Protein Microarrays and Biomarkers of Infectious Disease

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Proteome Microarrays and Host Antibody Responses
3. Protein-Protein Interactions (Non-Antibody)
4. Alternative Methods
5. Peptide and Protein Domain Microarrays
6. Quality Control
7. Antibody Microarrays
8. Microarray Printing
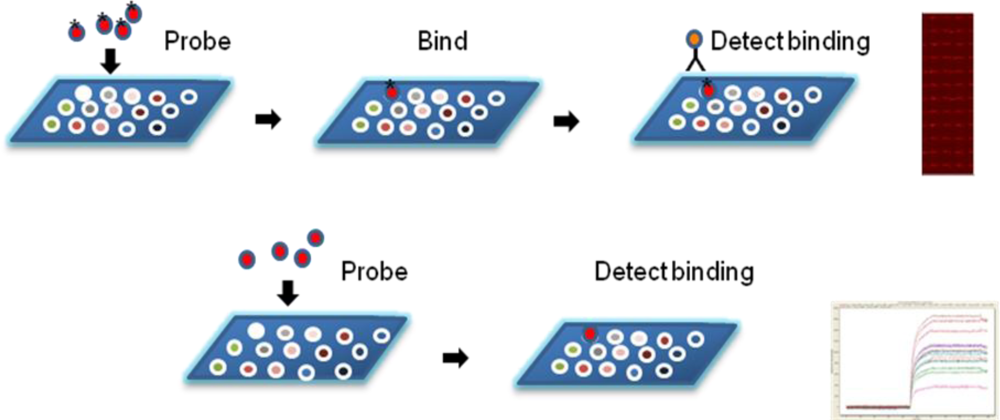
9. Labeled Detection of Binding Events
10. Label-Free Detection
11. Data Analysis
12. Conclusions
Acknowledgments
References
- Taniguchi, Y; Choi, PJ; Li, GW; Chen, H; Babu, M; Hearn, J; Emili, A; Xie, XS. Quantifying E. coli proteome and transcriptome with single-molecule sensitivity in single cells. Science 2010, 329, 533–538. [Google Scholar]
- Gygi, SP; Rochon, Y; Franza, BR; Aebersold, R. Correlation between protein and mRNA abundance in yeast. Mol. Cell Biol 1999, 19, 1720–1730. [Google Scholar]
- Yu, X; Xu, D; Cheng, Q. Label-free detection methods for protein microarrays. Proteomics 2006, 6, 5493–5503. [Google Scholar]
- Homola, J. Present and future of surface plasmon resonance biosensors. Anal. Bioanal. Chem 2003, 377, 528–539. [Google Scholar]
- Zheng, G; Patolsky, F; Cui, Y; Wang, WU; Lieber, CM. Multiplexed electrical detection of cancer markers with nanowire sensor arrays. Nat. Biotechnol 2005, 23, 1294–1301. [Google Scholar]
- Finnskog, D; Jaras, K; Ressine, A; Malm, J; Marko-Varga, G; Lilja, H; Laurell, T. High-speed biomarker identification utilizing porous silicon nanovial arrays and MALDI-TOF mass spectrometry. Electrophoresis 2006, 27, 1093–1103. [Google Scholar]
- Usui-Aoki, K; Kyo, M; Kawai, M; Murakami, M; Imai, K; Shimada, K; Koga, H. Protein and antibody microarrays: Clues towards biomarker discovery. Front. Drug Des. Disc 2006, 2, 23–33. [Google Scholar]
- Tomizaki, KY; Usui, K; Mihara, H. Protein-protein interactions and selection: Array-based techniques for screening disease-associated biomarkers in predictive/early diagnosis. FEBS J 2010, 277, 1996–2005. [Google Scholar]
- Vigil, A; Davies, DH; Felgner, PL. Defining the humoral immune response to infectious agents using high-density protein microarrays. Future Microbiol 2010, 5, 241–251. [Google Scholar]
- Borrebaeck, CA; Wingren, C. Design of high-density antibody microarrays for disease proteomics: Key technological issues. J. Proteomics 2009, 72, 928–935. [Google Scholar]
- Keasey, SL; Schmid, KE; Lee, MS; Meegan, J; Tomas, P; Minto, M; Tikhonov, AP; Schweitzer, B; Ulrich, RG. Extensive antibody cross-reactivity among infectious gram-negative bacteria revealed by proteome microarray analysis. Mol. Cell Proteomics 2009, 8, 924–935. [Google Scholar]
- Schmid, K; Keasey, SL; Pittman, P; Emerson, GL; Meegan, J; Tikonov, AP; Chen, G; Schweitzer, B; Ulrich, RG. Analysis of the human immune response to vacccina by use of novel protein microarray suggests that antibodies recognize less than 10% of the total viral proteome. Proteomics Clin. Appl 2008, 2, 1528–1538. [Google Scholar]
- Ramachandran, N; Srivastava, S; LaBaer, JL. Applications of protein microarrays for biomarker discovery. Proteomics-Clin. Appl 2008, 2, 1444–1459. [Google Scholar]
