Ginsenoside Rg3 Reduces Lipid Accumulation with AMP-Activated Protein Kinase (AMPK) Activation in HepG2 Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
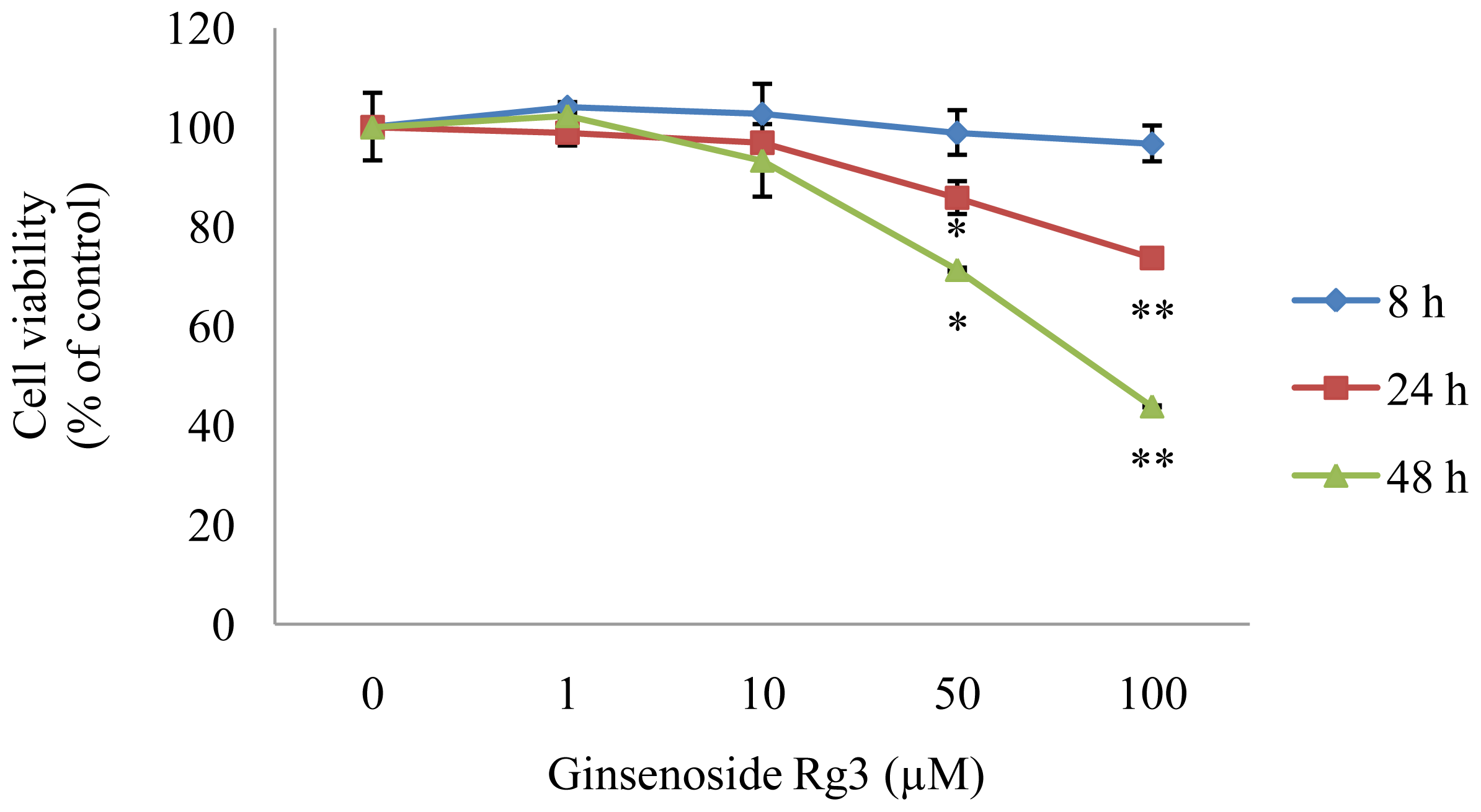
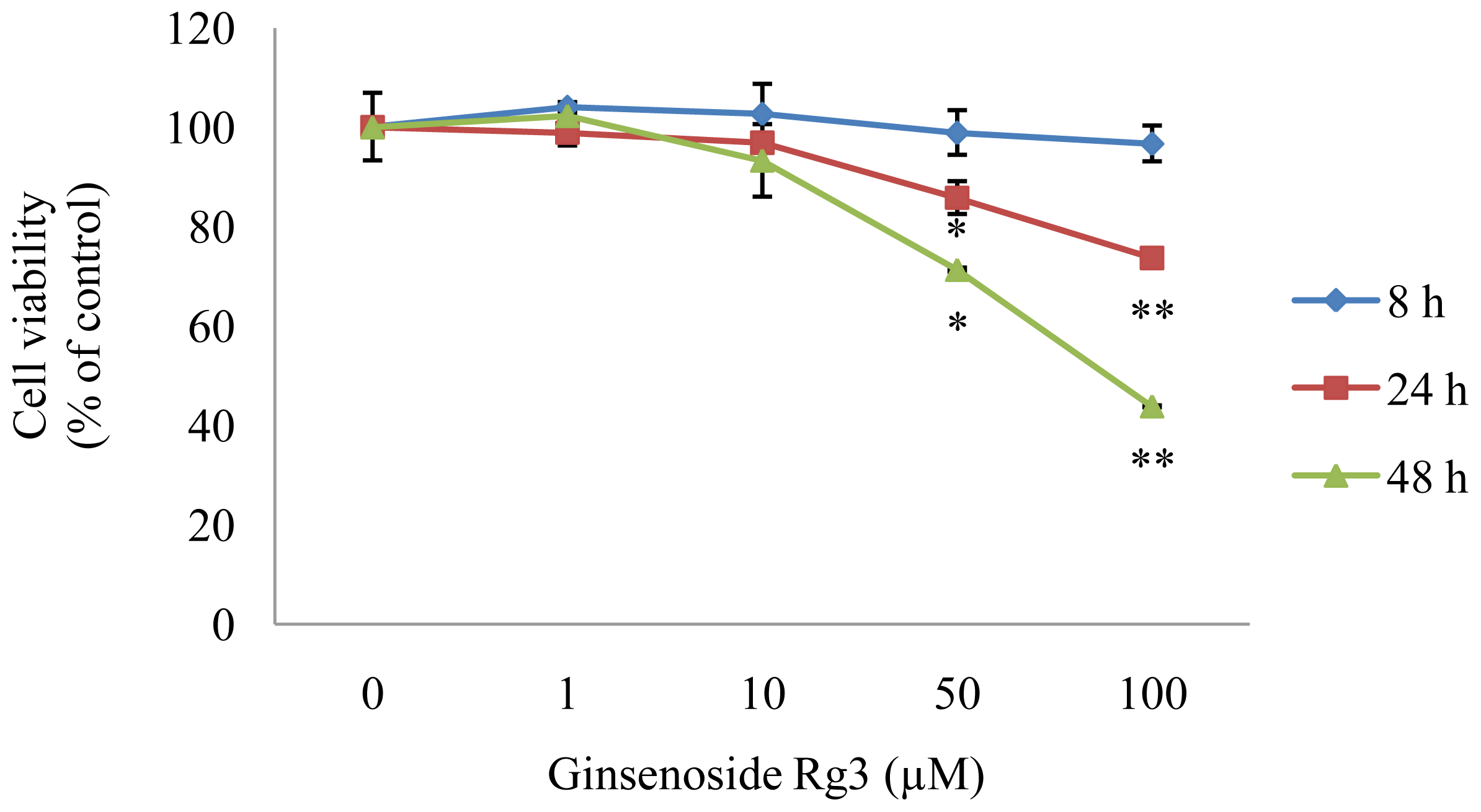
2.1. Effect of Ginsenoside Rg3 on HepG2 Cell Viability
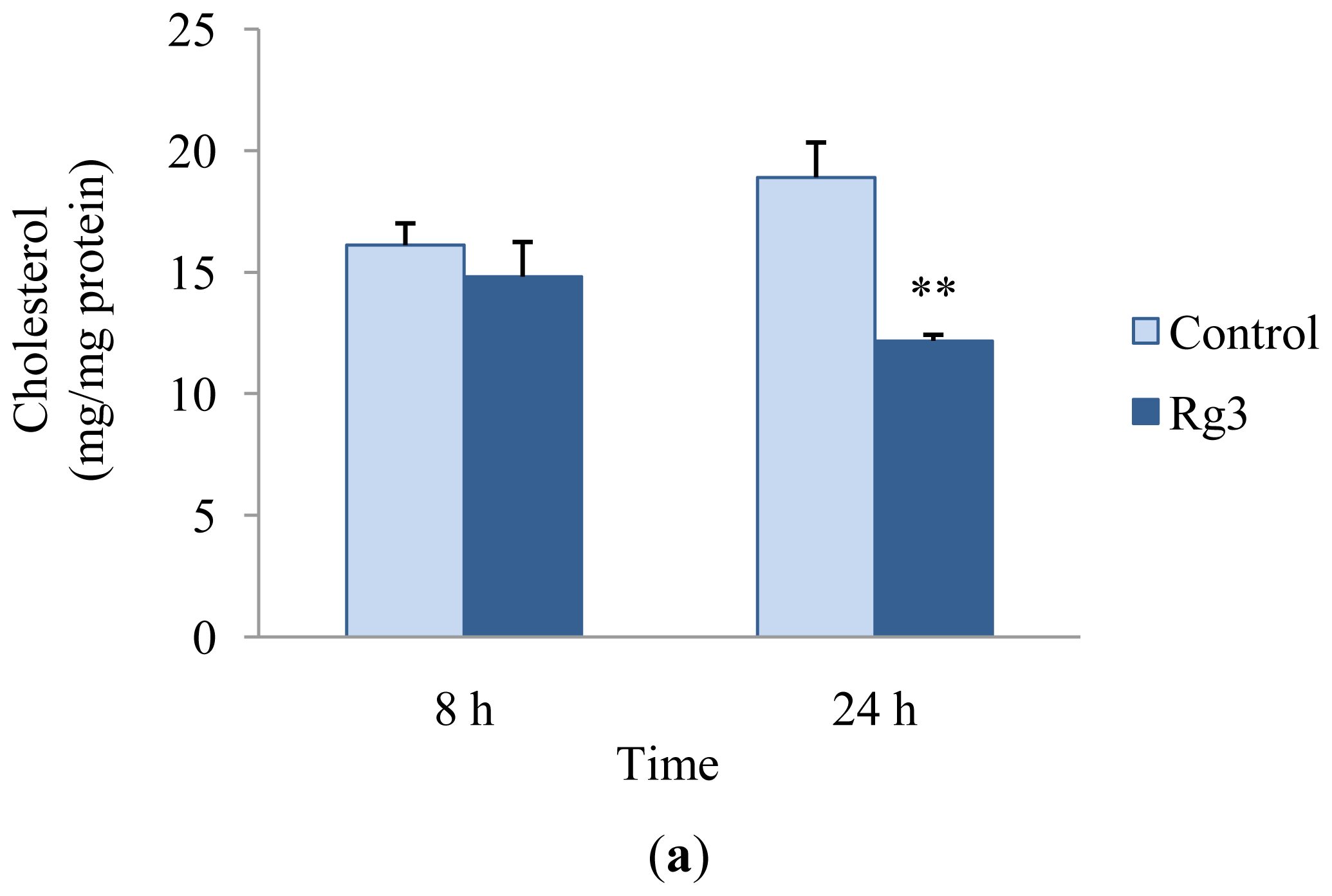
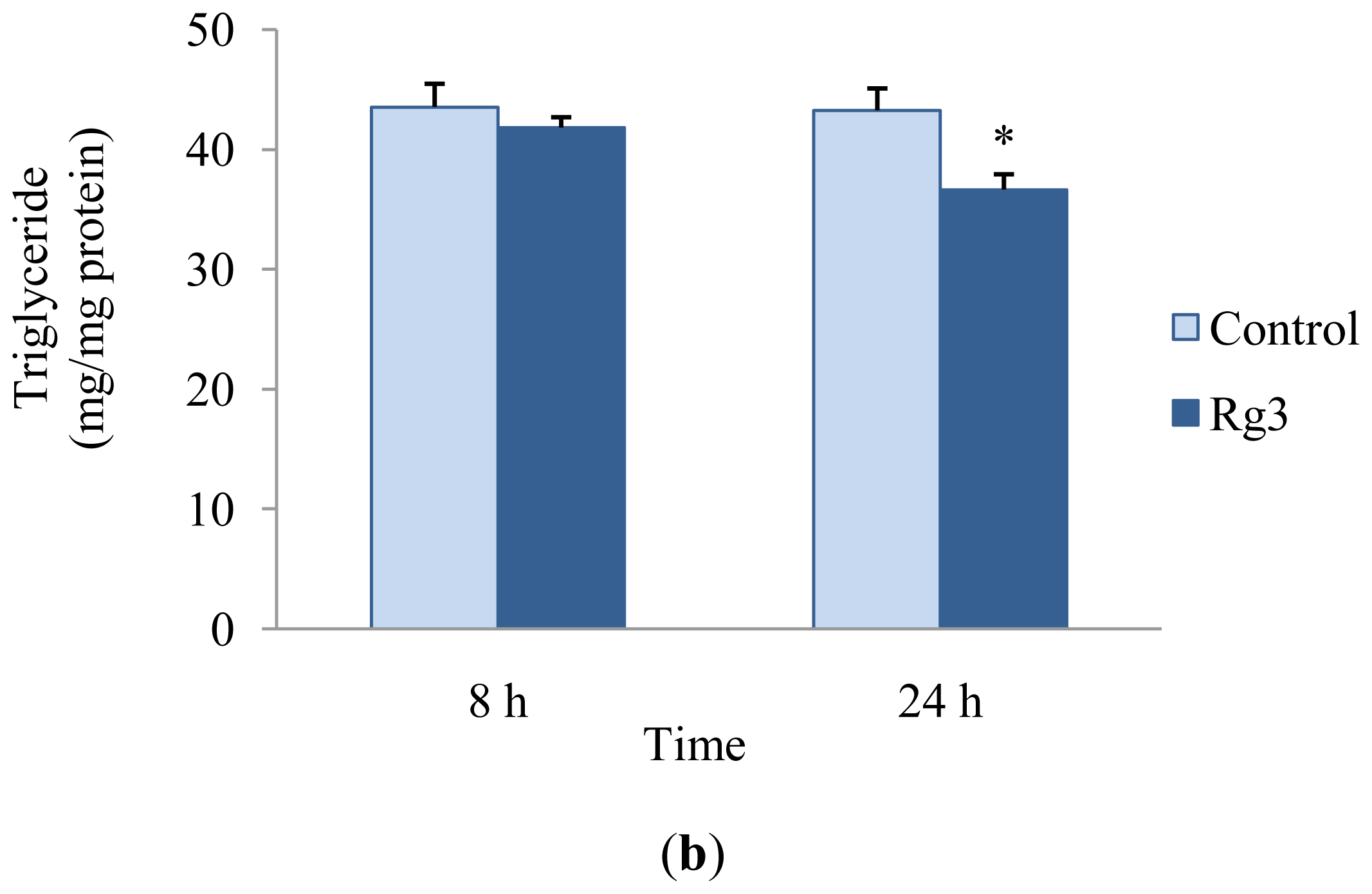
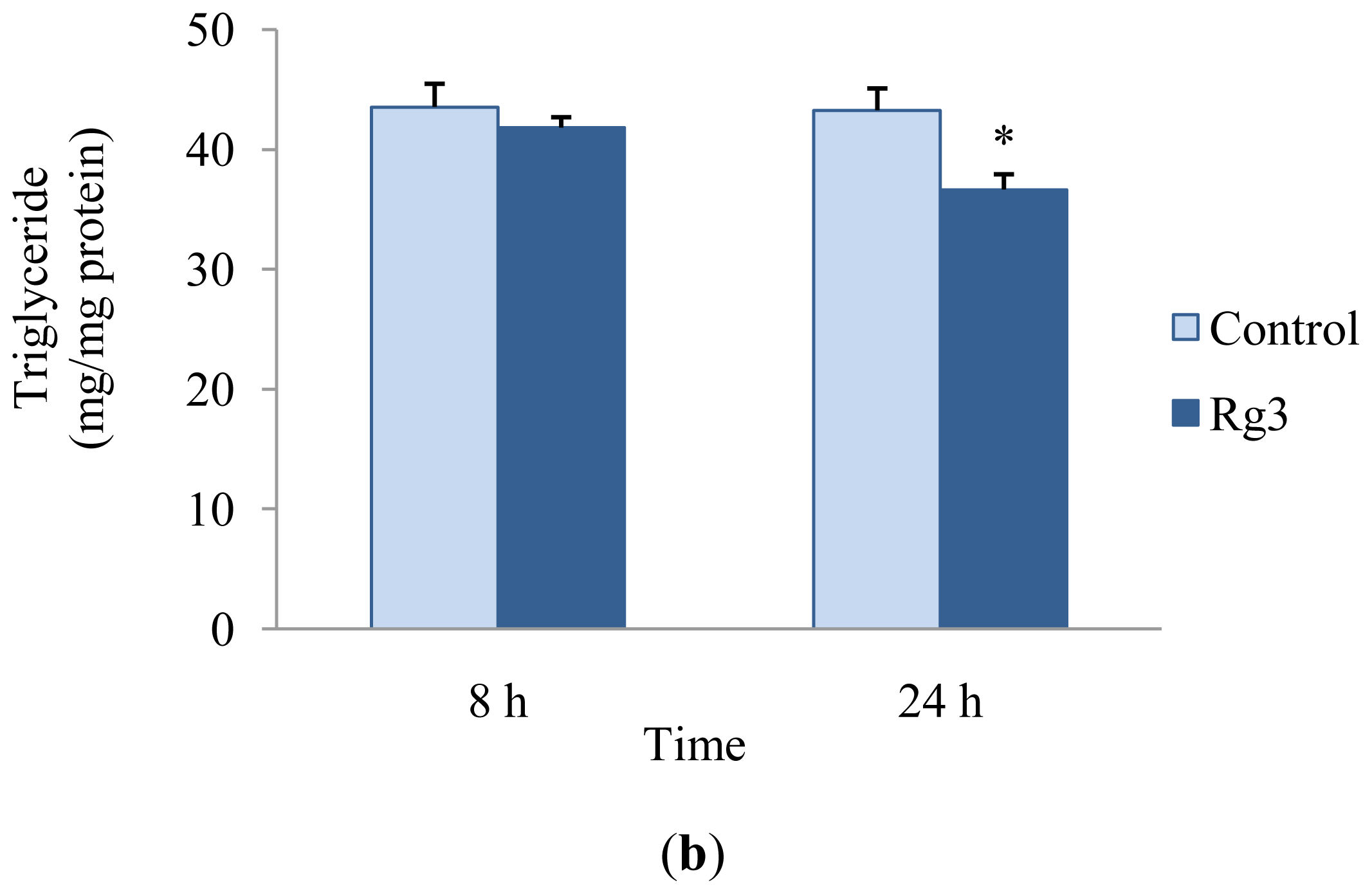
2.2. Effects of Ginsenoside Rg3 on Lipid Metabolism
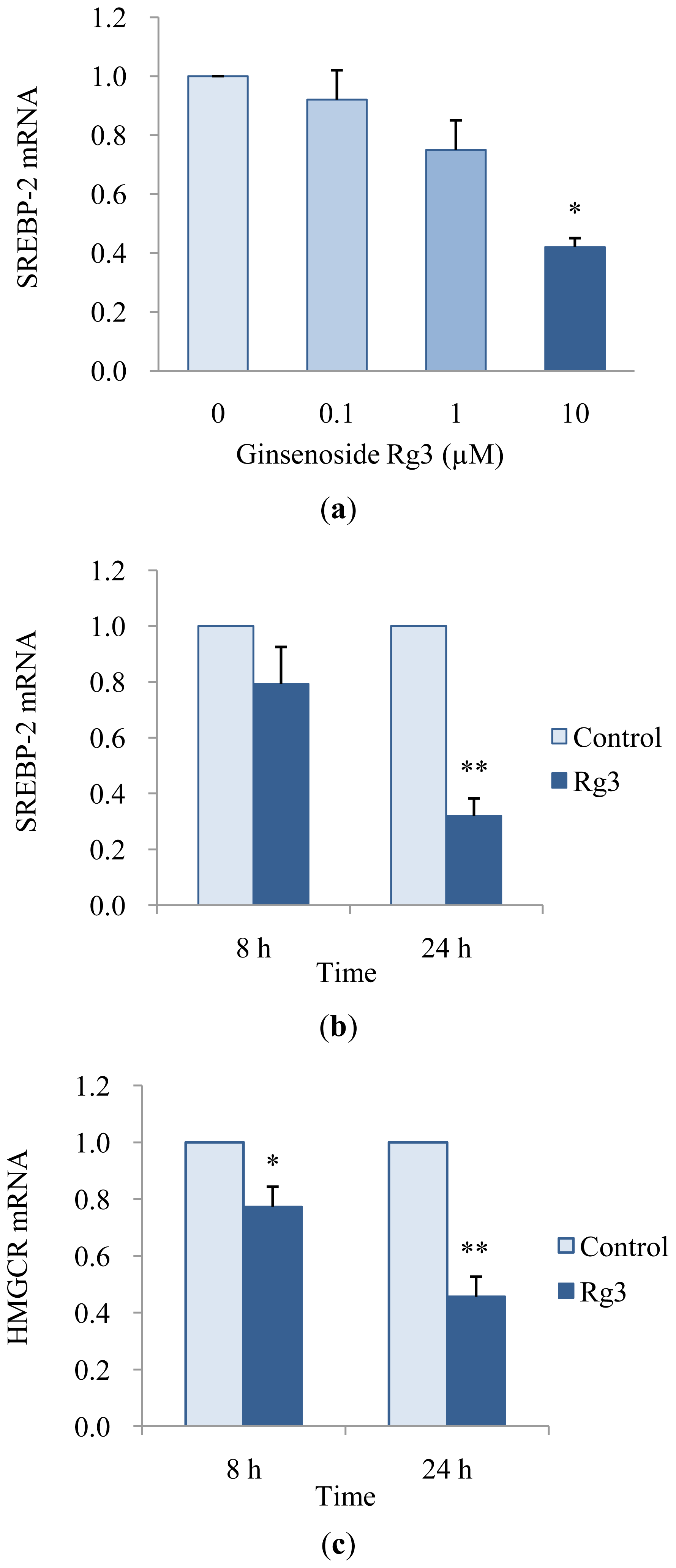
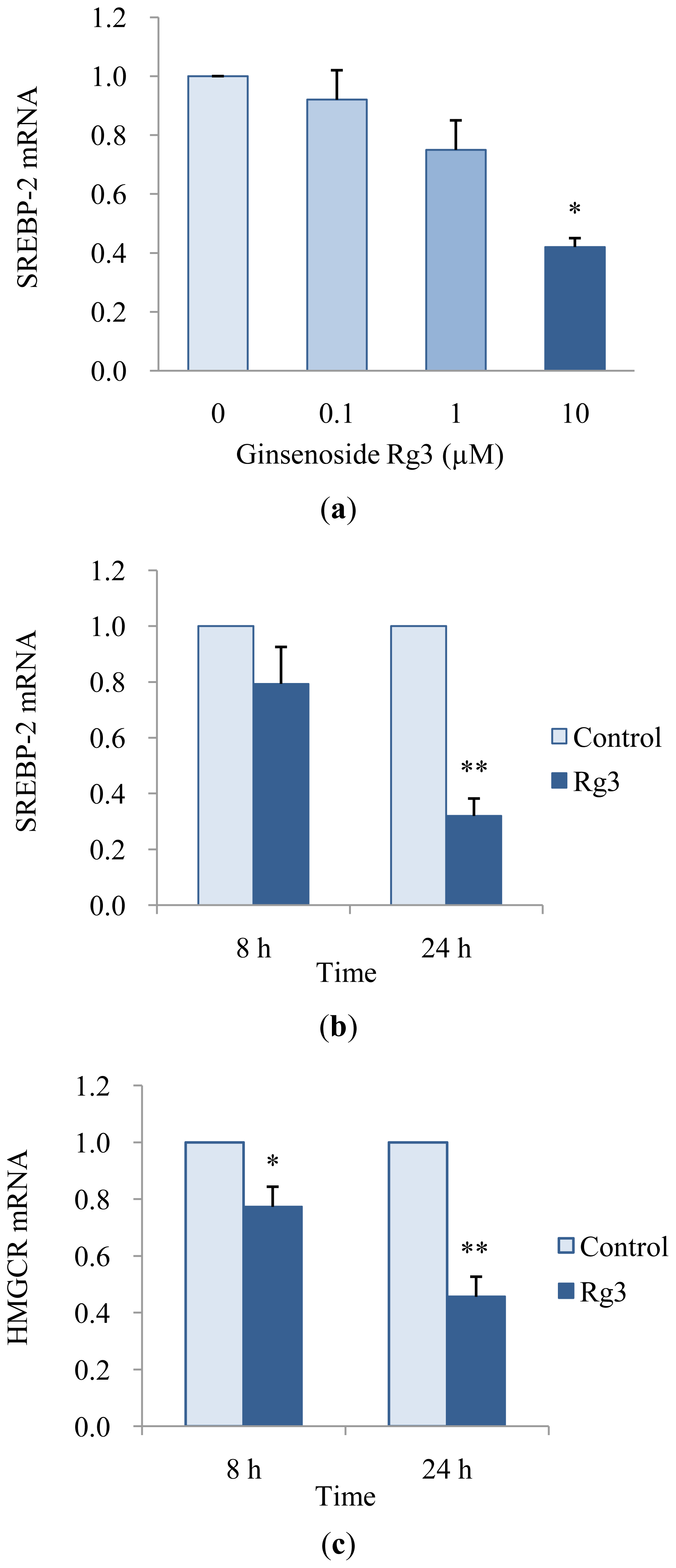
2.3. Effects of Ginsenoside Rg3 on Gene Expression of SREBP-2 and HMGCR
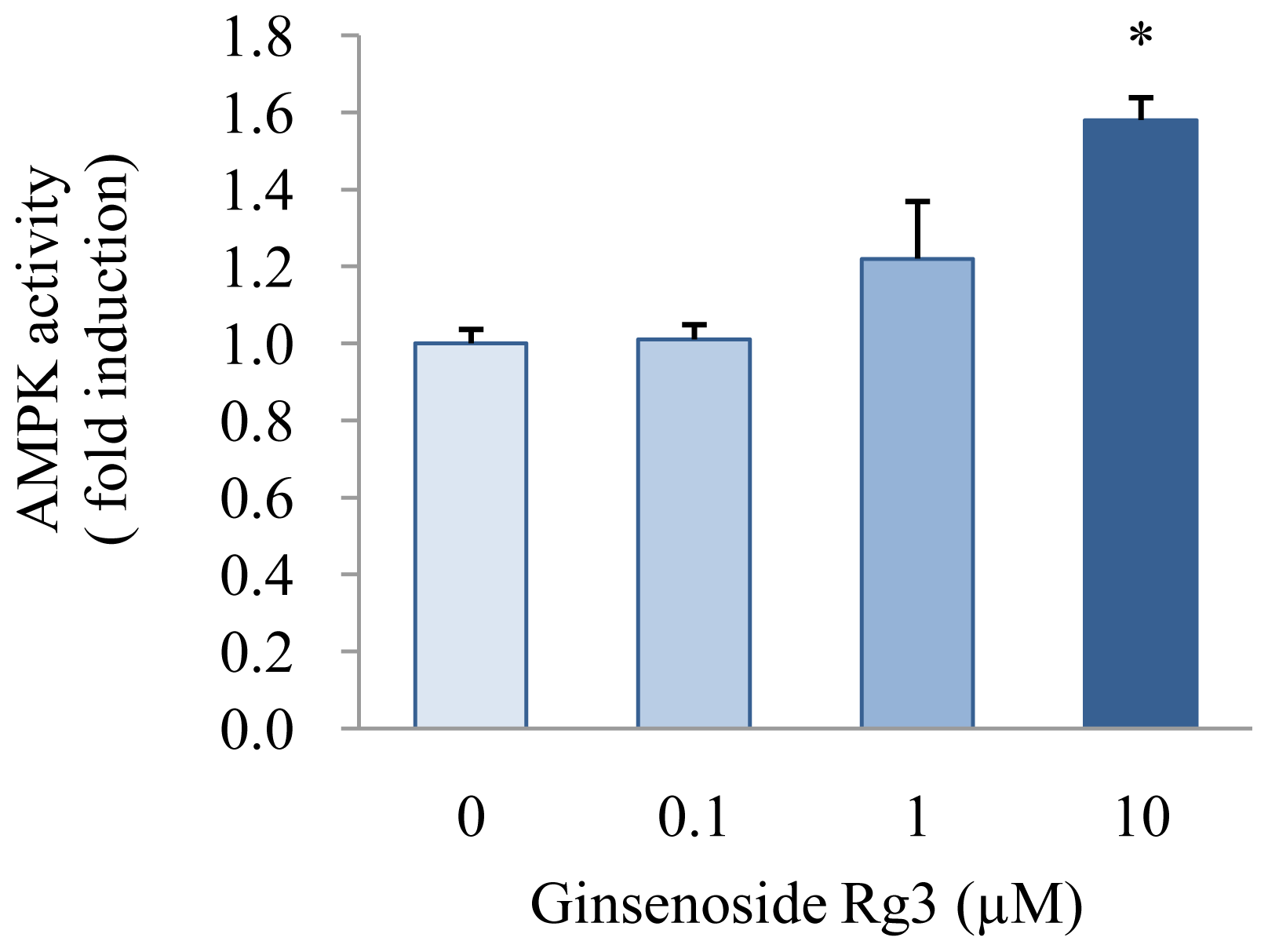
2.4. Effect of Ginsenoside Rg3 on AMPK Activation
3. Experimental Section
3.1. Reagents
3.2. Cell Culture
3.3. Cytotoxicity Assay
3.4. Cholesterol and Triglyceride Assay
3.5. Quantitative Real-Time Polymerase Chain Reaction (PCR)
3.6. AMPK Activity Assay
3.7. Statistical Analysis
