Cigarette Smoke Affects ABCAl Expression via Liver X Receptor Nuclear Translocation in Human Keratinocytes

Abstract
:1. Introduction
2. Results and Discussion
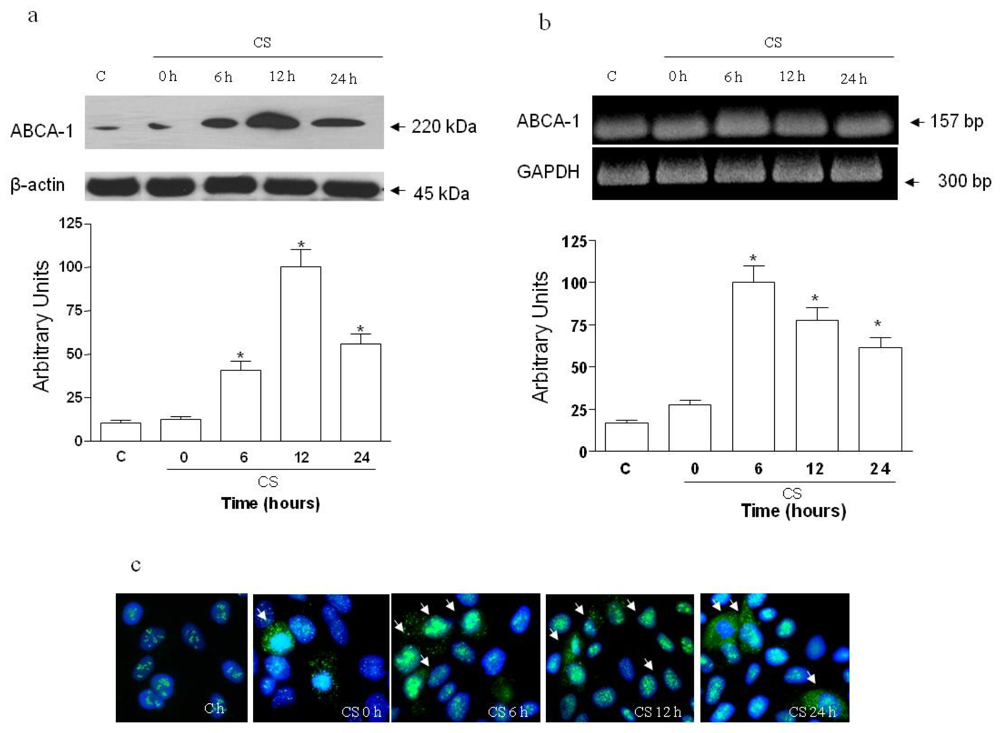
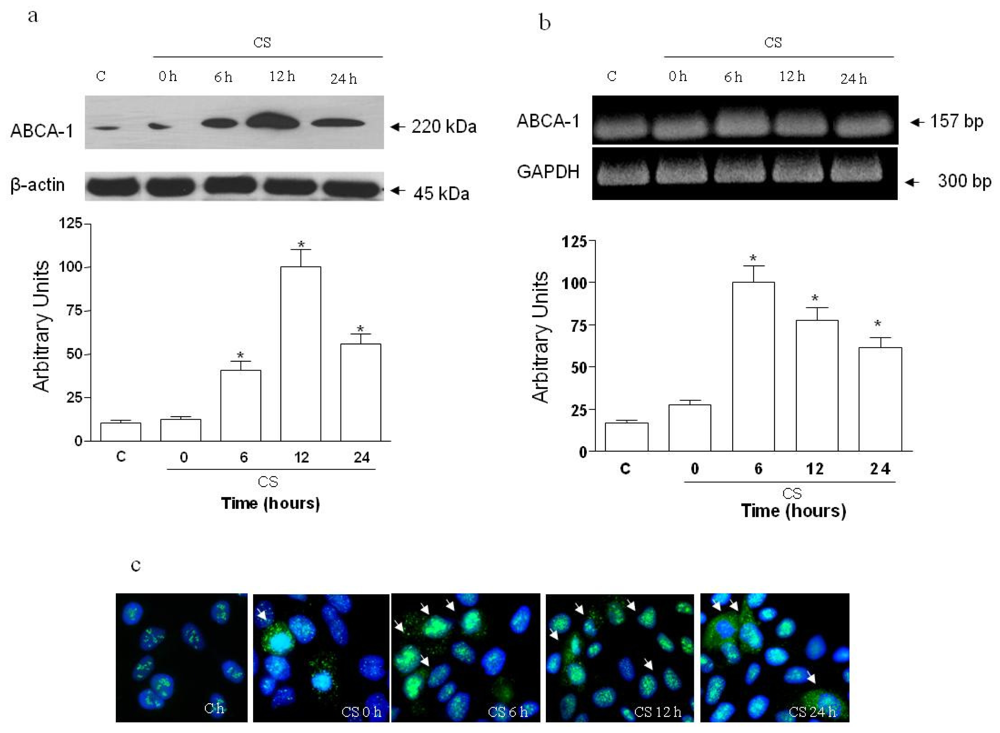
2.1. The Effect of CS on ABCA1 Levels
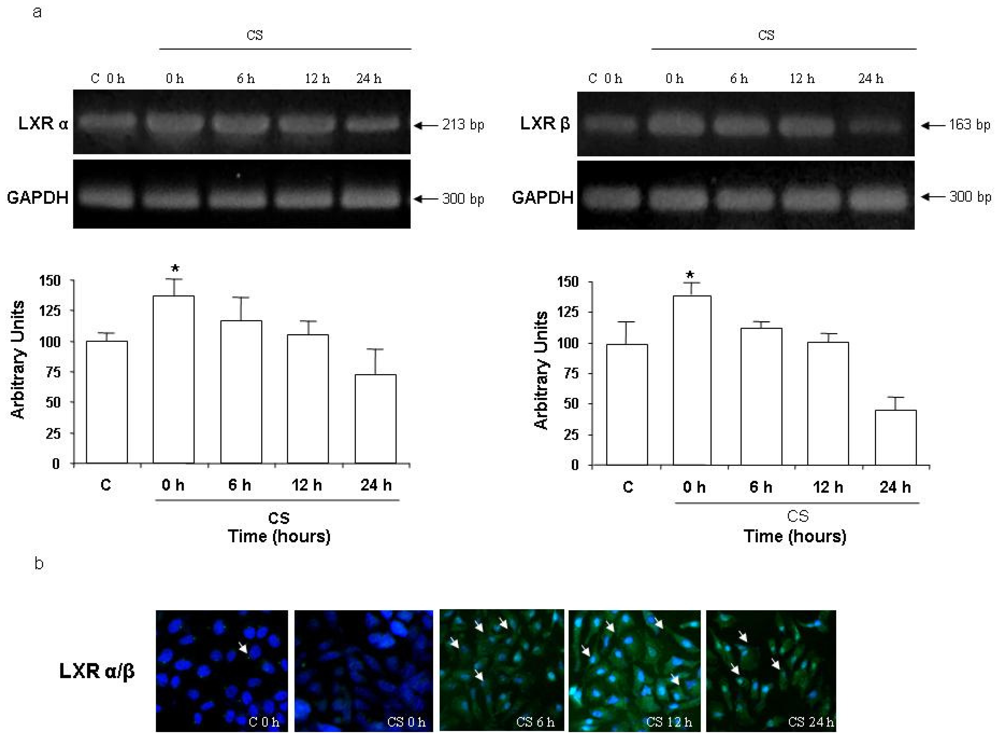
2.2 CS Induces LXR Activation
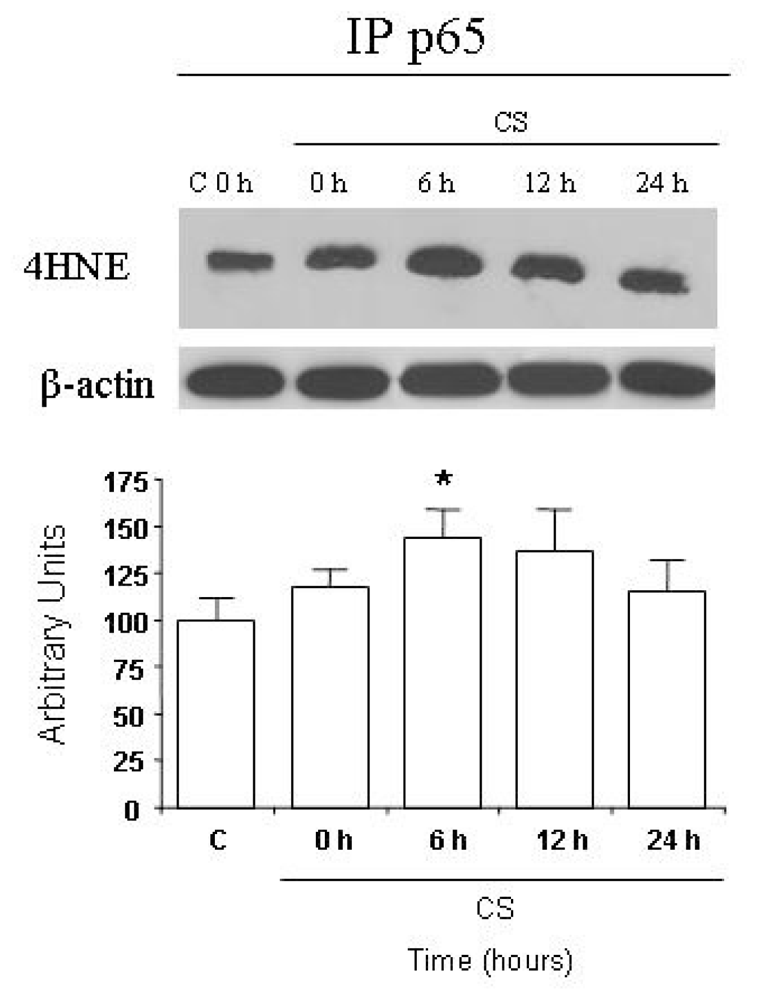
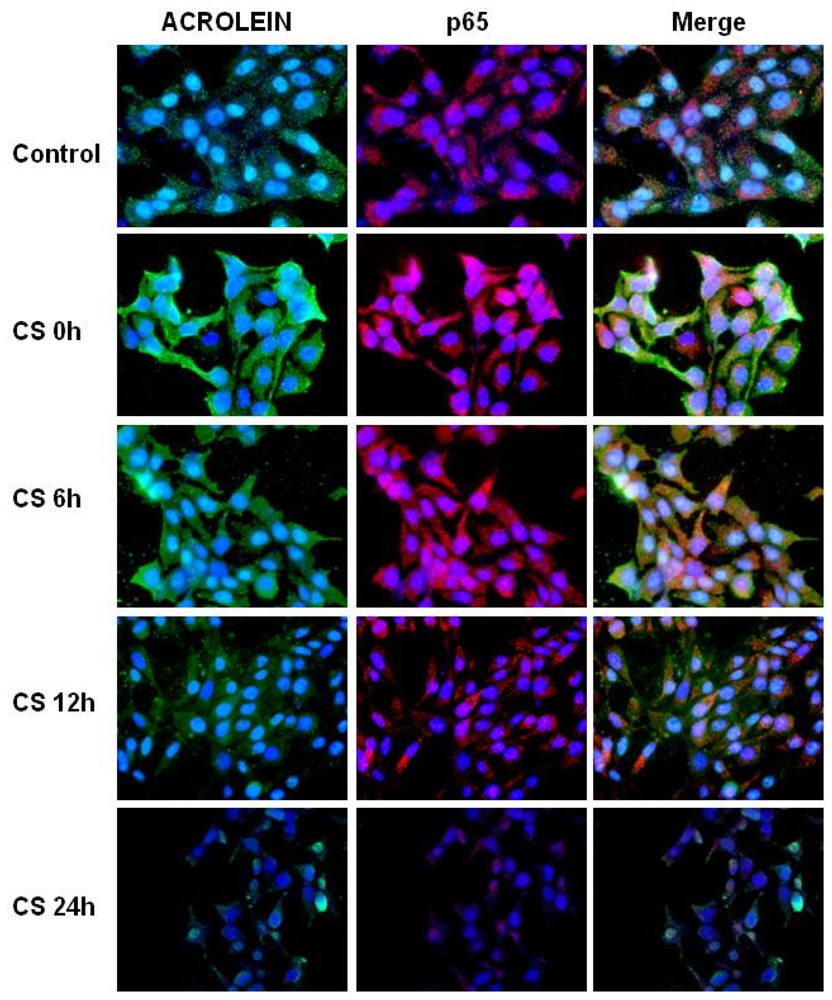
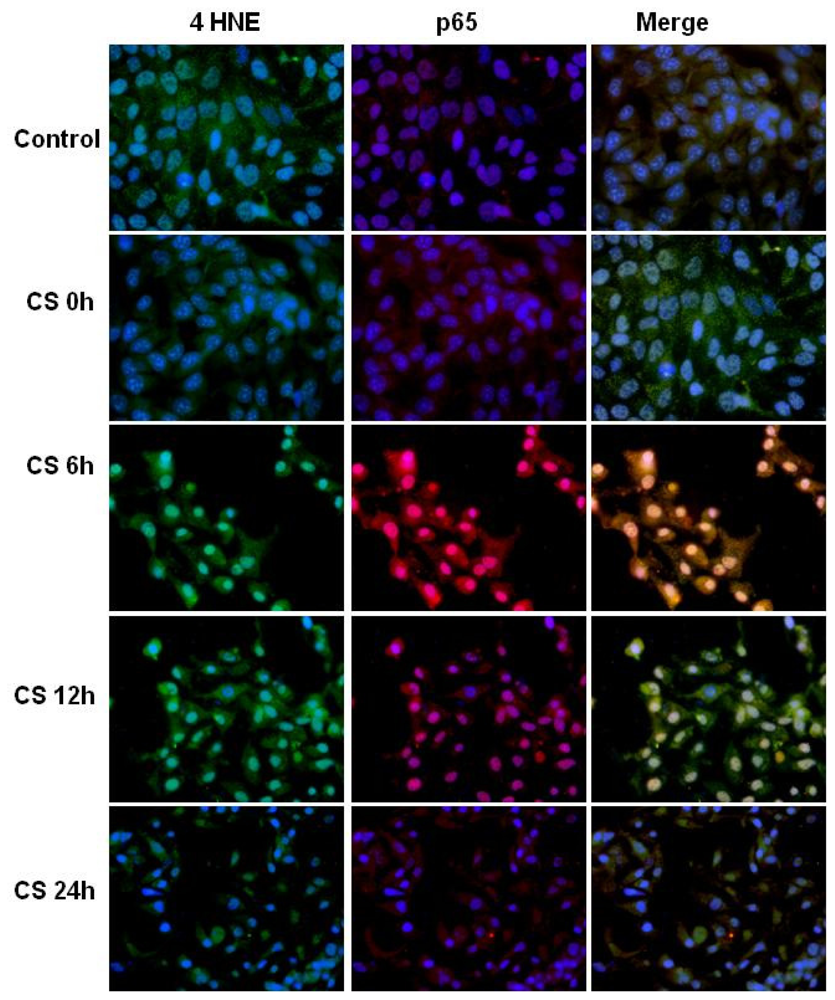
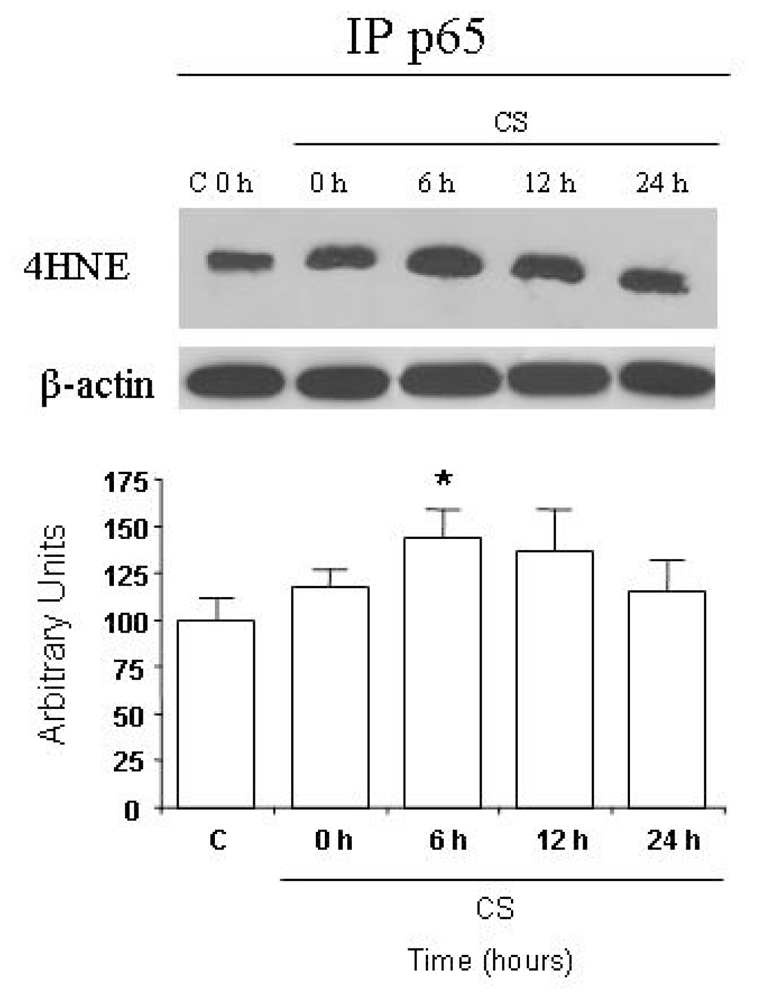
2.3. CS Induces NFκB-Aldehydes Adducts
3. Experimental Section
3.1. Cell Culture and Treatments
3.2. CS Exposure
3.3. Immunocytochemistry
3.4. Western Blot Analysis
3.5. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
3.6. Immunoprecipitation
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Schurer, NY; Elias, PM. The biochemistry and function of stratum corneum lipids. Adv. Lipid Res 1991, 24, 27–56. [Google Scholar]
- Freiman, A; Bird, G; Metelitsa, AI; Barankin, B; Lauzon, GJ. Cutaneous effects of smoking. J. Cutan. Med. Surg 2004, 8, 415–423. [Google Scholar]
