Lewis y Regulate Cell Cycle Related Factors in Ovarian Carcinoma Cell RMG-I in Vitro via ERK and Akt Signaling Pathways

Abstract
:Objective
Methods
Results
Conclusion
1. Introduction
2. Results
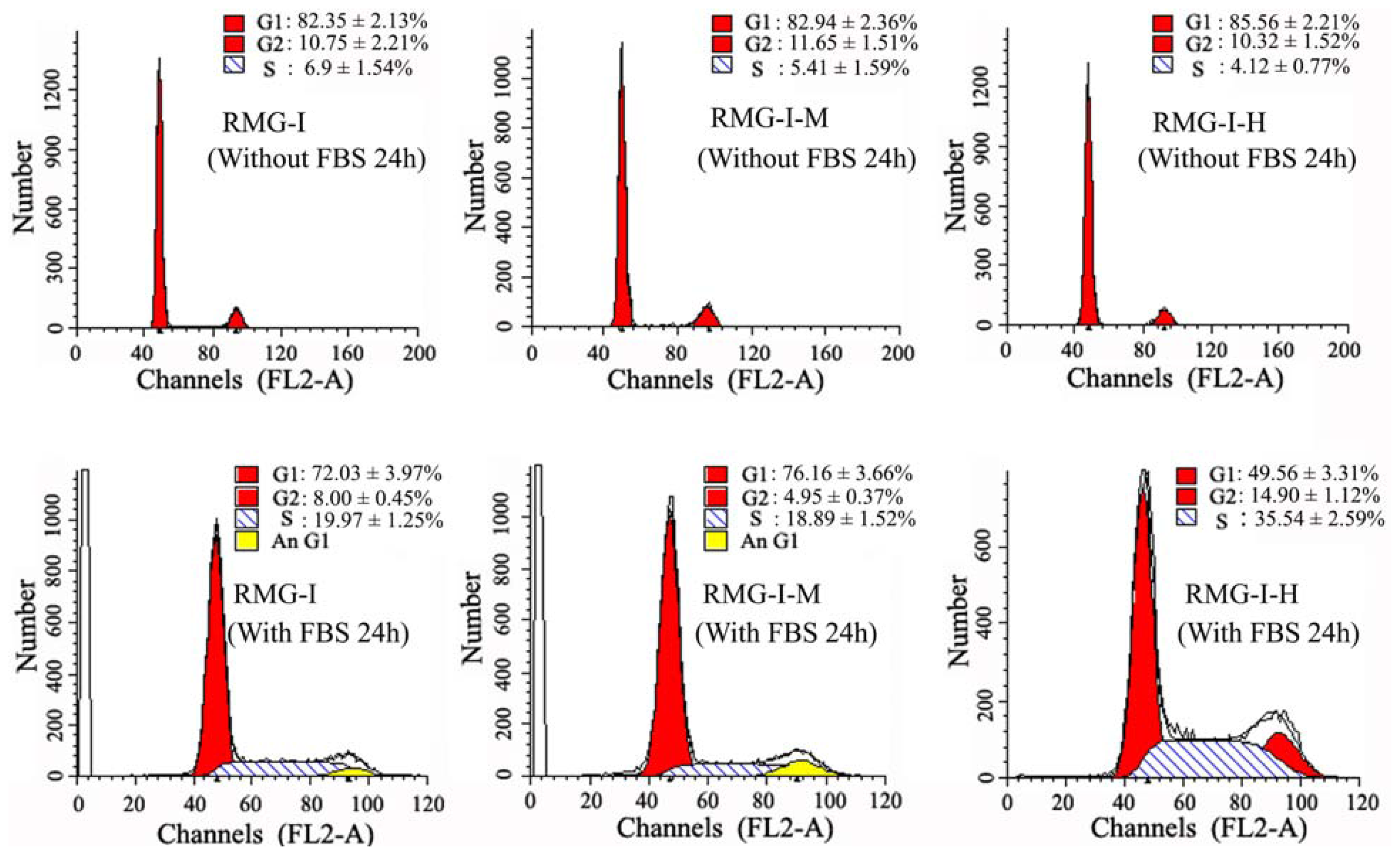
2.1. Lewis Y Overexpression Promoted Ovarian Cancer Cells to Enter S Phase
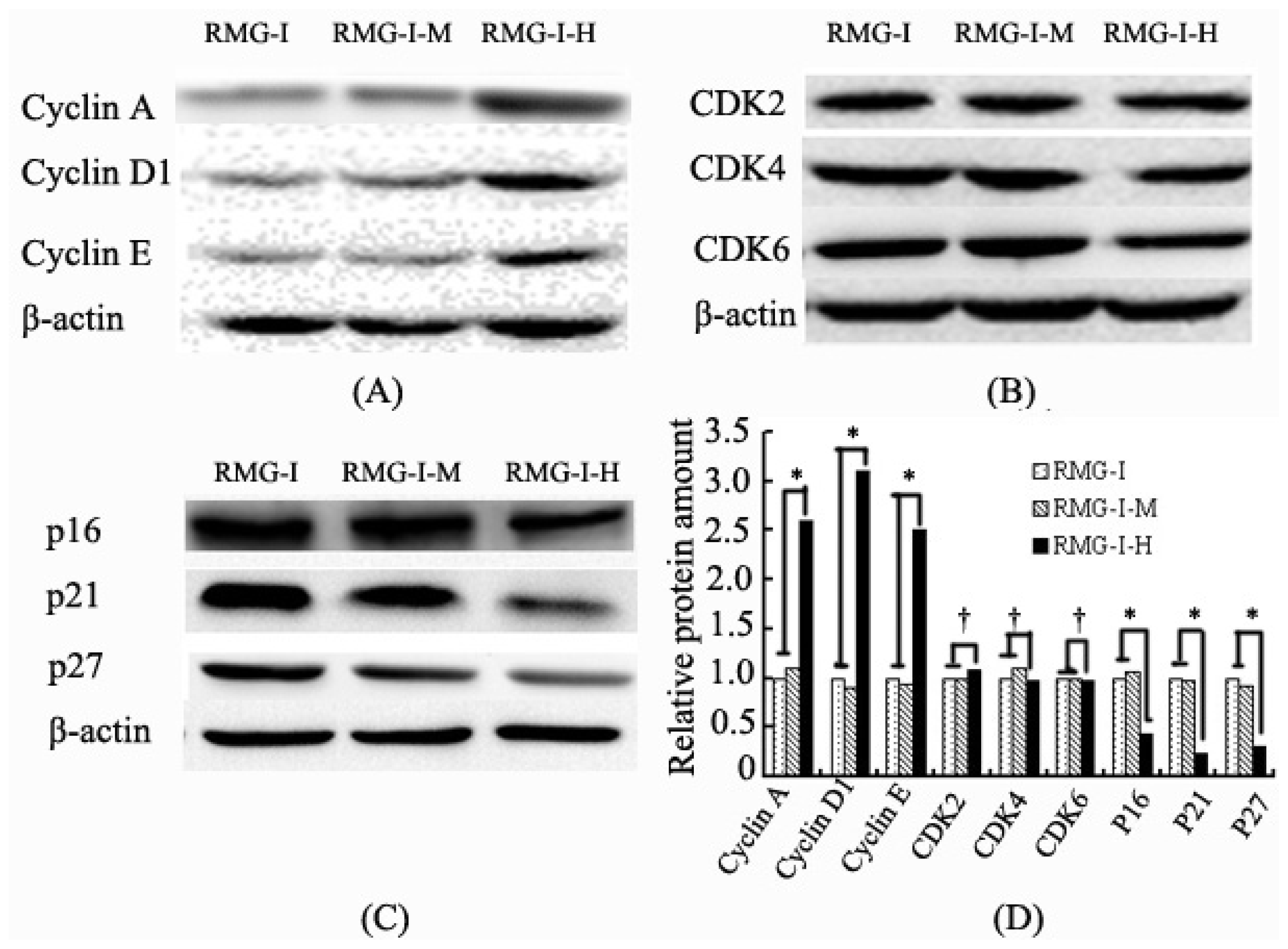
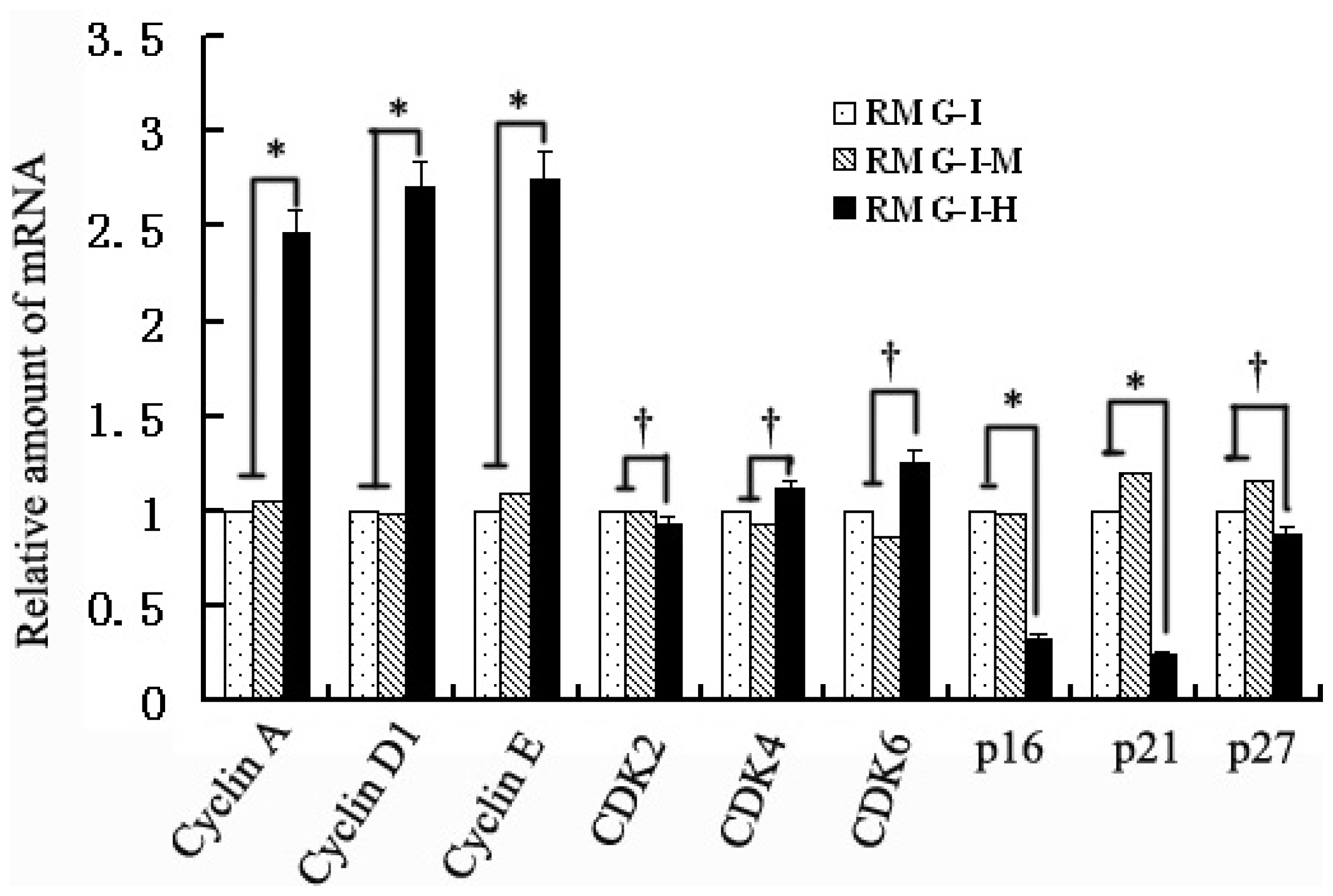
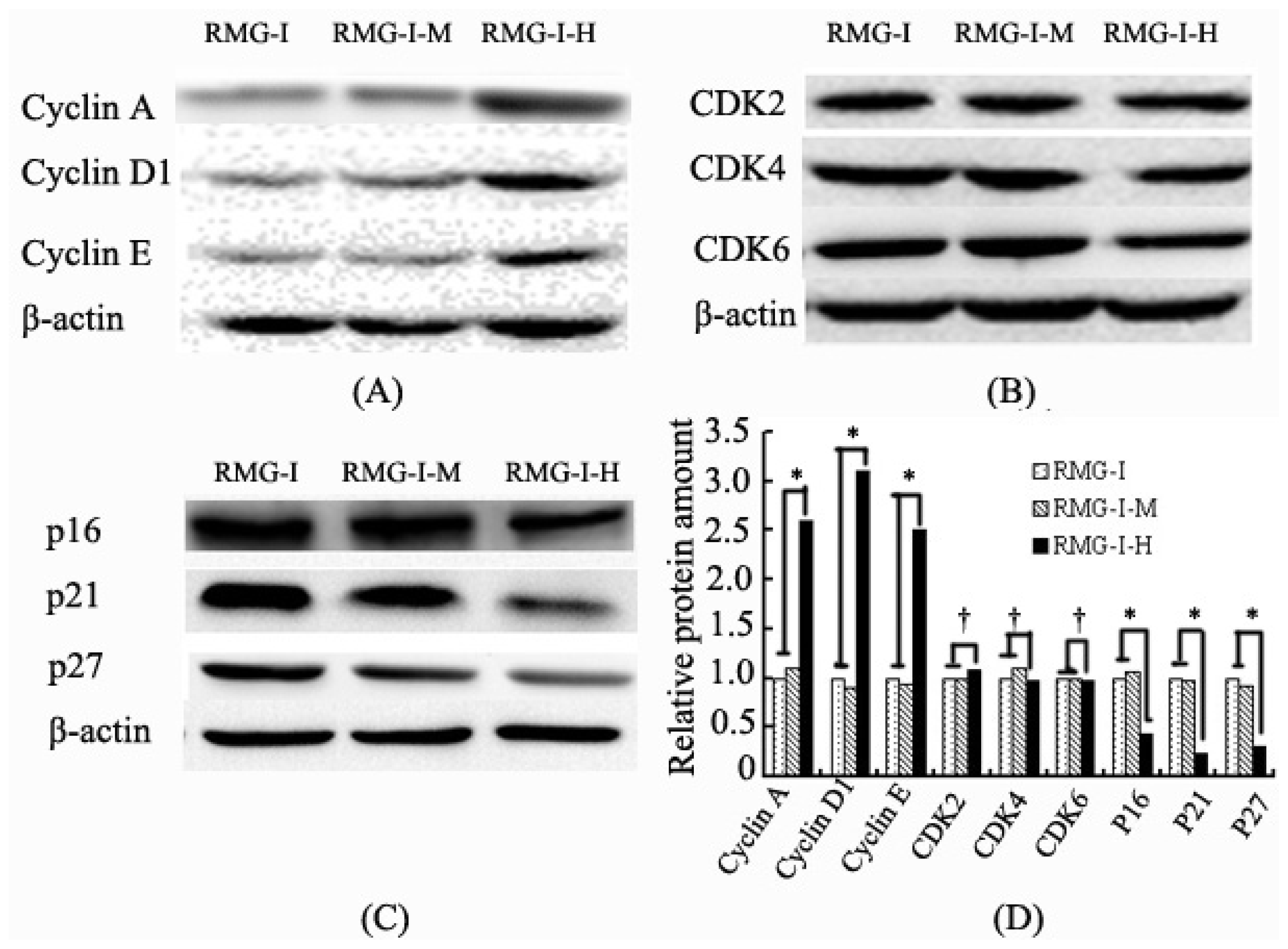
2.2. Lewis Y Overexpression Increased mRNA Expression Levels of Cyclins, p16 and p21 Without Affecting Both CDKs and p27 mRNA Expression in Ovarian Cancer Cells
2.3. Lewis Y Overexpression Promoted Cyclin and CKI Expression Without Affecting CDK Expression in Ovarian Cancer Cells
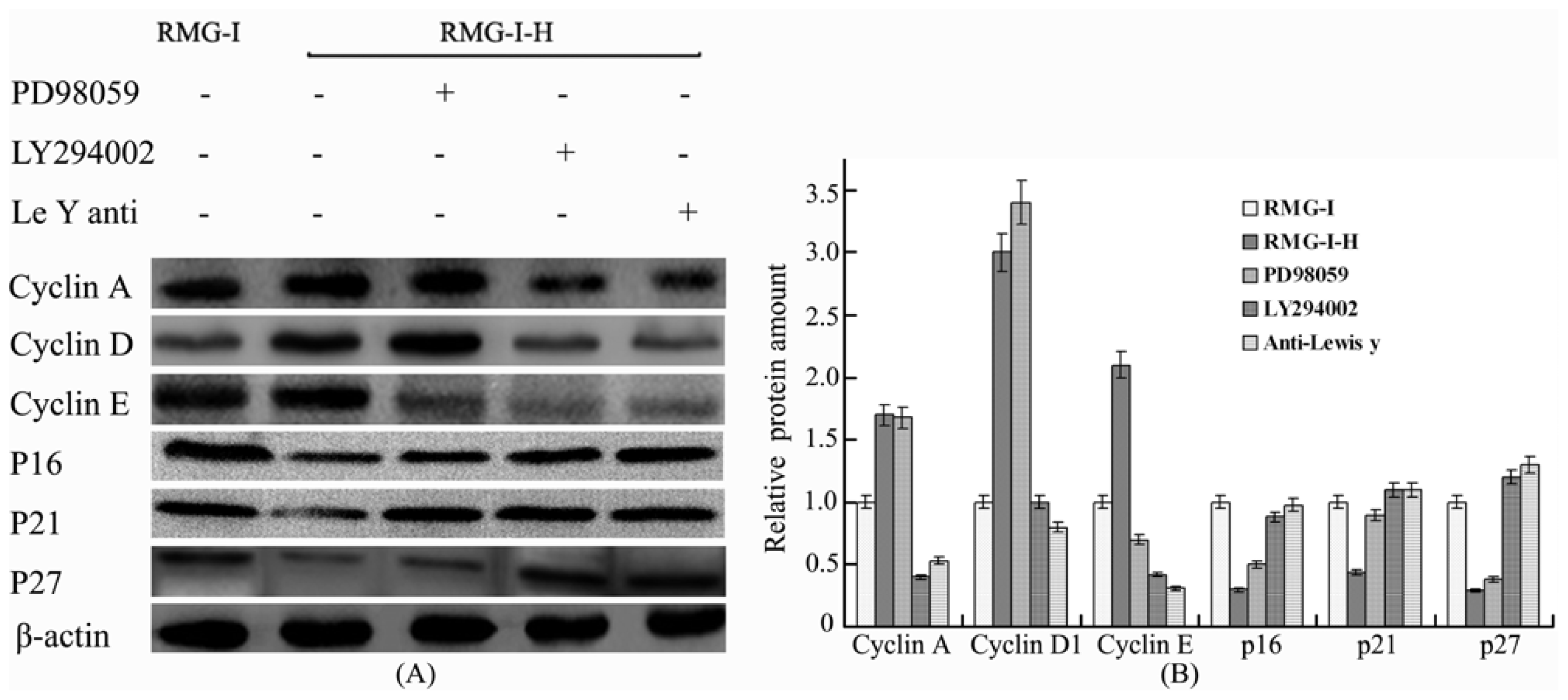
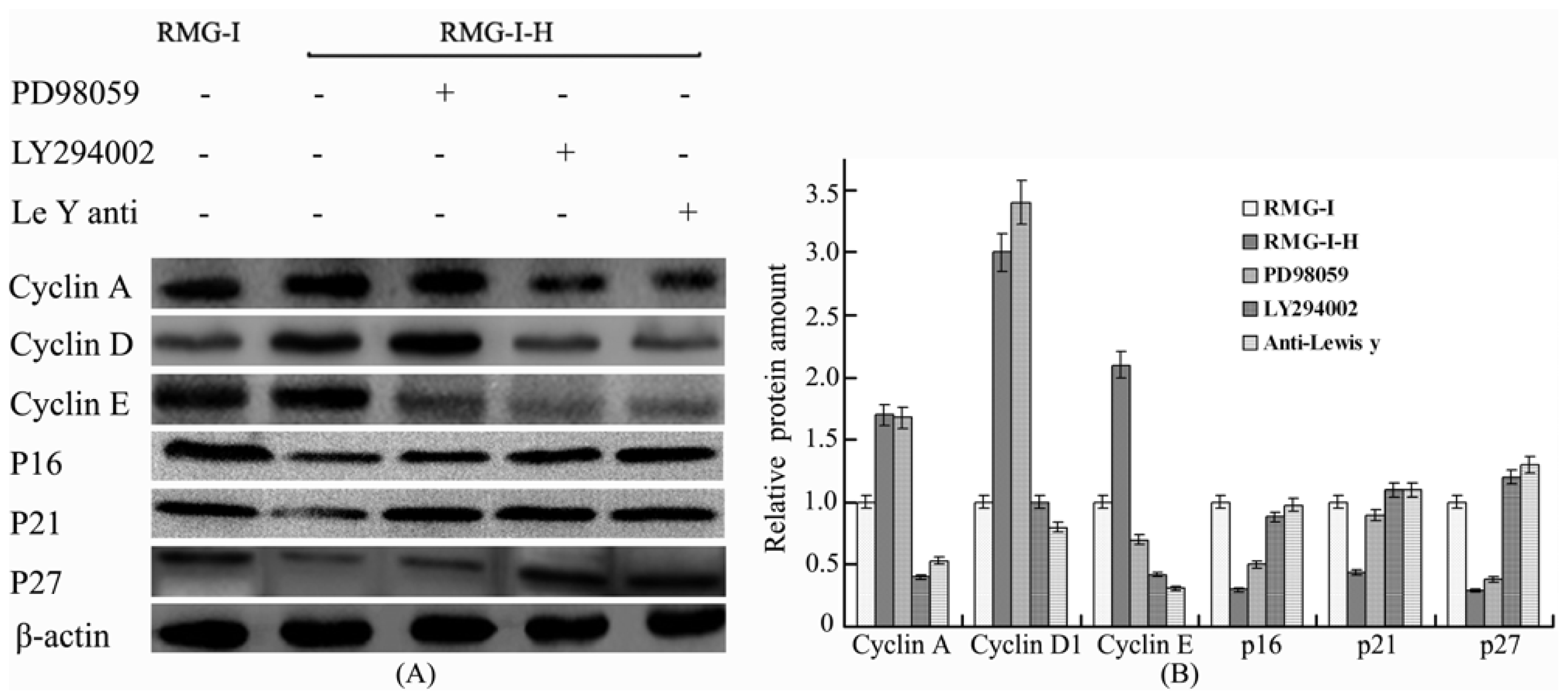
2.4. Lewis Y Antibody Blocking Reduced Lewis Y Overexpression-Mediated Differences in Cyclin and CKI Protein Expression
2.5. The Differences in Cyclin and CKI Protein Expression Levels Were Decreased by Treatment with LY294002 and PD98059 in Lewis Y Overexpressed Ovarian Cancer Cells
3. Discussion
4. Materials and Method
4.1. Reagents and Cell Lines
4.2. Cell Culture
