Rolly Protein (ROLP)-Epb4.1/3: A Potential Protein-Protein Interaction Relevant for the Maintenance of Cell Adhesion


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
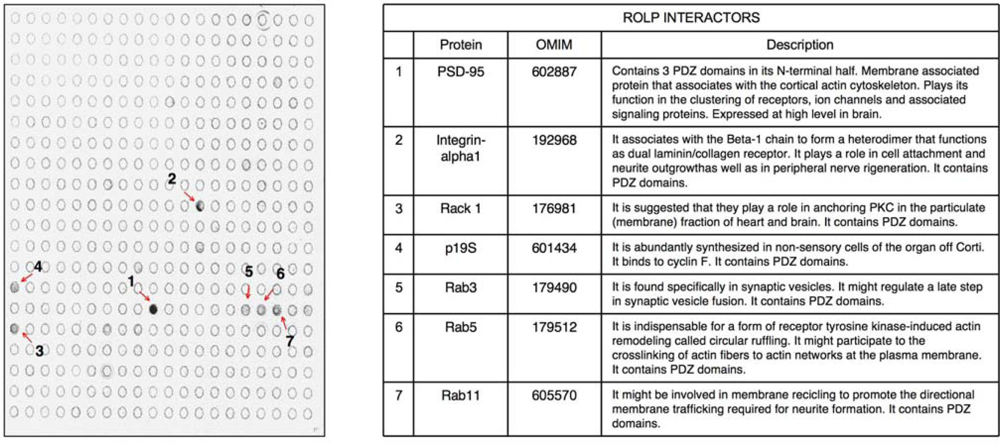
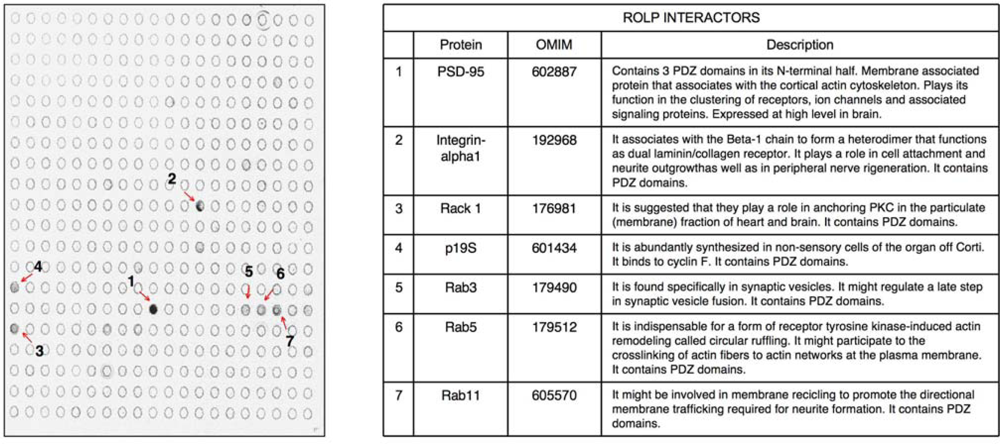
 kp1 (NP_035673), Rab3 (NP_848805), Rab5 (NP_080163) and Rab11 (NP_059078). Interestingly all these possible partners of ROLP share functional and structural common hallmarks: i) they all contain one or more PDZ domains ii) they are localized at the plasma membrane level where they play a role in membrane recycling, membrane fusion in synaptic vesicle formation, and cell anchoring at membrane level.
