Use of Parsimony Analysis to Identify Areas of Endemism of Chinese Birds: Implications for Conservation and Biogeography

Abstract
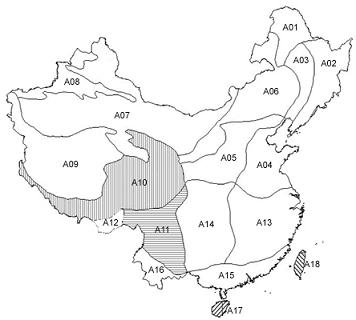
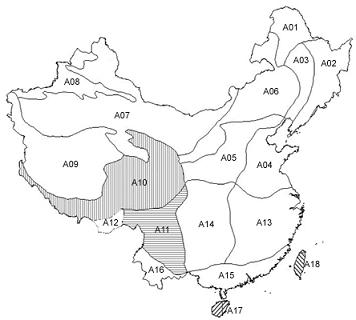
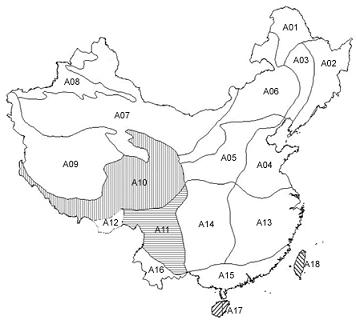
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. Results
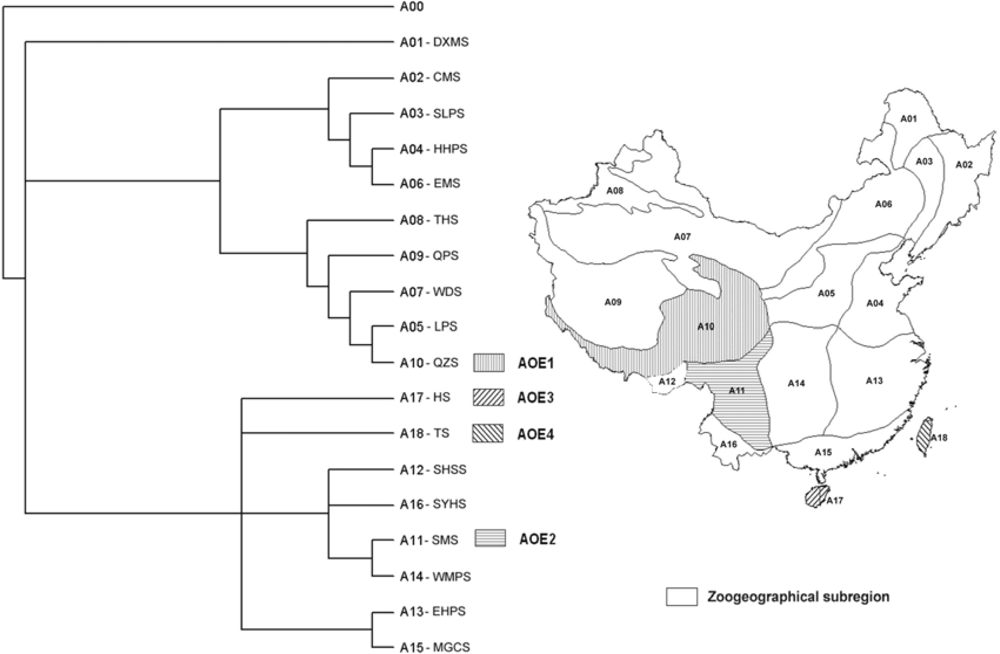
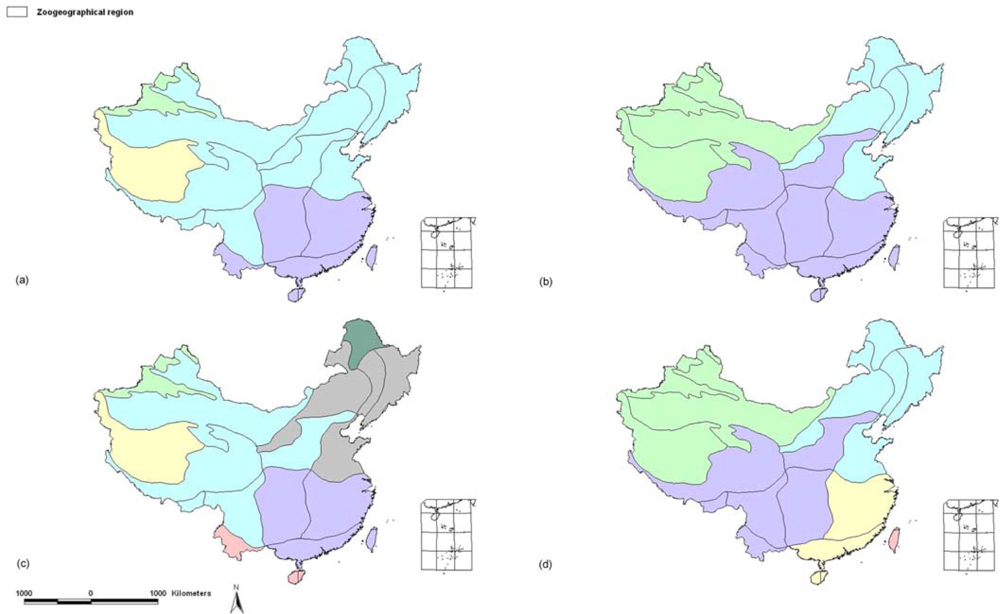
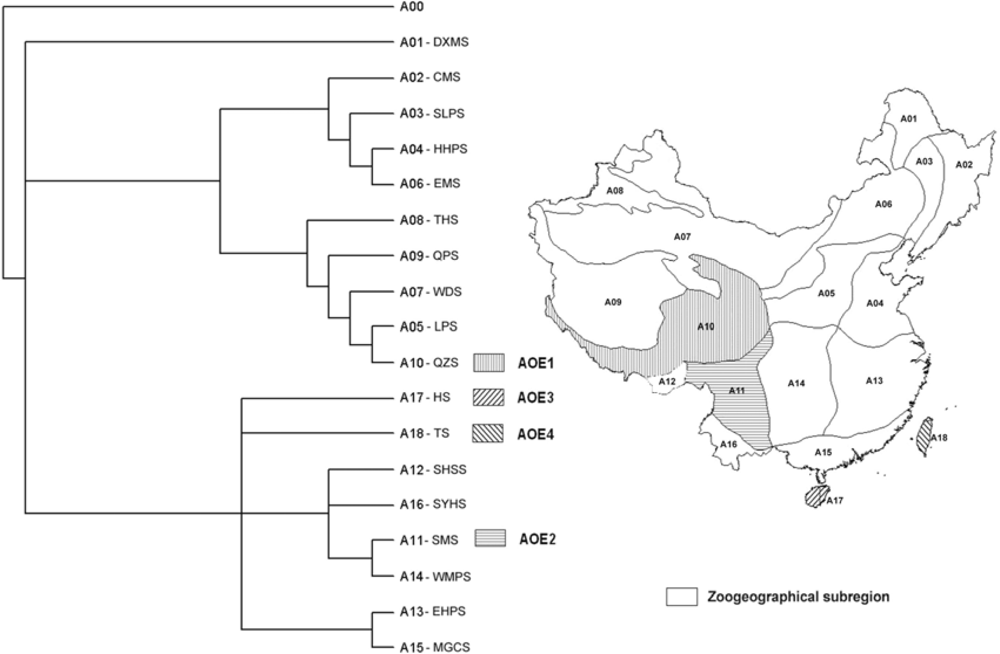
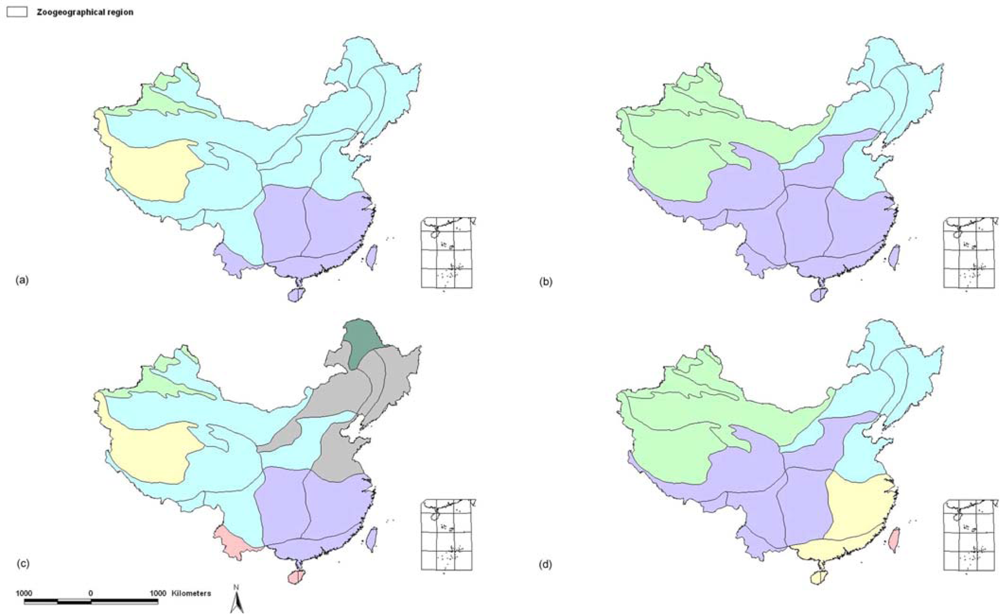
2.1.1. Subregional Avian AOEs
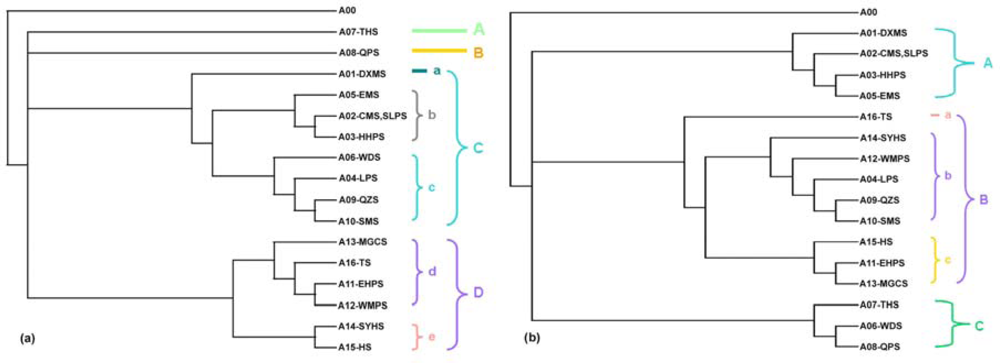
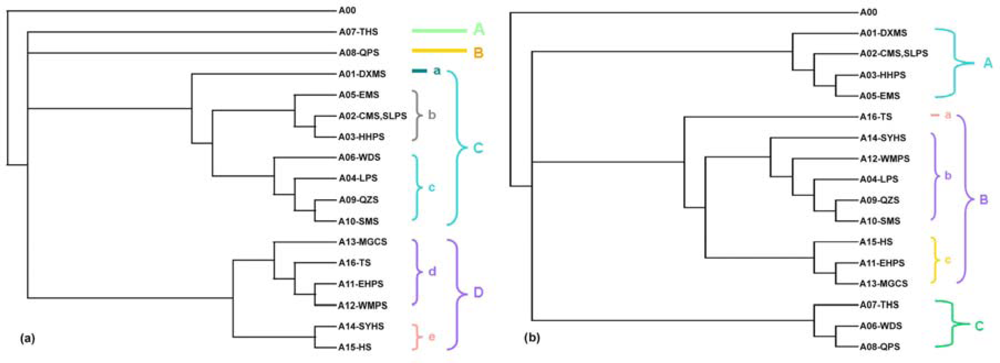
2.1.2. The 1976 and the 2007 Subregional Topologies
2.1.3. Topological Differences between the Two Trees
2.2. Discussion
2.2.1. Areas of Endemism at Subregional Scale
2.2.2. The Importance of AOEs for Avian Biodiversity Conservation
2.2.3. Can We Infer Species Distribution Changes by PAE?
3. Experimental Section
3.1. Species and Distributions
3.2. Parsimony Analysis of Endemicity
4. Conclusions
Acknowledgments
References and Notes
- Lei, FM; Qu, YH; Lu, JL; Yin, ZH. Conservation on diversity and distribution patterns of endemic birds in China. Biodivers. Conserv 2003, 12, 239–254. [Google Scholar]
- Lei, FM; Qu, YH; Tang, QQ; An, SC. Priorities for the conservation of avian biodiversity in China based on the distribution patterns of endemic bird genera. Biodivers. Conserv 2003, 12, 2487–2501. [Google Scholar]
- Cheng, TH; Zheng, GM; Zhang, FY. Fauna Sinica. Aves; Science Press: Beijing, China, 1997; Volume 1. [Google Scholar]
