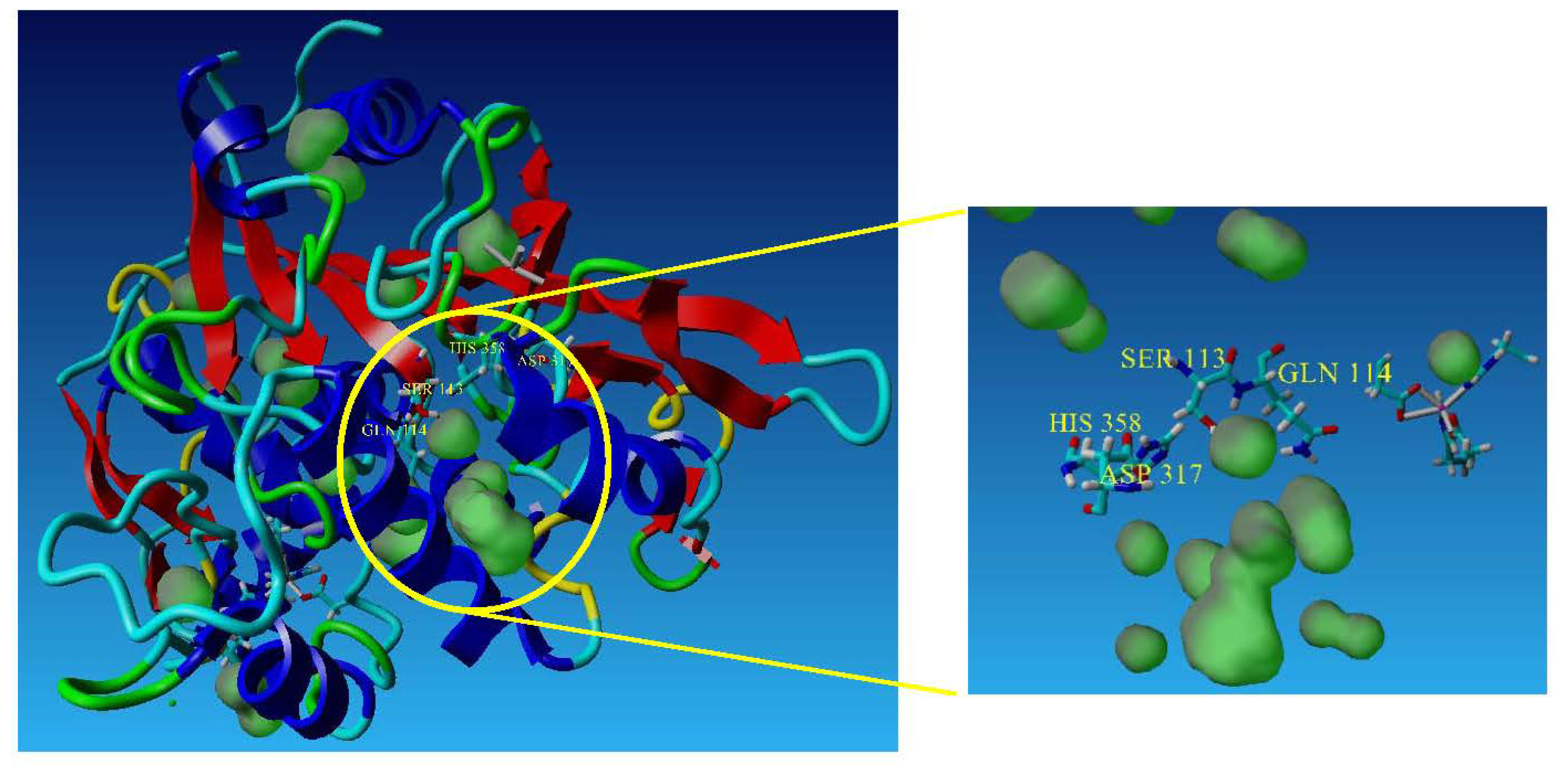
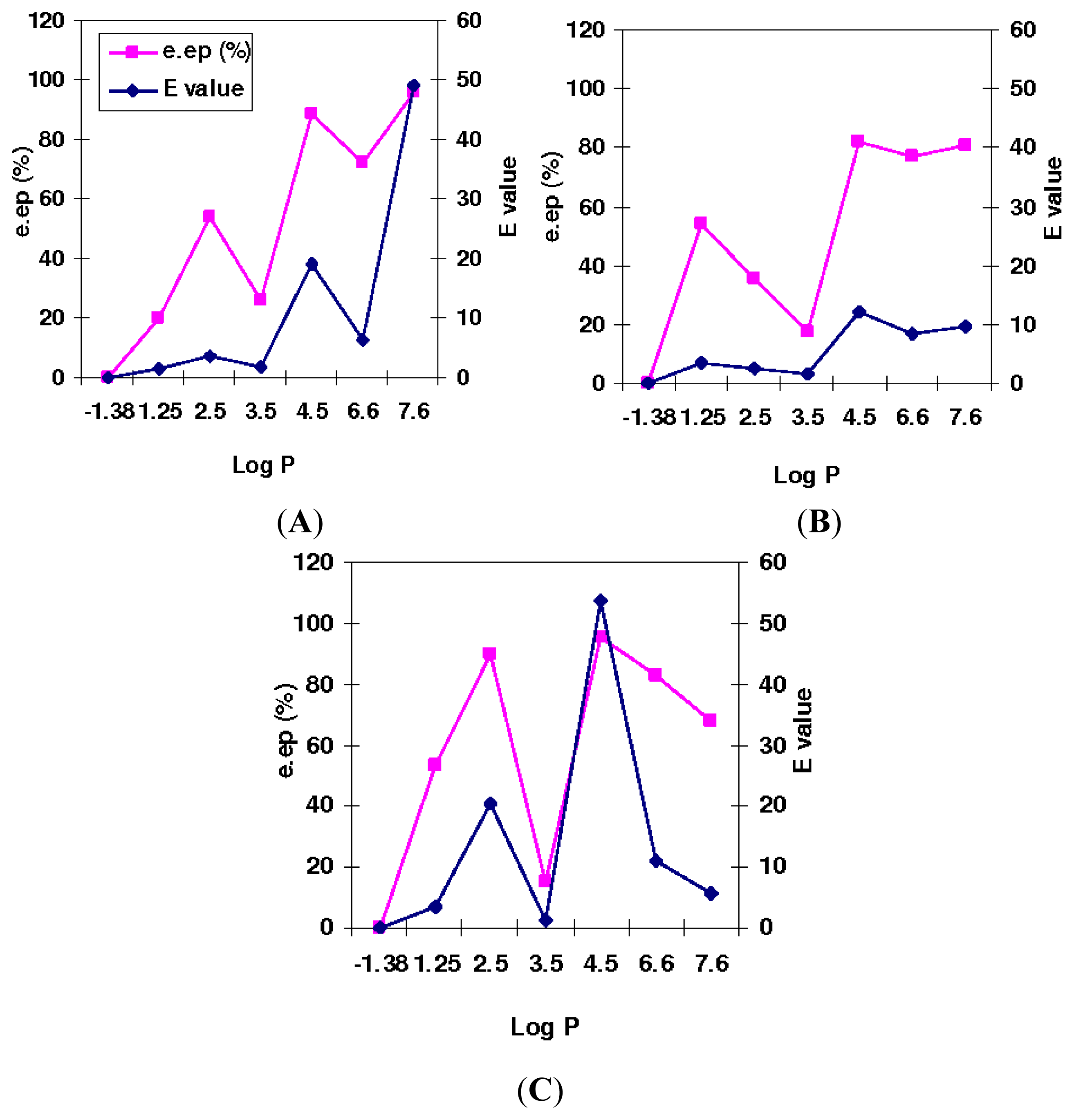
2.3. The Effect of Mutation on Enantioselectivity
3. Experimental Section
3.1. Materials
3.2. Substitution of Residues by Site-Directed Mutagenesis
3.3. Preparation of Stock and Working Culture
3.4. Partial Purification of Enzymes
3.5. Standard Lipase Assay and Protein Concentration
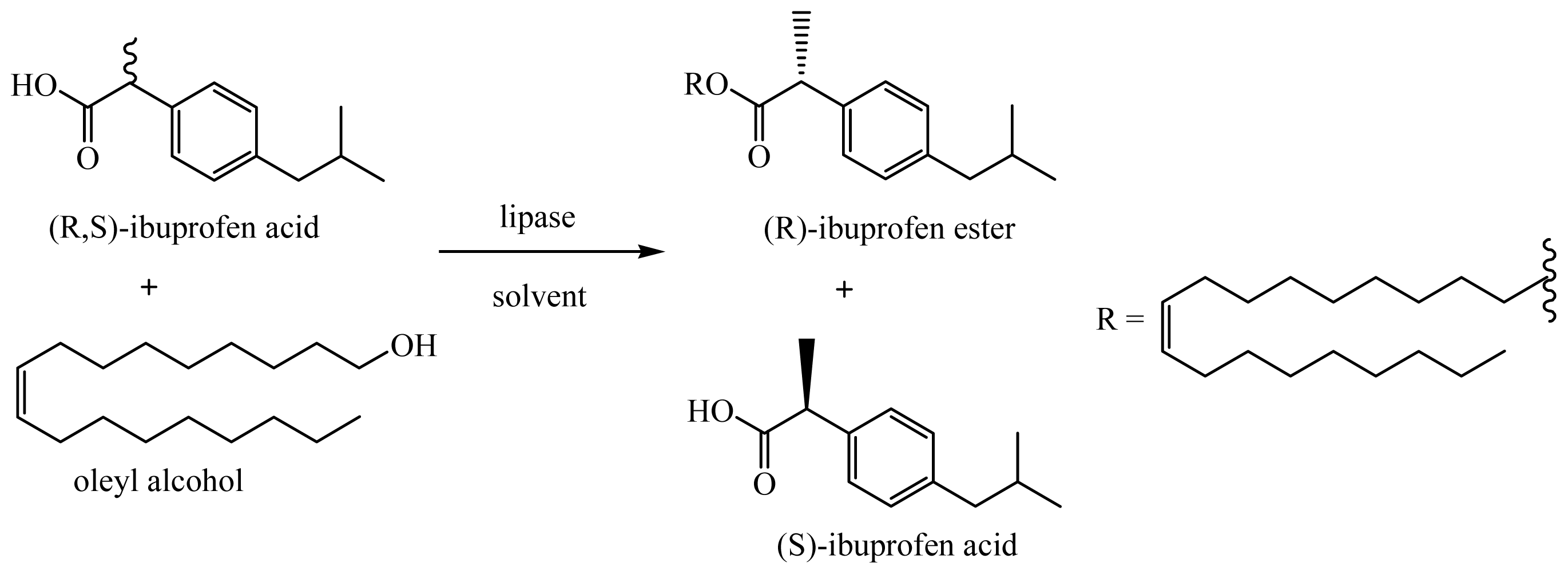
3.6. Enantioselective Esterification of (R-S)-Ibuprofen with Oleyl Alcohol
Effect of Reaction Conditions
3.7. Analysis and Determination of Ibuprofen Esters
4. Conclusions
Acknowledgments
References
- Forde, J.; Tully, E.; Vakurov, A.; Gibson, T.D.; Millner, P.; O’Fagain, C. Chemical modification and immobilization of laccase from Trametes hirsuta and from Myceliophthora thermophila. Enzym. Microb. Technol 2010, 46, 430–437. [Google Scholar]
- Cong, F.; Xing, K.; Gao, R.-J.; Cao, S.-G.; Zhang, G. Enhanced activity and enantioselectivity of a hyperthermophilic esterase from archeon Aeropyrum pernix K1 by acetone treatment. Appl. Biochem. Biotechnol 2011, 165, 795–801. [Google Scholar]
- Persson, M.; Costes, D.; Wehtje, E.; Adlercreutz, P. Preparation of lipases for use in organic solvents. Enzym. Microb. Technol 2002, 31, 916–923. [Google Scholar]
- Ghanem, A. Trends in lipase-catalyzed asymmetric access to enantiomerically pure/enriched compounds. Tetrahedron 2007, 63, 1721–1754. [Google Scholar]
- Overbeeke, P.L.A.; Jongejan, J.A.; Heijnen, J.J. Solvent effect on lipase enantioselectivity. Evidence for the presence of two thermodynamic states. Biotechnol. Bioeng 2000, 70, 278–290. [Google Scholar]
