Structures, Activities and Drug-Likeness of Anti-Infective Xanthone Derivatives Isolated from the Marine Environment: A Review


 ,
,  , and
, and
Abstract
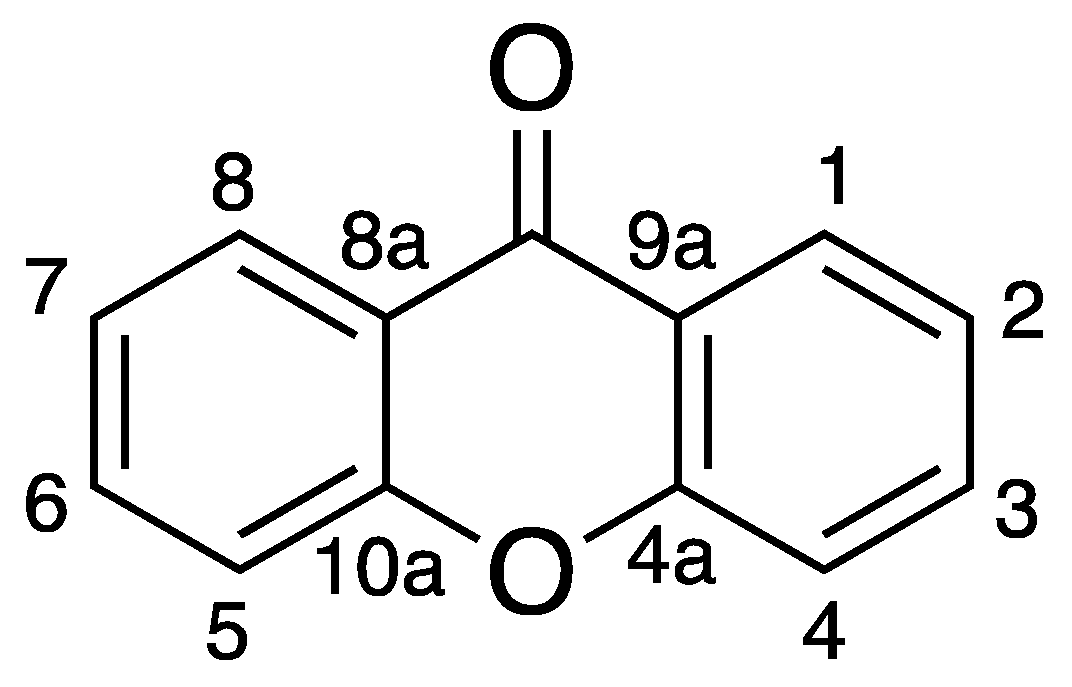
:1. Introduction
2. Anti-Infective Xanthones Isolated from Marine Environment
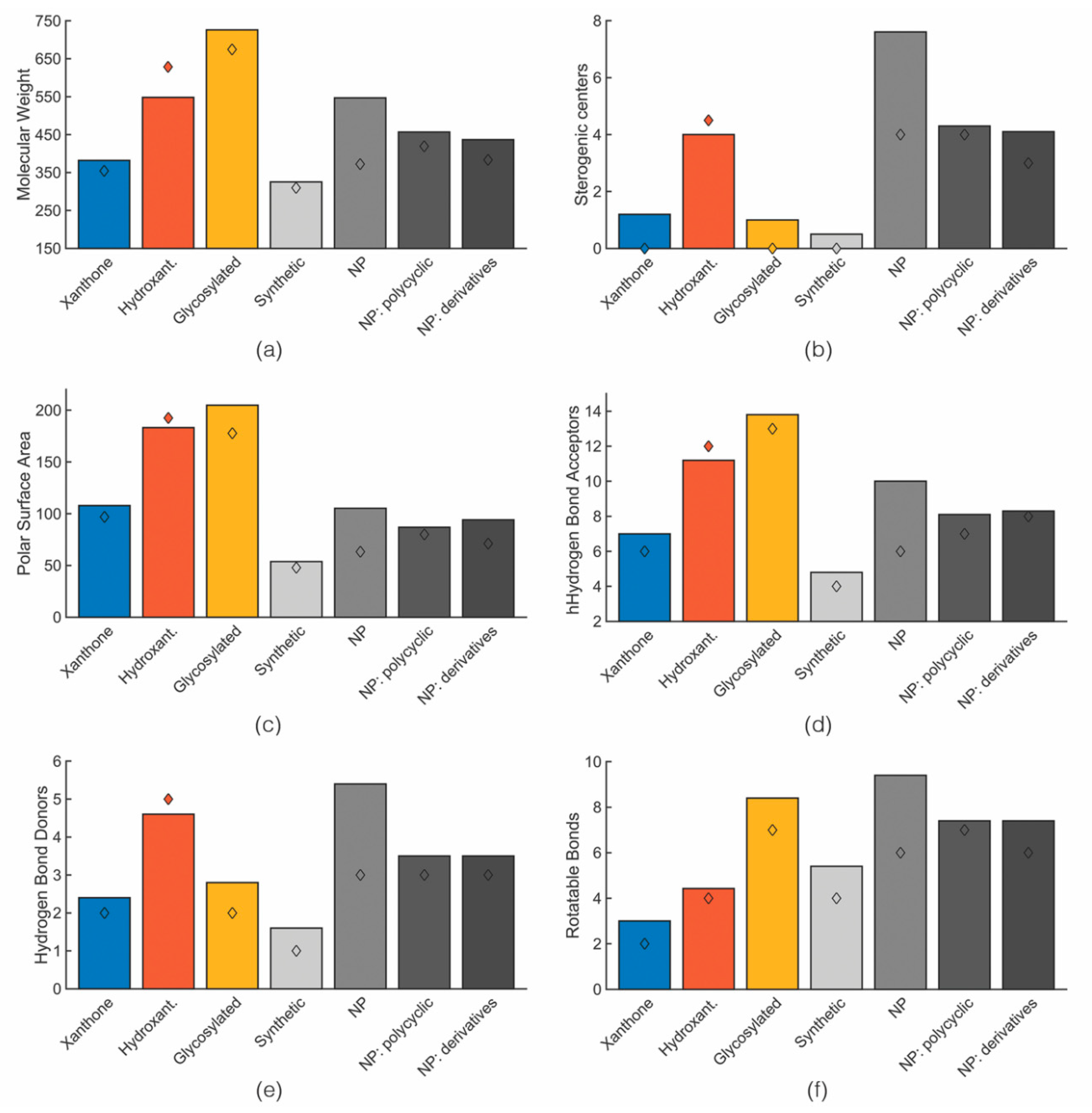
3. Comparison of Drug-Likeness of Marine Xanthone Derivatives with Marketed Drugs
- molecular weight (MW),
- number of stereogenic centers,
- number of hydrogen bond acceptors (HBA) and donors (HBD), described as the electrostatic bond between a hydrogen and a lone pair of electrons,
- number of rotatable bonds (RB),
- number of rings,
- fraction of sp3 carbons (Fsp3) defined as the ratio of sp3 hybridized carbons over the total number of carbons [33]
- fraction of aromatic heavy atoms (FAr), defined as the number of aromatic heavy atoms divided by the total number of heavy atoms.