- Menon, GK; Feingold, KR; Moser, AH; Brown, BE; Elias, PM. De novo sterologenesis in the skin. II. Regulation by cutaneous barrier requirements. J. Lipid Res 1985, 26, 418–427. [Google Scholar]
- Proksch, E; Brandner, JM; Jensen, JM. The skin: an indispensable barrier. Exp. Dermatol 2008, 17, 1063–1072. [Google Scholar]
- Oram, JF. ATP-binding cassette transporter A1 and cholesterol trafficking. Curr Opin. Lipidol 2002, 13, 373–381. [Google Scholar]
- Oram, JF. HDL apolipoproteins and ABCA1: Partners in the removal of excess cellular cholesterol. Arterioscler. Thromb. Vasc. Biol 2003, 23, 720–727. [Google Scholar]
- Rigamonti, E; Helin, L; Lestavel, S; Mutka, AL; Lepore, M; Fontaine, C; Bouhlel, MA; Bultel, S; Fruchart, JC; Ikonen, E; Clavey, V; Staels, B; Chinetti-Gbaguidi, G. Liver X receptor activation controls intracellular cholesterol trafficking and esterification in human macrophages. Circ. Res 2005, 97, 682–689. [Google Scholar]
- Egawa, M; Kohno, Y; Kumano, Y. Oxidative effects of cigarette smoke on the human skin. Int. J. Cosmet. Sci 1999, 21, 83–98. [Google Scholar]