4.3. Analysis the Effect of Lewis Y Antigen on Cell Proliferation
4.4. Determination and Real-Time PCR
4.5. Analysis of the Proteins of Cell Cycle Relation Factors with Western Blotting
4.6. Statistical Analysis
5. Conclusions
Acknowledgements
Reference
- Hao, Y.Y.; Lin, B.; Zhang, Y.H.; Li, F.F.; Diao, B.; Zhang, S.L. α1,2-Fucosyltransferase gene translection in fluences on biological behavior of ovarian carcinoma-derived RMG-1 cells (in Chinese). Fen Zi Xi Bao Sheng Wu Xue Bao 2008, 41, 435–442. [Google Scholar]
- Sherr, C.J. Mammalian G1 cyclins and cell cycle progression. Proc. Assoc. Am. Physicians 1995, 107, 181–186. [Google Scholar]
- Ruas, M.; Peters, G. The p16INK4a/CDKN2A tumor suppressor and its relatives. Biochim. Biophys. Acta 1998, 1378, F115–F177. [Google Scholar]
- Sherr, C.J. The Pezcoller lecture: Cancer cell cycles revisited. Cancer Res 2000, 60, 3689–3695. [Google Scholar]
- Liu, J.J.; Lin, B.; Hao, Y.Y.; Li, F.F.; Liu, D.W.; Qi, Y.; Zhu, L.C.; Zhang, S.L.; Iwamori, M. Lewis(y) antigen stimulates the growth of ovarian cancer cells via regulation of the epidermal growth factor receptor pathway. Oncol. Rep 2010, 23, 833–841. [Google Scholar]