- Zhang, RZ. Zoogeography of China; Science Press: Beijing, China, 1999. [Google Scholar]
- Lei, FM; Wei, GA; Zhao, HF; Yin, ZH. Subregional endemism of China endemic avian species. Biodivers. Conserv 2007, 16, 1119–1130. [Google Scholar]
- Stattersfield, AJ; Crowby, MJ; Long, AJ; Wege, DC. Endemic Bird Areas of the World: Priorities for Biodiversity Conservation; Birdlife International: Cambridge, UK, 1998. [Google Scholar]
- Myers, N; Mittermeier, RA; Mittermeier, CG; Fonseca, GAB; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar]
- Loyola, RD; Kubota, U; Lewinsohn, TM. Endemic vertebrates are the most effective surrogates for identifying conservation priorities among Brazilian ecoregions. Divers. Distrib 2007, 13, 389–306. [Google Scholar]
- Lamoreux, JF; Morrison, JC; Ricketts, TH; Olson, DM; Dinerstein, E; McKnoght, MW; Shugart, HH. Global tests of biodiversity concordance and the importance of endemism. Nature 2006, 440, 212–214. [Google Scholar]
- Rosen, BR. From fossils to earth history: applied historical biogeography. In Analytical Biogeography: An Integrated Approach to the Study of Animal and Plant Distributions; Myers, AA, Giller, P, Eds.; Chapman & Hall: London, UK, 1988; pp. 437–481. [Google Scholar]
- Morrone, JJ; Crisci, JV. Historical biogeography: Introduction to methods. Annu. Rev. Ecol. Syst 1995, 26, 373–401. [Google Scholar]