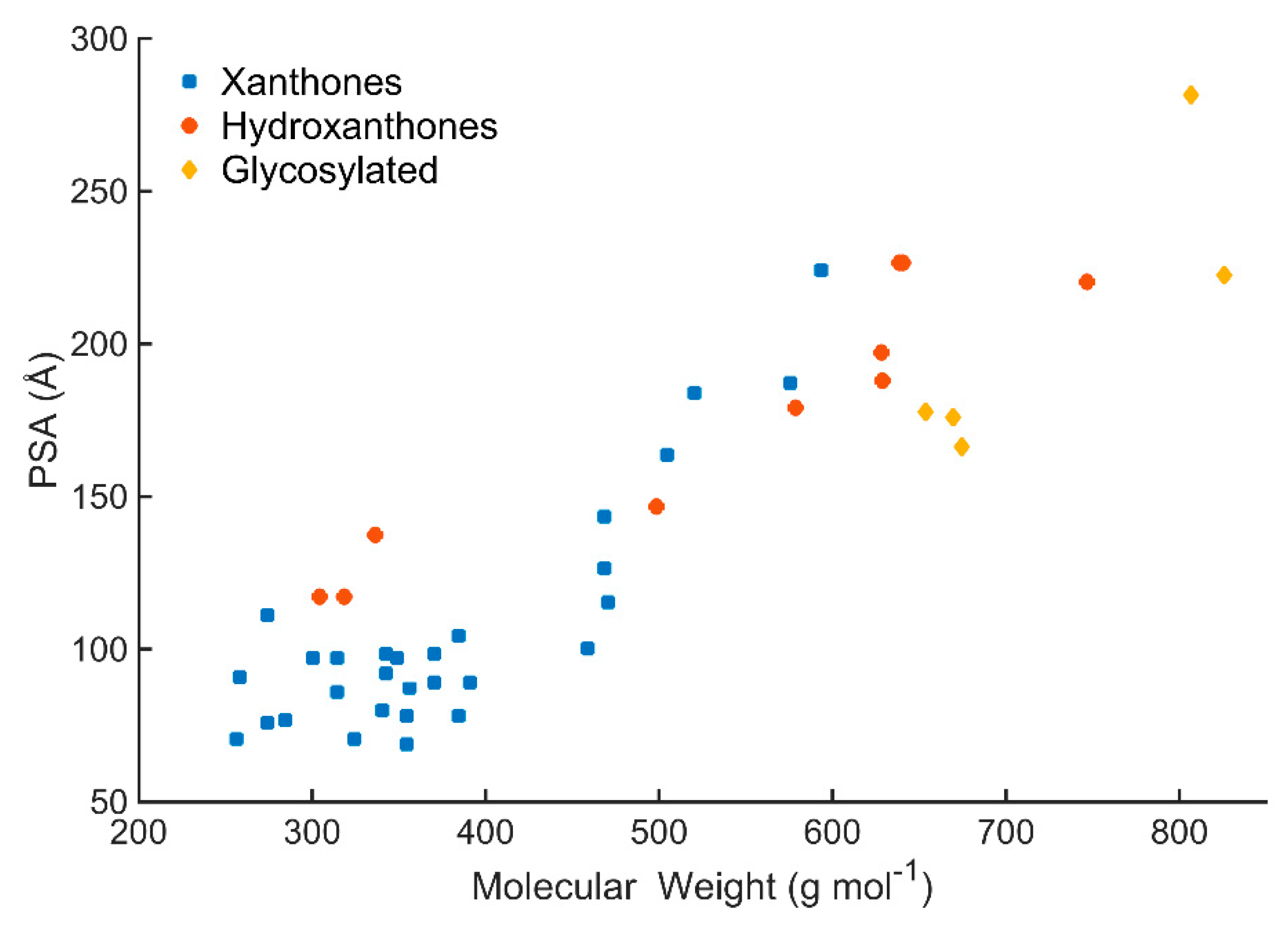
3.1. Size: Molecular Weight
3.2. Chirality: Number of Stereogenic Centers
3.3. Polarity: PSA and HBD/HBA
3.4. Molecular Flexibility: Rotatable Bonds and Aromatic Character
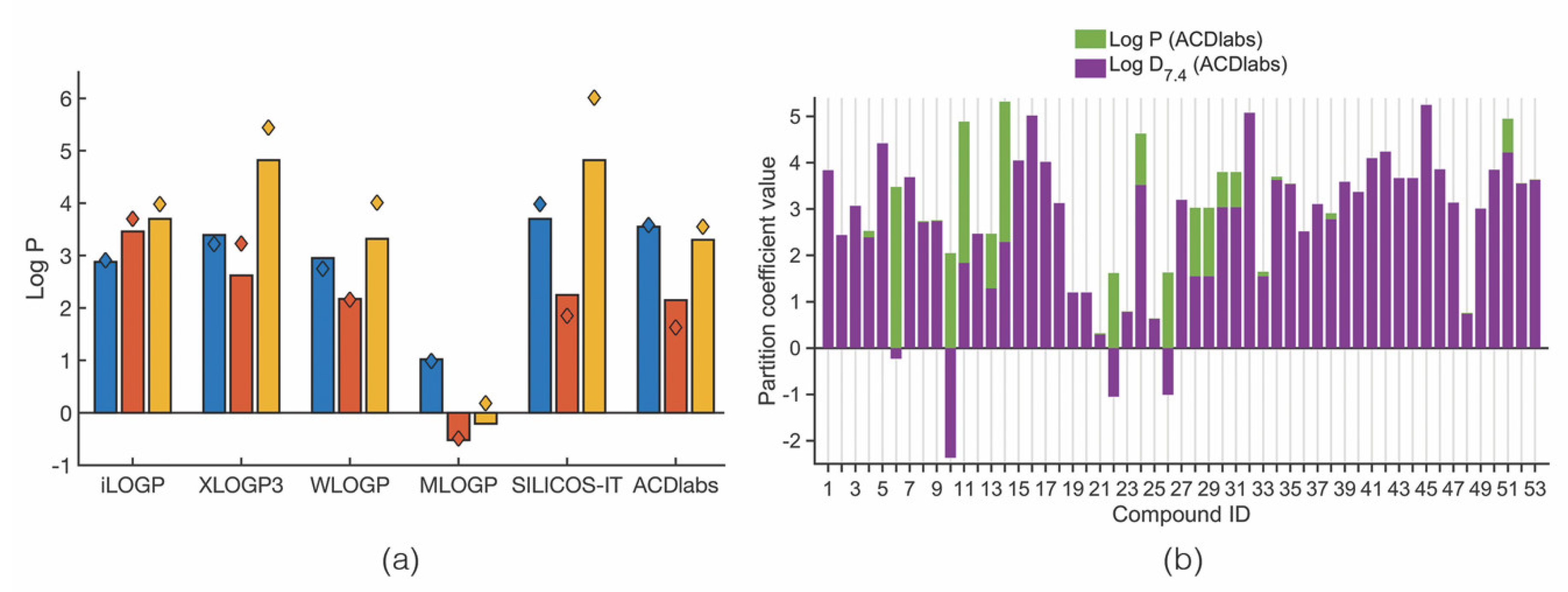
3.5. Lipophilicity: Log P
3.6. Solubility: Log S
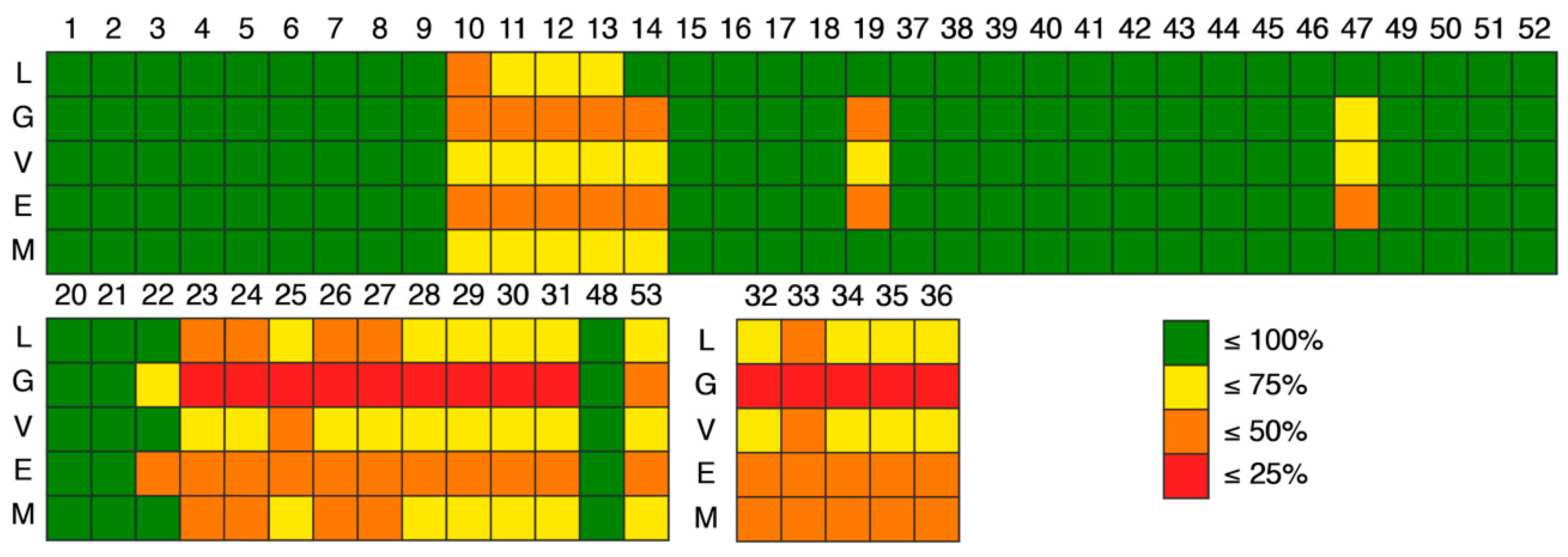
4. Compliance of Marine Xanthone Derivatives with the Rules of Drug-Likeness
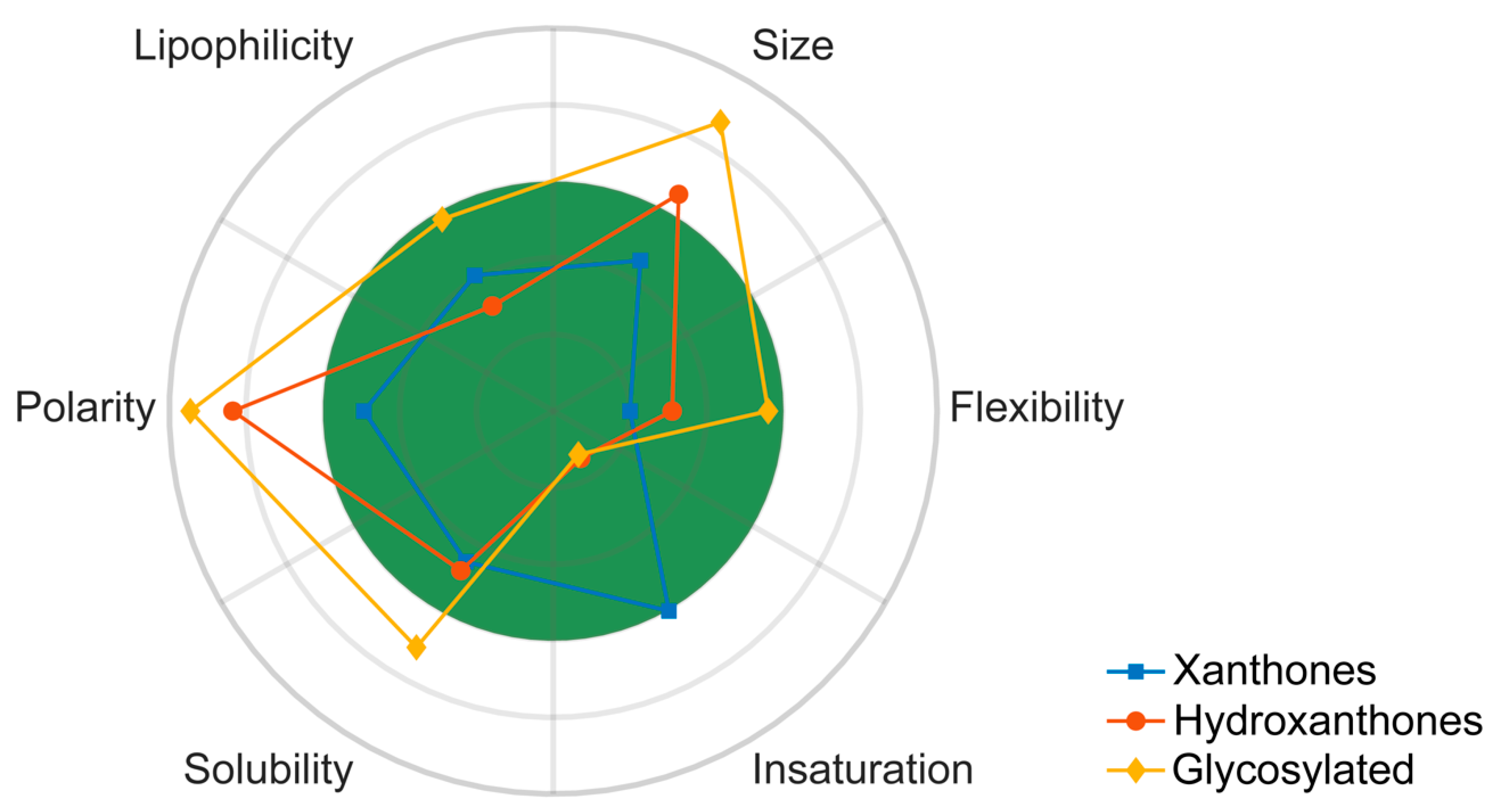
5. Trends on the PK Behavior of Marine Xanthone Derivatives
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hayashi, M.A.; Bizerra, F.C.; Da Silva, P.I.J. Antimicrobial compounds from natural sources. Front. Microbiol. 2013, 4, 195. [Google Scholar] [CrossRef]
- Furusawa, C.; Horinouchi, T.; Maeda, T. Toward prediction and control of antibiotic-resistance evolution. Curr. Opin. Biotechnol. 2018, 54, 45–49. [Google Scholar] [CrossRef]
- Shen, B. A New Golden Age of Natural Products Drug Discovery. Cell 2015, 163, 1297–1300. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef]
- Snelgrove, P.V.R. An Ocean of Discovery: Biodiversity Beyond the Census of Marine Life. Planta Med. 2016, 82, 790–799. [Google Scholar] [CrossRef] [Green Version]
- Lindequist, U. Marine-Derived Pharmaceuticals - Challenges and Opportunities. Biomol. Ther. 2016, 24, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Chen, J.; Hu, G.; Yu, J.; Zhu, X.; Lin, Y.; Chen, S.; Yuan, J. Statistical Research on the Bioactivity of New Marine Natural Products Discovered during the 28 Years from 1985 to 2012. Mar. Drugs 2015, 13, 202–221. [Google Scholar] [CrossRef] [Green Version]
- Penesyan, A.; Kjelleberg, S.; Egan, S. Development of novel drugs from marine surface associated microorganisms. Mar. Drugs 2010, 8, 438–459. [Google Scholar] [CrossRef]
- Kiuru, P.; D’Auria, M.V.; Muller, C.D.; Tammela, P.; Vuorela, H.; Yli-Kauhaluoma, J. Exploring Marine Resources for Bioactive Compounds. Planta Med. 2014, 80, 1234–1246. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Res. 2018, 35, 8–53. [Google Scholar] [CrossRef] [Green Version]
- Kong, D.X.; Jiang, Y.Y.; Zhang, H.Y. Marine natural products as sources of novel scaffolds: Achievement and concern. Drug Discovery Today 2010, 15, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Torres, V.; Encinar, J.; Herranz-López, M.; Pérez-Sánchez, A.; Galiano, V.; Barrajón-Catalán, E.; Micol, V. An Updated Review on Marine Anticancer Compounds: The Use of Virtual Screening for the Discovery of Small-Molecule Cancer Drugs. Molecules 2017, 22, 1037. [Google Scholar] [CrossRef]
- Rateb, M.E.; Ebel, R. Secondary metabolites of fungi from marine habitats. Nat. Prod. Rep. 2011, 28, 290–344. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.M.M.; Castanheiro, R.A.P.; Kijjoa, A. Xanthones from Marine-Derived Microorganisms: Isolation, Structure Elucidation and Biological Activities. Encycl. Anal. Chem. 2014, 1–21. [Google Scholar] [CrossRef]
- Xu, L.; Meng, W.; Cao, C.; Wang, J.; Shan, W.; Wang, Q. Antibacterial and antifungal compounds from marine fungi. Mar. Drugs 2015, 13, 3479–3513. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, C.M.G.; Afonso, C.M.M.; Pinto, M.M.M. Routes to Xanthones: An Update on the Synthetic Approaches. Curr. Org. Chem. 2012, 16. [Google Scholar] [CrossRef]
- Pinto, M.M.M.; Sousa, M.E.; Nascimento, M.S. Xanthone derivatives: New insights in biological activities. Curr. Med. Chem. 2005, 12, 2517–2538. [Google Scholar] [CrossRef] [PubMed]
- Resende, D.I.S.P.; Pereira-Terra, P.; Inácio, Â.S.; Costa, P.M.; Pinto, E.; Sousa, M.E.; Pinto, M.M.M. Lichen Xanthones as Models for New Antifungal Agents. Molecules 2018, 23, 2617. [Google Scholar] [CrossRef]
- Bedi, P.; Gupta, R.; Pramanik, T. Synthesis and biological properties of pharmaceutically important xanthones and benzoxanthone analogs: A brief review. Asian J. Pharm. Clin. Res. 2018, 11, 12–20. [Google Scholar] [CrossRef]
- Lipinski, C.A. Drug-like properties and the causes of poor solubility and poor permeability. J. Pharmacol. Toxicol. Methods 2000, 44, 235–249. [Google Scholar] [CrossRef]
- Meanwell, N.A. Improving drug candidates by design: A focus on physicochemical properties as a means of improving compound disposition and safety. Chem. Res. Toxicol. 2011, 24, 1420–1456. [Google Scholar] [CrossRef]
- Kerns, E. Pharmaceutical profiling in drug discovery. Drug Discovery Today 2003, 8, 316–323. [Google Scholar] [CrossRef]
- Ladbury, J.E.; Klebe, G.; Freire, E. Adding calorimetric data to decision making in lead discovery: A hot tip. Nat. Rev. Drug Discov. 2010, 9. [Google Scholar] [CrossRef] [PubMed]
- Hann, M.M.; Keseru, G.M. Finding the sweet spot: The role of nature and nurture in medicinal chemistry. Nat. Rev. Drug Discov. 2012, 11, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, D.R.P.; Soares, J.X.; Lopes, D.; Macedo, T.; Yordanova, D.; Jakobtorweihen, S.; Nunes, C.; Reis, S.; Pinto, M.M.M.; Afonso, C.M.M. Accessing lipophilicity of drugs with biomimetic models: A comparative study using liposomes and micelles. Eur. J. Pharm. Sci. 2018, 30, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Bade, R.; Chan, H.F.; Reynisson, J. Characteristics of known drug space. Natural products, their derivatives and synthetic drugs. Eur. J. Med. Chem. 2010, 45, 5646–5652. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.-Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular Properties That Influence the Oral Bioavailability of Drug Candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghose, A.K.; Viswanadhan, V.N.; Wendoloski, J.J. A Knowledge-Based Approach in Designing Combinatorial or Medicinal Chemistry Libraries for Drug Discovery. 1. A Qualitative and Quantitative Characterization of Known Drug Databases. J. Comb. Chem. 1999, 1, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Egan, W.J.; Merz, K.M.; Baldwin, J.J. Prediction of Drug Absorption Using Multivariate Statistics. J. Med. Chem. 2000, 43, 3867–3877. [Google Scholar] [CrossRef]