- Liu, D.W.; Liu, J.J.; Wang, C.Z.; Lin, B.; Liu, Q.; Hao, Y.Y.; Zhang, S.L.; Iwamori, M. The Stimulation of IGF-1R Expression by Lewis(y) Antigen Provides a Powerful Development Mechanism of Epithelial Ovarian Carcinoma. Int. J. Mol. Sci 2011, 12, 6781–6795. [Google Scholar]
- Yan, L.M.; Lin, B.; Zhu, L.C.; Hao, Y.Y.; Qi, Y.; Wang, C.Z.; Gao, S.; Liu, S.C.; Zhang, S.L.; Iwamori, M. Enhancement of the adhesive and spreading potentials of ovarian carcinoma RMG-1 cells due to increased expression of integrin alpha5 beta1 with the Lewis Y-structure on transfection of the alpha1,2-fucosyltransferase gene. Biochimie 2010, 92, 852–857. [Google Scholar]
- Liu, J.J.; Lin, B.; Hao, Y.Y.; Qi, Y.; Zhu, L.C.; Li, F.F.; Liu, D.W.; Cong, J.P.; Zhang, S.L.; Iwamori, M. Lewis; (y) antigen promotes the proliferation of ovarian carcinoma-derived RMG-I cell through the PI3K/Akt signaling pathway. In J. Exp. Clin. Cancer Res; 2009; Volume 28, pp. 154:1–154:12. [Google Scholar]
- Coolican, S.A.; Samuel, D.S.; Ewton, D.Z.; McWade, F.J.; Florini, J.R. The mitogenic and myogenic actions of insulin-like growth factors utilize distinct signaling pathways. J. Biol. Chem 1997, 272, 6653–6662. [Google Scholar]