- Stoevesandt, O; Taussig, MJ; He, M. Protein microarrays: High-throughput tools for proteomics. Expert Rev. Proteomics 2009, 6, 145–157. [Google Scholar]
- Rowe, W; Platt, M; Day, PJ. Advances and perspectives in aptamer arrays. Integr. Biol. (Camb.) 2009, 1, 53–58. [Google Scholar]
- Renberg, B; Nordin, J; Merca, A; Uhlen, M; Feldwisch, J; Nygren, PA; Karlstrom, AE. Affibody molecules in protein capture microarrays: Evaluation of multidomain ligands and different detection formats. J. Proteome Res 2007, 6, 171–179. [Google Scholar]
- Ramirez, AB; Loch, CM; Zhang, Y; Liu, Y; Wang, X; Wayner, EA; Sargent, JE; Sibani, S; Hainsworth, E; Mendoza, EA; Eugene, R; Labaer, J; Urban, ND; McIntosh, MW; Lampe, PD. Use of a single-chain antibody library for ovarian cancer biomarker discovery. Mol. Cell Proteomics 2010, 9, 1449–1460. [Google Scholar]
- vanMeter, A; Signore, M; Pierobon, M; Espina, V; Liotta, LA; Petricoin, EF, III. Reverse-phase protein microarrays: Application to biomarker discovery and translational medicine. Expert. Rev. Mol. Diagn 2007, 7, 625–633. [Google Scholar]
- Speer, R; Wulfkuhle, J; Espina, V; Aurajo, R; Edmiston, KH; Liotta, LA; Petricoin, EF, III. Development of reverse phase protein microarrays for clinical applications and patient-tailored therapy. Cancer Genomics Proteomics 2007, 4, 157–164. [Google Scholar]
- Paweletz, CP; Charboneau, L; Bischsel, VE; Simone, NL; Chen, T; Gillespie, JW; Emmert-Buck, MR; Roth, MJ; Petricoin, EF, III; Liotta, LA. Reverse phase protein microarrays which capture disease progression show activation of pro-survival pathways at the cancer invasion front. Oncogene 2001, 20, 1981–1989. [Google Scholar]
- Mendes, KN; Nicorici, D; Cogdell, D; Tabus, I; Yli-Harja, O; Guerra, R; Hamilton, SR; Zhang, W. Analysis of signaling pathways in 90 cancer cell lines by protein lysate array. J. Proteome Res 2007, 6, 2753–2767. [Google Scholar]
- Spurrier, B; Ramalingam, S; Nishizuka, S. Reverse-phase protein lysate microarrays for cell signaling analysis. Nat. Protoc 2008, 3, 1796–1808. [Google Scholar]
- Kersten, B; Wanker, EE; Hoheisel, JD; Angenendt, P. Multiplex approaches in protein microarray technology. Expert. Rev. Proteomics 2005, 2, 499–510. [Google Scholar]
- MacBeath, G; Schreiber, SL. Printing proteins as microarrays for high-throughput function determination. Science 2000, 289, 1760–1763. [Google Scholar]
- Ramachandran, N; Hainsworth, E; Bhullar, B; Eisenstein, S; Rosen, B; Lau, AY; Walter, JC; LaBaer, J. Self-assembling protein microarrays. Science 2004, 305, 86–90. [Google Scholar]
- Zhu, H; Bilgin, M; Bangham, R; Hall, D; Casamayor, A; Bertone, P; Lan, N; Jansen, R; Bidlingmaier, S; Houfek, T; Mitchell, T; Miller, P; Dean, RA; Gerstein, M; Snyder, M. Global analysis of protein activities using proteome chips. Science 2001, 293, 2101–2105. [Google Scholar]
- Davies, DH; Molina, DM; Wrammert, J; Miller, J; Hirst, S; Mu, Y; Pablo, J; Unal, B; Nakajima-Sasaki, R; Liang, X; Crotty, S; Karem, KL; Damon, IK; Ahmed, R; Villarreal, L; Felgner, PL. Proteome-wide analysis of the serological response to vaccinia and smallpox. Proteomics 2007, 7, 1678–1586. [Google Scholar]