4. Conclusions
Acknowledgements
References
- Deaton, C.; Froelicher, E.S.; Wu, L.H.; Ho, C.; Shishani, K.; Jaarsma, T. The global burden of cardiovascular disease. J. Cardiovasc Nurs 2011, 26, S5–S14. [Google Scholar]
- World Health Organization (WHO). Cardiovascular Diseases. 2011. Available online: http://www.who.int/mediacentre/factsheets/fs317/en/index.html accessed on 1 February 2012.
- Lü, J.-M.; Yao, Q.; Chen, C. Ginseng compounds: An update on their molecular mechanisms and medical applications. Curr. Vasc. Parmacol 2009, 7, 293–302. [Google Scholar]
- Hwang, J.-T.; Lee, M.-S.; Kim, H.-J.; Sung, M.-J.; Kim, H.Y.; Kim, M.S.; Kwon, D.Y. Antiobesity effect of ginsenoside Rg3 involves the AMPK and PPAR-γ signal pathways. Phytother. Res 2009, 23, 262–266. [Google Scholar]
- Yeo, C.; Lee, S.; Popovich, D.G. Ginseng (Panax quinquefolius) reduces cell growth, lipid acquisition and increases adiponectin expression in 3T3-L1 cells. Evid.-Based Complement. Alternat. Med 2011, 2011, 610625–610633. [Google Scholar]