- Fortino, V; Maioli, E; Torricelli, C; Davis, P; Valacchi, G. Cutaneous MMPs are differently modulated by environmental stressors in old and young mice. Toxicol. Lett 2007, 173, 73–79. [Google Scholar]
- Li, L; Holian, A. Acrolein: A respiratory toxin that suppresses pulmonary host defense. Rev. Environ. Health 1998, 13, 99–108. [Google Scholar]
- Finkelstein, EI; Ruben, J; Koot, CW; Hristova, M; van der Vliet, A. Regulation of constitutive neutrophil apoptosis by the alpha,beta-unsaturated aldehydes acrolein and 4-hydroxynonenal. Am. J. Physiol. Lung Cell Mol. Physiol 2005, 289, L1019–L1028. [Google Scholar]
- Petersen, DR; Doorn, JA. Reactions of 4-hydroxynonenal with proteins and cellular targets. Free Radic. Biol. Med 2004, 37, 937–945. [Google Scholar]
- Valacchi, G; Davis, PA; Khan, EM; Lanir, R; Maioli, E; Pecorelli, A; Cross, CE; Goldkorn, T. Cigarette smoke exposure causes changes in Scavenger Receptor B1 level and distribution in lung cells. Int. J. Biochem. Cell Biol 2009. [Epub ahead of print]. [Google Scholar]