- Davies, DH; Wyatt, LS; Newman, FK; Earl, PL; Chun, S; Hernandez, JE; Molina, DM; Hirst, S; Moss, B; Frey, SE; Felgner, PL. Antibody profiling by proteome microarray reveals the immunogenicity of the attenuated smallpox vaccine modified vaccinia virus ankara is comparable to that of Dryvax. J. Virol 2008, 82, 652–663. [Google Scholar]
- Felgner, PL; Kayala, MA; Vigil, A; Burk, C; Nakajima-Sasaki, R; Pablo, J; Molina, DM; Hirst, S; Chew, JS; Wang, D; Tan, G; Duffield, M; Yang, R; Neel, J; Chantratita, N; Bancroft, G; Lertmemongkolchai, G; Davies, DH; Baldi, P; Peacock, S; Titball, RW. A Burkholderia pseudomallei protein microarray reveals serodiagnostic and cross-reactive antigens. Proc. Natl. Acad. Sci. USA 2009, 106, 13499–13504. [Google Scholar]
- Li, B; Jiang, L; Song, Q; Yang, J; Chen, Z; Guo, Z; Zhou, D; Du, Z; Song, Y; Wang, J; Wang, H; Yu, S; Yang, R. Protein microarray for profiling antibody responses to Yersinia pestis live vaccine. Infect. Immun 2005, 73, 3734–3739. [Google Scholar]
- Li, B; Guo, Y; Guo, Z; Liang, Y; Zhu, Z; Zhou, Q; Yan, Y; Song, Z; Yang, R. Serologic survey of the sentinel animals for plague surveillance and screening for complementary diagnostic markers to F1 antigen by protein microarray. Am. J. Trop. Med. Hyg 2008, 79, 799–802. [Google Scholar]
- Li, B; Zhou, D; Wang, Z; Song, Z; Wang, H; Li, M; Dong, X; Wu, M; Guo, Z; Yang, R. Antibody profiling in plague patients by protein microarray. Microbes Infect 2008, 10, 45–51. [Google Scholar]
- Sundaresh, S; Randall, A; Unal, B; Petersen, JM; Belisle, JT; Hartley, MG; Duffield, M; Titball, RW; Davies, DH; Felgner, PL; Baldi, P. From protein microarrays to diagnostic antigen discovery: A study of the pathogen Francisella tularensis. Bioinformatics 2007, 23, 1508–1518. [Google Scholar]
- Eyles, JE; Unal, B; Hartley, MG; Newstead, SL; Flick-Smith, H; Prior, JL; Oyston, PC; Randall, A; Mu, Y; Hirst, S; Molina, DM; Davies, DH; Milne, T; Griffin, KF; Baldi, P; Titball, RW; Felgner, PL. Immunodominant Francisella tularensis antigens identified using proteome microarray. Proteomics 2007, 7, 2172–2183. [Google Scholar]
- Beare, PA; Chen, C; Bouman, T; Pablo, J; Unal, B; Cockrell, DC; Brown, WC; Barbian, KD; Porcella, SF; Samuel, JE; Felgner, PL; Heinzen, RA. Candidate antigens for Q fever serodiagnosis revealed by immunoscreening of a Coxiella burnetii protein microarray. Clin. Vaccine Immunol 2008, 15, 1771–1779. [Google Scholar]
- Liang, L; Leng, D; Burk, C; Nakajima-Sasaki, R; Kayala, MA; Atluri, VL; Pablo, J; Unal, B; Ficht, TA; Gotuzzo, E; Saito, M; Morrow, WJ; Liang, X; Baldi, P; Gilman, RH; Vinetz, JM; Tsolis, RM; Felgner, PL. Large scale immune profiling of infected humans and goats reveals differential recognition of Brucella melitensis antigens. PLoS Negl. Trop. Dis 2010, 4, e673. [Google Scholar]
- Barbour, AG; Jasinskas, A; Kayala, MA; Davies, DH; Steere, AC; Baldi, P; Felgner, PL. A genome-wide proteome array reveals a limited set of immunogens in natural infections of humans and white-footed mice with Borrelia burgdorferi. Infect. Immun 2008, 76, 3374–3389. [Google Scholar]
