Bacterial Stressors in Minimally Processed Food

Abstract
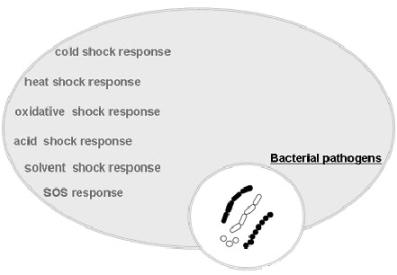
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Bacterial Pathogens
3. Key Factors Affecting the Survival and Growth of Pathogens
4. Stressors and Related Bacterial Stress Response
4.1. Cold Stress
4.2. Heat Stress
4.3. Acid and Solvent Stress
4.4. Oxidative Stress
4.5. Osmotic Stress
4.6. Irradiation
4.7. High Pressure Stress
4.8. Modified Atmosphere Packaging (MAP)
4.9. Biological Compounds
4.10. Antagonistic Microflora
4.11. Bacteriocins
4.12. Bacteriophages
5. Hurdle Technologies and Cross Protection
6. Conclusions
Acknowledgments
References
- Brackett, RE. Microbiological spoilage and pathogens in minimally processed refrigerated fruits and vegetables. In Minimally processed refrigerated fruits and vegetables; Wiley, RC, Ed.; Chapman and Hall: New York, NY, USA, 1994; pp. 269–312. [Google Scholar]
- Barry-Ryan, C; Pacussi, JM; O’Beirne, D. Quality of shredded carrots as affected by packaging film and storage temperature. J. Food Sci 2000, 65, 726–730. [Google Scholar]
- Beuchat, LR. Pathogenic microorganisms associated with fresh produce. J. Food Prot 1996, 59, 204–216. [Google Scholar]
- Parish, ME; Beuchat, LR; Suslow, TV; Harris, LJ; Garrett, EH; Farber, JN; Busta, FF. Methods to reduce/eliminate pathogens from fresh cut produce. Comp. Rev. Food Sci. Food Safety 2003, 2, 16–173. [Google Scholar]
- FDA, US Food and Drug Administration. Guidance for Industry. Guide to Minimize Microbial Food Safety Hazards of Fresh-cut Fruits and Vegetables. Guidance Contains Nonbinding Recommendations. Available online: http://www.cfsan.fda.gov/~dms/prodgui4.html#appe, Last updates: February 2008, accessed February 29, 2008.
- Ohlsson, T. Minimal processing technologies in the food industry; Ohlsson, T, Bengtsson, N, Eds.; Woodhead Publishing: Cambridge, UK, 2002. [Google Scholar]
- Leistner, L; Gould, GW. Hurdle Technologies: Combination Treatment for Food Stability, Safety and Quality; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002. [Google Scholar]
- Thomas, LV; Wimpenny, JWT. Investigation of the effect of combined variations in temperature, pH and NaCl concentrations on nisin inhibition of Listeria monocytogenes and Staphylococcus aureus. Appl. Environ. Microbiol 1996, 62, 2006–2012. [Google Scholar]
- McMeekin, TA; Presser, K; Ratkowsky, D; Ross, T; Salter, M; Tienungoon, S. Quantifying the hurdle concept by modelling the bacterial growth/no growth interface. Int. J. Food Microbiol 2000, 55, 93–98. [Google Scholar]
- Leistner, L. Basic aspects of food preservation by hurdle technology. Int. J. Food Microbiol 2000, 55, 181–186. [Google Scholar]
- Jones, K; Heaton, J. Microbial contamination of fruit and vegetables: evidence and issues. Microbiol 2006, 7, 28–31. [Google Scholar]
- Beneduce, L; Spano, G; Massa, S. Escherichia coli O157:H7 general characteristics, isolation and identification techniques. Annal. Microbiol 2003, 53, 511–527. [Google Scholar]
- Delaquis, P; Bach, S; Dinu, LD. Behavior of Escherichia coli O157:H7 in leafy vegetables. J. Food Prot 2007, 70, 1966–1974. [Google Scholar]
- Harris, LJ; Farber, JN; Beuchat, LR; Parish, ME; Suslow, TV; Garrett, EH; Busta, FF. Outbreaks associated with fresh produce: incidence, growth and survival of pathogens in fresh and fresh-cut produce. Comp. Rev. Food Sci. Food Safety 2003, 2, 78–141. [Google Scholar]
- Maki, D. Don’t eat the spinach - controlling foodborne infectious disease. New Engl. J. Med 2006, 355, 1952–1955. [Google Scholar]
- California Food Emergency Response Team 2007. Investigation of an Escherichia coli O157:H7 outbreak associated with dole pre-packaged spinach.
- Thurston-Enriquez, J; Watt, P; Dowd, SE; Enriquez, R; Pepper, IL; Gerbe, CP. Detection of protozoan parasites and microsporidia in irrigation waters for crop production. J. Food Prot 2002, 65, 378–382. [Google Scholar]
- Iwamoto, M; Taco, Bell; Taco, Johns. Escherichia coli in lettuce outbreaks 2006. CDC Outbreak Response and Surveillance Team Enteric Diseases Epidemiology Branch. Proceedings of the International Association of Food Protection’s 94th Annual Meeting (IAFP 2007), Disney's Contemporary Resort, Lake Buena Vista, FL, USA, July 8–11, 2007.