- Park, M.W.; Ha, J.; Chung, S.H. 20(S)-ginsenoside Rg3 enhances glucose-stimulated insulin secretion and activates AMPK. Biol. Pharm. Bull 2008, 31, 748–751. [Google Scholar]
- Lee, J.-Y.; Lim, K.-M.; Kim, S.-Y.; Bea, O.-N.; Noh, J.-Y.; Chung, S.-M.; Kim, K.; Shin, Y.-S.; Lee, M.-Y.; Chung, J.-H. Vascular smooth muscle dysfunction and remodeling induced by ginsenoside Rg3, a bioactive component of ginseng. Toxicol. Sci 2010, 117, 505–514. [Google Scholar]
- Lim, S.; Yoon, J.W.; Choi, S.H.; Cho, B.J.; Kim, J.T.; Chang, H.S.; Park, H.S.; Park, K.S.; Lee, H.K.; Kim, Y-B; et al. Effect of ginsam, a vinegar extract from Panax ginseng, on body weight and glucose homeostasis in an obese insulin-resistant rat model. Metabolism 2009, 58, 8–15. [Google Scholar]
- Kim, S.N.; Lee, J.H.; Shin, H.; Son, S.H.; Kim, Y.S. Effects of in vitro-digested ginsenosides on lipid accumulation in 3T3-L1 adipocytes. Planta Med 2009, 75, 596–601. [Google Scholar]