- Montor, WR; Huang, J; Hu, Y; Hainsworth, E; Lynch, S; Kronish, JW; Ordonez, CL; Logvinenko, T; Lory, S; LaBaer, J. Genome-wide study of Pseudomonas aeruginosa outer membrane protein immunogenicity using self-assembling protein microarrays. Infect. Immun 2009, 77, 4877–4886. [Google Scholar]
- Steller, S; Angenendt, P; Cahill, DJ; Heuberger, S; Lehrach, H; Kreutzberger, J. Bacterial protein microarrays for identification of new potential diagnostic markers for Neisseria meningitidis infections. Proteomics 2005, 5, 2048–2055. [Google Scholar]
- Li, Q; Jagannath, C; Rao, PK; Singh, CR; Lostumbo, G. Analysis of phagosomal proteomes: From latex-bead to bacterial phagosomes. Proteomics 2010, 10, 4098–4116. [Google Scholar]
- Popova, T; Espina, V; Bailey, C; Liotta, L; Petricoin, E; Popov, S. Anthrax infection inhibits the AKT signaling involved in the E-cadherin-mediated adhesion of lung epithelial cells. FEMS Immunol. Med. Microbiol 2009, 56, 129–142. [Google Scholar]
- Margarit, I; Bonacci, S; Pietrocola, G; Rindi, S; Ghezzo, C; Bombaci, M; Nardi-Dei, V; Grifantini, R; Speziale, P; Grandi, G. Capturing host-pathogen interactions by protein microarrays: Identification of novel streptococcal proteins binding to human fibronectin, fibrinogen, and C4BP. FASEB J 2009, 23, 3100–3112. [Google Scholar]
- Davies, DH; Liang, X; Hernandez, JE; Randall, A; Hirst, S; Mu, Y; Romero, KM; Nguyen, TT; Kalantari-Dehaghi, M; Crotty, S; Baldi, P; Villarreal, LP; Felgner, PL. Profiling the humoral immune response to infection by using proteome microarrays: high-throughput vaccine and diagnostic antigen discovery. Proc. Natl. Acad. Sci. USA 2005, 102, 547–552. [Google Scholar]
- Sundaresh, S; Doolan, DL; Hirst, S; Mu, Y; Unal, B; Davies, DH; Felgner, PL; Baldi, P. Identification of humoral immune responses in protein microarrays using DNA microarray data analysis techniques. Bioinformatics 2006, 22, 1760–1766. [Google Scholar]
- He, M; Taussig, MJ. Single step generation of protein arrays from DNA by cell-free expression and in situ immobilisation (PISA method). Nucleic Acids Res 2001, 29, E73–3. [Google Scholar]
- Ramachandran, N; Raphael, JV; Hainsworth, E; Demirkan, G; Fuentes, MG; Rolfs, A; Hu, Y; LaBaer, J. Next-generation high-density self-assembling functional protein arrays. Nat. Methods 2008, 5, 535–538. [Google Scholar]
- He, M; Stoevesandt, O; Palmer, EA; Khan, F; Ericsson, O; Taussig, MJ. Printing protein arrays from DNA arrays. Nat. Methods 2008, 5, 175–177. [Google Scholar]
- Ceroni, A; Sibani, S; Baiker, A; Pothineni, VR; Bailer, SM; Labaer, J; Haas, J; Campbell, CJ. Systematic analysis of the IgG antibody immune response against varicella zoster virus (VZV) using a self-assembled protein microarray. Mol. Biosyst 2010, 6, 1604–1610. [Google Scholar]
- Vizoso Pinto, MG; Pfrepper, KI; Janke, T; Noelting, C; Sander, M; Lueking, A; Haas, J; Nitschko, H; Jaeger, G; Baiker, A. A systematic approach for the identification of novel, serologically reactive recombinant Varicella-Zoster Virus (VZV) antigens. Virol. J 2010, 7, 165. [Google Scholar]
- Luevano, M; Bernard, HU; Barrera-Saldana, HA; Trevino, V; Garcia-Carranca, A; Villa, LL; Monk, BJ; Tan, X; Davies, DH; Felgner, PL; Kalantari, M. High-throughput profiling of the humoral immune responses against thirteen human papillomavirus types by proteome microarrays. Virology 2010, 2010. [Google Scholar]