- Bell, C; Kyriakides, A. Background, Listeria a practical approach to the organism and its control in foods; Bell, C, Kyriakides, A, Eds.; Chapman and Hall: London, UK, 1998; pp. 3–29. [Google Scholar]
- MacGowan, AP; Bowker, K; McLauchlin, J; Bennett, PM; Reeves, DS. The occurrence and seasonal changes in the isolation of Listeria spp. in shop bought food stuffs, human faeces, sewage and soil from urban sources. Int. J. Food Microbiol 1994, 21, 325–334. [Google Scholar]
- Ukuku, DO; Fett, W. 2002, Behaviour of Listeria monocytogenes inoculated on cantaloupe surfaces and efficacy of washing treatments to reduce transfer from rind to fresh-cut pieces. J. Food Prot 2002, 65, 924–930. [Google Scholar]
- Ho, JL; Shands, KN; Freidland, G; Eckind, P; Fraser, DW. An outbreak of type 4b Listeria monocytogenes infection involving patients from eight Boston hospitals. Arch. Internal. Med 1986, 146, 520–524. [Google Scholar]
- Schlech, WF; Lavigne, PM; Bortolussi, RA; Allen, AC; Haldane, EV; Wort, AJ; Hightower, AW; Johnson, SE; King, SH; Nicholls, ES; Broome, CV. Epidemic listeriosis-evidence for transmission by food. New Eng. J. Med 1983, 308, 203–206. [Google Scholar]
- Austin, JW; Dodds, KL; Blanchfield, B; Farber, JM. Growth and toxin production by Clostridium botulinum on inoculated fresh-cut packaged vegetables. J. Food Prot 1998, 61, 324–328. [Google Scholar]
- Jacxsens, L; Devlieghere, F; Falcato, P; Debevere, J. Behaviour of Listeria monocytogenes and Aeromonas spp. on fresh-cut produce packaged under equilibrium-modified atmosphere. J. Food Prot 1999, 62, 1128–1135. [Google Scholar]
- Bolin, HR; Stafford, AE; King, AD, Jr; Huxsoll, CC. Factors affecting the storage stability of shredded lettuce. J. Food Sci 1977, 42, 1319–1321. [Google Scholar]
- Beuchat, LR. Surface disinfection of raw produce. Dairy, Food Environ. Sanitat 1992, 12, 6–9. [Google Scholar]
- Nguyen-the, C; Carlin, F. The microbiology of minimally processed fresh fruits and vegetables. Crit. Rev. Food Sci 1994, 34, 371–401. [Google Scholar]
- Izumi, H. Electrolyzed water as a disinfectant for fresh-cut vegetables. J. Food Sci 1999, 64, 536–539. [Google Scholar]
- Beuchat, LR; Ryu, J-H. Produce handling and processing practices. Emerg. Infect. Dis 1997, 3, 459–465. [Google Scholar]
- Adams, MR; Hartley, AD; Cox, LJ. Factors affecting the efficacy of washing procedures used in the production of prepared salads. Food Microbiol 1989, 6, 69–77. [Google Scholar]
- Park, C-M; Hung, Y-C; Doyle, MP; Ezeike, GOI; Kim, C. Pathogen reduction and quality of lettuce treated with electrolysed oxidizing and acidified chlorinated water. J. Food Sci 2001, 66, 1368–1372. [Google Scholar]
- Park, C-M; Beuchat, LR. Evaluation of sanitizers for killing Escherichia coli O157:H7, Salmonella and naturally occurring microorganisms on cantaloupes, honeydew melons and asparagus. Dairy, Food Environ. Sanit 1999, 19, 842–847. [Google Scholar]
- Zhang, S; Farber, JM. The effects of various disinfectants against Listeria monocytogenes on fresh-cut vegetables. Food Microbiol 1996, 13, 311–321. [Google Scholar]
- Sapers, GM; Simmons, GF. Hydrogen peroxide disinfection of minimally processed fruits and vegetables. Food Technol 1998, 52, 48–52. [Google Scholar]
- Karapinar, M; Gonul, SA. Effects of sodium bicarbonate, vinegar, acetic and citric acids on growth and survival of Yersinia enterocolitica. Int. J. Food Microbiol 1992, 16, 343–347. [Google Scholar]
- Burrows, JP; Weber, M; Buchwitz, M; Rozanov, V; Ladsttter-Weienmayer, A; Richter, A; de Beek, R; Hoogen, R; Bramstedt, K; Eichmann, K-U; Eisinger, M; Perner, D. The Global Ozone Monitoring Experiment (GOME): Mission Concept and First Scientific Results. J. Atmos. Sci 1999, 56, 151–175. [Google Scholar]
- Beuchat, LR. Survival of enterohemorrhagic Escherichia coli O157:H7 in bovine feces applied to lettuce and the effectiveness of chlorinated water as a disinfectant. J. Food Prot 1999, 62, 845–849. [Google Scholar]
- Aureli, P; Costantini, A; Zolea, S. Antimicrobial activity of some plant essential oils against Listeria monocytogenes. J. Food Prot 1992, 55, 344–348. [Google Scholar]
- Skandamis, PN; Nychas, GJE. Effect of oregano essential oil on microbiological and physical-chemical attributes of minced meat stored in air and modified atmosphere. J. App. Microbiol 2001, 91, 1011–1022. [Google Scholar]
- Singh, N; Singh, RK; Bhunia, AK; Stroshine, RL. Efficacy of chlorine dioxide, ozone and thyme essential oil or a sequential washing in killing Escherichia coli O157:H7 on lettuce and baby carrots. Lebensm. Wiss. U. Technol 2002, 35, 720–729. [Google Scholar]
- James, SJ. Novel physical methods for decontamination of produce. Proceedings of IAFP’s Third European Symposium on Food Safety: Advancements in Food Safety, Rome, Italy, October 18–19, 2007.
- Francis, GA; Thomas, C; O’Beirne, D. Review paper: The microbiological safety of minimally processed vegetables. Int. J. Food Sci. Technol 1999, 34, 1–22. [Google Scholar]
- Price, RJ; Lee, JS. Inhibition of Pseudomonas species by hydrogen peroxide producing lactobacilli. J. Milk Food Technol 1970, 33, 13–18. [Google Scholar]
- Arihara, K; Cassens, RG; Luchansky, JB. Characterization of bacteriocins from Enterococcus faecium with activity against Listeria monocytogenes. Int. J. Food Microbiol 1993, 19, 123–134. [Google Scholar]
- Gahan, CGM; O’Driscoll, B; Hill, C. Acid adaptation of Listeria monocytogenes can enhance survival in acidic foods and during milk fermentation. Appl. Environ. Microbiol 1996, 62, 3128–3132. [Google Scholar]
- Leyer, GL; Johnson, EA. Acid adaptation promotes survival of Salmonella spp. in cheese. Appl. Environ. Microbiol 1992, 58, 2075–2080. [Google Scholar]
- Artés, F; Allende, A. Processing lines and alternative preservation techniques to prolong the shelf-life of minimally fresh processed leafy vegetables. Eur. J. Hortic. Sci 2005, 70, 231–245. [Google Scholar]
- Hodges, DM; Toivonen, PMA. Quality of fresh-cut fruits and vegetables as affected by exposure to abiotic stress. Postharvest Biol. Technol 2008, 48, 155–162. [Google Scholar]
- Aguayo, E; Escalona, V; Artés, F. Quality of fresh-cut tomato as affected by type of cut, packaging, temperature and storage time. Eur. Food Res. Technol 2004, 219, 492–499. [Google Scholar]
- Phadtare, S. Recent Developments in Bacterial Cold-Shock Response. Curr. Issues Mol. Biol 2004, 6, 125–136. [Google Scholar]
