Recombinant Cytokines from Plants


Abstract
:1. Introduction
2. Cytokines and Their Therapeutic Application
3. Recombinant Cytokines from Plant-Based Platforms
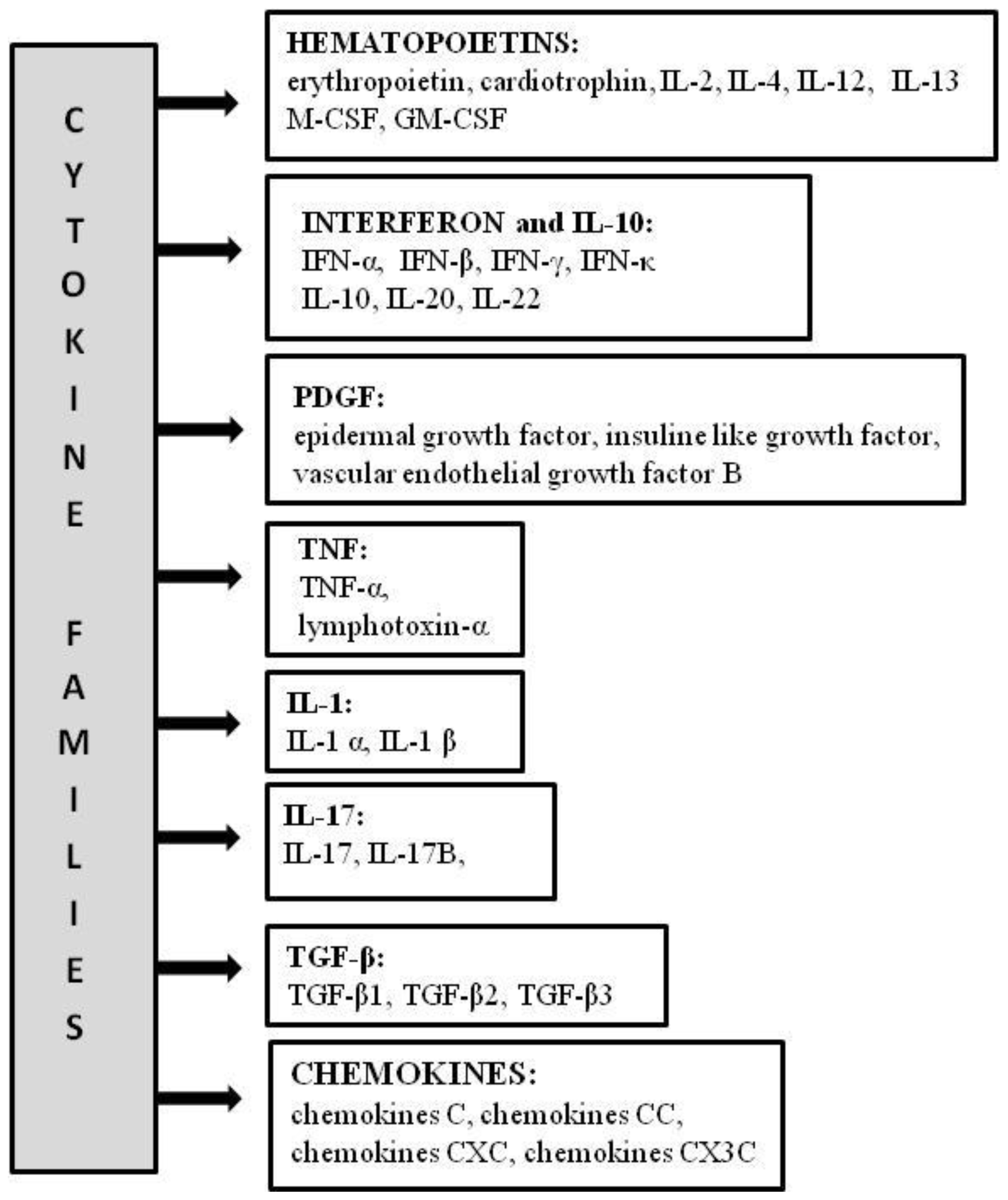
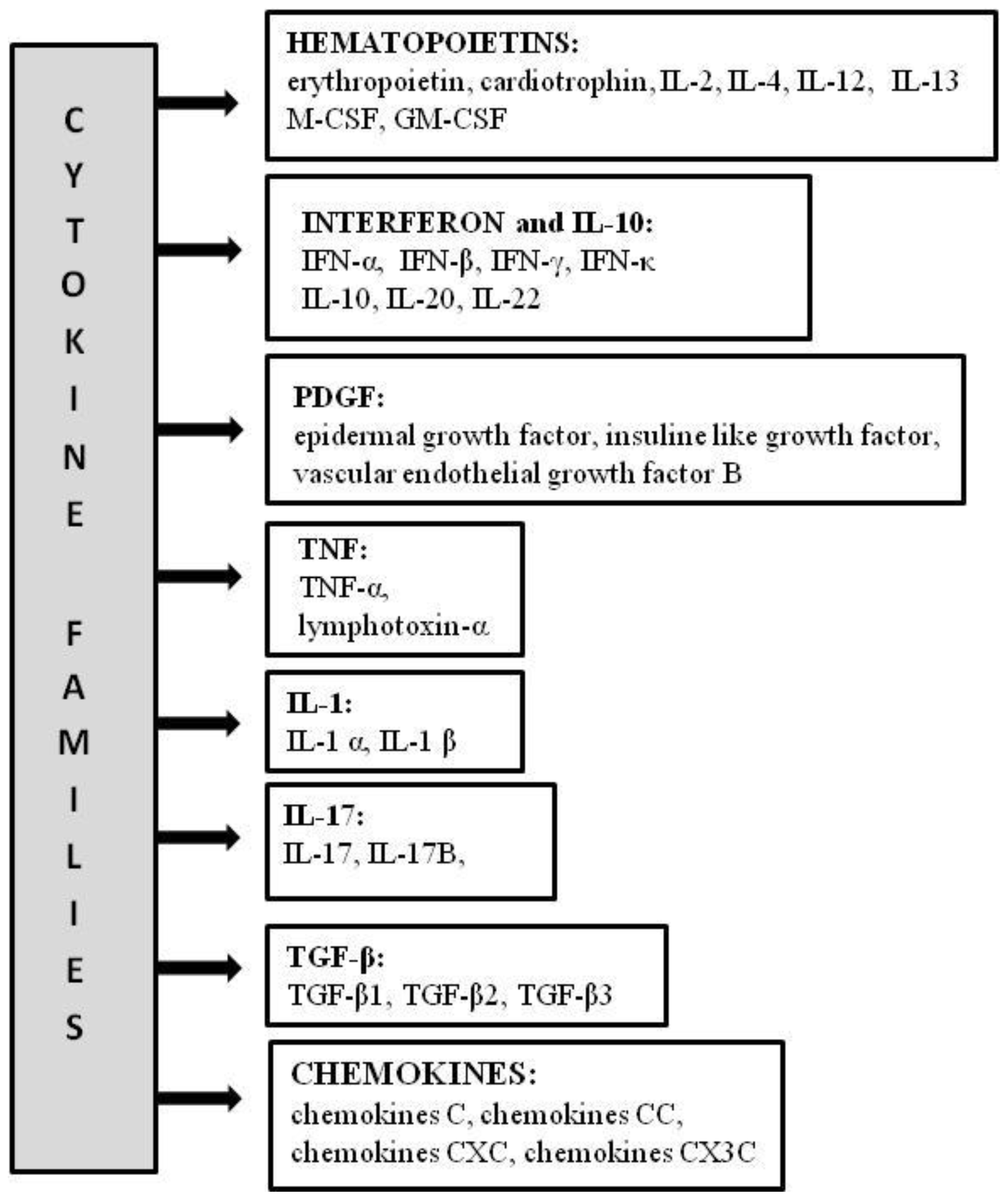
3.1. Hematopoietin Family
3.2. Interferon and IL-10 Family
3.3. Other Families
4. Prospects for Commercialization Plant-Produced Cytokines
4.1. Strategies Used to Improve the Performance of Plant-Base Production Platforms
4.2. Other Problems to Consider
5. Conclusions
Acknowledgements
References
- Barta, A; Sommergruber, K; Thompson, D; Hartmuth, K; Matzke, MA; Matzke, AJM. The expression of a nopaline synthase—human growth hormone chimaeric gene in transformed tobacco and sunflower callus tissue. Plant Mol. Biol 1986, 6, 347–357. [Google Scholar]
- Sijmons, PC; Dekker, BM; Schrammeijer, B; Verwoerd, TC; van den Elzen, PJ; Hoekema, A. Production of correctly processed human serum albumin in transgenic plants. Nat. Biotechnol 1990, 8, 217–221. [Google Scholar]
- Obembe, OO; Popoola, JO; Leelavathi, S; Reddy, SV. Advances in plant molecular farming. Biotechnol. Adv 2011, 29, 210–222. [Google Scholar]
- Homepage of KEGG: Koto Encyclopedia of Genes and Genomes. Available online: http://www.genome.jp/kegg/ accessed on 19 May 2011.