- Muegge, I.; Heald, S.L.; Brittelli, D. Simple Selection Criteria for Drug-like Chemical Matter. J. Med. Chem. 2001, 44. [Google Scholar] [CrossRef]
- Meanwell, N.A. Improving Drug Design: An Update on Recent Applications of Efficiency Metrics, Strategies for Replacing Problematic Elements, and Compounds in Nontraditional Drug Space. Chem. Res. Toxicol. 2016, 29, 564–616. [Google Scholar] [CrossRef]
- Daina, A.; Zoete, V. A BOILED-Egg To Predict Gastrointestinal Absorption and Brain Penetration of Small Molecules. Chemmedchem 2016, 11, 1117–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.S.; Brandão, P.; Fernandes, C.S.G.; Silva, M.R.P.C.; Sousa, M.E.D.; Pinto, M.M.M. Drug-like Properties and ADME of Xanthone Derivatives: The Antechamber of Clinical Trials. Curr. Med. Chem. 2016, 23, 3654–3686. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, C.M.G.; Afonso, C.M.M.; Soares, J.X.; Reis, S.; Sousa, D.; Lima, R.T.; Vasconcelos, M.H.; Pedro, M.; Barbosa, J.; Gales, L.; et al. Pyranoxanthones: Synthesis, growth inhibitory activity on human tumor cell lines and determination of their lipophilicity in two membrane models. Eur. J. Med. Chem. 2013, 69, 798–816. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Soares, J.X.; Cravo, S.; Tiritan, M.E.; Reis, S.; Afonso, C.; Fernandes, C.; Pinto, M.M.M. Lipophilicity assessement in drug discovery: Experimental and theoretical methods applied to xanthone derivatives. J. Chromatogr. B 2018, 1072, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Dharmaratne, H.R.; Wijesinghe, W.M.N.M.; Thevanasem, V. Antimicrobial activity of xanthones from Calophyllum species, against methicillin-resistant Staphylococcus aureus (MRSA). J. Ethnopharmacol. 1999, 66, 339–342. [Google Scholar] [CrossRef]
- Komguem, J.; Meli, A.; Manfouo, R.; Lontsi, D.; Ngounou, F.; Kuete, V.; Kamdem, H.W.; Tane, P.; Ngadjui, B.T.; Sondengam, B.L. Xanthones from Garcinia smeathmannii (Oliver) and their antimicrobial activity. Phytochemistry 2005, 66, 1713–1717. [Google Scholar] [CrossRef]
- Nguemeving, J.R.; Azebaze, A.G.B.; Kuete, V.; Carly, N.N.E.; Beng, V.P.; Meyer, M.; Blond, A.; Bodo, B.; Nkengfack, A.E. Laurentixanthones A and B, antimicrobial xanthones from Vismia laurentii. Phytochemistry 2006, 67, 1341–1346. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; El-Barbary, M.A.; El-Ghorab, D.M.H.; Bohlin, L.; Borg-Karlson, A.-K.; Goransson, U.; Verpoorte, R. Recent insights into the biosynthesis and biological activities of natural xanthones. Curr. Med. Chem. 2010, 17, 854–901. [Google Scholar] [CrossRef]
- Trisuwan, K.; Boonyaketgoson, S.; Rukachaisirikul, V.; Phongpaichit, S. Oxygenated xanthones and biflavanoids from the twigs of Garcinia xanthochymus. Tetrahedron Lett. 2014, 55, 3600–3602. [Google Scholar] [CrossRef]
- Ruan, J.; Zheng, C.; Liu, Y.; Qu, L.; Yu, H.; Han, L.; Zhang, Y.; Wang, T. Chemical and Biological Research on Herbal Medicines Rich in Xanthones. Molecules 2017, 22, 1698. [Google Scholar] [CrossRef]
- Song, F.; Ren, B.; Chen, C.; Yu, K.; Liu, X.; Zhang, Y.; Yang, N.; He, H.; Liu, X.; Dai, H.; et al. Three new sterigmatocystin analogues from marine-derived fungus Aspergillus versicolor MF359. Appl. Microbiol. Biotechnol. 2014, 98, 3753–3758. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Lateff, A.; Klemke, C.; Konig, G.M.; Wright, A.D. Two new xanthone derivatives from the algicolous marine fungus Wardomyces anomalus. J. Nat. Prod. 2003, 66, 706–708. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Cai, X.L.; Yang, H.; Xia, X.K.; Guo, Z.Y.; Yuan, J.; Li, M.F.; She, Z.G.; Lin, Y.C. The bioactive metabolites of the mangrove endophytic fungus Talaromyces sp. ZH-154 isolated from Kandelia candel (L.) Druce. Planta Med. 2010, 76, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Kossuga, M.H.; Romminger, S.; Xavier, C.; Milanetto, M.C.; Valle, M.Z.D.; Pimenta, E.F.; Morais, R.P.; Carvalho, E.d.; Mizuno, C.M.; Coradello, L.F.C. Evaluating methods for the isolation of marine-derived fungal strains and production of bioactive secondary metabolites. Rev. Bras. Farmacogn. 2012, 22, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Lin, Y.C.; She, Z.G.; Du, D.S.; Chan, W.L.; Zheng, Z.H. Paeciloxanthone, a new cytotoxic xanthone from the marine mangrove fungus Paecilomyces sp. (Tree1-7). J. Asian Nat. Prod. Res. 2008, 10, 133–137. [Google Scholar] [CrossRef]
- Sun, R.R.; Miao, F.P.; Zhang, J.; Wang, G.; Yin, X.L.; Ji, N.Y. Three new xanthone derivatives from an algicolous isolate of Aspergillus wentii. Magn. Reson. Chem. 2013, 51, 65–68. [Google Scholar] [CrossRef]
- Li, H.L.; Li, X.M.; Liu, H.; Meng, L.H.; Wang, B.G. Two new diphenylketones and a new xanthone from talaromyces islandicus EN-501, an endophytic fungus derived from the marine red alga Laurencia okamurai. Mar. Drugs 2016, 14. [Google Scholar] [CrossRef]