- El-Sharoud, WM; Graumann, PL. Cold shock proteins aid coupling of transcription and translation in bacteria. Sci. Prog 2007, 90, 15–27. [Google Scholar]
- Wang, N; Yamanaka, K; Inouye, M. CspI, the ninth member of the CspA family of Escherichia coli is induced upon cold shock. J. Bacteriol 1999, 181, 1603–1609. [Google Scholar]
- Bae, W; Xia, B; Inouye, M; Severinov, K. Escherichia coli CspA-family RNA chaperones are transcription antiterminators. Proc. Natl. Acad. Sci. USA 2000, 97, 7784–7789. [Google Scholar]
- Phadtare, S; Tyagi, S; Inouye, M; Severinov, K. Three amino acids in Escherichia coli CspE surface-exposed aromatic patch are critical for nucleic acid melting activity leading to transcription antitermination and cold acclimation of cells. J. Biol. Chem 2002, 277, 46706–46711. [Google Scholar]
- Graumann, P; Wendrich, TM; Weber, MHW; Schröder, K; Marahiel, MA. A family of cold shock proteins in Bacillus subtilis is essential for cellular growth and for efficient protein synthesis at optimal and low temperature. Mol Microbiol 1997, 25, 741–756. [Google Scholar]
- Graumann, P; Marahiel, MA. Cold shock proteins CspB and CspC are major stationary-phase-induced proteins in B. subtilis. Arch. Microbiol 1999, 171, 135–138. [Google Scholar]
- Martín-Diana, A; Rico, D; Barry-Ryan, C; Frías, J; Henehan, G; Barat, J. Efficacy of steamer jet-injection as alternative to chlorine in fresh-cut lettuce. Postharvest Biol. Technol 2007, 45, 97–107. [Google Scholar]
- Kim, Y; Lounds-Singleton, AJ; Talcott, ST. Antioxidant phytochemical and quality changes associated with hot water immersion treatment of mangoes (Mangifera indica L.). Food Chem 2009, 115, 989–993. [Google Scholar]
- Steiner, A; Abreu, M; Correia, L; Beirão-da-Costa, S; Leitão, E; Beirão-da-Costa, ML; Empis, J; Moldão-Martins, M. Metabolic response to combined mild heat pre-treatments and modified atmosphere packaging on fresh-cut peach. Eur. Food Res. Technol 2006, 222, 217–222. [Google Scholar]
- Fallik, E. Prestorage hot water treatments (immersion, rinsing and brushing). Postharvest Biol. Technol 2004, 32, 125–134. [Google Scholar]
- Ray, B. Methods to Detect Stressed Microorganisms. J. Food Prot 1979, 42, 346–355. [Google Scholar]
- Mackey, BM; Miles, CA; Parsons, SE; Seymour, DA. Thermal denaturation of whole cells and cell components of Escherishia coli examined by differential scanning calorimetry. J. Gen. Microbiol 1991, 137, 2361–2374. [Google Scholar]
- Rowbury, RJ. Basis of Stress Adaptation, with Particular Reference to the Subversion of Stress Adaptation, and to the Invovement of Extracellular Components in Adaptation. In Microbial Stress Adaptation and Food Safety; Yousef, AE, Juneja, VK, Eds.; CRC Press: Boca Raton, FL, USA, 2002; Chapter 8. [Google Scholar]
- Bardwell, JCA; Craig, EA. Major heat shock gene of Drosophila and the Escherichia coli heat inducible dnaK gene are homologous. Proc. Natl. Acad. Sci. USA 1984, 81, 848–852. [Google Scholar]
- Yura, T; Kanemori, M; Morita, MT. The heat shock response: regulation and function. In Bacterial Stress Responses; Stortz, G, Hengge-Aronis, R, Eds.; ASM Press: Washington, D.C., USA, 2000; pp. 3–18. [Google Scholar]
- Mackey, BM; Derrick, C. Heat shock protein synthesis and thermotolerance in Salmonella typhimurium. J. Appl. Bacteriol 1990, 69, 373–383. [Google Scholar]
- Mendoza, JA; Wilson, M; Joves, F; Ackermann, E. Thermostabilization of enzymes by the chaperonin GroEL. Biotechnol. Tech 1996, 10, 535–540. [Google Scholar]
- Uyttendaele, M; Neyts, K; Vanderswalmen, H; Notebaert, E; Debevere, J. Control of Aeromonas on minimally processed vegetables by decontamination with lactic acid, chlorinated water, or thyme essential oil solution. Int. J. Food Microbiol 2004, 90, 263–271. [Google Scholar]
- Soliva-Fortuny, RC; Martìn-Belloso, O. New advances in extending the shelflife of fresh-cut fruits: a review. Trends Food Sci. Tech 2003, 14, 341–353. [Google Scholar]
- Alandes, L; Hernando, I; Quiles, A; Perez-Munuera, I; Lluch, MA. Cell wall stability of fresh-cut Fuji apples treated with calcium lactate. J. Food Sci 2006, 71, 615–620. [Google Scholar]
- Pieterse, B; Leer, RJ; Schuren, FHJ; van der Werf, MJ. Unravelling the multiple effects of lactic acid stress on Lactobacillus plantarum by transcription profiling. Microbiol 2005, 151, 3881–3894. [Google Scholar]
- Hartke, A; Bouche, S; Giard, J-C; Benachour, A; Boutibonnes, P; Auffray, Y. The lactic acid stress response of Lactococcus lactis subsp. lactis. Curr. Microbiol 1996, 33, 194–199. [Google Scholar]
- Polen, T; Rittmann, D; Wendisch, VF; Sahm, H. DNA Microarray analyses of the long-term adaptive response of Escherichia coli to acetate and propionate. Appl. Environ. Microbiol 2003, 69, 1759–1774. [Google Scholar]
- Kirkpatrick, C; Maurer, LM; Oyelakin, NE; Yoncheva, YN; Maurer, R; Slonczewski, JL. Acetate and formate stress, opposite responses in the proteome of Escherichia coli. J. Bacteriol 2001, 183, 6466–6477. [Google Scholar]
- Cotter, PD; Hill, C. Surviving the acid test, responses of Gram-positive bacteria to low pH. Microbiol. Mol. Biol. Rev 2003, 67, 429–453. [Google Scholar]
- Cotter, PD; Gahan, CG; Hill, C. A glutamate decarboxylase system protects Listeria monocytogenes in gastric fluid. Mol Microbiol 2001, 40, 465–475. [Google Scholar]
- Azcarate-Peril, MA; Altermann, E; Hoover-Fitzula, RL; Cano, RJ; Klaenhammer, TR. Identification and inactivation of genetic loci involved with Lactobacillus acidophilus acid tolerance. Appl. Environ. Microbiol 2004, 70, 5315–5322. [Google Scholar]
- Axe, DD; Bailey, JE. Transport of lactate and acetate through the energized cytoplasmic membrane of Escherichia coli. Biotechnol. Bioeng 1995, 47, 8–19. [Google Scholar]
- Shelef, LA. Antimicrobial effects of lactates, a review. J. Food Prot 1994, 57, 445–450. [Google Scholar]
- Diez-Gonzalez, F; Russell, JB. The effects of carbonylcyanide-m-chlorophenylhydrazone (CCCP) and acetate on Escherichia coli O157, H7 and K-12, uncoupling versus anion accumulation. FEMS Microbiol. Lett 1997, 151, 71–76. [Google Scholar]
- Russell, JB; Diez-Gonzalez, F. The effects of fermentation acids on bacterial growth. Adv. Microbial. Physiol 1998, 39, 205–234. [Google Scholar]
- Lemos, JA; Chen, YY; Burne, RA. Genetic and physiologic analysis of the groE operon and role of the HrcA repressor in stress gene regulation and acid tolerance in Streptococcus mutans. J. Bacteriol 183, 6074–6084.