- Brown, M.S.; Goldstein, J.L. The SREBP pathway: Regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 1997, 89, 331–340. [Google Scholar]
- Kang, Q.; Chen, A. Curcumin suppresses expression of low-density lipoprotein (LDL) receptor, leading to the inhibition of LDL-induced activation of hepatic stellate cells. Br. J. Pharmacol 2009, 157, 1354–1367. [Google Scholar]
- Carling, D.; Mayer, F.V.; Sanders, M.J.; Gamblin, S.J. AMP-activated protein kinase: Nature’s energy sensor. Nat. Chem. Biol 2011, 18, 512–518. [Google Scholar]
- Zang, M.; Zuccollo, A.; Hou, X.; Nagate, D.; Walsh, K.; Herscovitz, H.; Brecher, P.; Ruderman, N.B.; Cohen, R.A. AMP-activated protein kinase is required for the lipid-lowering effect of Metformin in insulin-resistant human HepG2 cells. J. Biol. Chem 2004, 279, 47898–47905. [Google Scholar]
- Steinberg, G.R.; Kemp, B.E. AMPK in health and disease. Physiol. Rev 2009, 89, 1025–1078. [Google Scholar]
- Park, I.H.; Piao, L.Z.; Kwon, S.W.; Lee, Y.J.; Cho, S.Y.; Park, M.K.; Park, J.H. Cytotoxic dammarane glycosides from processed ginseng. Chem. Pharm. Bull 2002, 50, 538–540. [Google Scholar]
- Huang, J.; Tang, X.-H.; Ikejima, T.; Sun, X.-J.; Wang, X.-B.; Xi, R.-G.; Wu, L.-J. A new triterpenoid from Panax ginseng exhibits cytotoxicity through p53 and the caspase signaling pathway in the HepG2 cell line. Arch. Pharm. Res 2008, 31, 323–329. [Google Scholar]
- Jiang, J.-W.; Chen, X.-M.; Chen, X.-H.; Zhang, S.-S. Ginsenoside Rg3 inhibit hepatocellular carcinoma growth via intrinsic apoptotic pathway. World J. Gastroenterol 2011, 17, 3605–3613. [Google Scholar]
- Kang, K.S.; Yamabe, N.; Kim, H.Y.; Park, J.H.; Yokozawa, T. Effects of heat-processed ginseng and its active component ginsenoside 20(S)-Rg3 on the progression of renal damage and dysfunction in type 2 diabetic Otsuka Long-Evans Tokushima Fatty rats. Biol. Pharm. Bull 2010, 33, 1077–1081. [Google Scholar]