- Wang, Y.; Li, Q.; Zhang, Z.; Ma, J.; Feng, Y.J. Solvent effects on the enantioselectivity of the thermophilic lipase QLM in the resolution of (R,S)-2-octanol and (R,S)-2-pentanol. J. Mol. Catal. B 2009, 56, 146–150. [Google Scholar]
- Graber, M.; Irague, R.; Rosenfeld, E.; Lamare, S.; Franson, L.; Hult, K. Solvent as a competitive inhibitor for Candida antarctica lipase B. Biochim. Biophys. Acta 2007, 1774, 1052–1057. [Google Scholar]
- Wang, Z.; Feng, Y.; Cao, S.G. The effect of microwater on the enzyme-catalyzed reaction in organic media and its controlling methods. Progr. Natl. Sci. (China) 2002, 12, 130–134. [Google Scholar]
- Yu, S.S.; Yu, S.C.; Han, W.W.; Wang, H.L.; Zheng, B.S.; Feng, Y.J. A novel thermophilic lipase from Fervidobacterium nodosum Rt17-B1 representing a new subfamily of bacterial lipases. J. Mol. Catal. B 2010, 66, 81–89. [Google Scholar]
- Leow, T.C.; Rahman, R.N.Z.; Basri, M.; Salleh, A.B. High level expression of thermostable lipase from Geobacillus sp. strain T1. Biol. Biotechnol. Biochem 2004, 68, 96–103. [Google Scholar]
- Leow, T.C. Thermostable Lipase: Isolation, Gene Analysis, Expression, Characterization and Structure Elucidation. Ph.D. Thesis, Universiti Putra Malaysia, Serdang, Malaysia, October 2005. [Google Scholar]
- Carrasco-Lopez, C.; Godoy, C.; de las Rivas, G.; de las Rivas, B.; Fernandez-Lorente, G.; Palomo, J.M.; Guisan, J.M.; Fernandez-Lafuente, R.; Martinez-Ripoll, M.; Hermoso, J.A. Activation of bacterial thermoalkalophilic lipases is spurred by dramatic structural rearrangements. J. Biol. Chem 2009, 284, 4365–4372. [Google Scholar]
- Morley, K.L.; Kazlauskas, R.J. Improving enzyme properties: When are closer mutations better? Trends Biotechnol 2005, 23, 231–237. [Google Scholar]