- Meloche, S.; Pouysségur, J. The ERK1/2 mitogen-activated protein kinase pathway as a master regulator of the G1- to S-phase transition. Oncogene 2007, 26, 3227–3239. [Google Scholar]
- Girnita, L.; Shenoy, S.K.; Sehat, B.; Vasilcanu, R.; Vasilcanu, D.; Girnita, A.; Lefkowitz, RJ.; Larsson, O. Beta-arrestin and Mdm2 mediate IGF-1 receptor-stimulated ERK activation and cell cycle progression. J. Biol. Chem 2007, 282, 11329–11338. [Google Scholar]
- Halevy, O.; Cantley, L.C. Differential regulation of the phosphoinositide 3-kinase and MAP kinase pathways by hepatocyte growth factor vs. insulin-like growth factor-I in myogenic cells. Exp. Cell Res 2004, 297, 224–234. [Google Scholar]
- Rommel, C.; Bodine, S.C.; Clarke, B.A.; Rossman, R.; Nunez, L.; Stitt, T.N.; Yancopoulos, G.D.; Glass, DJ. Mediation of IGF-1-induced skeletal myotube hypertrophy by PI(3)K/Akt/mTOR and PI(3)K/Akt/GSK3 pathways. Nat. Cell Biol 2001, 3, 1009–1013. [Google Scholar]
- Kornasio, R.; Riederer, I.; Butler-Browne, G.; Mouly, V.; Uni, Z.; Halevy, O. β-hydroxy-β-methylbutyrate (HMB) stimulates myogenic cell proliferation, differentiation and survival via the MAPK/ERK and PI3K/Akt pathways. Biochim. Biophys. Acta 2009, 1793, 755–763. [Google Scholar]