- Morrone, JJ; Escalante, T. Parsimony analysis of endemicity of Mexican terrestrial mammals at different area units: when size matters. J. Biogeogr 2002, 29, 1095–1104. [Google Scholar]
- Morrone, JJ. On the identification of areas of endemism. Syst. Biol 1994, 43, 438–441. [Google Scholar]
- Hubert, N; Renno, JF. Historical biogeography of South American freshwater fishes. J. Biogeogr 2006, 33, 1414–1436. [Google Scholar]
- Huang, XL; Qiao, GX; Lei, FM. Areas of endemism and patterns of diversity for aphids of the Qinghai-Tibetan Plateau and the Himalayas. J. Biogeogr 2008, 35, 230–240. [Google Scholar]
- Linder, HP. On areas of endemism, with an example from the African Restionaceae. Syst. Biol 2001, 50, 892–912. [Google Scholar]
- Szumik, CA; Cuezzo, F; Goloboff, PA; Chalup, AE. An optimality criterion to determine areas of endemism. Syst. Biol 2002, 51, 806–816. [Google Scholar]
- Szumik, CA; Goloboff, PA. Areas of endemism: an improved optimality criterion. Syst. Biol 2004, 53, 968–977. [Google Scholar]
- Mast, AR; Nyffeler, R. Using a null model to recognize significant co-occurrence prior to identifying candidate areas of endemism. Syst. Biol 2003, 52, 271–280. [Google Scholar]
- Hausdorf, B; Hennig, C. Biotic element analysis in biogeography. Syst. Biol 2003, 52, 717–723. [Google Scholar]
- Moline, PM; Linder, HP. Input data, analytical methods and biogeography of Elegia (Restionaceae). J. Biogeogr 2006, 33, 47–62. [Google Scholar]