- Moon, K.; Chung, B.; Shin, Y.; Rheingold, A.L.; Moore, C.E.; Park, S.J.; Park, S.; Lee, S.K.; Oh, K.B.; Shin, J.; et al. Pentacyclic antibiotics from a tidal mud flat-derived actinomycete. J. Nat. Prod. 2015, 78, 524–529. [Google Scholar] [CrossRef]
- Liu, L.L.; Xu, Y.; Han, Z.; Li, Y.X.; Lu, L.; Lai, P.Y.; Zhong, J.L.; Guo, X.R.; Zhang, X.X.; Qian, P.Y. Four New Antibacterial Xanthones from the Marine-Derived Actinomycetes Streptomyces caelestis. Mar. Drugs 2012, 10, 2571–2583. [Google Scholar] [CrossRef] [Green Version]
- Barrero, A.F.; Oltra, J.E.; Herrador, M.M.; Cabrera, E.; Sanchez, J.F.; Quílez, J.F.; Rojas, F.J.; Reyes, J.F. Gibepyrones: α-pyrones from Gibberella fujikuroi. Tetrahedron 1993, 49, 141–150. [Google Scholar] [CrossRef]
- Malmstrom, J.; Christophersen, C.; Barrero, A.F.; Oltra, J.E.; Justicia, J.; Rosales, A. Bioactive metabolites from a marine-derived strain of the fungus Emericella variecolor. J. Nat. Prod. 2002, 65, 364–367. [Google Scholar] [CrossRef] [PubMed]
- Fredimoses, M.; Zhou, X.; Lin, X.; Tian, X.; Ai, W.; Wang, J.; Liao, S.; Liu, J.; Yang, B.; Yang, X.; et al. New Prenylxanthones from the Deep-Sea Derived Fungus Emericella sp. SCSIO 05240. Mar. Drugs 2014, 12, 3190–3202. [Google Scholar] [CrossRef] [Green Version]
- Erbert, C.; Lopes, A.A.; Yokoya, N.S.; Furtado, N.A.; Conti, R.; Pupo, M.T.; Lopes, J.L.C.; Debonsi, H.M. Antibacterial compound from the endophytic fungus Phomopsis longicolla isolated from the tropical red seaweed Bostrychia radicans. Bot. Mar. 2012, 55, 435–440. [Google Scholar] [CrossRef]
- Yao, Q.; Wang, J.; Zhang, X.; Nong, X.; Xu, X.; Qi, S. Cytotoxic Polyketides from the Deep-Sea-Derived Fungus Engyodontium album DFFSCS021. Mar. Drugs 2014, 12, 5902–5915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ding, W.; Wang, R.; Du, Y.; Liu, H.; Kong, X.; Li, C. Identification and Bioactivity of Compounds from the Mangrove Endophytic Fungus Alternaria sp. Mar. Drugs 2015, 13, 4492–4504. [Google Scholar] [CrossRef] [Green Version]
- Fredimoses, M.; Zhou, X.; Ai, W.; Tian, X.; Yang, B.; Lin, X.; Liu, J.; Liu, Y. Emerixanthone E, a new xanthone derivative from deep sea fungus Emericella sp SCSIO 05240. Nat. Prod. Res. 2018, 17, 1–7. [Google Scholar] [CrossRef]
- Bao, J.; Sun, Y.L.; Zhang, X.Y.; Han, Z.; Gao, H.C.; He, F.; Qian, P.Y.; Qi, S.H. Antifouling and antibacterial polyketides from marine gorgonian coral-associated fungus Penicillium sp. SCSGAF 0023. J. Antibiot. (Tokyo) 2013, 66, 219–223. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, X.P.; Ju, Z.R.; Liao, X.J.; Huang, X.J.; Zhang, C.; Zhao, B.X.; Xu, S.H. Aspergchromones A and B, two new polyketides from the marine sponge-associated fungus Aspergillus sp. SCSIO XWS03F03. J. Asian Nat. Prod. Res. 2017, 19, 684–690. [Google Scholar] [CrossRef]
- Zhang, W.; Krohn, K.; Florke, U.; Pescitelli, G.; Di Bari, L.; Antus, S.; Kurtan, T.; Rheinheimer, J.; Draeger, S.; Schulz, B. New mono- and dimeric members of the secalonic acid family: Blennolides A-G isolated from the fungus Blennoria sp. Chemistry 2008, 14, 4913–4923. [Google Scholar] [CrossRef]
- Eltamany, E.E.; Abdelmohsen, U.R.; Ibrahim, A.K.; Hassanean, H.A.; Hentschel, U.; Ahmed, S.A. New antibacterial xanthone from the marine sponge-derived Micrococcus sp. EG45. Bioorg. Med. Chem. Lett. 2014, 24, 4939–4942. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Wiese, J.; Wenzel-Storjohann, A.; Malien, S.; Schmaljohann, R.; Imhoff, J.F. Engyodontochones, Antibiotic Polyketides from the Marine Fungus Engyodontium album Strain LF069. Chem. Eur. J. 2016, 22, 7452–7462. [Google Scholar] [CrossRef] [PubMed]
- Malet-Cascon, L.; Romero, F.; Espliego-Vazquez, F.; Gravalos, D.; Fernandez-Puentes, J.L. IB-00208, a new cytotoxic polycyclic xanthone produced by a marine-derived Actinomadura. I. Isolation of the strain, taxonomy and biological activites. J. Antibiot. 2003, 56, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Peoples, A.J.; Zhang, Q.; Millett, W.P.; Rothfeder, M.T.; Pescatore, B.C.; Madden, A.A.; Ling, L.L.; Moore, C.M. Neocitreamicins I and II, Novel Antibiotics with Activity Against Methicillin Resistant Staphylococcus aureus and Vancomycin-Resistant Enterococci. J. Antibiot. (Tokyo) 2008, 61, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Maiese, W.M.; Lechevalier, M.P.; Lechevalier, H.A.; Korshalla, J.; Goodman, J.; Wildey, M.J.; Kuck, N.; Greenstein, M. LL-E19085 alpha, a novel antibiotic from Micromonospora citrea: Taxonomy, fermentation and biological activity. J. Antibiot. (Tokyo) 1989, 42, 846–851. [Google Scholar] [CrossRef]
- Frenz, J.L.; Kohl, A.C.; Kerr, R.G. Marine natural products as therapeutic agents: Part 2. Expert. Opin. Ther. Pat. 2004, 14, 17–33. [Google Scholar] [CrossRef]
- Höller, U.; König, G.M.; Wright, A.D. A New Tyrosine Kinase Inhibitor from a Marine Isolate of Ulocladium botrytis and New Metabolites from the Marine Fungi Asteromyces cruciatus and Varicosporina ramulosa. Eur. J. Org. Chem. 1999, 1999, 2949–2955. [Google Scholar] [CrossRef]
- Marmann, A.; Aly, A.; Lin, W.; Wang, B.; Proksch, P. Co-Cultivation—A Powerful Emerging Tool for Enhancing the Chemical Diversity of Microorganisms. Mar. Drugs 2014, 12, 1043–1065. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhang, J.; Shao, C.; Ding, W.; She, Z.; Lin, Y. A new xanthone derivative from the co-culture broth of two marine fungi (strain No. E33 and K38). Chem. Nat. Compd. 2011, 47, 382–384. [Google Scholar] [CrossRef]