- Mcdonnell, G; Denver, RA. Antiseptics and Disinfectants: Activity, Action, and Resistance. Clin. Microbiol. Rev 1999, 12, 147–179. [Google Scholar]
- Hallsworth, JE. Ethanol-Induced Water Stress in Yeast. J. Ferment. Bioeng 1998, 85, 125–137. [Google Scholar]
- Plotto, A; Baib, J; Narciso, JA; Brecht, JK; Baldwin, EA. Ethanol vapor prior to processing extends fresh-cut mango storage by decreasing spoilage, but does not always delay ripening. Postharvest Biol. Technol 2006, 39, 134–145. [Google Scholar]
- Bai, J; Baldwin, EA; Soliva Fortuny, RC; Mattheis, JP; Stanley, R; Perera, C; Brecht, JK. Effect of pretreatment of intact ‘Gala’ apple with ethanol vapor, heat, or 1 methylcyclopropene on quality and shelf life of fresh-cut slices. J. Am. Soc. Hort. Sci 2004, 129, 583–593. [Google Scholar]
- Xie, F. Disinfection byproducts in drinking water: form, analysis and control. In Technology & Industrial Arts; CRC Press: Boca Raton, FL, USA, 2003; p. 176. [Google Scholar]
- Khadre, MA; Yousef, AE. Sporicidal action of ozone and hydrogen peroxide: a comparative study. Int. J. Food Microbiol 2001, 71, 131–138. [Google Scholar]
- Ongeng, D; Devlieghere, F; Devevere, J; Coosemans, J; Ryckeboer, J. The efficacy of electrolysed oxidising water for inactivating spoilage microorganisms in process water on minimally processed vegetables. Int. J. Food Microbiol 2006, 109, 187–197. [Google Scholar]
- Guzel-Seydim, Z; Bever, PI; Greene, A. Efficacy of ozone to reduce bacterial populations in the presence of food components. Food Microbiol 2004, 21, 475–479. [Google Scholar]
- Parish, M; Beuchat, L; Suslow, T; Harris, L; Garret, E; Farber, J; Busta, F. Methods to reduce or eliminate pathogens from fresh and fresh-cut produce. Comp. Rev. Food Sci. Food Safety 2003, 2, 161–173. [Google Scholar]
- Storz, G; Imlay, JA. Oxidative stress. Curr. Opin. Microbiol 1999, 2, 188–194. [Google Scholar]
- Thybert, D; Avner, S; Lucchetti-Miganeh, C; Chéron, A; Barloy-Hubler, F. OxyGene: an innovative platform for investigating oxidative-response genes in whole prokaryotic genomes. BMC Genomics 2008, 9, 637. [Google Scholar]
- Cabiscol, E; Tamarit, J; Ros, J. Oxidative stress in bacteria and protein damage by reactive oxygen species. Int. Microbiol 2000, 3, 3–8. [Google Scholar]
- Poole, LB; Karplus, PA; Claiborne, A. Protein sulfenic acids in redox signaling. Annu. Rev. Pharmacol. Toxicol 2004, 44, 325–347. [Google Scholar]
- Aydogan, B; Marshall, DT; Swarts, SG; Turner, JE; Boone, AJ; Richards, NG; Bolch, WE. Site-specific OH attack to the sugar moiety of DNA: a comparison of experimental data and computational simulation. Radiat. Res 2002, 157, 38–44. [Google Scholar]
- Rivett, AJ. Regulation of intracellular protein turnover: covalent modification as a mechanism of marking proteins for degradation. Curr. Top. Cell. Regul 1986, 28, 291–337. [Google Scholar]
- Ritz, D; Patel, H; Doan, B; Zheng, M; Aslund, F; Storz, G; Beckwith, J. Thioredoxin 2 is involved in the oxidative stress response in Escherichia coli. J. Biol. Chem 2000, 275, 2505–2512. [Google Scholar]
- Smulevich, G; Jakopitsch, C; Droghetti, E; Obinger, C. Probing the structure and bifunctionality of catalase-peroxidase (KatG). J. Inorg. Biochem 2006, 100, 568–585. [Google Scholar]
- Jonsson, TJ; Lowther, WT. The peroxiredoxin repair proteins. Subcell. Biochem 2007, 44, 115–141. [Google Scholar]
- Putz, S; Gelius-Dietrich, G; Piotrowski, M; Henze, K. Rubrerythrin and peroxiredoxin: two novel putative peroxidases in the hydrogenosomes of the microaerophilic protozoon Trichomonas vaginalis. Mol. Biochem. Parasitol 2005, 142, 212–223. [Google Scholar]
- Smith, J; Shrift, A. Phylogenetic distribution of glutathione peroxidase. Comp. Biochem. Physiol. B 1979, 63, 39–44. [Google Scholar]
- van Pee, KH. Bacterial haloperoxidases and their role in secondary metabolism. Biotechnol. Adv 1990, 8, 185–205. [Google Scholar]
- Wuerges, J; Lee, JW; Yim, YI; Yim, HS; Kang, SO; Djinovic Carugo, K. Crystal structure of nickel-containing superoxide dismutase reveals another type of active site. Proc. Natl. Acad. Sci. USA 2004, 101, 8569–8574. [Google Scholar]
- Lombard, M; Touati, D; Fontecave, M; Niviere, V. Superoxide reductase as a unique defense system against superoxide stress in the microaerophile Treponema pallidum. J. Biol. Chem 2000, 275, 27021–27026. [Google Scholar]
- Heylen, K; Vanparys, B; Gevers, D; Wittebolle, L; Boon, N; De Vos, P. Nitric oxide reductase (norB) gene sequence analysis reveals discrepancies with nitrite reductase (nir) gene phylogeny in cultivated denitrifiers. Environ. Microbiol 2007, 9, 1072–1077. [Google Scholar]
- Abee, T; Wouters, JA. Microbial stress response in minimal processing. Int. J. Food Microbiol 1999, 50, 65–91. [Google Scholar]
- Wood, JM; Bremer, E; Csonka, LN; Kraemer, R; Poolman, B; van der Heide, T; Smith, LT. Osmosensing and osmoregulatory compatible solute accumulation by bacteria. Comp. Biochem. Physiol. A Mol. Integr. Physiol 2001, 130, 437–460. [Google Scholar]
- Csonka, LN. Physiological and Genetic Responses of Bacteria to Osmotic-Stress. Microbiol. Rev 1989, 53, 121–147. [Google Scholar]
- Corry, JEL. The Water Relations and Heat Resistance of Micro-organisms. Prog. Ind. Microbiol 1972, 12, 73–108. [Google Scholar]
- Poolman, B; Spitzer, JJ; Wood, JM. Bacterial osmosensing: roles of membrane structure and electrostatics in lipid-protein and protein-protein interactions. Biochim Biophys Acta 2004, 1666, 88–104. [Google Scholar]