- Kattan, GH; Franco, P; Rojas, V; Morales, G. Biological diversification in a complex region: a spatial analysis of faunistic diversity and biogeography of the Andes of Colombia. J. Biogeogr 2004, 31, 1829–1839. [Google Scholar]
- Brommer, JE. The range margins of northern birds shift polewards. Ann. Zool. Fenn 2004, 41, 391–397. [Google Scholar]
- Thomas, CD; Lennon, JJ. Birds extend their ranges northwards. Nature 1999, 399, 213. [Google Scholar]
- Johnson, NK. Pioneering and natural expansion of breeding distributions in western North American birds. Stud. Avian Biol 1994, 15, 27–46. [Google Scholar]
- Valiela, I; Bowen, JL. Shifts in winter distribution in birds: effects of global warming and local habitat change. Ambio 2003, 32, 76–480. [Google Scholar]
- Wehtje, W. The range expansion of the great-tailed grackle (Quiscalus mexicanus Gmelin) in North America since 1880. J. Biogeogr 2003, 30, 1593–1607. [Google Scholar]
- Maclean, IMD; Austin, GE; Rehfisch, MM; Blew, J; Crowe, O; Delany, S; Devos, K; Deceuninck, B; Gunther, K; Laursen, K; van Roomen, M; Wahl, J. Climate change causes rapid changes in the distribution and site abundance of birds in winter. Global Change Biol 2008, 14, 2489–2500. [Google Scholar]
- Zuckerberg, B; Woods, AM; Porter, WF. Poleward shifts in breeding bird distributions in New York State. Global Change Biol 2009, 15, 1866–1883. [Google Scholar]
- Sun, QH; Zhang, ZW. The impact of climate warming on the distribution of Chinese birds. Chin. J. Zool 2000, 35, 45–48. [Google Scholar]
- Cheng, TH. A Checklist of the Avifauna of China; Science Press: Beijing, China, 1976. [Google Scholar]