- Homepage of Cytokines & Cells Online Pathfinder Encyclopedia. Available online: http://www.copewithcytokines.org/cope.cgi accessed on 19 May 2011.
- Andersson, U; Tracey, KJ. Molecular basis of applied biological therapeutics. J. Intern. Med 2011, 269, 2–7. [Google Scholar]
- Cutler, A; Brombacher, F. Cytokine therapy. Ann. N. Y. Acad. Sci 2005, 1056, 16–29. [Google Scholar]
- Tayal, V; Kalra, BS. Cytokines and anti-cytokines as therapeutics-an update. Eur. J. Pharmacol 2008, 579, 1–12. [Google Scholar]
- Toka, FN; Pack, CD; Rouse, BT. Molecular adjuvants for mucosal immunity. Immunol. Rev 2004, 199, 100–112. [Google Scholar]
- Elias, EG; Zapas, JL; McCarron, EC; Beam, SL; Hasskamp, JH; Culpepper, WJ. Sequential administration of GM-CSF (Sargramostim) and IL-2 +/− autologous vaccine as adjuvant therapy in cutaneous melanoma: An interim report of a phase II clinical trial. Cancer Biother. Radiopharm 2008, 23, 285–291. [Google Scholar]
- Batista-Duharte, A; Lindblad, EB; Oviedo-Orta, E. Progress in understanding adjuvant immunotoxicity mechanisms. In Toxicol Lett; 2011; Volume 203, pp. 97–105. [Google Scholar]
- Williams, P; Galipeau, J. GMCSF-Interleukin fusion cytokines induce novel immune effectors that can serve as biopharmaceuticals for treatment of autoimmunity and cancer. J. Intern. Med 2011, 269, 74–84. [Google Scholar]
- Matsumoto, S; Ikura, K; Ueda, M; Sasaki, R. Characterization of a human glycoprotein (erythropoietin) produced in cultured tobacco cells. Plant Mol. Biol 1995, 27, 1163–1172. [Google Scholar]
- Hong, S-Y; Kwon, T-H; Lee, J-H; Jang, Y-S; Yang, M-S. Production of biologically active hG-CSF by transgenic plant cell suspension culture. Enzyme Microb. Technol 2002, 30, 762–767. [Google Scholar]
- James, EA; Wang, C; Wang, Z; Reeves, R; Shin, JH; Magnuson, NS; Lee, JM. Production and characterization of biologically active human GM-CSF secreted by genetically modified plant cells. Protein Expr. Purif 2000, 19, 131–138. [Google Scholar]
- Lee, JH; Kim, NS; Kwon, TH; Jang, YS; Yang, MS. Increased production of human granulocyte-macrophage colony stimulating factor (hGM-CSF) by the addition of stabilizing polymer in plant suspension cultures. J. Biotechnol 2002, 96, 205–211. [Google Scholar]
- Shin, YJ; Hong, SY; Kwon, TH; Jang, YS; Yang, MS. High level of expression of recombinant human granulocyte-macrophage colony stimulating factor in transgenic rice cell suspension culture. Biotechnol. Bioeng 2003, 82, 778–783. [Google Scholar]
- Kim, NS; Kim, TG; Jang, YS; Shin, YJ; Kwon, TH; Yang, MS. Amylase gene silencing by RNA interference improves recombinant hGM-CSF production in rice suspension culture. Plant Mol. Biol 2008, 68, 369–377. [Google Scholar]
- Kim, NS; Kim, TG; Kim, OH; Ko, EM; Jang, YS; Jung, ES; Kwon, TH; Yang, MS. Improvement of recombinant hGM-CSF production by suppression of cysteine proteinase gene expression using RNA interference in a transgenic rice culture. Plant Mol. Biol 2008, 68, 263–275. [Google Scholar]
- Kim, TG; Lee, HJ; Jang, YS; Shin, YJ; Kwon, TH; Yang, MS. Co-expression of proteinase inhibitor enhances recombinant human granulocyte-macrophage colony stimulating factor production in transgenic rice cell suspension culture. Protein Expr. Purif 2008, 61, 117–121. [Google Scholar]
- Wang, ML; Goldstein, C; Su, W; Moore, PH; Albert, HH. Production of biologically active GM-CSF in sugarcane: A secure biofactory. Transgenic Res 2005, 14, 167–178. [Google Scholar]