- Valacchi, G; Rimbach, G; Saliou, C; Weber, SU; Packer, L. Effect of benzoyl peroxide on antioxidant status, NF-kappaB activity and interleukin-1alpha gene expression in human keratinocytes. Toxicology 2001, 165, 225–234. [Google Scholar]
- Khan, EM; Lanir, R; Danielson, AR; Goldkorn, T. Epidermal growth factor receptor exposed to cigarette smoke is aberrantly activated and undergoes perinuclear trafficking. FASEB J 2008, 22, 910–917. [Google Scholar]
- Valacchi, G; Pagnin, E; Corbacho, AM; Olano, E; Davis, PA; Packer, L; Cross, CE. In vivo ozone exposure induces antioxidant/stress-related responses in murine lung and skin. Free Radic. Biol. Med 2004, 36, 673–681. [Google Scholar]
- Silverstein, P. Smoking and wound healing. Am. J. Med 1992, 93, 22S–24S. [Google Scholar]
- Schick, S; Glantz, S. Philip Morris toxicological experiments with fresh sidestream smoke: More toxic than mainstream smoke. Tob. Control 2005, 14, 396–404. [Google Scholar]
- Cai, J; Bhatnagar, A; Pierce, WM, Jr. Protein modification by acrolein: Formation and stability of cysteine adducts. Chem. Res. Toxicol 2009, 22, 708–716. [Google Scholar]