- Shi, YF; Li, JJ; Li, BY. Uplift and Environmental Change of Qinghai-Xizang (Tibetan) Plateau in the Late Cenozoic; Guangdong Science and Technology Press: Guangzhou, China, 1998. [Google Scholar]
- Cao, WX; Chen, YY; Wu, YF; Zhu, SQ. Origin and evolution of Schizothoracine fishes in relation to the upheaval of the Qinghai-Xizang Plateau. In Studies on the Period, Amplitude and Type of the Uplift of the Qinghai-Xizang Plateau; Li, QF, Ed.; Science Press: Beijing, China, 1981; pp. 118–129. [Google Scholar]
- Zou, FS; Lim, HC; Marks, BD; Moyle, RG; Sheldon, FH. Molecular phylogenetic analysis of the Grey-cheeked Fulvetta (Alcippe morrisonia) of China and Indochina: a case of remarkable genetic divergence in a “species”. Mol. Phylogenet. Evol 2007, 44, 165–174. [Google Scholar]
- Song, G; Qu, YH; Yin, ZH; Li, SH; Liu, NF; Lei, FM. Phylogeography of the Alcippe morrisonia (Aves: Timaliidae): Long population history beyond late Pleistocene glaciations. BMC Evol Biol 2009, 9, 143:1–143:11. [Google Scholar]
- Li, SH; Li, JW; Han, LX; Yao, CT; Shi, HT; Lei, FM. Species delimitation in the Hwamei Garrulax canorus. Ibis 2006, 148, 698–706. [Google Scholar]
- Brown, JH; Limolino, MV. Biogeography; Sinauer Associates: Sunderland, UK, 1998. [Google Scholar]
- Zhao, HF; Lei, FM; Qu, YH; Lu, JL. Conservation priority based on avian subspecific differentiation of endemic species. Acta Zool. Sinica 2007, 53, 378–382. [Google Scholar]
- .