kp1 (NP_035673), Rab3 (NP_848805), Rab5 (NP_080163) and Rab11 (NP_059078). Interestingly all these possible partners of ROLP share functional and structural common hallmarks: i) they all contain one or more PDZ domains ii) they are localized at the plasma membrane level where they play a role in membrane recycling, membrane fusion in synaptic vesicle formation, and cell anchoring at membrane level.2. Experimental Section
2.1. Cell adhesion assay
2.2. Real time quantitative RT-PCR
2.3. Gene silencing
2.4. Cell culture and transient transfection
2.5. Antibody array
2.6. Immunoprecipitation
2.7. Western blot analysis
3. Results
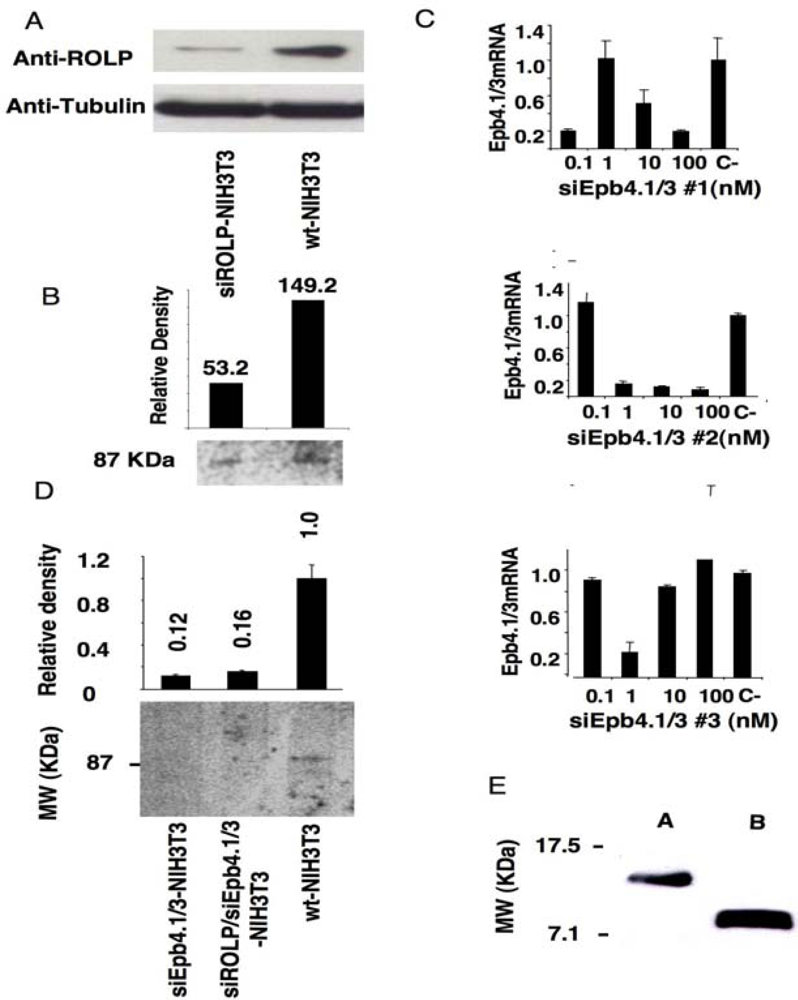
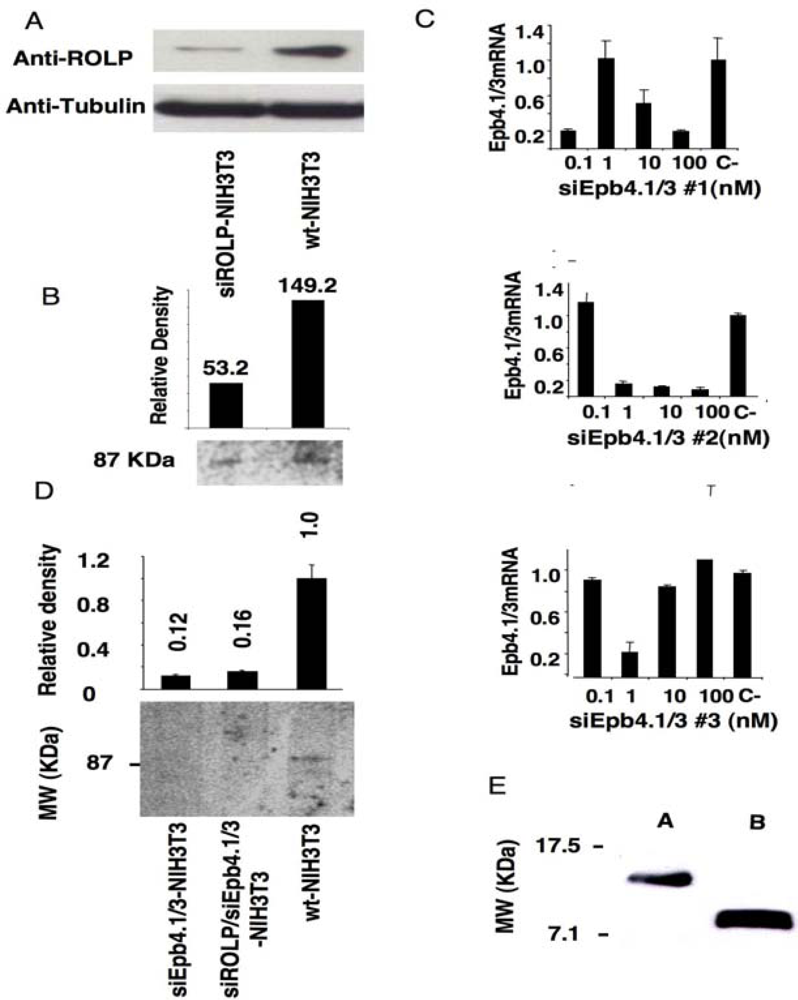
3.1. Immunoprecipitation assay coupled to gene silencing technology in NIH3T3 cells suggests a possible interaction of ROLP with Epb4.1/3
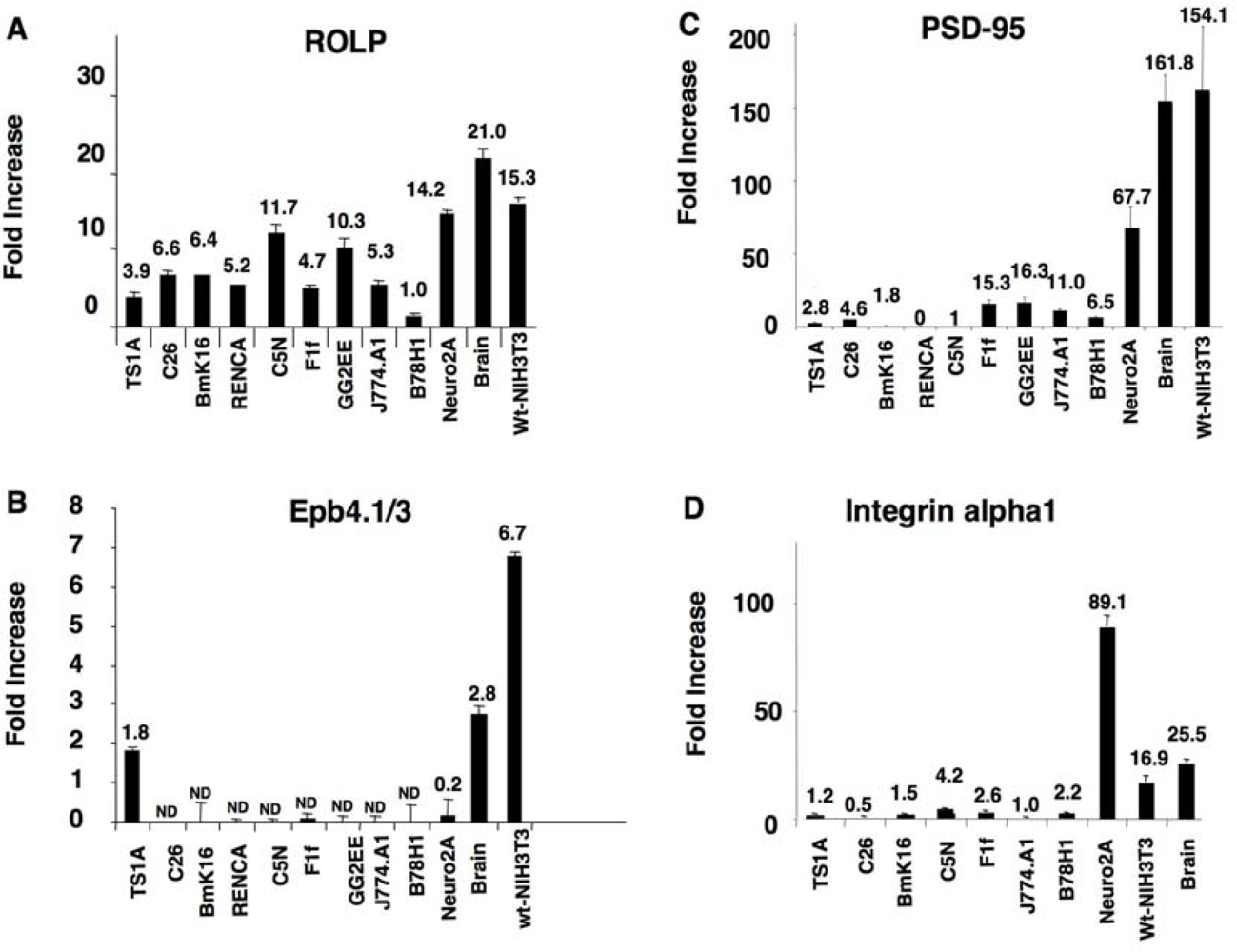
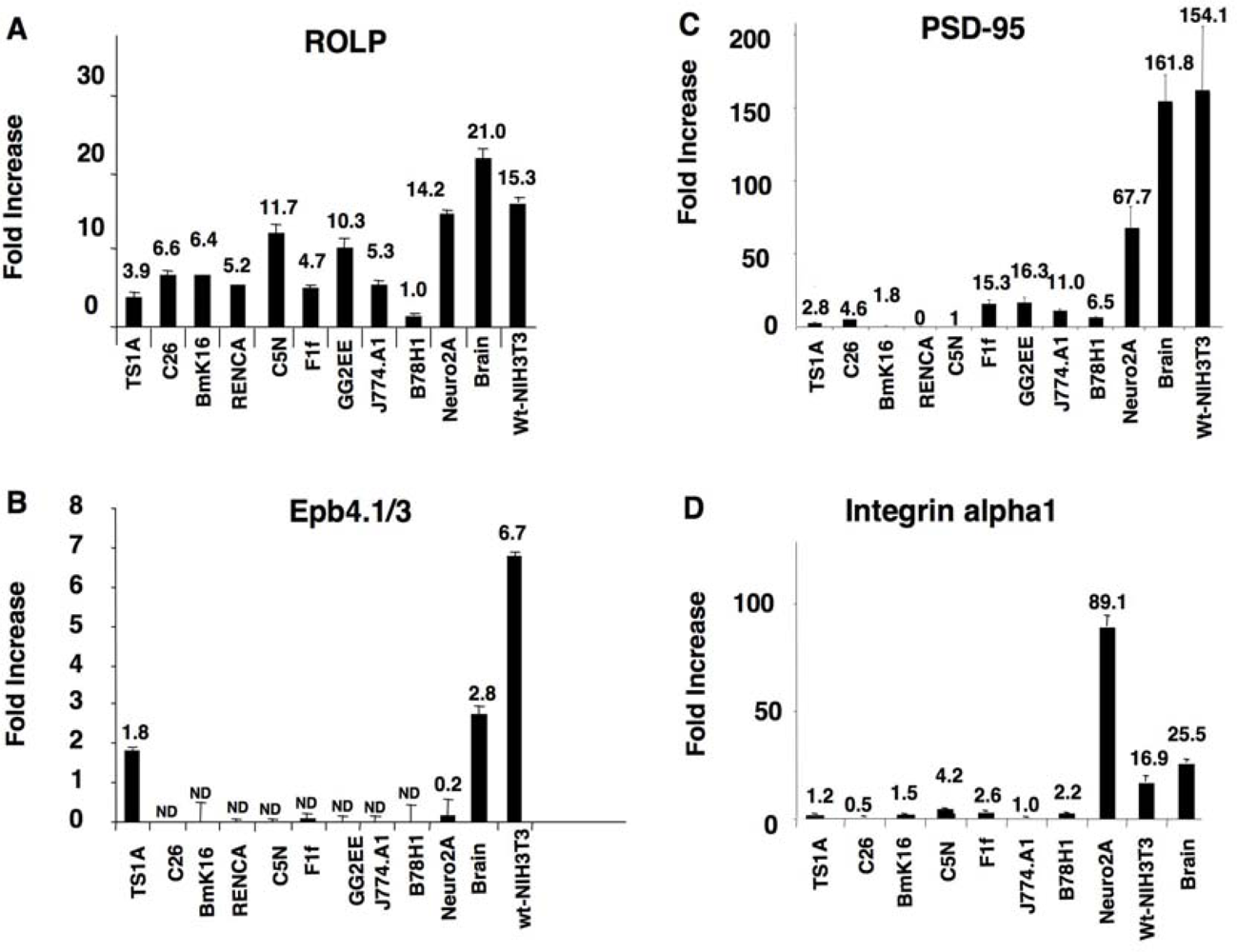
kp1 (NP_035673), Rab3 (NP_848805), Rab5 (NP_080163) and Rab11 (NP_059078) (Figure 2 and Table 1). Among these proteins Integrin-alpha 1 is of particular interest due to its key role in cell adhesion and to the intensity of its signal. Altogether the above results keep in line with the possible interaction of ROLP with PSD95 although its potential functional contact with Integrin-alpha1 and/or one of the other five proteins in a more physiological context cannot be excluded a priori by this approach.3.2. The tissue expression profile of ROLP is compatible with its interaction with Epb4.1/3, PSD95 and Integrin-alpha1