- Waterboer, T; Sehr, P; Michael, KM; Franceschi, S; Nieland, JD; Joos, TO; Templin, MF; Pawlita, M. Multiplex human papillomavirus serology based on in situ-purified glutathione s-transferase fusion proteins. Clin. Chem 2005, 51, 1845–1853. [Google Scholar]
- Zhu, H; Hu, S; Jona, G; Zhu, X; Kreiswirth, N; Willey, BM; Mazzulli, T; Liu, G; Song, Q; Chen, P; Cameron, M; Tyler, A; Wang, J; Wen, J; Chen, W; Compton, S; Snyder, M. Severe acute respiratory syndrome diagnostics using a coronavirus protein microarray. Proc. Natl. Acad. Sci. USA 2006, 103, 4011–4016. [Google Scholar]
- Duburcq, X; Olivier, C; Malingue, F; Desmet, R; Bouzidi, A; Zhou, F; Auriault, C; Gras-Masse, H; Melnyk, O. Peptide-protein microarrays for the simultaneous detection of pathogen infections. Bioconjug. Chem 2004, 15, 307–316. [Google Scholar]
- List, C; Qi, W; Maag, E; Gottstein, B; Muller, N; Felger, I. Serodiagnosis of Echinococcus spp. infection: Explorative selection of diagnostic antigens by peptide microarray. PLoS Negl. Trop. Dis 2010, 4, e771. [Google Scholar]
- Espejo, A; Cote, J; Bednarek, A; Richard, S; Bedford, MT. A protein-domain microarray identifies novel protein-protein interactions. Biochem. J 2002, 367, 697–702. [Google Scholar]
- Kaushansky, A; Allen, JE; Gordus, A; Stiffler, MA; Karp, ES; Chang, BH; MacBeath, G. Quantifying protein-protein interactions in high throughput using protein domain microarrays. Nat. Protoc 2010, 5, 773–790. [Google Scholar]
- Kijanka, G; Murphy, D. Protein arrays as tools for serum autoantibody marker discovery in cancer. J. Proteomeics 2009, 72, 936–944. [Google Scholar]
- Predki, PF; Mattoon, D; Bangham, R; Schweitzer, B; Michaud, G. Protein microarrays: A new tool for profiling antibody cross-reactivity. Hum. Antibodies 2005, 14, 7–15. [Google Scholar]
- Michaud, GA; Salcius, M; Zhou, F; Bangham, R; Bonin, J; Guo, H; Snyder, M; Predki, PF; Schweitzer, BI. Analyzing antibody specificity with whole proteome microarrays. Nat. Biotechnol 2003, 21, 1509–1512. [Google Scholar]
- Haab, BB; Dunham, MJ; Brown, PO. Protein microarrays for highly parallel detection and quantitation of specific proteins and antibodies in complex solutions. Genome Biol 2001, 2. RESEARCH0004. [Google Scholar]
- Larkin, EA; Stiles, BG; Ulrich, RG. Inhibition of toxic shock by human monoclonal antibodies against staphylococcal enterotoxin B. PLoS One 2010, 5. in press.. [Google Scholar]
- Rucker, VC; Havenstrite, KL; Herr, AE. Antibody microarrays for native toxin detection. Anal. Biochem 2005, 339, 262–270. [Google Scholar]
- Gehring, AG; Albin, DM; Bhunia, AK; Reed, SA; Tu, SI; Uknalis, J. Antibody microarray detection of Escherichia coli O157:H7: Quantification, assay limitations, and capture efficiency. Anal. Chem 2006, 78, 6601–6007. [Google Scholar]
- Gehring, AG; Albin, DM; Reed, SA; Tu, SI; Brewster, JD. An antibody microarray, in multiwell plate format, for multiplex screening of foodborne pathogenic bacteria and biomolecules. Anal. Bioanal. Chem 2008, 391, 497–506. [Google Scholar]
