RORα, a Potential Tumor Suppressor and Therapeutic Target of Breast Cancer

Abstract
:1. Introduction
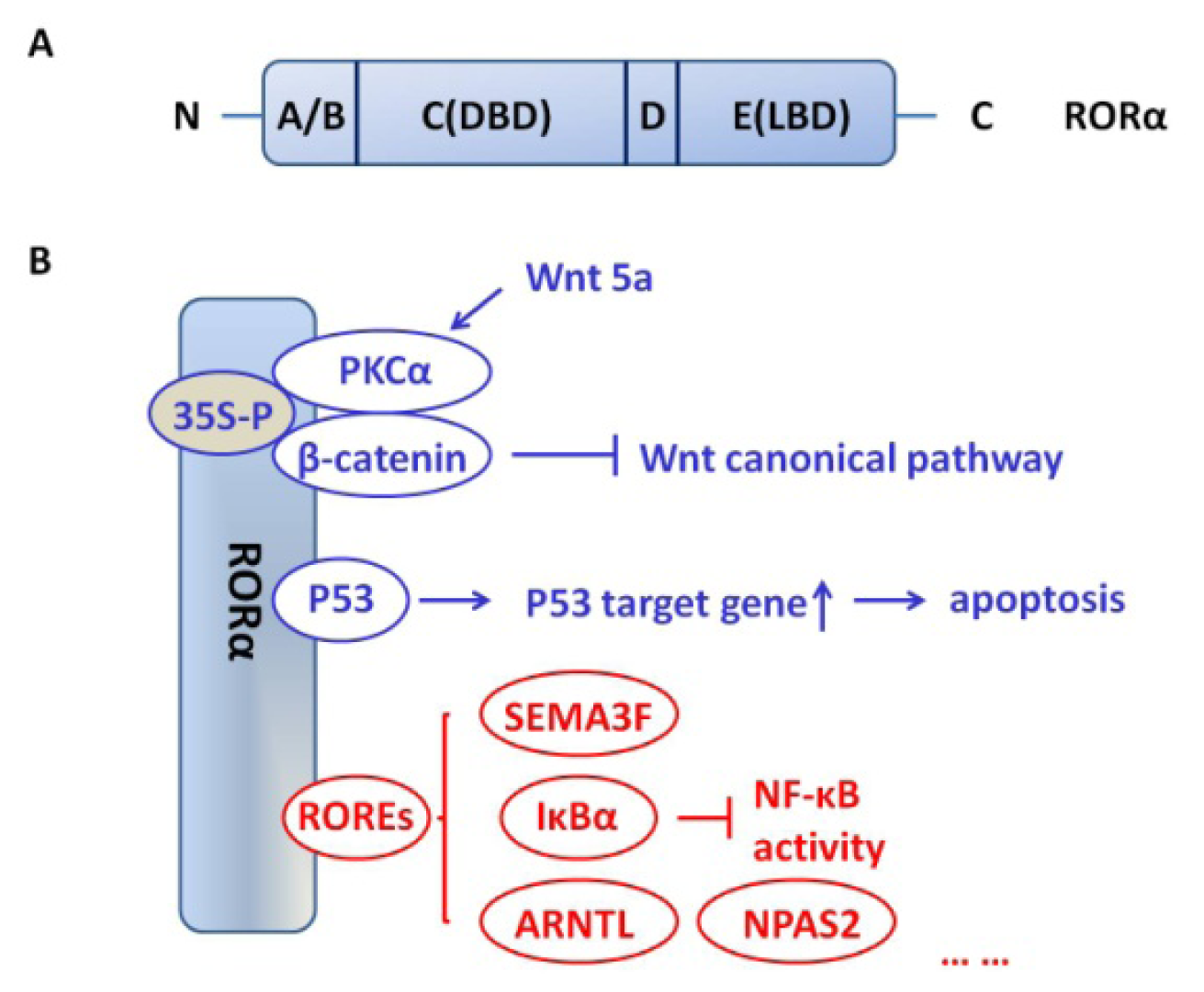
2. RORα Structure
3. RORα Function in Human Breast Cancer
4. Potential Pathways that Mediate the Tumor Suppressive Activities of RORα
4.1. Canonical versus Non-Canonical Pathways
4.2. Role of SEMA3F
4.3. Role of Wnt/β-Catenin
4.4. Role of p53
4.5. Role of Hypoxia/Angiogenesis
4.6. Role of NF-κβ
4.7. Role of Circadian-Related Genes
4.8. Interaction with Other NR
5. ROR α as a Drug Target
6. Conclusions
Acknowledgments
References
- Harris, C.C. Structure and function of the p53 tumor suppressor gene: Clues for rational cancer therapeutic strategies. J. Natl. Cancer Inst 1996, 88, 1442–1455. [Google Scholar]
- Saito, Y.; Gopalan, B.; Mhashilkar, A.M.; Roth, J.A.; Chada, S.; Zumstein, L.; Ramesh, R. Adenovirus-mediated PTEN treatment combined with caffeine produces a synergistic therapeutic effect in colorectal cancer cells. Cancer Gene Ther 2003, 10, 803–813. [Google Scholar]
- Wiggins, J.F.; Ruffino, L.; Kelnar, K.; Omotola, M.; Patrawala, L.; Brown, D.; Bader, A.G. Development of a lung cancer therapeutic based on the tumor suppressor microRNA-34. Cancer Res 2010, 70, 5923–5930. [Google Scholar]
- Forouzanfar, M.H.; Foreman, K.J.; Delossantos, A.M.; Lozano, R.; Lopez, A.D.; Murray, C.J.; Naghavi, M. Breast and cervical cancer in 187 countries between 1980 and 2010: A systematic analysis. Lancet 2011, 378, 1461–1484. [Google Scholar]
- Xiong, G.; Wang, C.; Evers, B.M.; Zhou, B.P.; Xu, R. RORalpha suppresses breast tumor invasion by inducing SEMA3F expression. Cancer Res 2012, 72, 1728–1739. [Google Scholar]
- Kim, H.; Lee, J.M.; Lee, G.; Bhin, J.; Oh, S.K.; Kim, K.; Pyo, K.E.; Lee, J.S.; Yim, H.Y.; Kim, K.I.; et al. DNA damage-induced RORalpha is crucial for p53 stabilization and increased apoptosis. Mol. Cell 2011, 44, 797–810. [Google Scholar]
- Moretti, R.M.; Marelli, M.M.; Motta, M.; Polizzi, D.; Monestiroli, S.; Pratesi, G.; Limonta, P. Activation of the orphan nuclear receptor RORalpha induces growth arrest in androgen-independent DU 145 prostate cancer cells. Prostate 2001, 46, 327–335. [Google Scholar]
- Ariazi, E.A.; Ariazi, J.L.; Cordera, F.; Jordan, V.C. Estrogen receptors as therapeutic targets in breast cancer. Curr. Top. Med. Chem 2006, 6, 181–202. [Google Scholar]
- Widschwendter, M.; Berger, J.; Hermann, M.; Muller, H.M.; Amberger, A.; Zeschnigk, M.; Widschwendter, A.; Abendstein, B.; Zeimet, A.G.; Daxenbichler, G.; et al. Methylation and silencing of the retinoic acid receptor-beta2 gene in breast cancer. J. Natl. Cancer Inst 2000, 92, 826–832. [Google Scholar]
- Liu, Y.; Lee, M.O.; Wang, H.G.; Li, Y.; Hashimoto, Y.; Klaus, M.; Reed, J.C.; Zhang, X. Retinoic acid receptor beta mediates the growth-inhibitory effect of retinoic acid by promoting apoptosis in human breast cancer cells. Mol. Cell. Biol 1996, 16, 1138–1149. [Google Scholar]