3.3. ROLP silencing prevents cell adhesion
4. Discussion
5. Conclusions
Acknowledgments
- Authors’ ContributionsM.C. has been involved in both the identification of ROLP protein interactors by antibody array and in testing cell adhesion capacity of cells. M.M. determined the expression profile of ROLP, PSD-95 and Epb4.1/3 by Real Time RT-PCR. S.M. characterized the expression of ROLP by immunofluorescence. AP performed ROLP silencing procedures and identified Epb4.1/3 as ROLP partner. I.V. and F.T. partecipated to the preparation of some molecular reagents needed for this project. R.C. provided conceptual suggestions and critically read the manuscript. A.P. conceived the project, designed the experiments and wrote the paper.
References
- Pagano, A; Tonachini, L; Monticone, M; Tortelli, F; Castelnuovo, M; Randazzo, N; Tavella, S; Di Marco, E; Cancedda, R; Castagnola, P. Proliferative arrest and activation of apoptosis related genes in Rolly Protein-silenced cells. Gene 2006, 382, 79–87. [Google Scholar]
- Lue, RA; Marfatia, SM; Branton, D; Chishti, AH. Cloning and characterization of hdlg: the human homologue of the Drosophila discs large tumor suppressor binds to protein 4.1. Proc. Natl. Acad. Sci. USA 1994, 91, 9818–9822. [Google Scholar]
- Marfatia, SM; Morais Cabral, JH; Lin, L; Hough, C; Bryant, PJ; Stolz, L; Chishti, AH. Modular organization of the PDZ domains in the human discs-large protein suggests a mechanism for coupling PDZ domain-binding proteins to ATP and the membrane cytoskeleton. J. Cell Biol 1996, 135, 753–766. [Google Scholar]