- Yang, T.Y.; Chang, G.C.; Chen, K.C.; Hung, H.W.; Hsu, K.H.; Sheu, G.T.; Hsu, S.L. Sustained activation of ERK and Cdk2/cyclin-A signaling pathway by pemetrexed leading to S-phase arrest and apoptosis in human non-small cell lung cancer A549 cells. Eur. J. Pharmacol 2011, 663, 17–26. [Google Scholar]
- Chen, J.; Zeng, J.; Xin, M.; Huang, W.; Chen, X. Formononetin induces cell cycle arrest of human breast cancer cells via IGF1/PI3K/Akt pathways in vitro and in vivo. Horm. Metab. Res 2011, 43, 681–686. [Google Scholar]
- Tocchi, A.; Mazzoni, G.; Fornasari, V.; Miccini, M.; Daddi, G.; Tagliacozzo, S. Preservation of the inferior mesenteric artery in colorectal resection for complicated diverticular disease. Am. J. Surg 2001, 182, 162–167. [Google Scholar]
- Koroleva, O.A.; Tomlinson, M.; Parinyapong, P.; Sakvarelidze, L.; Leader, D.; Shaw, P.; Doonan, J.H. CycD1, a putative G1 cyclin from Antirrhinum majus, accelerates the cell cycle in cultured tobacco BY-2 cells by enhancing both G1/S entry and progression through S and G2 phases. Plant Cell 2004, 16, 2364–2379. [Google Scholar]