- Poolman, B; Blount, P; Folgering, JHA; Friesen, RHE; Moe, PC; van der Heide, T. How do membrane proteins sense water stress. Mol. Microbiol 2002, 44, 889–902. [Google Scholar]
- McLaggan, D; Naprstek, J; Buurman, E; Epstein, W. Interdependence of K+ and glutamate accumulation during osmotic adaptation of Escherichia coli. J. Biol. Chem 1994, 269, 1911–1917. [Google Scholar]
- Samelis, J; Sofos, JN. Chapter 9: Strategies to Control Stress-Adapted Pathogens. In Microbial Stress Adaptation and Food Safety; Yousef, AE, Juneja, VK, Eds.; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Canovas, D; Fletcher, SA; Hayashi, M; Csonka, LN. Role of Trehalose in Growth at High Temperature of Salmonella enterica Serovar Typhimurium. J. Bacteriol 2001, 183, 3365–3371. [Google Scholar]
- González-Aguilar, GA; Ruiz-Cruz, S; Cruz-Valenzuela, R; Ayala-Zavala, JF; de la Rosa, LA; Alvarez-Parrilla, E; Gutiérrez-López, GF. New Technologies to Preserve Quality of Fresh-Cut Produce. In Food Engineering: Integrated Approaches; Barbosa-Cánovas, GV, Welti-Chanes, J, Parada-Arias, E, Eds.; Springer: Berlin, Germany, 2008. [Google Scholar]
- Lado, B; Yousef, A. Alternative food-preservation technologies: efficacy and mechanisms. Microbes Infect 2002, 4, 433–440. [Google Scholar]
- Bintsis, T; Litopoulou-Tzanetaki, E; Robinson, R. Existing and potential applications of ultraviolet light in the food industry—a critical review. J. Sci. Food Agric 2000, 80, 637–645. [Google Scholar]
- Gómez-López, V; Devlieghere, F; Bonduellea, V; Debevere, J. Intense light pulses decontamination of minimally processed vegetables and their shelf-life. Int. J. Food Microbiol 2005, 103, 79–89. [Google Scholar]
- Takeshita, K; Shibato, J; Sameshima, T; Fukunaga, S; Isobe, S; Arihara, K; Itoh, M. Damage of yeast cells induced by pulsed light irradiation. Int. J. Food Microbiol 2003, 85, 151–158. [Google Scholar]
- Kiyosawa, K; Tanaka, M; Matsunaga, T; Nikaido, O; Yamamoto, K. Amplified UvrA protein can ameliorate the ultraviolet sensitivity of an Escherichia coli recA mutant. Mutat. Rese 2001, 487, 149–156. [Google Scholar]
- Lopez-Malo, A; Palou, E; Barboza-Canovas, GV; Swanson, BG; Welit-Chanes, J. Minimally processed foods with high hydrostatic pressure. In Trends in Food Engineering; Lozano, JE, Ed.; Technomic Pub Co: Lancaster, PA, USA, 2000; pp. 267–286. [Google Scholar]
- Ikeda, F; Baba, T; Como, G; Ohtsubo, T; Lizada, MCC. Effect of hydrostatic pressure on postharvest physiology in fruit. Acta. Hortic 2000, 518, 101–106. [Google Scholar]
- Dede, S; Alps, H; Bayindirli, A. High hydrostatic pressure treatment and storage of carrot and tomato juices: antioxidant activity and microbial safety. J. Sci. Food Agric 2007, 87, 773–782. [Google Scholar]
- Yanga, DS; Balandrán-Quintanab, RR; Ruizc, CF; Toledoc, RT; Kaysa, SJ. Effect of hyperbaric, controlled atmosphere, and UV treatments on peach volatiles. Postharvest Biol. Technol 2009, 51, 334–341. [Google Scholar]
- Pradillon, F; Gaill, F. Pressure and life: some biological strategies. Rev. Environ. Sci. Biotechnol 2007, 6, 181–195. [Google Scholar]
- Balny, C; Masson, P; Heremans, K. High pressure effects on biological macromolecules: from structural changes to alteration of cellular processes. Biochim. Biophys. Acta 2002, 1595, 3–10. [Google Scholar]
- Balny, C; Mozhaev, VV; Lange, R. Hydrostatic pressure and proteins: Basic concepts and new data. Comp. Biochem. Physiol 1997, 116, 299–304. [Google Scholar]
- Malone, AS; Chung, Y-K; Yousef, AE. Genes of Escherichia coli O157:H7 that are involved in high-pressure resistance. Appl. Environ. Microbiol 2006, 72, 2661–2671. [Google Scholar]
- Rico, D; Martin-Diana, AB; Barat, JM; Barry-Ryan, C. Extending and measuring the quality of fresh-cut fruit and vegetables: a review. Trends Food Sci. Technol 2007, 18, 373–386. [Google Scholar]
- Novak, JS; Sapers, GM; Juneja, VK. Microbial Safety of Minimally Processed Foods; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Matthews, KR. Microbiology of Fresh Produce; ASM Press: Washington, D.C., USA, 2006. [Google Scholar]
- Dixon, NM; Kell, DB. The inhibition by CO2 of the growth and metabolism of micro-organisms: a review. J. Appl. Bacteriol 1989, 67, 109–136. [Google Scholar]
- Hudson, JA; Mott, SJ; Penney, N. Growth of Listeria monocytogenes, Aeromonas hydrophila and Yersinia enterocolitica on vacuum and saturated carbon dioxide controlled atmosphere packaged sliced roast beef. J. Food Prot 1994, 57, 204–208. [Google Scholar]
- Marino, M; Bersani, C; Comi, G. Impedance measurement to study antimicrobial activity of essential oils from Lamiaceae and Compositae. Int. J. Food Microbiol 2001, 67, 187–195. [Google Scholar]
- Burt, S. Essential oils: their antibacterial properties and potential applications in foods - a review. Int. J. Food Microbiol 2004, 94, 223–253. [Google Scholar]
- Lanciotti, R; Gianotti, A; Patrignani, F; Belletti, N; Guerzoni, ME; Gardini, F. Use of natural aroma compounds to improve shelflife and safety of minimally processed fruits. Trends Food Sci. Technol 2004, 15, 201–208. [Google Scholar]
- Gutierrez, J; Rodriguez, G; Barry-Ryan, C; Bourke, P. Efficacy of plant essential oils against food-borne pathogens and spoilage bacteria associated with ready to eat vegetables: antimicrobial and sensory screening. J. Food Prot 2008, 71, 1846–1854. [Google Scholar]