- Song, T.-B.; An, Y.R.; Kim, S.J.; Park, H.-W.; Jung, J.-W.; Kyung, J.-S.; Hwang, S.Y.; Kim, Y.-S. Lipid metabolic effect of Korean red ginseng extract in mice fed on a high-fat diet. J. Sci. Food Agric 2012, 92, 388–396. [Google Scholar]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Invest 2002, 109, 1125–1131. [Google Scholar]
- Horton, J.D.; Shimomura, I.; Brown, M.S.; Hammer, R.E.; Goldstein, J.L.; Shimano, H. Activation of cholesterol synthesis in preference to fatty acid synthesis in liver and adipose tissue of transgenic mice overproducing sterol regulatory element-binding protein-2. J. Clin. Invest 1998, 101, 2331–2339. [Google Scholar]
- Matsda, M.; Korn, B.S.; Hammer, R.E.; Moon, Y.-A.; Komuro, R.; Horton, J.D.; Goldstein, J.L.; Brown, M.S.; Shimomura, I. SREBP cleavage-activating protein (SCAP) is required for increased lipid synthesis in liver induced by cholesterol deprivation and insulin elevation. Genes Dev 2001, 15, 1206–1216. [Google Scholar]
- Chen, Q.; Reimer, R.A. Dairy protein and leucine alter GLP-1 release and mRNA of genes involved in intestinal lipid metabolism in vitro. Nutrition 2009, 25, 340–349. [Google Scholar]
- Viollet, B.; Guigas, B.; Leclerc, J.; Hébrard, S.; Lantier, L.; Mounier, R.; Andreelli, F.; Foretz, M. AMP-activated protein kinase in the regulation of hepatic energy metabolism: From physiology to therapeutic perspectives. Acta Physiol 2009, 196, 81–98. [Google Scholar]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.-J.; et al. AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab 2011, 13, 376–388. [Google Scholar]
- Rozen, S.; Skaletsky, H.J. Primer3 on the www for General Users and for Biologist Programmers. In Bioinformatics Methods and Protocols: Methods in Molecular Biology; Krawetz, S., Misener, S., Eds.; Humana Press: Totowa, NJ, USA, 1999; Volume 132, pp. 365–386. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar]


© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, S.; Lee, M.-S.; Kim, C.-T.; Kim, I.-H.; Kim, Y. Ginsenoside Rg3 Reduces Lipid Accumulation with AMP-Activated Protein Kinase (AMPK) Activation in HepG2 Cells. Int. J. Mol. Sci. 2012, 13, 5729-5739. https://doi.org/10.3390/ijms13055729
Lee S, Lee M-S, Kim C-T, Kim I-H, Kim Y. Ginsenoside Rg3 Reduces Lipid Accumulation with AMP-Activated Protein Kinase (AMPK) Activation in HepG2 Cells. International Journal of Molecular Sciences. 2012; 13(5):5729-5739. https://doi.org/10.3390/ijms13055729
Chicago/Turabian StyleLee, Seohyun, Mak-Soon Lee, Chong-Tai Kim, In-Hwan Kim, and Yangha Kim. 2012. "Ginsenoside Rg3 Reduces Lipid Accumulation with AMP-Activated Protein Kinase (AMPK) Activation in HepG2 Cells" International Journal of Molecular Sciences 13, no. 5: 5729-5739. https://doi.org/10.3390/ijms13055729