- Jetten, A.M.; Kurebayashi, S.; Ueda, E. The ROR nuclear orphan receptor subfamily: Critical regulators of multiple biological processes. Prog. Nucleic Acid Res. Mol. Biol 2001, 69, 205–247. [Google Scholar]
- Becker-Andre, M.; Andre, E.; DeLamarter, J.F. Identification of nuclear receptor mRNAs by RT-PCR amplification of conserved zinc-finger motif sequences. Biochem. Biophys. Res. Commun 1993, 194, 1371–1379. [Google Scholar]
- Giguere, V.; Tini, M.; Flock, G.; Ong, E.; Evans, R.M.; Otulakowski, G. Isoform-specific amino-terminal domains dictate DNA-binding properties of ROR alpha, a novel family of orphan hormone nuclear receptors. Genes Dev 1994, 8, 538–553. [Google Scholar]
- Hamilton, B.A.; Frankel, W.N.; Kerrebrock, A.W.; Hawkins, T.L.; FitzHugh, W.; Kusumi, K.; Russell, L.B.; Mueller, K.L.; van Berkel, V.; Birren, B.W.; et al. Disruption of the nuclear hormone receptor RORalpha in staggerer mice. Nature 1996, 379, 736–739. [Google Scholar]
- Steinmayr, M.; Andre, E.; Conquet, F.; Rondi-Reig, L.; Delhaye-Bouchaud, N.; Auclair, N.; Daniel, H.; Crepel, F.; Mariani, J.; Sotelo, C.; et al. staggerer phenotype in retinoid-related orphan receptor alpha-deficient mice. Proc. Natl. Acad. Sci. USA 1998, 95, 3960–3965. [Google Scholar]
- Jarvis, C.I.; Staels, B.; Brugg, B.; Lemaigre-Dubreuil, Y.; Tedgui, A.; Mariani, J. Age-related phenotypes in the staggerer mouse expand the RORalpha nuclear receptor’s role beyond the cerebellum. Mol. Cell. Endocrinol 2002, 186, 1–5. [Google Scholar]
- Wang, N.S.; McHeyzer-Williams, L.J.; Okitsu, S.L.; Burris, T.P.; Reiner, S.L.; McHeyzer-Williams, M.G. Divergent transcriptional programming of class-specific B cell memory by T-bet and RORalpha. Nat. Immunol 2012, 13, 604–611. [Google Scholar]
- Wong, S.H.; Walker, J.A.; Jolin, H.E.; Drynan, L.F.; Hams, E.; Camelo, A.; Barlow, J.L.; Neill, D.R.; Panova, V.; Koch, U.; et al. Transcription factor RORalpha is critical for nuocyte development. Nat. Immunol 2012, 13, 229–236. [Google Scholar]
- Kang, H.S.; Okamoto, K.; Takeda, Y.; Beak, J.Y.; Gerrish, K.; Bortner, C.D.; DeGraff, L.M.; Wada, T.; Xie, W.; Jetten, A.M. Transcriptional profiling reveals a role for RORalpha in regulating gene expression in obesity-associated inflammation and hepatic steatosis. Physiol. Genomics 2011, 43, 818–828. [Google Scholar]
- Jetten, A.M. Retinoid-related orphan receptors (RORs): Critical roles in development, immunity, circadian rhythm, and cellular metabolism. Nucl. Recept. Signal 2009, 7, e003. [Google Scholar]
- Kallen, J.; Schlaeppi, J.M.; Bitsch, F.; Delhon, I.; Fournier, B. Crystal structure of the human RORalpha Ligand binding domain in complex with cholesterol sulfate at 2.2 A. J. Biol. Chem 2004, 279, 14033–14038. [Google Scholar]