- Abdul Rahman, M.Z.; Salleh, A.B.; Raja Abdul Rahman, R.N.Z.; Abdul Rahman, M.B.; Basri, M.; Leow, T.C. Unlocking the mystery behind the activation phenomenon of T1 lipase: A molecular dynamics simulations approach. Protein Sci 2012, 21, 1210–1221. [Google Scholar]
- Klibanov, A. Improving enzymes by using them in organic solvents. Process Biochem 2001, 43, 1259–1264. [Google Scholar]
- Wen, S.; Tan, T.; Yu, M. Immobilised lipase YILip2-catalyzed resolution of (±)-phenylethylamine in a medium with organic solvent. Process Biochem 2008, 43, 1259–1264. [Google Scholar]
- Pilissido, C.; de Oliveira, C.; da Graca, N.M. Enantioselective acylation of (R,S)-phenylethylamine catalyzed by lipases. Process Biochem 2009, 44, 1352–1357. [Google Scholar]
- Fitzpatrick, P.A.; Klibanov, A.M. How can the solvent affect enzyme enantioselectivity. J. Am. Chem. Soc 1991, 113, 3166–3171. [Google Scholar]
- Ghanem, A.; Aboul-Enein, H.Y. Lipase-Mediated chiral resolution by racemates in organic solvents. Tetrahedron Asymmetry 2004, 15, 3331–3351. [Google Scholar]
- Zhao, D.; Xun, E.; Wang, J.; Wang, R.; We, X.; Wang, H.; Wang, Z. Enantioselective esterification of ibuprofen by a novel thermophilic biocatalyst: APE1547. Biotechnol. Bioprocess Eng 2011, 16, 638–644. [Google Scholar]
- Gorman, L.A.S.; Dordick, J.S. Organic solvents strips water off enzymes. Biotechnol. Bioeng 1992, 39, 392–397. [Google Scholar]
- Jongejan, J.A. Effects of Organic Solvents on Enzyme Selectivity. In Organic Synthesis with Enzymes in Non-Aqueous Media; Wiley: Weinheim, Germany, 2008; pp. 50–51. [Google Scholar]
- Cheong, K.W.; Leow, T.C.; Rahman, R.N.Z.; Basri, M.; Rahman, M.B.; Salleh, A.B. Reductive alkylation causes the formation of a molten globule-like intermediate structure in Geobacillus zalihae T1 thermostable lipase. Appl. Biochem. Biotechnol 2011, 164, 362–375. [Google Scholar]