- LoPachin, RM; Gavin, T; Petersen, DR; Barber, DS. Molecular mechanisms of 4-hydroxy-2- nonenal and acrolein toxicity: Nucleophilic targets and adduct formation. Chem. Res Toxicol 2009, 22, 1499–1508. [Google Scholar]
- Freiman, A; Bird, G; Metelitsa, AI; Barankin, B; Lauzon, GJ. Cutaneous effects of smoking. J. Cutan. Med. Surg 2004, 8, 415–423. [Google Scholar]
- Jung, CG; Horike, H; Cha, BY; Uhm, KO; Yamauchi, R; Yamaguchi, T; Hosono, T; Iida, K; Woo, JT; Michikawa, M. Honokiol increases ABCA1 expression level by activating retinoid X receptor beta. Biol. Pharm. Bull 2010, 33, 1105–1111. [Google Scholar]
- Chen, M; Li, W; Wang, N; Zhu, Y; Wang, X. ROS and NF-kappaB but not LXR mediate IL-1beta signaling for the downregulation of ATP-binding cassette transporter A1. Am. J. Physiol. Cell Physiol 2007, 292, C1493–C1501. [Google Scholar]
- Joseph, SB; Castrillo, A; Laffitte, BA; Mangelsdorf, DJ; Tontonoz, P. Reciprocal regulation of inflammation and lipid metabolism by liver X receptors. Nat. Med 2003, 9, 213–219. [Google Scholar]