- Kallen, J.A.; Schlaeppi, J.M.; Bitsch, F.; Geisse, S.; Geiser, M.; Delhon, I.; Fournier, B. X-ray structure of the hRORalpha LBD at 1.63 A: Structural and functional data that cholesterol or a cholesterol derivative is the natural ligand of RORalpha. Structure 2002, 10, 1697–1707. [Google Scholar]
- Zenri, F.; Hiroi, H.; Momoeda, M.; Tsutsumi, R.; Hosokawa, Y.; Koizumi, M.; Nakae, H.; Osuga, Y.; Yano, T.; Taketani, Y. Expression of retinoic acid-related orphan receptor alpha and its responsive genes in human endometrium regulated by cholesterol sulfate. J. Steroid Biochem. Mol. Biol 2012, 128, 21–28. [Google Scholar]
- Zhu, Y.; McAvoy, S.; Kuhn, R.; Smith, D.I. RORA, a large common fragile site gene, is involved in cellular stress response. Oncogene 2006, 25, 2901–2908. [Google Scholar]
- Yamashita, S.; Tsujino, Y.; Moriguchi, K.; Tatematsu, M.; Ushijima, T. Chemical genomic screening for methylation-silenced genes in gastric cancer cell lines using 5-aza-2′-deoxycytidine treatment and oligonucleotide microarray. Cancer Sci 2006, 97, 64–71. [Google Scholar]
- Lee, J.M.; Kim, I.S.; Kim, H.; Lee, J.S.; Kim, K.; Yim, H.Y.; Jeong, J.; Kim, J.H.; Kim, J.Y.; Lee, H.; et al. RORalpha attenuates Wnt/beta-catenin signaling by PKCalpha-dependent phosphorylation in colon cancer. Mol. Cell 2010, 37, 183–195. [Google Scholar]
- Dai, J.; Ram, P.T.; Yuan, L.; Spriggs, L.L.; Hill, S.M. Transcriptional repression of RORalpha activity in human breast cancer cells by melatonin. Mol. Cell. Endocrinol 2001, 176, 111–120. [Google Scholar]
- Lu, Y.; Yi, Y.; Liu, P.; Wen, W.; James, M.; Wang, D.; You, M. Common human cancer genes discovered by integrated gene-expression analysis. PLoS One 2007, 2, e1149. [Google Scholar]
- Shimizu, A.; Mammoto, A.; Italiano, J.E., Jr; Pravda, E.; Dudley, A.C.; Ingber, D.E.; Klagsbrun, M. ABL2/ARG tyrosine kinase mediates SEMA3F-induced RhoA inactivation and cytoskeleton collapse in human glioma cells. J. Biol. Chem. 2008, 283, 27230–27238. [Google Scholar]
- Potiron, V.A.; Sharma, G.; Nasarre, P.; Clarhaut, J.A.; Augustin, H.G.; Gemmill, R.M.; Roche, J.; Drabkin, H.A. Semaphorin SEMA3F affects multiple signaling pathways in lung cancer cells. Cancer Res 2007, 67, 8708–8715. [Google Scholar]
- Benhaj, K.; Akcali, K.C.; Ozturk, M. Redundant expression of canonical Wnt ligands in human breast cancer cell lines. Oncol. Rep 2006, 15, 701–707. [Google Scholar]
- Yan, L.; Della Coletta, L.; Powell, K.L.; Shen, J.; Thames, H.; Aldaz, C.M.; MacLeod, M.C. Activation of the canonical Wnt/beta-catenin pathway in ATF3-induced mammary tumors. PLoS One 2011, 6, e16515. [Google Scholar]
- Kremenevskaja, N.; von Wasielewski, R.; Rao, A.S.; Schofl, C.; Andersson, T.; Brabant, G. Wnt-5a has tumor suppressor activity in thyroid carcinoma. Oncogene 2005, 24, 2144–2154. [Google Scholar]