- Poetz, O; Ostendorp, R; Brocks, B; Schwenk, JM; Stoll, D; Joos, TO; Templin, MF. Protein microarrays for antibody profiling: Specificity and affinity determination on a chip. Proteomics 2005, 5, 2402–2411. [Google Scholar]
- Angenendt, P; Glokler, J. Evaluation of antibodies and microarray coatings as a prerequisite for the generation of optimized antibody microarrays. Methods Mol. Biol 2004, 264, 123–134. [Google Scholar]
- Wingren, C; Steinhauer, C; Ingvarsson, J; Persson, E; Larsson, K; Borrebaeck, CA. Microarrays based on affinity-tagged single-chain Fv antibodies: Sensitive detection of analyte in complex proteomes. Proteomics 2005, 5, 1281–1291. [Google Scholar]
- Rubina, AY; Kolchinsky, A; Makarov, AA; Zasedatelev, AS. Why 3-D? Gel-based microarrays in proteomics. Proteomics 2008, 8, 817–831. [Google Scholar]
- Natarajan, S; Katsamba, PS; Miles, A; Eckman, J; Papalia, GA; Rich, RL; Gale, BK; Myszka, DG. Continuous-flow microfluidic printing of proteins for array-based applications including surface plasmon resonance imaging. Anal. Biochem 2008, 373, 141–146. [Google Scholar]
- Lizardi, PM; Huang, X; Zhu, Z; Bray-Ward, P; Thomas, DC; Ward, DC. Mutation detection and single-molecule counting using isothermal rolling-circle amplification. Nat. Genet 1998, 19, 225–232. [Google Scholar]
- Schweitzer, B; Roberts, S; Grimwade, B; Shao, W; Wang, M; Fu, Q; Shu, Q; Laroche, I; Zhou, Z; Tchernev, VT; Christiansen, J; Velleca, M; Kingsmore, SF. Multiplexed protein profiling on microarrays by rolling-circle amplification. Nat. Biotechnol 2002, 20, 359–365. [Google Scholar]
- Zajac, A; Song, D; Qian, W; Zhukov, T. Protein microarrays and quantum dot probes for early cancer detection. Colloids Surf. B Biointerfaces 2007, 58, 309–314. [Google Scholar]
- Schwenk, JM; Gry, M; Rimini, R; Uhlen, M; Nilsson, P. Antibody suspension bead arrays within serum proteomics. J. Proteome Res 2008, 7, 3168–3179. [Google Scholar]
- Clifford, GM; Shin, HR; Oh, JK; Waterboer, T; Ju, YH; Vaccarella, S; Quint, W; Pawlita, M; Franceschi, S. Serologic response to oncogenic human papillomavirus types in male and female university students in Busan, South Korea. Cancer Epidemiol. Biomarkers Prev 2007, 16, 1874–1879. [Google Scholar]
- Altuglu, I; Bozkurt, H; Samlioglu, P; Zeytinoglu, A. Evaluation of three different assays for the assessment of Epstein Barr-Virus immunological status. New Microbiol 2007, 30, 393–398. [Google Scholar]
- Klutts, JS; Liao, RS; Dunne, WM, Jr; Gronowski, AM. Evaluation of a multiplexed bead assay for assessment of Epstein-Barr virus immunologic status. J. Clin. Microbiol 2004, 42, 4996–5000. [Google Scholar]
- Khan, IH; Ravindran, R; Yee, J; Ziman, M; Lewinsohn, DM; Gennaro, ML; Flynn, JL; Goulding, CW; DeRiemer, K; Lerche, NW; Luciw, PA. Profiling antibodies to Mycobacterium tuberculosis by multiplex microbead suspension arrays for serodiagnosis of tuberculosis. Clin. Vaccine Immunol 2008, 15, 433–438. [Google Scholar]
- Ro, HS; Koh, BH; Jung, SO; Park, HK; Shin, YB; Kim, MG; Chung, BH. Surface plasmon resonance imaging protein arrays for analysis of triple protein interactions of HPV, E6, E6AP, and p53. Proteomics 2006, 6, 2108–2111. [Google Scholar]