- Parra, M; Gascard, P; Walensky, LD; Gimm, JA; Blackshaw, S; Chan, N; Takakuwa, Y; Berger, T; Lee, G; Chasis, JA; et al. Molecular and functional characterization of protein 4.1B, a novel member of the protein 4.1 family with high level, focal expression in brain. J. Biol. Chem 2000, 275, 3247–3255. [Google Scholar]
- Hoover, KB; Bryant, PJ. The genetics of the protein 4.1 family: organizers of the membrane and cytoskeleton. Curr. Opin. Cell Biol 2000, 12, 229–234. [Google Scholar]
- Woods, DF; Hough, C; Peel, D; Callaini, G; Bryant, PJ. Dlg protein is required for junction structure, cell polarity, and proliferation control in Drosophila epithelia. J. Cell Biol 1996, 134, 1469–1482. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar]
- Luque, CM; Perez-Ferreiro, CM; Perez-Gonzalez, A; Englmeier, L; Koffa, MD; Correas, I. An alternative domain containing a leucine-rich sequence regulates nuclear cytoplasmic localization of protein 4.1R. J. Biol. Chem 2003, 278, 2686–2691. [Google Scholar]
- Sun, CX; Robb, VA; Gutmann, DH. Protein 4.1 tumor suppressors: getting a FERM grip on growth regulation. J. Cell Sci 2002, 115, 3991–4000. [Google Scholar]
- Tran, YK; Bogler, O; Gorse, KM; Wieland, I; Green, MR; Newsham, IF. A novel member of the NF2/ERM/4.1 superfamily with growth suppressing properties in lung cancer. Cancer Res 1999, 59, 35–43. [Google Scholar]