- Leris, A.C.; Roberts, T.R.; Jiang, W.G.; Newbold, R.F.; Mokbel, K. WNT5A expression in human breast cancer. Anticancer Res 2005, 25, 731–734. [Google Scholar]
- Knappskog, S.; Lonning, P.E. p53 and its molecular basis to chemoresistance in breast cancer. Expert Opin. Ther. Targets 2012, 16, S23–S30. [Google Scholar]
- Lacroix, M.; Toillon, R.A.; Leclercq, G. p53 and breast cancer, an update. Endocr. Relat. Cancer 2006, 13, 293–325. [Google Scholar]
- Wang, Y.; Solt, L.A.; Kojetin, D.J.; Burris, T.P. Regulation of p53 stability and apoptosis by a ROR agonist. PLoS One 2012, 7, e34921. [Google Scholar]
- Pugh, C.W.; Gleadle, J.; Maxwell, P.H. Hypoxia and oxidative stress in breast cancer. Hypoxia signalling pathways. Breast Cancer Res 2001, 3, 313–317. [Google Scholar]
- Besnard, S.; Silvestre, J.S.; Duriez, M.; Bakouche, J.; Lemaigre-Dubreuil, Y.; Mariani, J.; Levy, B.I.; Tedgui, A. Increased ischemia-induced angiogenesis in the staggerer mouse, a mutant of the nuclear receptor Roralpha. Circ. Res 2001, 89, 1209–1215. [Google Scholar]
- Dales, J.P.; Garcia, S.; Meunier-Carpentier, S.; Andrac-Meyer, L.; Haddad, O.; Lavaut, M.N.; Allasia, C.; Bonnier, P.; Charpin, C. Overexpression of hypoxia-inducible factor HIF-1alpha predicts early relapse in breast cancer: retrospective study in a series of 745 patients. Int. J. Cancer 2005, 116, 734–739. [Google Scholar]
- Miki, N.; Ikuta, M.; Matsui, T. Hypoxia-induced activation of the retinoic acid receptor-related orphan receptor alpha4 gene by an interaction between hypoxia-inducible factor-1 and Sp1. J. Biol. Chem 2004, 279, 15025–15031. [Google Scholar]
- Chauvet, C.; Bois-Joyeux, B.; Danan, J.L. Retinoic acid receptor-related orphan receptor (ROR) alpha4 is the predominant isoform of the nuclear receptor RORalpha in the liver and is up-regulated by hypoxia in HepG2 human hepatoma cells. Biochem. J 2002, 364, 449–456. [Google Scholar]
- Journiac, N.; Jolly, S.; Jarvis, C.; Gautheron, V.; Rogard, M.; Trembleau, A.; Blondeau, J.P.; Mariani, J.; Vernet-der Garabedian, B. The nuclear receptor ROR(alpha) exerts a bi-directional regulation of IL-6 in resting and reactive astrocytes. Proc. Natl. Acad. Sci. USA 2009, 106, 21365–21370. [Google Scholar]
- Delerive, P.; Monte, D.; Dubois, G.; Trottein, F.; Fruchart-Najib, J.; Mariani, J.; Fruchart, J.C.; Staels, B. The orphan nuclear receptor ROR alpha is a negative regulator of the inflammatory response. EMBO Rep 2001, 2, 42–48. [Google Scholar]
- Migita, H.; Morser, J.; Kawai, K. Rev-erbalpha upregulates NF-kappaB-responsive genes in vascular smooth muscle cells. FEBS Lett 2004, 561, 69–74. [Google Scholar]
- Guillaumond, F.; Dardente, H.; Giguere, V.; Cermakian, N. Differential control of Bmal1 circadian transcription by REV-ERB and ROR nuclear receptors. J. Biol. Rhythms 2005, 20, 391–403. [Google Scholar]
- Hayden, M.S.; Ghosh, S. Signaling to NF-kappaB. Genes Dev 2004, 18, 2195–2224. [Google Scholar]