- Gutierrez, J; Barry-Ryan, C; Bourke, P. Antimicrobial activity of plant essential oils using food model media: Efficacy, synergistic potential and interactions with food components. Food Microbiol 2009, 26, 142–150. [Google Scholar]
- Campaniello, D; Bevilacqua, A; Sinigaglia, M; Corbo, MR. Chitosan: Antimicrobial activity and potential applications for preservino minimally processed strawberries. Food Microbiol 2009, 25, 992–1000. [Google Scholar]
- Chien, PJ; Sheu, F; Lin, HR. Coating citrus (Murcott tangor) fruit with low molecular weight chitosan increases postharvest quality and shelf life. Food Chem 2007, 100, 1160–1164. [Google Scholar]
- Martin-Diana, AB; Rico, D; Frias, J; Mulcahy, J; Henehan, GTM; Barry-Ryan, C. Whey permeate as a bio-preservative for shelf life maintenance of fresh-cut vegetables. Inn. Food Sci. Emerg. Tech 2006, 7, 112–123. [Google Scholar]
- Cai, Y; Ng, LK; Farber, JM. Isolation and characterization of nisin-producing Lactococcus lactis subsp. lactis from bean-sprouts. J. Appl. Microbiol 1997, 83, 499–507. [Google Scholar]
- Vescovo, M; Torriani, S; Orsi, C; Macchiarolo, F; Scolari, G. Application of antimicrobial-producing lactic acid bacteria to control pathogens in ready-to-use vegetables. J. Appl. Bacteriol 1996, 81, 113–119. [Google Scholar]
- Liao, C-H; Sapers, GM. Influence of soft rot bacteria on growth of Listeria monocytogenes on potato tuber slices. J. Food Prot 1999, 62, 343–348. [Google Scholar]
- Allende, A; Martinez, B; Selma, V; Gil, MI; Suarez, JE; Rodriguez, A. Growth and bacteriocin production by lactic acid bacteria in vegetable broth and their effectiveness at reducing Listeria monocytogenes in vitro and in fresh-cut lettuce. Food Microbiol 2007, 24, 759–766. [Google Scholar]
- Carlin, F; Nguyen-the, C; Morris, CE. The influence of the background microflora on the fate of Listeria monocytogenes on minimally processed fresh broad leaved endive (Cichorium endivia var. latifolia). J. Food Prot 1996, 59, 698–703. [Google Scholar]
- Francis, GA; O’Beirne, D. Effects of storage atmosphere on Listeria monocytogenes and competing microflora using a surface model system. Int. J. Food Sci. Technol 1998, 33, 465–476. [Google Scholar]
- Duffy, G; Whiting, RC; Sheridan, JJ. The effects of a competitive microflora, pH and temperature on the growth kenetics of Escherichia coli O157:H7. Food Microbiol 1999, 16, 299–307. [Google Scholar]
- Liao, C-H; Cooke, PH. Response to trisodium phosphate treatment of Salmonella Chester attached to fresh-cut green pepper slices. Can. J. Microbiol 2001, 47, 25–32. [Google Scholar]
- Galvez, A; Lopez, RL; Abriouel, H; Valdivia, E; Omar, NB. Application of bacteriocins in the control of foodborne pathogenic and spoilage bacteria. Crit. Rev. Biotechnol 2008, 28, 125–152. [Google Scholar]
- Bennik, MHJ; Vorstman, W; Smid, EJ; Gorris, LGM. The influence of oxygen and carbon dioxide on the growth of prevalent Enterobacteriaceae and Pseudomonas species isolated from fresh and controlled-atmosphere-stored vegetables. Food Microbiol 1998, 15, 459–469. [Google Scholar]
- Barry-Ryan, C; Pacussi, JM; O’Beirne, D. Quality of shredded carrots as affected by packaging film and storage temperature. J. Food Sci 2000, 65, 726–730. [Google Scholar]
- Beuchat, LR. Surface decontamination of fruits and vegetables eaten raw: a review; World Health Organization, Food Safety Unit: Geneva, Switzerland, 1998. WHO/FSF/FOS/98.2, Available online: http://www.who.int/foodsafety/publications/fs_management/en/surface_decon.pdf/ (accessed May 25, 2009).
- Sun, Z; Zhong, J; Liang, X; Liu, J; Chen, X; Huan, L. Novel Mechanism for Nisin Resistance via Proteolytic Degradation of Nisin by the Nisin Resistance Protein NSR. Antimicrob. Agents Chemother 2009, 53, 1964–1973. [Google Scholar]
- Leverentz, B; Conway, WS; Alavidze, Z; Janisiewicz, WJ; Fuchs, Y; Camp, MJ; Chighladze, E; Sulakvelidze, A. Examination of bacteriophage as a biocontrol method for Salmonella on fresh-cut fruit—a model study. J. Food Prot 2001, 64, 1116–1121. [Google Scholar]
- Barry-Ryan, C; O’Beirne, D. Effects of slicing method on the quality and storage-life of modified atmosphere packaged carrot discs. J. Food Sci 1998, 63, 851–856. [Google Scholar]
- Leverentz, B; Conway, WS; Camp, MJ; Janisiewicz, WJ; Abuladze, T; Yang, M; Saftner, R; Sulakvelidze, A. Biocontrol of Listeria monocytogenes on Fresh-Cut Produce by Treatment with Lytic Bacteriophages and a Bacteriocin. Appl. Environ. Microbiol 2003, 69, 4519–4526. [Google Scholar]
- Coffey, A; Ross, RP. Bacteriophage-resistance systems in dairy starter strains: molecular analysis to application. Antonie Leeuwenhoek 2002, 82, 303–321. [Google Scholar]
- Yang, JM; DeUrraza, PJ; Matvienko, N; O’Sullivan, DJ. Involvement of the LlaKR2I Methylase in Expression of the AbiR Bacteriophage Defense System in Lactococcus lactis subsp. lactis biovar diacetylactis KR2. J. Bacteriol 2006, 188, 1920–1928. [Google Scholar]
- Chopin, M-C; Chopin, A; Bidnenko, E. Phage abortive infection in lactococci: variations on a theme. Curr. Opin. Microbiol 2005, 8, 473–479. [Google Scholar]
- Hazan, R; Engelberg-Kulka, H. Escherichia coli mazEF-mediated cell death as a defense mechanism that inhibits the spread of phage P1. Mol. Gen. Genomics 2004, 272, 227–234. [Google Scholar]