- Shao, C.; Wang, C.; Wei, M.; Gu, Y.; Xia, X.; She, Z.; Lin, Y. Structure elucidation of two new xanthone derivatives from the marine fungus Penicillium sp. (ZZF 32#) from the South China Sea. Magn. Reson. Chem. 2008, 46, 1066–1069. [Google Scholar] [CrossRef]
- Wang, X.; Mao, Z.-G.; Song, B.-B.; Chen, C.-H.; Xiao, W.-W.; Hu, B.; Wang, J.-W.; Jiang, X.-B.; Zhu, Y.-H.; Wang, H.-J. Advances in the Study of the Structures and Bioactivities of Metabolites Isolated from Mangrove-Derived Fungi in the South China Sea. Mar. Drugs 2013, 11, 3601–3616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; He, W.; Huang, X.; Tian, X.; Liao, S.; Yang, B.; Wang, F.; Zhou, X.; Liu, Y. Antifungal New Oxepine-Containing Alkaloids and Xanthones from the Deep-Sea-Derived Fungus Aspergillus versicolor SCSIO 05879. J. Agric. Food. Chem. 2016, 64, 2910–2916. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Rotinsulu, H.; Kaneko, T.; Murakami, K.; Fujiwara, H.; Ukai, K.; Namikoshi, M. A new dibenz[b,e]oxepine derivative, 1-hydroxy-10-methoxy-dibenz[b,e]oxepin-6,11-dione, from a marine-derived fungus, Beauveria bassiana TPU942. Mar. Drugs 2012, 10, 2691–2697. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Huang, J.; Zhou, X.; Lin, X.; Liu, J.; Liao, S.; Wang, J.; Liu, F.A.; Tao, H.; Liu, Y. The Fungal Metabolites with Potential Antiplasmodial Activity. Curr. Med. Chem. 2018, 25, 3796–3825. [Google Scholar] [CrossRef] [PubMed]
- Pontius, A.; Krick, A.; Kehraus, S.; Brun, R.; Konig, G.M. Antiprotozoal activities of heterocyclic-substituted xanthones from the marine-derived fungus Chaetomium sp. J. Nat. Prod. 2008, 71, 1579–1584. [Google Scholar] [CrossRef] [PubMed]
- Tempone, A.G.; Martins de Oliveira, C.; Berlinck, R.G.S. Current Approaches to Discover Marine Antileishmanial Natural Products. Planta Med. 2011, 77, 572–585. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Yang, J.; Chen, F.; Lin, X.; Chen, C.; Zhou, X.; Liu, S.; Liu, Y. Structurally diverse polyketides from the mangrove-derived fungus diaporthe sp. SCSIO 41011 with their anti-influenza A virus activities. Front. Chem. 2018, 6. [Google Scholar] [CrossRef]
- Tan, S.; Yang, B.; Liu, J.; Xun, T.; Liu, Y.; Zhou, X. Penicillixanthone A, a marine-derived dual-coreceptor antagonist as anti-HIV-1 agent. Nat. Prod. Res. 2017, 1–5. [Google Scholar] [CrossRef]
- Fang, W.; Lin, X.; Zhou, X.; Wan, J.; Lu, X.; Yang, B.; Ai, W.; Lin, J.; Zhang, T.; Tu, Z.; et al. Cytotoxic and antiviral nitrobenzoyl sesquiterpenoids from the marine-derived fungus Aspergillus ochraceus Jcma1F17. MedChemComm 2014, 5, 701–705. [Google Scholar] [CrossRef]
- Liu, F.A.; Lin, X.; Zhou, X.A.-O.; Chen, M.; Huang, X.; Yang, B.; Tao, H. Xanthones and Quinolones Derivatives Produced by the Deep-Sea-Derived Fungus Penicillium sp. SCSIO Ind16F01. Mar. Drugs 2017, 22. [Google Scholar] [CrossRef]
- SIB. SwissADME. Available online: http://www.swissadme.ch (accessed on 19 October 2018).
- ACD/I-Labs. ACD/I-Lab Prediction Modules. Available online: https://ilab.acdlabs.com/iLab2/ (accessed on 14 September 2018).
- Camp, D.; Garavelas, A.; Campitelli, M. Analysis of Physicochemical Properties for Drugs of Natural Origin. J. Nat. Prod. 2015, 78, 1370–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feher, M.; Schmidt, J.M. Property Distributions: Differences between Drugs, Natural Products, and Molecules from Combinatorial Chemistry. J. Chem. Inf. Comput. Sci. 2003, 43, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Caron, G.; Ermondi, G. Molecular descriptors for polarity: The need for going beyond polar surface area. Future Medicinal Chemistry 2016, 8, 2013–2016. [Google Scholar] [CrossRef] [PubMed]
- Freeman-Cook, K.D.; Hoffman, R.L.; Johnson, T.W. Lipophilic efficiency: The most important efficiency metric in medicinal chemistry. Future Medicinal Chemistry 2013, 5, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Kerns, E.H.; Carter, G.T. Drug-Like Property Concepts in Pharmaceutical Design. Curr. Pharm. Des. 2009, 15, 2184–2194. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Name | Activity | Method | Source | Ref. |
|---|---|---|---|---|---|
| antibacterial activity | |||||
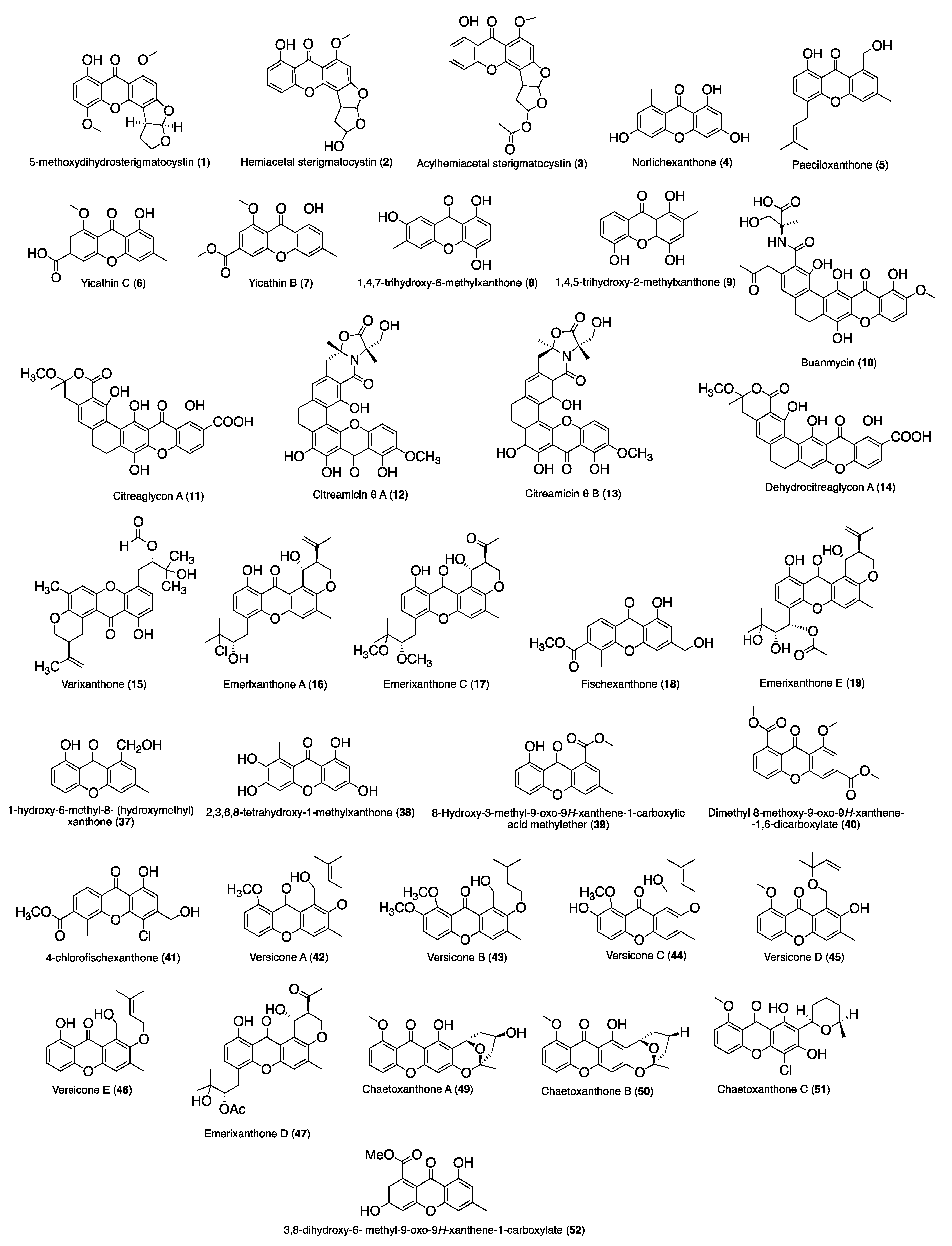
| 1 | 5-Methoxydihydrosterigmatocystin | S. aureus (ATCC 6538) (MIC, 12.5 μg/mL), B. subtilis (ATCC 6633) (MIC, 3.125 μg/mL), MRSA (MIC, >100 μg/mL), P. aeruginosa (ATCC 15692) (MIC, >100 μg/mL) | Antimicrobial Susceptibility Testing Standards outlined | Aspergillus versicolor MF359 isolated from a marine sponge sample of Hymeniacidon perleve collected from the Bohai Sea | [43] |
| 2 | Hemiacetal sterigmatocystin | S. aureus (ATCC 6538) (MIC, >100 μg/mL), B. subtilis (ATCC 6633) (MIC, >100 μg/mL), MRSA (MIC, >100 μg/mL), P. aeruginosa (ATCC 15692) (MIC, >100 μg/mL) | |||
| 3 | Acylhemiacetal sterigmatocystin | S. aureus (ATCC 6538) (MIC, >100 μg/mL), B. subtilis (ATCC 6633) (MIC, >100 μg/mL), MRSA (MIC, >100 μg/mL), P. aeruginosa (ATCC 15692) (MIC, >100 μg/mL) | |||
| 4 | Norlichexanthone (3,6,8-trihydroxy-1-methylxanthone or 1,3,6- trihydroxy-8-methyl-9H-xanthen-9-one) | S. aureus (ATCC 27154) (MIC, 12.5 µg/mL), S. ventriculi (ATCC 29068) (MIC, 25.0 µg/mL), P. aeruginosa (ATCC 25668) (MIC, 25.0 µg/mL) | Microdilution assay | P. raistrikii obtained from the sponge Axinella cf. corrugate or the mangrove endophytic fungus Talaromyces sp. ZH-154 | [44,45,46] |