- Nakshatri, H.; Bhat-Nakshatri, P.; Martin, D.A.; Goulet, R.J., Jr; Sledge, G.W., Jr. Constitutive activation of NF-kappaB during progression of breast cancer to hormone-independent growth. Mol. Cell. Biol. 1997, 17, 3629–3639. [Google Scholar]
- Huber, M.A.; Azoitei, N.; Baumann, B.; Grunert, S.; Sommer, A.; Pehamberger, H.; Kraut, N.; Beug, H.; Wirth, T. NF-kappaB is essential for epithelial-mesenchymal transition and metastasis in a model of breast cancer progression. J. Clin. Invest 2004, 114, 569–581. [Google Scholar]
- Cao, Y.; Karin, M. NF-kappaB in mammary gland development and breast cancer. J. Mammary Gland Biol. Neoplasia 2003, 8, 215–223. [Google Scholar]
- Sahar, S.; Sassone-Corsi, P. Circadian clock and breast cancer: A molecular link. Cell Cycle 2007, 6, 1329–1331. [Google Scholar]
- Stevens, R.G. Circadian disruption and breast cancer: from melatonin to clock genes. Epidemiology 2005, 16, 254–258. [Google Scholar]
- Zhu, Y.; Stevens, R.G.; Leaderer, D.; Hoffman, A.; Holford, T.; Zhang, Y.; Brown, H.N.; Zheng, T. Non-synonymous polymorphisms in the circadian gene NPAS2 and breast cancer risk. Breast Cancer Res. Treat 2008, 107, 421–425. [Google Scholar]
- Winter, S.L.; Bosnoyan-Collins, L.; Pinnaduwage, D.; Andrulis, I.L. Expression of the circadian clock genes Per1 and Per2 in sporadic and familial breast tumors. Neoplasia 2007, 9, 797–800. [Google Scholar]
- Fu, L.; Pelicano, H.; Liu, J.; Huang, P.; Lee, C. The circadian gene Period2 plays an important role in tumor suppression and DNA damage response in vivo. Cell 2002, 111, 41–50. [Google Scholar]
- Sato, T.K.; Panda, S.; Miraglia, L.J.; Reyes, T.M.; Rudic, R.D.; McNamara, P.; Naik, K.A.; FitzGerald, G.A.; Kay, S.A.; Hogenesch, J.B. A functional genomics strategy reveals Rora as a component of the mammalian circadian clock. Neuron 2004, 43, 527–537. [Google Scholar]
- Akashi, M.; Takumi, T. The orphan nuclear receptor RORalpha regulates circadian transcription of the mammalian core-clock Bmal1. Nat. Struct. Mol. Biol 2005, 12, 441–448. [Google Scholar]
- Crumbley, C.; Wang, Y.; Kojetin, D.J.; Burris, T.P. Characterization of the core mammalian clock component, NPAS2, as a REV-ERBalpha/RORalpha target gene. J. Biol. Chem 2010, 285, 35386–35392. [Google Scholar]
- Dong, C.; Yuan, L.; Dai, J.; Lai, L.; Mao, L.; Xiang, S.; Rowan, B.; Hill, S.M. Melatonin inhibits mitogenic cross-talk between retinoic acid-related orphan receptor alpha (RORalpha) and ERalpha in MCF-7 human breast cancer cells. Steroids 2010, 75, 944–951. [Google Scholar]
- Odawara, H.; Iwasaki, T.; Horiguchi, J.; Rokutanda, N.; Hirooka, K.; Miyazaki, W.; Koibuchi, Y.; Shimokawa, N.; Iino, Y.; Takeyoshi, I.; et al. Activation of aromatase expression by retinoic acid receptor-related orphan receptor (ROR) alpha in breast cancer cells: Identification of a novel ROR response element. J. Biol. Chem 2009, 284, 17711–17719. [Google Scholar]