- Ohtsubo, M.; Theodoras, A.M.; Schumacher, J.; Roberts, J.M.; Pagano, M. Human cyclin E, a nuclear protein essential for the G1-to-S phase transition. Mol. Cell. Biol 1995, 15, 2612–2624. [Google Scholar]
- Lee, M.H.; Yang, H.Y. Regulators of G1 cyclin-dependent kinases and cancers. Cancer Metastasis Rev 2003, 22, 435–449. [Google Scholar]
- Vidal, A.; Koff, A. Cell-cyclin inhibitors: Three families united by common cause. Gene 2000, 247, 1–15. [Google Scholar]
- Polyak, K.; Lee, M.H.; Erdjurment-Bromage, H.; Koff, A.; Roberts, J.M.; Tempst, P.; Massagué, J. Cloning of p27 Kip1, a cyclin-dependent kinase inhibitor and a potential mediator of extracellular antimitogenic signals. Cell 1994, 78, 59–66. [Google Scholar]
- Abbas, T.; Dutta, A. p21 in cancer: Intricate networks and multiple activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar]
- Kashima, H.; Shiozawa, T.; Miyamoto, T.; Suzuki, A.; Uchikawa, J.; Kurai, M.; Konishi, I. Autocrine stimulation of IGF1 in estrogen-induced growth of endometrial carcinoma cells: Involvement of the mitogen-activated protein kinase pathway followed by up-regulation of cyclin D1 and cyclin E. Endocr. Relat. Cancer 2009, 16, 113–122. [Google Scholar]