- Sardana, RK; Alli, Z; Dudani, A; Tackaberry, E; Panahi, M; Narayanan, M; Ganz, P; Altosaar, I. Biological activity of human granulocyte-macrophage colony stimulating factor is maintained in a fusion with seed glutelin peptide. Transgenic Res 2002, 11, 521–531. [Google Scholar]
- Sardana, R; Dudani, AK; Tackaberry, E; Alli, Z; Porter, S; Rowlandson, K; Ganz, P; Altosaar, I. Biologically active human GM-CSF produced in the seeds of transgenic rice plants. Transgenic Res 2007, 16, 713–721. [Google Scholar]
- Ning, T; Xie, T; Qiu, Q; Yang, W; Zhou, S; Zhou, L; Zheng, C; Zhu, Y; Yang, D. Oral administration of recombinant human granulocyte-macrophage colony stimulating factor expressed in rice endosperm can increase leukocytes in mice. Biotechnol. Lett 2008, 30, 1679–1686. [Google Scholar]
- Gora-Sochacka, A; Redkiewicz, P; Napiorkowska, B; Gaganidze, D; Brodzik, R; Sirko, A. Recombinant mouse granulocyte-macrophage colony-stimulating factor is glycosylated in transgenic tobacco and maintains its biological activity. J. Interferon Cytokine Res 2010, 30, 135–142. [Google Scholar]
- Zhou, F; Wang, ML; Albert, HH; Moore, PH; Zhu, YJ. Efficient transient expression of human GM-CSF protein in Nicotiana benthamiana using potato virus X vector. Appl. Microbiol. Biotechnol 2006, 72, 756–762. [Google Scholar]
- Magnuson, NS; Linzmaier, PM; Reeves, R; An, G; HayGlass, K; Lee, JM. Secretion of biologically active human interleukin-2 and interleukin-4 from genetically modified tobacco cells in suspension culture. Protein Expr. Purif 1998, 13, 45–52. [Google Scholar]
- Park, Y; Cheong, H. Expression and production of recombinant human interleukin-2 in potato plants. Protein Expr. Purif 2002, 25, 160–165. [Google Scholar]
- Morandini, F; Avesani, L; Bortesi, L; van Droogenbroeck, B; de Wilde, K; Arcalis, E; Bazzoni, F; Santi, L; Brozzetti, A; Falorni, A; et al. Non-food/feed seeds as biofactories for the high-yield production of recombinant pharmaceuticals. Plant Biotechnol J 2011. [Google Scholar] [CrossRef]
- Patel, J; Zhu, H; Menassa, R; Gyenis, L; Richman, A; Brandle, J. Elastin-like polypeptide fusions enhance the accumulation of recombinant proteins in tobacco leaves. Transgenic Res 2007, 16, 239–249. [Google Scholar]
- Ma, S; Huang, Y; Davis, A; Yin, Z; Mi, Q; Menassa, R; Brandle, JE; Jevnikar, AM. Production of biologically active human interleukin-4 in transgenic tobacco and potato. Plant Biotechnol. J 2005, 3, 309–318. [Google Scholar]
- Menassa, R; Kennette, W; Nguyen, V; Rymerson, R; Jevnikar, A; Brandle, J. Subcellular targeting of human interleukin-10 in plants. J. Biotechnol 2004, 108, 179–183. [Google Scholar]
- Fujiwara, Y; Aiki, Y; Yang, L; Takaiwa, F; Kosaka, A; Tsuji, NM; Shiraki, K; Sekikawa, K. Extraction and purification of human interleukin-10 from transgenic rice seeds. Protein Expr. Purif 2010, 72, 125–130. [Google Scholar]
- Bortesi, L; Rossato, M; Schuster, F; Raven, N; Stadlmann, J; Avesani, L; Falorni, A; Bazzoni, F; Bock, R; Schillberg, S; et al. Viral and murine interleukin-10 are correctly processed and retain their biological activity when produced in tobacco. BMC Biotechnol 2009, 9, 22. [Google Scholar] [Green Version]
- Gutierrez-Ortega, A; Avila-Moreno, F; Saucedo-Arias, LJ; Sanchez-Torres, C; Gomez-Lim, MA. Expression of a single-chain human interleukin-12 gene in transgenic tobacco plants and functional studies. Biotechnol. Bioeng 2004, 85, 734–740. [Google Scholar]
- Gutierrez-Ortega, A; Sandoval-Montes, C; de Olivera-Flores, TJ; Santos-Argumedo, L; Gomez-Lim, MA. Expression of functional interleukin-12 from mouse in transgenic tomato plants. Transgenic Res 2005, 14, 877–885. [Google Scholar]