| B. agisterium (inhibition zone 1 mm), B. megaterium (inhibition zone 1 mm) | Diffusion assay | Wardomyces anomalu isolated from the green alga Enteromorpha sp. (Ulvaceae) collected in the Baltic Sea | |||
| 5 | Paeciloxanthone | E. coli (inhibitory zones of 12 mm) | Standard disk assay | Paecilomyces sp. was isolated from an estuarine mangrove from the Taiwan Strait | [47] |
| 6 | Yicathin C | E. coli (inhibition diameter 12.0 mm), S. aureus (inhibition diameter 7.5 mm) | Standard agar diffusion assay | Aspergillus wentii (red alga Gymnogongrus flabelliformis) collected from the coast of Pingtan Island, China | [48] |
| 7 | Yicathin B | E. coli (inhibition diameter 9 mm) | |||
| 8 | 1,4,7-Trihydroxy-6-methylxanthone | E. coli (MIC, 32 μg/mL), P. aeruginosa (MIC, 32 μg/mL), S. aureus (MIC, >64 μg/mL), V. alginolyticus (MIC, 32 μg/mL), V. harveyi (MIC, 32 μg/mL), V. parahaemolyticus (MIC, 32 μg/mL) | Microplate assay | Talaromyces islandicus EN-501, an endophytic fungus obtained from the fresh collected marine red alga Laurencia okamurai | [49] |
| 9 | 1,4,5-Trihydroxy-2-methylxanthone | E. coli (MIC, 4 μg/mL), P. aeruginosa (MIC, 4 μg/mL), S. aureus (MIC, 8 μg/mL), V. alginolyticus (MIC, 4 μg/mL), V. harveyi (MIC, 8 μg/mL), V. parahaemolyticus (MIC, 4 μg/mL) | |||
| 10 | Buanmycin | S. aureus (MIC, 10.5 μM), B. subtilis (MIC, 0.7 μM), K. rhizophila (MIC, 10.5 μM), S. enterica (MIC, 0.7 μM), P. hauseri (MIC, 21.1 μM), S. aureus sortase A (IC50: 43.2 μM) | Microdilution assay | Streptomyces strain from a tidal mudflat in Buan, Republic of Korea. | [50] |
| 11 | Citreaglycon A | S. haemolyticus (MIC, 8.0 μg/mL), S. aureus UST950701-005 (MIC, 16 μg/mL), B. subtillis 769 (MIC, 8.0 μg/mL), S. aureus ATCC43300 (MIC, 8.0 μg/mL) | Microdilution assay | Streptomyces caelestis from Red Sea | [51] |
| 12 | Citreamicin θ A | S. haemolyticus (MIC, 0.5 μg/mL), S. aureus UST950701-005 (MIC, 1.0 μg/mL), B. subtillis 769 (MIC, 0.25 μg/mL), S. aureus ATCC43300 (MIC, 0.25 μg/mL) | |||
| 13 | Citreamicin θ B | S. haemolyticus UST950701-004 (MIC, 0.5 μg/mL), S. aureus UST950701-005 (MIC, 1.0 μg/mL), B. subtillis 769 (MIC, 0.25 μg/mL), S. aureus ATCC43300 (MIC, 0.25 μg/mL) | |||
| 14 | Dehydrocitreaglycon A | S. haemolyticus UST950701-004 (MIC, 8.0 μg/mL), S. aureus UST950701-005 (MIC, 16 μg/mL), B. subtillis 769 (MIC, 8.0 μg/mL) | |||
| 15 | Varixanthone | E. coli (MIC, 12.5 µg/mL), Proteus sp. (MIC, 12.5 µg/mL), B. subtilis (MIC, 12.5 µg/mL), S. aureus (MIC, 12.5 µg/mL), E. faecalis (MIC, 50 µg/mL) | Method [52] | Emericella variecolor was isolated from a sponge (Porifera) collected in the Caribbean Sea | [53] |
| 16 | Emerixanthone A | E. coli (ATCC 29922), K. pneumoniae (ATCC 13883), S. aureus (ATCC 29213), E. faecalis (ATCC 29212), A. bacterbaumannii (ATCC 19606), A. hydrophila (ATCC 7966): Diameters of inhibition zones were all 4–6 mm | Filter paper disc agar diffusion method | Emericella sp. SCSIO 05240 from South China Sea | [54] |
| 17 | Emerixanthone C | E. coli (ATCC 29922), K. pneumoniae (ATCC 13883), S. aureus (ATCC 29213), E. faecalis (ATCC 29212), A. bacterbaumannii (ATCC 19606), A. hydrophila (ATCC 7966): Diameters of inhibition zones were all 4–6 mm | [54,55,56] | ||
| 18 | Fischexanthone | E. coli (MIC, >1265.82 µM), S. aureus (MIC, >1265.82 µM) | Broth tube dilution method | Mangrove endophytic fungus Alternaria sp. R6 collected from the mangrove in Leizhou peninsula, Guangdong Province, China | [57] |
| 19 | Emerixanthone E | E. coli (ATCC 29922), K. pneumoniae (ATCC 13883), S. aureus (ATCC 29213), E. faecalis (ATCC 29212), A. baumannii (ATCC 19606), and A. hydrophila (ATCC 7966): Diameters of the inhibition zones ranged between 9 and 11 mm | Diffusion method | Marine fungus Emericella sp. was isolated from the South China Sea | [58] |
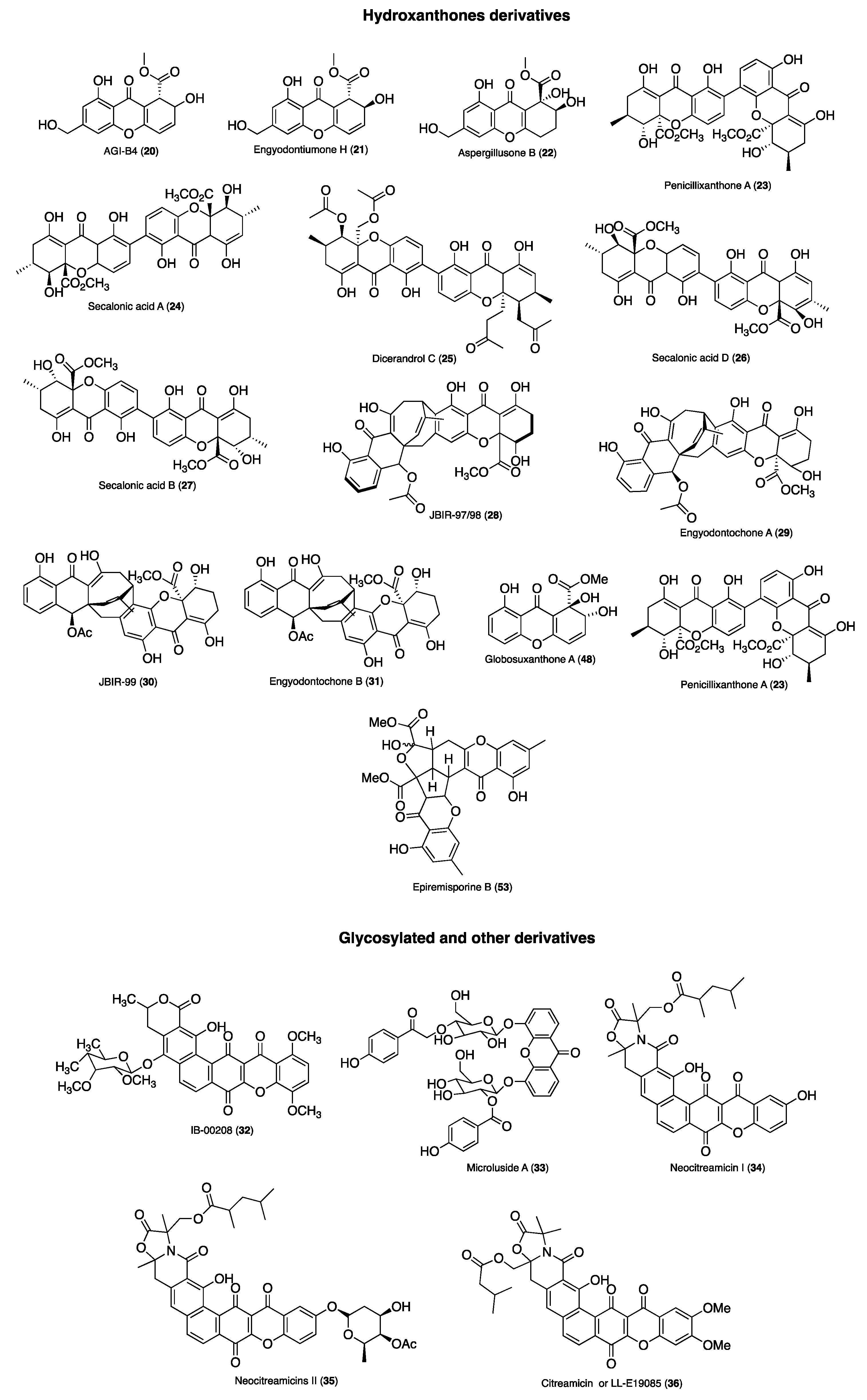
| 20 | AGI-B4 | E. coli (zone of inhibition 13.8 mm), B. subtilis (zone of inhibition 16.5 mm) | Standard disc diffusion assay | Engyodontium album DFFSCS021 from a marine sediment sample collected in the South China Sea | [56] |
| E. coli (MIC, 64 μg/mL), B. subtilis (MIC, 64 μg/mL) | Microbroth dilution method | ||||
| 21 | Engyodontiumone H | E. coli (zone of inhibition 15.8 mm), B. subtilis (zone of inhibition 17.5 mm) | Standard disc diffusion assay | ||
| E. coli (MIC, 64 μg/mL), B. subtilis (MIC, 32 μg/mL) | Microbroth dilution method | ||||
| 22 | Aspergillusone B | E. coli (zone of inhibition 11.0 mm), B. subtilis (zone of inhibition 14.4 mm) | Standard disc diffusion assay | ||
| E. coli (MIC, 64 μg/mL), B. subtilis (MIC, 64 μg/mL) | Microbroth dilution method | ||||
| 23 | Penicillixanthone A | B. subtilis (MIC, 24.4 µg/mL), E. coli JVC1228 (MIC, 24.4 µg/mL), M. luteus UST950701-006 (MIC, 24.4 µg/mL), P. nigrifaciens UST010620-005 (MIC, 97.5 µg/mL) | Standard disc diffusion assay | Penicillium sp. SCSGAF 0023 isolated from South China Sea gorgonian coral Dichotella gemmacea | [59] |
| 24 | Secalonic acid A | S. aureus (ATCC 27154) (MIC 12.5 μg/mL), E. coli (ATCC 25922) (MIC 25 μg/mL), S. ventriculi (ATCC 29068) (MIC 12.5 μg/mL), P. aeruginosa (ATCC 25668) (MIC, 12.5 μg/mL) | - | Talaromyces sp. ZH-154 from the South-China Sea | [45] |
| 25 | Dicerandrol C | S. aureus (ATCC 6538) (MIC, 1/1.33 µg/mL), S. saprophyticus (ATCC 15305) (MIC, 2/2.66 µg/mL) | Microdilution broth method | Endophytic fungus Phomopsis longicolla isolated from the tropical red seaweed Bostrychia radicans from Brazil | [55] |