- Burch, L.R.; Scott, M.; Pohler, E.; Meek, D.; Hupp, T. Phage-peptide display identifies the interferon-responsive, death-activated protein kinase family as a novel modifier of MDM2 and p21WAF1. J. Mol. Biol 2004, 337, 115–128. [Google Scholar]
- Polyak, K.; Kato, J.Y.; Solomon, M.J.; Sherr, C.J.; Massague, J.; Roberts, J.M.; Koff, A. p27Kip1, a cyclin-CDK inhibitor; links transforming growth factor-b and contact inhibiton to cell cycle arrest. Genes Dev 1994, 8, 9–22. [Google Scholar]
- Toyoshima, H.; Hunter, T. P27, a novel inhibitor of G1 cyclin-cdk protein kinase activity, is related to p21. Cell 1994, 78, 67–74. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene name | Sequence of primers(5′-3′) | Product length (bp) | Annealing temperature (°C) |
|---|---|---|---|
| Cyclin A | F: AGGTACTGAAGTCCGGGAAC R: GTGACATGCTCATCATTTACAGGAA | 106 | 62 |
| Cyclin D1 | F: TGATGCTGGGCACTTCATCTG R: TCCAATCATCCCGAATGAGAGTC | 177 | 63 |
| Cyclin E | F: TTTGCAGGATCCAGATCAAGA R: CACAGACTGCATTATTGTCCCAAG | 92 | 60 |
| CDK2 | F: CTCCACCGAGACCTTAAACCTCAG R: TCGGTACCACAGGGTCACCA | 138 | 60 |
| CDK4 | F: CTTCTGCAGTCCACATATGCAACA R: CAACTGGTCGGCTTCAGAGTTTC | 114 | 60 |
| CDK6 | F: GATCTCTGGAGTGTTGGCTGCATA R: GGCAACATCTCTAGGCCAGTCTTC | 144 | 62 |