- Guo, ZM; Miao, QL; Li, X. Variation characteristics of temperature over northern China in recent 50 years. Scientia Geogr. Sinica 2005, 25, 448–454. [Google Scholar]
- Miao, QL; Ding, YY; Wang, Y. Impact of climate warming on the northern boundary of subtropical zone of China. Geogr. Res 2009, 28, 634–642. [Google Scholar]
- Chen, SY; Guo, ZX; Gao, R; Han, T. The climatic variety and anomaly of the winter mean air temperature of the east monsoon area in China. J. Appl. Meteor. Sci 2009, 20, 478–485. [Google Scholar]
- Ma, ZF; Peng, J; Gao, WL; Tian, H. Climate variation of southwest China in recent 40 years. Plateau Meteor 2006, 25, 633–642. [Google Scholar]
- Price, JT; Root, TL. Climate change and Neotropical migrants. Presented at the 66th Annual North American Wildlife and Natural Resources Conference, Washington, DC, USA, March 2001.
- Webb, T, III. Past changes in vegetation and climate: lessons for the future. In Global Warming and Biological Diversity; Peters, RL, Lovejoy, TE, Eds.; Yale University Press: New Haven, CT, USA, 1992; pp. 59–72. [Google Scholar]
- Nihei, SS. Misconceptions about parsimony analysis of endemicity. J. Biogeogr 2006, 33, 2099–2106. [Google Scholar]
- Lei, FM; Lu, TC. China Endemic Birds; Science Press: Beijing, China, 2006. [Google Scholar]
- Hausdorf, B. Units in biogeography. Syst. Biol 2002, 51, 648–652. [Google Scholar]
- Swofford, DL. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); , ver 4.0. Sinauer Associates: Sunderland, UK, 1998. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huang, X.-L.; Qiao, G.-X.; Lei, F.-M. Use of Parsimony Analysis to Identify Areas of Endemism of Chinese Birds: Implications for Conservation and Biogeography. Int. J. Mol. Sci. 2010, 11, 2097-2108. https://doi.org/10.3390/ijms11052097
Huang X-L, Qiao G-X, Lei F-M. Use of Parsimony Analysis to Identify Areas of Endemism of Chinese Birds: Implications for Conservation and Biogeography. International Journal of Molecular Sciences. 2010; 11(5):2097-2108. https://doi.org/10.3390/ijms11052097
Chicago/Turabian StyleHuang, Xiao-Lei, Ge-Xia Qiao, and Fu-Min Lei. 2010. "Use of Parsimony Analysis to Identify Areas of Endemism of Chinese Birds: Implications for Conservation and Biogeography" International Journal of Molecular Sciences 11, no. 5: 2097-2108. https://doi.org/10.3390/ijms11052097