- Leto, TL; Pratt, BM; Madri, JA. Mechanisms of cytoskeletal regulation: modulation of aortic endothelial cell protein band 4.1 by the extracellular matrix. J. Cell Physiol 1986, 127, 423–431. [Google Scholar]
- Correas, I. Characterization of isoforms of protein 4.1 present in the nucleus. Biochem. J 1991, 279, 581–585. [Google Scholar]
- De Carcer, G; Lallena, MJ; Correas, I. Protein 4.1 is a component of the nuclear matrix of mammalian cells. Biochem. J 1995, 312, 871–877. [Google Scholar]
- Krauss, SW; Chasis, JA; Rogers, C; Mohandas, N; Krockmalnic, G; Penman, S. Structural protein 4.1 is located in mammalian centrosomes. Proc. Natl. Acad. Sci. USA 1997, 94, 7297–7302. [Google Scholar]
- Charboneau, AL; Singh, V; Yu, T; Newsham, IF. Suppression of growth and increased cellular attachment after expression of DAL-1 in MCF-7 breast cancer cells. Int. J. Cancer 2002, 100, 181–188. [Google Scholar]
- Feller, SM; Ren, R; Hanafusa, H; Baltimore, D. SH2 and SH3 domains as molecular adhesives: the interactions of Crk and Abl. Trends Biochem. Sci 1994, 19, 453–458. [Google Scholar]
- Pawson, T. Protein modules and signalling networks. Nature 1995, 373, 573–580. [Google Scholar]
- McClay, DR; Wessel, GM; Marchase, RB. Intercellular recognition: quantitation of initial binding events. Proc. Natl. Acad. Sci. USA 1981, 78, 4975–4979. [Google Scholar]
- Cancedda, R; Castagnola, P; Cancedda, FD; Dozin, B; Quarto, R. Developmental control of chondrogenesis and osteogenesis. Int. J. Dev. Biol 2000, 44, 707–714. [Google Scholar]