- Sanchez-Hernandez, C; Gutierrez-Ortega, A; Aguilar-Leon, D; Hernandez-Pando, R; Gomez-Lim, M; Gomez-Garcia, B. In vivo activity of plant-based interleukin-12 in the lung of Balb/c mouse. BMC Res. Notes 2010, 3, 151. [Google Scholar]
- Kwon, TH; Seo, JE; Kim, J; Lee, JH; Jang, YS; Yang, MS. Expression and secretion of the heterodimeric protein interleukin-12 in plant cell suspension culture. Biotechnol. Bioeng 2003, 81, 870–875. [Google Scholar]
- Wang, DJ; Brandsma, M; Yin, Z; Wang, A; Jevnikar, AM; Ma, S. A novel platform for biologically active recombinant human interleukin-13 production. Plant Biotechnol. J 2008, 6, 504–515. [Google Scholar]
- Zhang, B; Yang, YH; Lin, YM; Rao, Q; Zheng, GG; Wu, KF. Expression and production of bioactive human interleukin-18 in transgenic tobacco plants. Biotechnol. Lett 2003, 25, 1629–1635. [Google Scholar]
- Farran, I; Rio-Manterola, F; Iniguez, M; Garate, S; Prieto, J; Mingo-Castel, AM. High-density seedling expression system for the production of bioactive human cardiotrophin-1, a potential therapeutic cytokine, in transgenic tobacco chloroplasts. Plant Biotechnol. J 2008, 6, 516–527. [Google Scholar]
- Ohya, K; Matsumura, T; Ohashi, K; Onuma, M; Sugimoto, C. Expression of two subtypes of human IFN-alpha in transgenic potato plants. J. Interferon Cytokine Res 2001, 21, 595–602. [Google Scholar]
- Sawahel, WA. The production of transgenic potato plants expressing human alpha-interferon using lipofectin-mediated transformation. Cell. Mol. Biol. Lett 2002, 7, 19–29. [Google Scholar]
- Arazi, T; Slutsky, SG; Shiboleth, YM; Wang, Y; Rubinstein, M; Barak, S; Yang, J; Gal-On, A. Engineering zucchini yellow mosaic potyvirus as a non-pathogenic vector for expression of heterologous proteins in cucurbits. J. Biotechnol 2001, 87, 67–82. [Google Scholar]
- Arlen, PA; Falconer, R; Cherukumilli, S; Cole, A; Cole, AM; Oishi, KK; Daniell, H. Field production and functional evaluation of chloroplast-derived interferon-alpha2b. Plant Biotechnol. J 2007, 5, 511–525. [Google Scholar]
- Luchakivskaya, Y; Kishchenko, O; Gerasymenko, I; Olevinskaya, Z; Simonenko, Y; Spivak, M; Kuchuk, M. High-level expression of human interferon alpha-2b in transgenic carrot (Daucus carota L.) plants. Plant Cell Rep 2011, 30, 407–415. [Google Scholar]
- Li, J; Chen, M; Liu, X-W; Zhang, H-C; Shen, FF; Wang, GP. Transient expression of an active human interferon-beta in lettuce. Sci. Hortic 2007, 112, 258–265. [Google Scholar]
- Chen, TL; Lin, YL; Lee, YL; Yang, NS; Chan, MT. Expression of bioactive human interferon-gamma in transgenic rice cell suspension cultures. Transgenic Res 2004, 13, 499–510. [Google Scholar]
- Song, L; Zhao, DG; Wu, YJ; Li, Y. Transient expression of chicken alpha interferon gene in lettuce. J. Zhejiang Univ. Sci. B 2008, 9, 351–355. [Google Scholar]
- Fukuzawa, N; Tabayashi, N; Okinaka, Y; Furusawa, R; Furuta, K; Kagaya, U; Matsumura, T. Production of biologically active Atlantic salmon interferon in transgenic potato and rice plants. J. Biosci. Bioeng 2010, 110, 201–207. [Google Scholar]
- Ohya, K; Itchoda, N; Ohashi, K; Onuma, M; Sugimoto, C; Matsumura, T. Expression of biologically active human tumor necrosis factor-alpha in transgenic potato plant. J. Interferon Cytokine Res 2002, 22, 371–378. [Google Scholar]
- Potula, HH; Kathuria, SR; Ghosh, AK; Maiti, TK; Dey, S. Transient expression, purification and characterization of bioactive human fibroblast growth factor 8b in tobacco plants. Transgenic Res 2008, 17, 19–32. [Google Scholar]