| 26 | Secalonic acid D | B. subtilis (MIC, 24.4 µg/mL), E. coli JVC1228 (MIC, 24.4 µg/mL), M. luteus UST950701-006 (MIC, 24.4 µg/mL), P. nigrifaciens UST010620-005 (MIC, 97.5 µg/mL) | Standard disc diffusion assay | Penicillium sp. SCSGAF 0023 isolated from South China Sea gorgonian coral Dichotella gemmacea | [59] |
| S. aureus ATCC 29,213 (IC50 7.19 μM), M. tuberculosis (IC50 1.26 μM) | Standardized single disk method | Marine sponge-derived fungus Aspergillus sp. SCSIO XWS03F03 | [60] | ||
| 27 | Secalonic acid B | B. megaterium (15 mm), B. subtilis (MIC, 97.5 µg/mL), E. coli JVC1228 (MIC, 97.5 µg/mL), M. luteus UST950701-006 (MIC, 97.5 µg/mL), P. nigrifaciens UST010620-005 (MIC, 390.5 µg/mL) | Standard disc diffusion assay | Blennoria sp. and Penicillium sp. SCSGAF 0023 isolated from South China Sea gorgonian coral Dichotella gemmacea | [59,61,62] |
| 28 | JBIR-97/98 | S. epidermidis (IC50 0.20 (±0.04) μM), MRSA (IC50 0.19 (±0.02) μM), P. acnes (IC50 11.0 (±1.3) μM) | Microbroth dilution method | LF069 was isolated from the marine sponge Cacospinga scalaris sampled at the Limski Fjord, Croatia and classified as Engyodontium album | [63] |
| 29 | Engyodontochone A | S. epidermidis (IC50 0.19 (±0.04) μM), MRSA (IC50 0.17 (±0.02) μM), P. acnes (IC50 13.8 (±1.7) μM) | |||
| 30 | JBIR-99 | S. epidermidis (IC50 0.21 (± 0.04) μM), MRSA (IC50 0.25 (± 0.07)μM), P. acnes (IC50 14.1 (±2.7) μM) | |||
| 31 | Engyodontochone B | S. epidermidis (IC50 0.22 (±0.03) μM), MRSA (IC50 0.24 (±0.04) μM), P. acnes (IC50 11.7 (±2.4) μM) | |||
| 32 | IB-00208 | E. coli (ATCC 10536) (MIC, >150 nM), K. pneumonie (ATCC 29665) (MIC, >150 nM), P. aerigona (ATCC 10145) (MIC, >150 nM), B. subtilis (ATCC 6051) (MIC, 1.4 nM), S. aureus (ATCC 6538P) (MIC, 1.4 nM), M. luteus (ATCC 9341) (MIC, 0.09 nM) | - | Actinomadura sp. collected from northern coast of Spain | [64] |
| 33 | Microluside A | E. faecalis JH212 (MIC, 10 μM), S. aureus NCTC 8325 (MIC, 13 μM) | Microdilution assay | Micrococcus sp. EG45 was cultivated from the Red Sea sponge Spheciospongia vagabunda | [62] |
| 34 | Neocitreamicin I | B. subtilis 1A1 (MIC, 0.06 μg/mL), MRSA NRS1 (MIC, 0.50 μg/mL), MRSA NRS2 (MIC, 0.12 μg/mL), MRSA NRS71 (MIC, 0.12 μg/mL), E. faecalis (VRE 51299) (MIC, 0.06 μg/mL), E. faecalis (VRE 51575) (MIC, 0.12 μg/mL), E. coli K-12 (MIC, >8.0 μg/mL) | Liquid growth medium | Nocardia strain (G0655) isolated from a sandy soil sample collected in Falmouth, Massachusetts (USA) | [65] |
| 35 | Neocitreamicins II | B. subtilis 1A1 (MIC, 0.12 μg/mL), MRSA NRS1 (MIC, 1.0 μg/mL), MRSA NRS2 (MIC, 0.50 μg/mL), MRSA NRS71 (MIC, 0.50 μg/mL), E. faecalis (VRE 51299) (MIC, 0.06 μg/mL), E. faecalis (VRE 51575) (MIC, 0.25 μg/mL), E. coli K-12 (MIC, >8.0 μg/mL) | |||
| 36 | Citreamicin α or LL-E19085α | E. coli (MIC, >128 μg/mL), K. pneumoniae (MIC, >128 μg/mL), Serratia sp. (MIC, >128 μg/mL), Citrobacter sp. (MIC, >128 μg/mL), P. aeruginosa (MIC, >128 μg/mL), S. aureus (MIC, <0.06–0.12 μg/mL), S. epidermidis (MIC, <0.06 μg/mL), Enterococcus sp. (MIC, <0.06–0.12 μg/mL), Streptococcus sp. (MIC, <0.06 μg/mL), S. pneumoniae (MIC, <0.06 μg/mL), B. fragilis (MIC, 16 μg/mL), B. thetaiotaomicron (MIC, 4 μg/mL), Clostridium perfringens (MIC, <0.06 μg/mL), C. difficile (MIC, <0.06 μg/mL) [66] | Agar diffusion method | Marine Micromonospora sp. [67] | [66,67] |
| antifungal activity | |||||
| 4 | Norlichexanthone (3,6,8-trihydroxy-1-methylxanthone) | C. albicans (ATCC 10231) (MIC, 6.25 μg/mL), A. niger (ATCC 13496) (MIC, 25.0 μg/mL), F. oxysporum f. sp. Cubense (MIC, 50.0 μg/mL) | Agar diffusion assay | P. raistrikii (obtained from the sponge Axinella cf. corrugate) or Talaromyces sp. ZH-154 from the South-China Sea | [44,45,46] |
| E. repens (inhibition zone 1 mm) | Wardomyces anomalus isolated from the green alga Enteromorpha sp. (Ulvaceae) collected in the Baltic Sea | [47] | |||
| 6 | Yicathin C | C. lagenarium (inhibition zone 11.0 mm) | Standard agar diffusion test | Aspergillus wentii (red alga Gymnogongrus flabelliformis) collected from the coast of Pingtan Island, China | [48] |
| 10 | Buanmycin | C. albicans (MIC, 21.1 μM), A. fumigatus (MIC, 84.3 μM) | Microdilution method | Streptomyces strain from a tidal mudflat in Buan, Republic of Korea | [50] |
| 18 | Fischexanthone | F. graminearum (MIC, 474.68 µM), C. musae (MIC, 474.68 µM) | Broth tube dilution method | Mangrove endophytic fungus Alternaria sp. R6 collected in Leizhou peninsula, Guangdong Province, China | [57] |
| 37 | 1-Hydroxy-6-methyl-8-(hydroxymethyl) xanthone | E. repens (inhibition zone 2 mm), U. violacea (inhibition zone 2 mm) | Agar diffusion assay | Ulocladium botrytis (strain no. 193A4), isolated from the marine sponge Callyspongia vaginalis, collected from Dominica, Caribbean | [68] |
| 38 | 2,3,6,8-Tetrahydroxy-1-methylxanthone | M. violaceum (inhibition zone 1 mm) | Agar diffusion assay | Wardomyces anomalus Brooks & Hansford (Microascaceae, As-comycetes), isolated from the green alga Enteromorpha sp. collected around Fehmarn island in the Baltic Sea | [44] |
| 39 | 8-Hydroxy-3-methyl-9-oxo-9H-xanthene-1-carboxylic acid methylether | G. musae (Rate of inhibition 53%), P. cichoralearum (Rate of inhibition 48%), C. glocosporioides (Rate of inhibition 28%), B. graminearum (Rate of inhibition 4.6%), F. exysporum (Rate of inhibition 9.5%) | Disk assay method | Co-culture broth of two mangrove fungi (strain No. K38 and E33) collected in South China Sea coast | [69,70,71] |