- Gu, F.; Hsu, H.K.; Hsu, P.Y.; Wu, J.; Ma, Y.; Parvin, J.; Huang, T.H.; Jin, V.X. Inference of hierarchical regulatory network of estrogen-dependent breast cancer through ChIP-based data. BMC Syst. Biol 2010, 4, 170. [Google Scholar]
- Wang, Y.; Kumar, N.; Nuhant, P.; Cameron, M.D.; Istrate, M.A.; Roush, W.R.; Griffin, P.R.; Burris, T.P. Identification of SR1078, a synthetic agonist for the orphan nuclear receptors RORalpha and RORgamma. ACS Chem. Biol 2010, 5, 1029–1034. [Google Scholar]
- Wiesenberg, I.; Missbach, M.; Kahlen, J.P.; Schrader, M.; Carlberg, C. Transcriptional activation of the nuclear receptor RZR alpha by the pineal gland hormone melatonin and identification of CGP 52608 as a synthetic ligand. Nucleic Acids Res 1995, 23, 327–333. [Google Scholar]
- Carlberg, C. Gene regulation by melatonin. Ann. NY Acad. Sci 2000, 917, 387–396. [Google Scholar]
- Korkmaz, A.; Sanchez-Barcelo, E.J.; Tan, D.X.; Reiter, R.J. Role of melatonin in the epigenetic regulation of breast cancer. Breast Cancer Res. Treat 2009, 115, 13–27. [Google Scholar]
- Winczyk, K.; Pawlikowski, M.; Karasek, M. Melatonin and RZR/ROR receptor ligand CGP 52608 induce apoptosis in the murine colonic cancer. J. Pineal Res 2001, 31, 179–182. [Google Scholar]
- Winczyk, K.; Pawlikowski, M.; Guerrero, J.M.; Karasek, M. Possible involvement of the nuclear RZR/ROR-alpha receptor in the antitumor action of melatonin on murine Colon 38 cancer. Tumor Biol 2002, 23, 298–302. [Google Scholar]


{kind=link}
| Analysis type by cancer | Normal vs. Cancer | |
|---|---|---|
| bladder cancer | 1 | |
| brain and CNS cancer | 2 | |
| breast cancer | 9 | 2 |
| cervical cancer | 3 | |
| colorectal cancer | 5 | |
| esophageal cancer | 7 | |
| gastric cancer | 2 | |
| head and neck cancer | 5 | |
| kidney cancer | 1 | 1 |
| leukemia | 9 | 2 |
| liver cancer | 1 | |
| lung cancer | 2 | 1 |
| lymphoma | 4 | 4 |
| melanoma | 3 | 1 |
| myeloma | 1 | 1 |
| other cancer | 9 | |
| ovarian cancer | 1 | |
| pancreatic cancer | 2 | |
| prostate cancer | 2 | |
| sarcoma | 1 | 1 |
| significant unique analyses | 68 | 15 |
| total unique analyses | 381 | |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Du, J.; Xu, R. RORα, a Potential Tumor Suppressor and Therapeutic Target of Breast Cancer. Int. J. Mol. Sci. 2012, 13, 15755-15766. https://doi.org/10.3390/ijms131215755
Du J, Xu R. RORα, a Potential Tumor Suppressor and Therapeutic Target of Breast Cancer. International Journal of Molecular Sciences. 2012; 13(12):15755-15766. https://doi.org/10.3390/ijms131215755
Chicago/Turabian StyleDu, Jun, and Ren Xu. 2012. "RORα, a Potential Tumor Suppressor and Therapeutic Target of Breast Cancer" International Journal of Molecular Sciences 13, no. 12: 15755-15766. https://doi.org/10.3390/ijms131215755