- Daniell, H; Ruiz, G; Denes, B; Sandberg, L; Langridge, W. Optimization of codon composition and regulatory elements for expression of human insulin like growth factor-1 in transgenic chloroplasts and evaluation of structural identity and function. BMC Biotechnol 2009, 9, 33. [Google Scholar]
- Xie, T; Qiu, Q; Zhang, W; Ning, T; Yang, W; Zheng, C; Wang, C; Zhu, Y; Yang, D. A biologically active rhIGF-1 fusion accumulated in transgenic rice seeds can reduce blood glucose in diabetic mice via oral delivery. Peptides 2008, 29, 1862–1870. [Google Scholar]
- Medrano, J; Reidy, MJ; Liu, J; Ayala, M; Dolan, MC; Cramer, CL. Faye, L, Gomord, V, Eds.; Rapid system for evaluating bioproduction capacity of complex pharmaceutical proteins in plants. In Methods in Molecular Biology, Recombinant Proteins from Plants; Humana Press: New York, NY, USA, 2009; Volume 483, pp. 41–67. [Google Scholar]
- Edelbaum, O; Stein, D; Holland, N; Gafni, Y; Livneh, O; Novick, D; Rubinstein, M; Sela, I. Expression of active human interferon-beta in transgenic plants. J. Interferon Res 1992, 12, 449–453. [Google Scholar]
- Davies, HM. Review article: Commercialization of whole-plant systems for biomanufacturing of protein products: Evolution and prospects. Plant Biotechnol. J 2010, 8, 845–861. [Google Scholar]
- Ramessar, K; Capell, T; Christou, P. Molecular pharming in cereal crops. Phytochem. Rev 2008, 7, 579–592. [Google Scholar]
- Schillberg, S; Twyman, RM; Fischer, R. Opportunities for recombinant antigen and antibody expression in transgenic plants-technology assessment. Vaccine 2005, 23, 1764–1769. [Google Scholar]
- Sharma, AK; Sharma, MK. Plants as bioreactors: Recent developments and emerging opportunities. Biotechnol. Adv 2009, 27, 811–832. [Google Scholar]
- Luo, J; Ning, T; Sun, Y; Zhu, J; Zhu, Y; Lin, Q; Yang, D. Proteomic analysis of rice endosperm cells in response to expression of hGM-CSF. J. Proteome Res 2009, 8, 829–837. [Google Scholar]
- Sparrow, PA; Irwin, JA; Dale, PJ; Twyman, RM; Ma, JK. Pharma-Planta: Road testing the developing regulatory guidelines for plant-made pharmaceuticals. Transgenic Res 2007, 16, 147–161. [Google Scholar]
- Spok, A; Twyman, RM; Fischer, R; Ma, JK; Sparrow, PA. Evolution of a regulatory framework for pharmaceuticals derived from genetically modified plants. Trends Biotechnol 2008, 26, 506–517. [Google Scholar]
- Lutz, KA; Maliga, P. Construction of marker-free transplastomic plants. Curr. Opin. Biotechnol 2007, 18, 107–114. [Google Scholar]
- Barampuram, S; Zhang, ZJ. Recent advances in plant transformation. Methods Mol. Biol 2011, 701, 1–35. [Google Scholar]
- Paul, M; Ma, JK. Plant-made pharmaceuticals: Leading products and production platforms. Biotechnol. Appl. Biochem 2011, 58, 58–67. [Google Scholar]

{kind=link}
| Cytokine | Disease or Indication | Drug Name (Company) |
|---|---|---|
| G-CSF | Neutropenia | Neupogen/Filgrastim (Hoffmann-La Roche) |
| GM-CSF | Leukemia Bone marrow Stem cell transplants | Leukine/Sargramostin (Bayer) |
| Interferon-α (INF-α) | Chronic hepatitis B Chronic hepatitis C Hairy cell leukemia Chronic myeloid leukemia Condyloma acuminate AIDS-related Kaposi’s sarcoma genital warts | Intron A (Schering Plough) Roferon A (Hoffman La Roche) Infergen (Three Rivers Pharmaceuticals) Alferon N (HEMISPHERx Biopharma) Pegasys (Genentech USA/Roche) Pegintron (Merck) |
| Interferon-β (INF-β) | Relapsing multiple sclerosis | Betaseron (Bayer) Avonex (Biogen Idec) |
| Interferon-γ (INF-γ) | Malignant osteopetrosis Chronic granulomatous disease | Actimmune (Intermune Pharma) |
| Erythropoietin-α (EPO-α) | Anemia due to chronic renal failure HIV infected patients Chemotherapy Primary bone marrow disorders | Eprex (Cilag Jansen) Epogen (Amgen) Procrit (Ortho Biotech) |