| 40 | Dimethyl 8-methoxy-9-oxo-9H-xanthene-1, 6-dicarboxylat | F. oxysporum f. sp. Cubense (MIC, 12.5 µg/mL) | - | Penicillium sp. (ZZF 32#) isolated from the South China Sea | [71,72] |
| 41 | 4-Chlorofischexanthone | F. graminearum (MIC, 107 µM), C. musae (MIC, 214 µM) | Broth tube dilution method | Mangrove endophytic fungus Alternaria sp. R6 collected from the mangrove in Leizhou peninsula, Guangdong Province, China | [57] |
| 42 | Versicone A | C. cutatum (MIC, 32 μg/mL), F. oxysporum (MIC, 128 μg/mL), M. oryzae (MIC, >200 μg/mL) | Broth microdilution method | Aspergillus versicolor SCSIO 05879 collected from the Indian Ocean | [73] |
| 43 | Versicone B | C. cutatum (MIC, >200 μg/mL), F. oxysporum (MIC, >200 μg/mL), M. oryzae (MIC, >200 μg/mL) | |||
| 44 | Versicone C | C. cutatum (MIC, >200 μg/mL), F. oxysporum (MIC, >200 μg/mL), M. oryzae (MIC, >200 μg/mL) | |||
| 45 | Versicone D | C. cutatum (MIC, >200 μg/mL), F. oxysporum (MIC, >200 μg/mL), M. oryzae (MIC, >200 μg/mL) | |||
| 46 | Variecoxanthone A | C. cutatum (MIC, >200 μg/mL), F. oxysporum (MIC, >200 μg/mL), M. oryzae (MIC, >200 μg/mL) | |||
| 47 | Emerixanthones D | Fusarium sp., Penicillium sp., A. niger, R. solani, F. sporium f. sp. niveum, F. sporium f. sp. Cucumeris: Diameters of inhibition zones of which were both 3–4 mm | Filter paper discagar diffusion method | Emericella sp. SCSIO 05240 from South China Sea | [54] |
| 24 | Secalonic acid A | C. albicans (ATCC 10231) (MIC, 6.25 μg/mL), A. niger (ATCC 13496) (MIC, 6.25 μg/mL), F. oxysporum f. sp. Cubense (MIC, 12.5 μg/mL) | Microdilution assay | Talaromyces sp. ZH-154 from the South-China Sea | [45] |
| 27 | Secalonic acid B | M. violaceum (inhibition zone 13 mm) | Standard disc diffusion assay | Blennoria sp. and Penicillium sp. SCSGAF 0023 isolated from South China Sea gorgonian coral Dichotella gemmacea | [59,61] |
| 28 | JBIR-97/98 | C. albicans (IC50 4.6 (±0.5) μM), T. rubrum (IC50 4.1 (±0.8) μM) | Microbroth dilution method | LF069 was isolated from the marine sponge Cacospinga scalaris sampled at the Limski Fjord, Croatia and classified as Engyodontium album | [63] |
| 29 | Engyodontochone A | C. albicans (IC50 6.1 (±4.5) μM), T. rubrum (IC50 6.0 (±1.7) μM) | |||
| 30 | JBIR-99 | C. albicans (IC50 13.5 (±0.9) μM), T. rubrum (IC50 5.3 (±1.0) μM) | |||
| 31 | Engyodontochone B | C. albicans (IC50 4.6 (±0.7) μM), T. rubrum (IC50 4.3 (±0.9) μM) | |||
| 48 | Globosuxanthone A | C. albicans IFM 4954 (7 mm inhibition zone) | Paper disk method | B. bassiana TPU942, was isolated from a piece of an unidentified marine sponge collected at Iriomote Island | [74] |
| antiparasitic activity | |||||
| 49 | Chaetoxanthone A | T. brucei rhodesiense (strain STIB 900) (IC50 4.7 μg/mL), T. cruzi (strain Tulahuen C4) (IC50 > 10 μg/mL), L. donoVani (strain MHOM-ET-67/L82) (IC50 5.3 μg/mL), P. falciparum (IC50 3.5 μg/mL) | Modified [3H]hypoxanthine incorporation assay | Chaetomium sp. from the Greek alga originated from Kamari on the island Santorini. | [75,76,77] |
| 50 | Chaetoxanthone B | T. brucei rhodesiense (strain STIB 900) (IC50 9.3 μg/mL), T. cruzi (strain Tulahuen C4) (IC50 7.1 μg/mL), L. donoVani (strain MHOM-ET-67/L82) (IC50 3.4 μg/mL), P. falciparum (IC50 0.5 μg/mL) | |||
| 51 | Chaetoxanthone C | T. brucei rhodesiense (strain STIB 900) (IC50 42.6 μg/mL), T. cruzi (strain Tulahuen C4) (IC50 1.5 μg/mL), L. donoVani (strain MHOM-ET-67/L82) (IC50 3.1 μg/mL), P. falciparum (IC50 4.0 μg/mL) | |||
| antiviral activity | |||||
| 52 | 3,8-Dihydroxy-6- methyl-9-oxo-9H-xanthene-1-carboxylate | A/FM-1/1/47 (H1N1) (IC50 4.80 ± 1.28 μM), A/Puerto Rico/8/34 H274Y (H1N1) (IC50 9.40 ± 1.96 μM), A/Aichi/2/68 (H3N2) (IC50 5.12 ± 1.49 μM) | 3-(4,5-Dimethylthiazol-2yl)-2,5-diphenyltetrazolium bromide (MTT) colorimetric assay | Mangrove-derived fungus Diaporthe sp. SCSIO 41011from Rhizophora stylosa, which was collected in Sanya city, Hainan Province, China | [78] |
| 23 | Penicillixanthone A | HIV-1 SF162 (10 μM, 90.86 ± 0.82%) | TZM-bl cells | Jellyfish-derived fungus Aspergillus fumigatus | [79] |
| 53 | Epiremisporine B | anti-EV71 (IC50 19.8 μM), H3N2 (IC50 24.1 μM) | CPE inhibition assay [80] | Penicillium sp. SCSIO Ind16F01 was isolated from a deep-sea sediment sample collected in the Indian Ocean | [81] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loureiro, D.R.P.; Soares, J.X.; Costa, J.C.; Magalhães, Á.F.; Azevedo, C.M.G.; Pinto, M.M.M.; Afonso, C.M.M. Structures, Activities and Drug-Likeness of Anti-Infective Xanthone Derivatives Isolated from the Marine Environment: A Review. Molecules 2019, 24, 243. https://doi.org/10.3390/molecules24020243
Loureiro DRP, Soares JX, Costa JC, Magalhães ÁF, Azevedo CMG, Pinto MMM, Afonso CMM. Structures, Activities and Drug-Likeness of Anti-Infective Xanthone Derivatives Isolated from the Marine Environment: A Review. Molecules. 2019; 24(2):243. https://doi.org/10.3390/molecules24020243
Chicago/Turabian StyleLoureiro, Daniela R. P., José X. Soares, Joana C. Costa, Álvaro F. Magalhães, Carlos M. G. Azevedo, Madalena M. M. Pinto, and Carlos M. M. Afonso. 2019. "Structures, Activities and Drug-Likeness of Anti-Infective Xanthone Derivatives Isolated from the Marine Environment: A Review" Molecules 24, no. 2: 243. https://doi.org/10.3390/molecules24020243