| P16 | F: GGCACCAGAGGCAGTAACCA R: GGACCTTCGGTGACTGATGATCTAA | 126 | 63 |
| P21 | F: AAGACCATGTGGACCTGTCACTGT R: GAAGATCAGCCGGCGTTTG | 155 | 62 |
| P27 | F: CAAATGCCGGTTCTGTGGAG R: TCCATTCCATGAAGTCAGCGATA | 177 | 63 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, D.; Liu, J.; Lin, B.; Liu, S.; Hou, R.; Hao, Y.; Liu, Q.; Zhang, S.; Iwamori, M. Lewis y Regulate Cell Cycle Related Factors in Ovarian Carcinoma Cell RMG-I in Vitro via ERK and Akt Signaling Pathways. Int. J. Mol. Sci. 2012, 13, 828-839. https://doi.org/10.3390/ijms13010828
Liu D, Liu J, Lin B, Liu S, Hou R, Hao Y, Liu Q, Zhang S, Iwamori M. Lewis y Regulate Cell Cycle Related Factors in Ovarian Carcinoma Cell RMG-I in Vitro via ERK and Akt Signaling Pathways. International Journal of Molecular Sciences. 2012; 13(1):828-839. https://doi.org/10.3390/ijms13010828
Chicago/Turabian StyleLiu, Dawo, Juanjuan Liu, Bei Lin, Shuice Liu, Rui Hou, Yingying Hao, Qing Liu, Shulan Zhang, and Masao Iwamori. 2012. "Lewis y Regulate Cell Cycle Related Factors in Ovarian Carcinoma Cell RMG-I in Vitro via ERK and Akt Signaling Pathways" International Journal of Molecular Sciences 13, no. 1: 828-839. https://doi.org/10.3390/ijms13010828