- Camandola, S; Poli, G; Mattson, MP. The lipid peroxidation product 4-hydroxy-2,3-nonenal inhibits constitutive and inducible activity of nuclear factor kappa B in neurons. Brain Res. Mol. Brain Res 2000, 85, 53–60. [Google Scholar]
- Valacchi, G; Pagnin, E; Phung, A; Nardini, M; Schock, BC; Cross, CE; van der Vliet, A. Inhibition of NFκB activation and IL-8 expression in human bronchial epithelial cells by acrolein. Antioxid. Redox. Signal 2005, 7, 25–31. [Google Scholar]
- Page, S; Fischer, C; Baumgartner, B; Haas, M; Kreusel, U; Loidl, G; Hayn, M; Ziegler-Heitbrock, HW; Neumeier, D; Brand, K. 4-Hydroxynonenal prevents NF-κB activation and tumor necrosis factor expression by inhibiting IkappaB phosphorylation and sub- sequent proteolysis. J. Biol. Chem 1999, 274, 11611–11618. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target sequence | Primer | Sequence (5′→3′) | Amplicon length |
|---|---|---|---|
| ABCA-1 | Sense | AACAGTTTGTGGCCCTTTTG | 157 bp |
| Antisense | AGTTCCAGGCTGGGGTACTT | ||
| LXRα | Sense | CGGGCTTCCACTACAATGTT | 213 bp |