| IL-2 | Metastatic renal cell cancer Metastatic melanoma | Aldesleukin (Novartis) Proleukin (Prometheus Laboratories) |
| IL-11 | Thrombocytopenia | Oprelvekin/Neumega (Genetics Institute, Inc./Wyeth/Pfizer) |
| Cytokine * | Method/Plant Material | Selected Elements of the Expression Cassette | Expression Level/Yield | Reference |
|---|---|---|---|---|
| erythropoietin | T/tobacco (BY2) cells suspension | 35S promoter and terminator | 0.0026% TSP; 25 pg/L | [13] |
| G-CSF | T/tobacco cells suspension | 35S promoter with the double enhancer, Ω-translation enhancer; nos terminator | 105 μg/L | [14] |
| GM-CSF | T/tobacco cells suspension | 35S promoter, translation enhancer from TEV, His tag, T7 terminator | 150 μg/L (intercellular); 250 μg/L (secretory) | [15] |
| GM-CSF | T/tobacco cells suspension | 35S promoter with the double enhancer, nos terminator | 180–780 μg/L | [16] |
| GM-CSF | M/rice cells suspension | rice amylase promoter and signal peptide | 129 mg/L (25% secreted proteins) | [17] |
| GM-CSF | M/rice cells suspension rice | amylase promoter and signal peptide, RNAi-mediated silencing of α-amylase gene to 8.2% | 280 mg/L | [18] |
| GM-CSF | M/rice cells suspension | rice amylase promoter and signal peptide, RNAi-mediated silencing of cysteine proteinase | 290 mg/L | [19] |
| GM-CSF | M/rice cells suspension | rice amylase promoter and signal peptide; co-expression of gene encoding synthetic protease inhibitor (SPI-II) | 250 mg/L | [20] |
| GM-CSF | M/sugarcane leaves | MUbi-1 promoter from maize or SCubi-9 from sugarcane | 0.02% TSP | [21] |
| GM-CSF | T/tobacco seeds | Gt1, Gt3 (glutelin) promoters and signal peptide, nos terminator | 0.005–0.03% TSP | [22] |
| GM-CSF | T/tobacco seeds | Gt1 (glutelin) promoter and signal peptide, nos terminator | 1.3% TSP | [23] |
| GM-CSF | T/rice seeds | Gt13a (glutelin) promoter (specific for seed endosperm) and glutelin signal peptide, nos terminator, codon optimalization | 0.5–14 μg/seed | [24] |
| Murine GM-CSF | T/tobacco leaves | RbcS1 Promoter; signal peptide, KDEL | 19 μg/g fresh leaves; 0.22% | [25] |
| GM-CSF | V/N. benthamiana leaves | PVX-derived vector: 35S promoter, His tag | 0.2–2% TSP | [26] |
| IL-2 | T/tobacco cells suspension | 35S promoter, T7 terminator | 0.09 mg/L | [27] |
| IL-2 | T/potato tubers | patatin promoter; nos terminator | 115 U/mg TSP | [28] |
| Murine IL-2 | T/Arabidopsis seeds T/tobacco seeds | novel binary Gateway vector (pPphasGW) containing β-phaseolin promoter from common bean and the signal peptide of the Arabidopsis 2S2 seed storage protein gene; KDEL | Much higher yield in Arabidopsis than in tobacco: 0.3 mg/g of seeds (0.7% TSP); biologically active in vitro | [29] |
| IL-4 | T/tobacco leaves | 35S promoter with double enhancer, t-CUP-translation enhancer, ELP, KDEL, nos terminator | 0.086% TSP | [30] |
| IL-4 | T/tobacco leaves, T/potato tubers | 35S promoter, KDEL sequence | 0.1% TSP in tobacco; 0.08% TSP in potato | [31] |
| IL-4 | T/tobacco cells suspension | 35S CaMV promoter, T7 terminator | 0.45 mg/L | [27] |
| IL-10 | T/tobacco leaves | 35S promoter with double enhancer, t-CUP-translation enhancer, ELP, KDEL, nos terminator | 0.27% TSP | [30] |
| IL-10 | T/tobacco leaves |
|
| [32] |
| IL-10 | T/rice seeds | GluB-1 promoter and signal peptide, His tag, KDEL, codon optimization | 2 mg of pure IL-10 per 40 g of rice powder | [33] |