- Johnson, EA. Chapter 4: Microbial Adaptation and Survival in Foods. In Microbial Stress Adaptation and Food Safety; Yousef, AE, Juneja, VK, Eds.; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Rowe, MT; Kirk, RB. Cross-protection phenomenon in Escherichia coli strains harbouring cytotoxic necrotizing factors and cytolethal distending toxins.”. Lett. Appl. Microbiol 2001, 32, 67–70. [Google Scholar]
- Lou, YQ; Yousef, AE. Adaptation to sublethal environmental stresses protects Listeria monocytogenes against lethal preservation factors. Appl. Environ. Microbiol 1997, 63, 1252–1255. [Google Scholar]
- Periago, PM; Schaik, WV; Abee, T; Wouters, JA. Identification of Proteins Involved in the Heat Stress Response of Bacillus cereus ATCC 14579. Appl. Environ. Microbiol 2002, 68, 3486–3495. [Google Scholar]
- Leyer, GJ; Johnson, EA. Acid Adaptation Induces Cross-Protection against Environmental Stresses in Salmonella Typhimurium. Appl. Environ. Microbiol 1993, 59, 1842–1847. [Google Scholar]
- Mattick, KL; Jorgensen, F; Legan, JD; Lappin-Scott, HM; Humphrey, TJ. Habituation of Salmonella spp. at Reduced Water Activity and Its Effect on Heat Tolerance. Appl. Environ. Microbiol 2000, 66, 4921–4925. [Google Scholar]
- Phadtare, S; Inouye, M. Role of CspC and CspE in regulation of expression of RpoS and UspA, the stress response proteins in Escherichia coli. J. Bacteriol 2001, 183, 1205–1214. [Google Scholar]
- Derzelle, S; Hallet, B; Ferain, T; Delcour, J; Hols, P. Improved adaptation to cold-shock, stationary phase, and freezing stresses in Lactobacillus plantarum overproducing cold-shock proteins. Appl. Environ. Microbiol 2003, 69, 4285–4290. [Google Scholar]
- Katzif, S; Danavall, D; Bowers, S; Balthazar, JT; Shafer, WM. The major cold shock gene, cspA, is involved in the susceptibility of Staphylococcus aureus to an antimicrobial peptide of human cathepsin. G Infect. Immun 2003, 71, 4304–4312. [Google Scholar]
- Mangoli, S; Sanzgiri, VR; Mahajan, SK. A common regulator of cold and radiation response in Escherichia coli. J. Environ. Pathol. Toxicol. Oncol 2001, 20, 23–26. [Google Scholar]
- Yamanaka, K; Inouye, M. Growth-phasedependent expression of cspD, encoding a member of the CspA family in Escherichia coli. J. Bacteriol 1997, 179, 5126–5130. [Google Scholar]
- Schmid, B; Klumpp, J; Raimann, E; Loessner, MJ; Stephan, R; Tasara, T. Role of cold shock proteins in growth of Listeria monocytogenes under cold and osmotic stress conditions. Appl. Environ. Microbiol 2009, 75, 1621–1627. [Google Scholar]
- Ravishankar, S; Juneja, VK. Adaptation or Resistance Responses of Microorganisms to Stresses in the Food Processing Environment. In Microbial Stress Adaptation and Food Safety; Yousef, AE, Juneja, VK, Eds.; CRC Press: Boca Raton, FL, USA, 2002; Chapter 5. [Google Scholar]
- Ferrer, M; Chernikova, TN; Yakimov, MM; Golyshin, PN; Timmis, KN. Chaperonins govern growth of Escherichia coli at low temperatures. Nat. Biotechnol 2003, 21, 1266–1267. [Google Scholar]
- Juneja, VK; Klein, PG; Marmer, BS. Heat shock and thermotolerance of Escherichia coli O157:H7 in a model beef gravy system and ground beef. J. Appl. Microbiol 1998, 84, 677–684. [Google Scholar]
- Foster, JW. Low pH adaptation and the acid tolerance response of Salmonella typhimurium. Crit. Rev. Microbiol 1995, 21, 215–237. [Google Scholar]
- Masters, CI; Miles, CA; Mackey, BM. Survival and biological activity of heat damaged DNA. Lett. Appl. Microbiol 1998, 27, 279–282. [Google Scholar]
- El-Sharoud, WM. Ribosome inactivation for preservation: concepts and reservations. Sci. Progr 2004, 87, 137–152. [Google Scholar]
- Kim, YJ; Moon, MH; Song, JY; Smith, CP; Hong, S-K; Chang, YK. Acidic pH shock induces the expressions of a wide range of stress-response genes. BMC Genomics 2008, 9, 604. [Google Scholar] [Green Version]
- Abram, F; Starr, E; Karatzas, KAG; Matlawska-Wasowska, K; Boyd, A; Wiedmann, M; Boor, KJ; Connally, D; O’Byrne, CP. Identification of Components of the Sigma B Regulon in Listeria monocytogenes That Contribute to Acid and Salt Tolerance. Appl. Environ. Microbiol 2008, 74, 6848–6858. [Google Scholar]
- Streit, F; Delettre, J; Corrieu, G; Béal, C. Acid adaptation of Lactobacillus delbrueckii subsp. bulgaricus induces physiological responses at membrane and cytosolic levels that improves cryotolerance. J. Appl. Microbiol 2008, 105, 1071–1080. [Google Scholar]
- Bang, I-S; Frye, JG; McClelland, M; Velayudhan, J; Fang, FC. Alternative sigma factor interactions in Salmonella : σE and σH promote antioxidant defences by enhancing σS levels. Mol. Microbiol 2005, 56, 811–823. [Google Scholar]
- Gebhard, S; Hümpel, A; McLellan, AD; Cook, GM. The alternative sigma factor SigF of Mycobacterium smegmatis is required for survival of heat shock, acidic pH and oxidative stress. Microbiol 2008, 154, 2786–2795. [Google Scholar]
- Smirnova, GV; Zakirova, ON; Oktyabrskii, ON. The Role of Antioxidant Systems in the Cold Stress Response of Escherichia coli. Microbiology 2001, 70, 45–50. [Google Scholar]
- Gottesman, S. Proteolysis in bacterial regulatory circuits. Annu. Rev. Cell Dev. Biol 2003, 19, 565–587. [Google Scholar]
- Sauer, RT; Bolon, DN; Burton, BM; Burton, RE; Flynn, JM; Grant, RA. Sculpting the proteome with AAA(+) proteases and disassembly machines. Cell 2004, 119, 9–18. [Google Scholar]
- Hanson, PI; Whiteheart, SW. AAA+ proteins: have engine, will work. Nat. Rev. Mol. Cell Biol 2005, 6, 519–529. [Google Scholar]