- Liu, Z.-Q.; Zheng, X.-B.; Zhang, S.-P.; Zheng, Y.-G. Cloning, expression and characterization of a lipase gene from the Candida antarctica ZJB09193 and its application in biosynthesis of vitamin A esters. Microbiol. Rev 2012, 167, 452–460. [Google Scholar]
- Ong, A.L.; Kamaruddin, A.H.; Long, W.S.; Lim, S.T.; Kumari, R. Performance of free Candida antarctica lipase B in the enantioselective esterification of (R)-ketoprofen. Enzym. Microb. Technol 2006, 39, 924–929. [Google Scholar]
- Serdakowski, A.L.; Dordick, J.S. Enzyme activation for organic solvents made easy. Trends Biotechnol 2008, 25, 48–54. [Google Scholar]
- Wang, Y.; Wei, D.Q.; Wang, J.F. Molecular dynamics studies on T1 lipase: Insight into a double-flap mechanism. J. Chem. Inf. Model 2010, 24, 875–878. [Google Scholar]
- Antona, N.D.; Lombardi, P.; Nicolosi, G.; Salvo, G. Large scale preparation of enantiopure S-ketoprofen by biocatalyzed kinetic resolution. Process Biochem 2002, 38, 373–377. [Google Scholar]
- Tawaki, S.; Klibanov, A.M. Inversion of enzyme enantioselectivity mediated by solvent. J. Am. Chem. Soc 1992, 114, 1882–1884. [Google Scholar]
- Ducret, A.; Trani, M.; Lortie, R. Lipase-Catalyzed enantioselective esterification of ibuprofen in organic solvents under controlled water activity. Enzym. Microb. Technol 1998, 22, 212–216. [Google Scholar]
- Zhang, G.; Gao, R.; Zheng, L.; Zhang, A.; Wang, Y.; Wang, Q.; Feng, Y.; Cao, S. Study of the relationship between structure and enantioselectivity of a hyperthermophilic esterase from Archeon aeropyrum pernix K1. J. Mol. Catal. B 2006, 38, 148–153. [Google Scholar]