| IL-10 | T/tobacco leaves | 35S promoter with double enhancer; three constructs for each viral IL-10 or murine IL-10, ER-targeted, plasma membrane (IL-10 facing the apoplast), ER-membrane (IL-10 facing the cytosol) assayed in transient expression system, cassettes for ER-targeted cytokines were used for the stable expression | Viral: 10.8 μg/g fresh leaves Murine: 37.0 μg/g fresh leaves | [34] |
| IL-12 | T/tobacco leaves | 35S promoter and terminator | 40 ng/g | [35] |
| IL-12 | T/tomato leaves and fruits | 35S promoter with double enhancer; 35S terminator | 7.3 μg/g leaves 4.3 μg/g fruits | [36,37] |
| IL-12 | T/tobacco cells suspension | 35S promoter with double enhancer; Ω-translation enhancer | 175 μg/L | [38] |
| IL-13 | T/tobacco leaves | double 35S promoter; translation enhancer from AMV; KDEL; nos terminator | 0.15% TSP | [39] |
| IL-18 | T/tobacco leaves | 35S promoter with double enhancer, Ω-translation enhancer, nos terminator | 0.004–0.051% TSP; 351 ng/g | [40] |
| cardiotrophin-1 | M/tobacco leaves, chloroplasts transformation |
|
| [41] |
| IFN-α2b IFN-α8 | T/potato | - | 560 IU/g of tissue | [42] |
| IFNα | L/tomato (leaf tissue and cells suspension) | P1 portion of the dual “bi-directional” promoter from A. tumefaciens cDNA, polyadenylation signal from A. tumefaciens gene-7 | 923–3029 U/g FW tissue | [43] |
| IFN-α2 | V/squash (Cucurbita pepo) and cucumber (Cucumis sativus) | Viral vector dirived from attenuated zucchini yellow mosaic potyvirus (AG) | max. 430,000 IU/FW of leaves | [44] |
| IFN-α2b | M/tobacco leaves, chloroplasts transformation | Cassette: 5′UTR/HIS/THR/IFNα2b cloned into the chloroplast vector pLD-CtV | 3 mg/g, 20% TSP | [45] |
| IFN-α2b | T/carrot leaves |
| Biological activity on average:
| [46] |
| IFNβ | transient, agroinfiltration of the leaves of lettuce | 35S promoter, nos terminator | 3.1 × 104 IU/mL | [47] |
| IFNγ | T/rice cells suspension |
|
| [48] |
| Chicken IFN-α | Transient expression, agroinfiltration of the leaves of lettuce | 35S promoter, nos terminator | 0.393 μg/kg tissue, 0.0004% TSP | [49] |
| Fish IFN-α1 | T/rice T/potato | 35S promoter with double enhancer, nos terminator | Biological activity: in rice-up to 0.82 U/mg leaves; in potato-up to 5.4 U/mg leaves | [50] |
| TNF-α | T/potato | 35S promoter, Ω-translation enhancer, SEKDEL sequence | 15 μg/g tissue | [51] |
| Fibroblast growth factor 8 isoform b (FGF8b) | T/tobacco leaves | 35S promoter with double enhancer; 35S terminator, c-myc, His, KDEL | 4.1% TSP | [52] |
| Insulin like growth factor 1 (IGF-1) | M/tobacco, transplastomic | psbA promoter, translantion enhancer (5′UTR psbA) and ZZ-tag from S. aureus, codon optimization | up to 32% TSP | [53] |
| Insulin like growth factor 1 (IGF-1) | M/rice seeds | Glutelin (Gt13a) promoter, the Gt13a signal peptide in frame with the fusion protein containing IGF-1 attached to the C-terminus of ER luminal binding protein (BipC), nos terminator | up to 6.8% of total seed protein; biologically active in vivo (effectively reduced blood glucose in diabetic mice) | [54] |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sirko, A.; Vaněk, T.; Góra-Sochacka, A.; Redkiewicz, P. Recombinant Cytokines from Plants. Int. J. Mol. Sci. 2011, 12, 3536-3552. https://doi.org/10.3390/ijms12063536
Sirko A, Vaněk T, Góra-Sochacka A, Redkiewicz P. Recombinant Cytokines from Plants. International Journal of Molecular Sciences. 2011; 12(6):3536-3552. https://doi.org/10.3390/ijms12063536
Chicago/Turabian StyleSirko, Agnieszka, Tomas Vaněk, Anna Góra-Sochacka, and Patrycja Redkiewicz. 2011. "Recombinant Cytokines from Plants" International Journal of Molecular Sciences 12, no. 6: 3536-3552. https://doi.org/10.3390/ijms12063536