- Neher, SB; Villen, J; Oakes, EC; Bakalarski, CE; Sauer, RT; Gygi, SP; Baker, TA. Proteomic profiling of ClpXP substrates after DNA damage reveals extensive instability within SOS regulon. Mol. Cell 2006, 22, 193–204. [Google Scholar]
- Pruteanu, M; Baker, TA. Controlled degradation by ClpXP protease tunes the levels of the excision repair protein UvrA to the extent of DNA damage. Mol. Microbiol 2009, 71, 912–924. [Google Scholar]
- Spano, G; Capozzi, V; Vernile, A; Massa, S. Cloning, molecular characterization and expression analysis of two small heat shock genes isolated from wine Lactobacillus plantarum. J. Appl. Microbiol 2004, 97, 774–782. [Google Scholar]
- Spano, G; Beneduce, L; Perrotta, C; Massa, S. Cloning and characterization of the hsp 18.55 gene, a new member of the small heat shock genes family isolated from wine Lactobacillus plantarum. Res. Microbiol 2005, 156, 219–224. [Google Scholar]
- Fiocco, D; Capozzi, V; Goffin, P; Hols, P; Spano, G. Improved adaptation to heat, cold, and solvent tolerance in Lactobacillus plantarum. Appl. Microbiol. Biotechnol 2007, 77, 909–915. [Google Scholar]
- Welch, TJ; Farewell, A; Neidhardt, FC; Bartlett, DH. Stress response of Escherichia coli to elevated hydrostatic pressure. J. Bacteriol 1993, 175, 7170–7177. [Google Scholar]
- Chen, H; Neetoo, H; Ye, M; Joerger, RD. Differences in pressure tolerance of Listeria monocytogenes strains are not correlated with other stress tolerances and are not based on differences in CtsR. Food Microbiol 2009, 26, 404–408. [Google Scholar]
- McDougald, D; Gong, L; Srinivasan, S; Hild, E; Thompson, L; Takayama, K; Rice, SA; Kjelleberg, S. Defences against oxidative stress during starvation in bacteria. Antonie van Leeuwenhoek 2002, 81, 3–13. [Google Scholar]
- Lucchetti-Miganeh, C; Burrowes, E; Baysse, C; Ermel, G. The post-transcriptional regulator CsrA plays a central role in the adaptation of bacterial pathogens to different stages of infection in animal hosts. Microbiology 2008, 154, 16–29. [Google Scholar]
- Giotis, ES; Julotok, M; Wilkinson, BJ; Blair, IS; McDowell, DA. Role of sigma B factor in the alkaline tolerance response of Listeria monocytogenes 10403S and cross-protection against subsequent ethanol and osmotic stress. J. Food Prot 2008, 71, 1481–1485. [Google Scholar]
- Shahjee, HM; Banerjee, K; Ahmad, F. Comparative analysis of naturally occurring L-amino acid osmolytes and their D-isomers on protection of Escherichia coli against environmental stresses. J. Biosci 2002, 27, 515–520. [Google Scholar]
- Chattopadhyay, MK. The cryoprotective effects of glycine betaine on bacteria. Trends Microbiol 2002, 10, 311. [Google Scholar]
- Chattopadhyay, MK; Kern, R; Mistou, MY; Dandekar, AM; Uratsu, SL; Richarme, G. The chemical chaperone proline relieves the thermosensitivity of a dnaK deletion mutant at 42°C. J. Bacteriol 2004, 186, 8149–8152. [Google Scholar]
- VanBogelen, RA; Neidhardt, FC. Ribosomes as sensors of heat and cold shock in Escherichia coli. Proc. Natl. Acad. Sci. USA 1990, 87, 5589–5593. [Google Scholar]
- Brissette, JL; Russel, M; Weiner, L; Model, P. Phage shock protein, a stress protein of Escherichia coli. Proc. Natl. Acad. Sci. USA 1990, 87, 862–866. [Google Scholar]
- Lloyd, LJ; Jones, SE; Jovanovic, G. Identification of a new member of the phage shock protein response in Escherichia coli, the phage shock protein G (PspG). J. Biol. Chem 2004, 279, 55707–55714. [Google Scholar]
- Hassani, AS; Malekzadeh, F; Amirmozafari, N; Hamdi, K; Ordouzadeh, N; Ghaemi, A. Phage Shock Protein G. A Novel Ethanol-Induced Stress Protein in Salmonella typhimurium. Curr. Microbiol 2009, 58, 239–244. [Google Scholar]
- Hussain, I; Mahmood, MS; Akhtar, M; Khan, A. Prevalence of Campylobacter species in meat, milk and other food commodities in Pakistan. Food Microbiol 2007, 24, 219–222. [Google Scholar]
- Bhagwat, AA. Microbiological safety of fresh-cut produce: where are we now? In Microbiology of Fresh Produce; Matthews, KR, Ed.; American Society for Microbiology: Washington, D.C., USA, 2006; pp. 121–165. [Google Scholar]
- Skandamisa, PN; Yoona, Y; Stopfortha, JD; Kendallb, PA; Sofos, JN. Heat and acid tolerance of Listeria monocytogenes after exposure to single and multiple sublethal stresses. Food Microbiol 2008, 25, 294–303. [Google Scholar]
- Chua, DK; Goh, R; Saftner, A; Bhagwat, AA. Fresh-Cut Lettuce in Modified Atmosphere Packages Stored at Improper Temperatures Supports Enterohemorrhagic E. coli Isolates to Survive Gastric Acid Challenge. J. Food Sci 2008, 73, M148–153. [Google Scholar]
- Artés, F; Gómez, P; Aguayo, E; Escalona, V; Artés-Hernández, F. Sustainable sanitation techniques for keeping quality and safety of fresh-cut plant commodities. Postharvest Biol. Technol 2009, 51, 287–296. [Google Scholar]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Capozzi, V.; Fiocco, D.; Amodio, M.L.; Gallone, A.; Spano, G. Bacterial Stressors in Minimally Processed Food. Int. J. Mol. Sci. 2009, 10, 3076-3105. https://doi.org/10.3390/ijms10073076
Capozzi V, Fiocco D, Amodio ML, Gallone A, Spano G. Bacterial Stressors in Minimally Processed Food. International Journal of Molecular Sciences. 2009; 10(7):3076-3105. https://doi.org/10.3390/ijms10073076
Chicago/Turabian StyleCapozzi, Vittorio, Daniela Fiocco, Maria Luisa Amodio, Anna Gallone, and Giuseppe Spano. 2009. "Bacterial Stressors in Minimally Processed Food" International Journal of Molecular Sciences 10, no. 7: 3076-3105. https://doi.org/10.3390/ijms10073076