- Yuk, JS; Ha, KS. Array-based spectral SPR biosensor: Analysis of mumps virus infection. Methods Mol. Biol 2009, 503, 37–47. [Google Scholar]
- Ndao, M; Rainczuk, A; Rioux, MC; Spithill, TW; Ward, J. Is SELDI-TOF a valid tool for diagnostic biomarkers? Trends Parasitol 2010, 26, 561–567. [Google Scholar]
- Ndao, M; Spithill, TW; Caffrey, R; Li, H; Podust, VN; Perichon, R; Santamaria, C; Ache, A; Duncan, M; Powell, MR; Ward, BJ. Identification of novel diagnostic serum biomarkers for Chagas’ disease in asymptomatic subjects by mass spectrometric profiling. J. Clin. Microbiol 2010, 48, 1139–1149. [Google Scholar]
- Deckers, N; Dorny, P; Kanobana, K; Vercruysse, J; Gonzalez, AE; Ward, B; Ndao, M. Use of ProteinChip technology for identifying biomarkers of parasitic diseases: The example of porcine cysticercosis (Taenia solium). Exp. Parasitol 2008, 120, 320–329. [Google Scholar]
- Gavin, IM; Kukhtin, A; Glesne, D; Schabacker, D; Chandler, DP. Analysis of protein interaction and function with a 3-dimensional MALDI-MS protein array. Biotechniques 2005, 39, 99–107. [Google Scholar]
- Cui, Y; Wei, Q; Park, H; Lieber, CM. Nanowire nanosensors for highly sensitive and selective detection of biological and chemical species. Science 2001, 293, 1289–1292. [Google Scholar]
- Li, Y; Yang, HH; You, QH; Zhuang, ZX; Wang, XR. Protein recognition via surface molecularly imprinted polymer nanowires. Anal. Chem 2006, 78, 317–320. [Google Scholar]
- Gantelius, J; Hartmann, M; Schwenk, JM; Roeraade, J; Andersson-Svahn, H; Joos, TO. Magnetic bead-based detection of autoimmune responses using protein microarrays. N. Biotechnol 2009, 26, 269–276. [Google Scholar]
- Marina, O; Biernacki, MA; Brusic, V; Wu, CJ. A concentration-dependent analysis method for high density protein microarrays. J. Proteome Res 2008, 7, 2059–2068. [Google Scholar]
- Olle, EW; Sreekumar, A; Warner, RL; McClintock, SD; Chinnaiyan, AM; Bleavins, MR; Anderson, TD; Johnson, KJ. Development of an internally controlled antibody microarray. Mol. Cell Proteomics 2005, 4, 1664–1672. [Google Scholar]
- Hamelinck, D; Zhou, H; Li, L; Verweij, C; Dillon, D; Feng, Z; Costa, J; Haab, BB. Optimized normalization for antibody microarrays and application to serum-protein profiling. Mol. Cell Proteomics 2005, 4, 773–784. [Google Scholar]
- Andersson, O; Kozlowski, M; Garachtchenko, T; Nikoloff, C; Lew, N; Litman, DJ; Chaga, G. Determination of relative protein abundance by internally normalized ratio algorithm with antibody arrays. J. Proteome Res 2005, 4, 758–767. [Google Scholar]

© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Natesan, M.; Ulrich, R.G. Protein Microarrays and Biomarkers of Infectious Disease. Int. J. Mol. Sci. 2010, 11, 5165-5183. https://doi.org/10.3390/ijms11125165
Natesan M, Ulrich RG. Protein Microarrays and Biomarkers of Infectious Disease. International Journal of Molecular Sciences. 2010; 11(12):5165-5183. https://doi.org/10.3390/ijms11125165
Chicago/Turabian StyleNatesan, Mohan, and Robert G. Ulrich. 2010. "Protein Microarrays and Biomarkers of Infectious Disease" International Journal of Molecular Sciences 11, no. 12: 5165-5183. https://doi.org/10.3390/ijms11125165