- Yageta, M; Kuramochi, M; Masuda, M; Fukami, T; Fukuhara, H; Maruyama, T; Shibuya, M; Murakami, Y. Direct association of TSLC1 and DAL-1, two distinct tumor suppressor proteins in lung cancer. Cancer Res 2002, 62, 5129–5133. [Google Scholar]
- Gutmann, DH; Donahoe, J; Perry, A; Lemke, N; Gorse, K; Kittiniyom, K; Rempel, SA; Gutierrez, JA; Newsham, IF. Loss of DAL-1, a protein 4.1-related tumor suppressor, is an important early event in the pathogenesis of meningiomas. Hum. Mol. Genet 2000, 9, 1495–1500. [Google Scholar]

© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Castelnuovo, M.; Monticone, M.; Massone, S.; Vassallo, I.; Tortelli, F.; Cancedda, R.; Pagano, A. Rolly Protein (ROLP)-Epb4.1/3: A Potential Protein-Protein Interaction Relevant for the Maintenance of Cell Adhesion. Int. J. Mol. Sci. 2009, 10, 2054-2065. https://doi.org/10.3390/ijms10052054
Castelnuovo M, Monticone M, Massone S, Vassallo I, Tortelli F, Cancedda R, Pagano A. Rolly Protein (ROLP)-Epb4.1/3: A Potential Protein-Protein Interaction Relevant for the Maintenance of Cell Adhesion. International Journal of Molecular Sciences. 2009; 10(5):2054-2065. https://doi.org/10.3390/ijms10052054
Chicago/Turabian StyleCastelnuovo, Manuele, Massimiliano Monticone, Sara Massone, Irene Vassallo, Federico Tortelli, Ranieri Cancedda, and Aldo Pagano. 2009. "Rolly Protein (ROLP)-Epb4.1/3: A Potential Protein-Protein Interaction Relevant for the Maintenance of Cell Adhesion" International Journal of Molecular Sciences 10, no. 5: 2054-2065. https://doi.org/10.3390/ijms10052054