- Broos, J. Impact of the enzyme flexibility on the enantioselectivity in organic media towards specific and non-specific substrates. Biocatal. Biotrans 2002, 20, 291–295. [Google Scholar]
- Nayeem, A.; Chiang, S.J.; Liu, S.W.; Sun, Y.; You, L.; Basch, J. Engineering enzymes for improved catalytic efficiency: A computational study of site mutagenesis in epothilone-B hydroxylase. Protein Eng. Des. Sel 2009, 22, 257–266. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Chen, C.S.; Fujimoto, Y.; Girdaukas, G.; Sih, C.J. Quantitative analyses of biochemical kinetic resolutions of enantiomers. J. Am. Chem. Soc 1982, 104, 7294–7299. [Google Scholar]



© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wahab, R.A.; Basri, M.; Rahman, M.B.A.; Rahman, R.N.Z.R.A.; Salleh, A.B.; Leow, T.C. Combination of Oxyanion Gln114 Mutation and Medium Engineering to Influence the Enantioselectivity of Thermophilic Lipase from Geobacillus zalihae. Int. J. Mol. Sci. 2012, 13, 11666-11680. https://doi.org/10.3390/ijms130911666
Wahab RA, Basri M, Rahman MBA, Rahman RNZRA, Salleh AB, Leow TC. Combination of Oxyanion Gln114 Mutation and Medium Engineering to Influence the Enantioselectivity of Thermophilic Lipase from Geobacillus zalihae. International Journal of Molecular Sciences. 2012; 13(9):11666-11680. https://doi.org/10.3390/ijms130911666
Chicago/Turabian StyleWahab, Roswanira Abdul, Mahiran Basri, Mohd Basyaruddin Abdul Rahman, Raja Noor Zaliha Raja Abdul Rahman, Abu Bakar Salleh, and Thean Chor Leow. 2012. "Combination of Oxyanion Gln114 Mutation and Medium Engineering to Influence the Enantioselectivity of Thermophilic Lipase from Geobacillus zalihae" International Journal of Molecular Sciences 13, no. 9: 11666-11680. https://doi.org/10.3390/ijms130911666