| Antisense | TCAGGCGGATCTGTTCTTCT | ||
| LXRβ | Sense | CCTCCTGAAGGCATCCACTA | 163 bp |
| Antisense | GAACTCGAAGATGGGGTTGA | ||
| GAPDH | Sense | ATGGGGAAGGTGAAGGTCGG | 300 bp |
| Antisense | TGGTGAAGACGCCAGTGGAC |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sticozzi, C.; Pecorelli, A.; Belmonte, G.; Valacchi, G. Cigarette Smoke Affects ABCAl Expression via Liver X Receptor Nuclear Translocation in Human Keratinocytes. Int. J. Mol. Sci. 2010, 11, 3375-3386. https://doi.org/10.3390/ijms11093375
Sticozzi C, Pecorelli A, Belmonte G, Valacchi G. Cigarette Smoke Affects ABCAl Expression via Liver X Receptor Nuclear Translocation in Human Keratinocytes. International Journal of Molecular Sciences. 2010; 11(9):3375-3386. https://doi.org/10.3390/ijms11093375
Chicago/Turabian StyleSticozzi, Claudia, Alessandra Pecorelli, Giuseppe Belmonte, and Giuseppe Valacchi. 2010. "Cigarette Smoke Affects ABCAl Expression via Liver X Receptor Nuclear Translocation in Human Keratinocytes" International Journal of Molecular Sciences 11, no. 9: 3375-3386. https://doi.org/10.3390/ijms11093375