
Eruca sativa Meal against Diabetic Neuropathic Pain: An H2S-Mediated Effect of Glucoerucin

 , , ,
, , ,  and
and 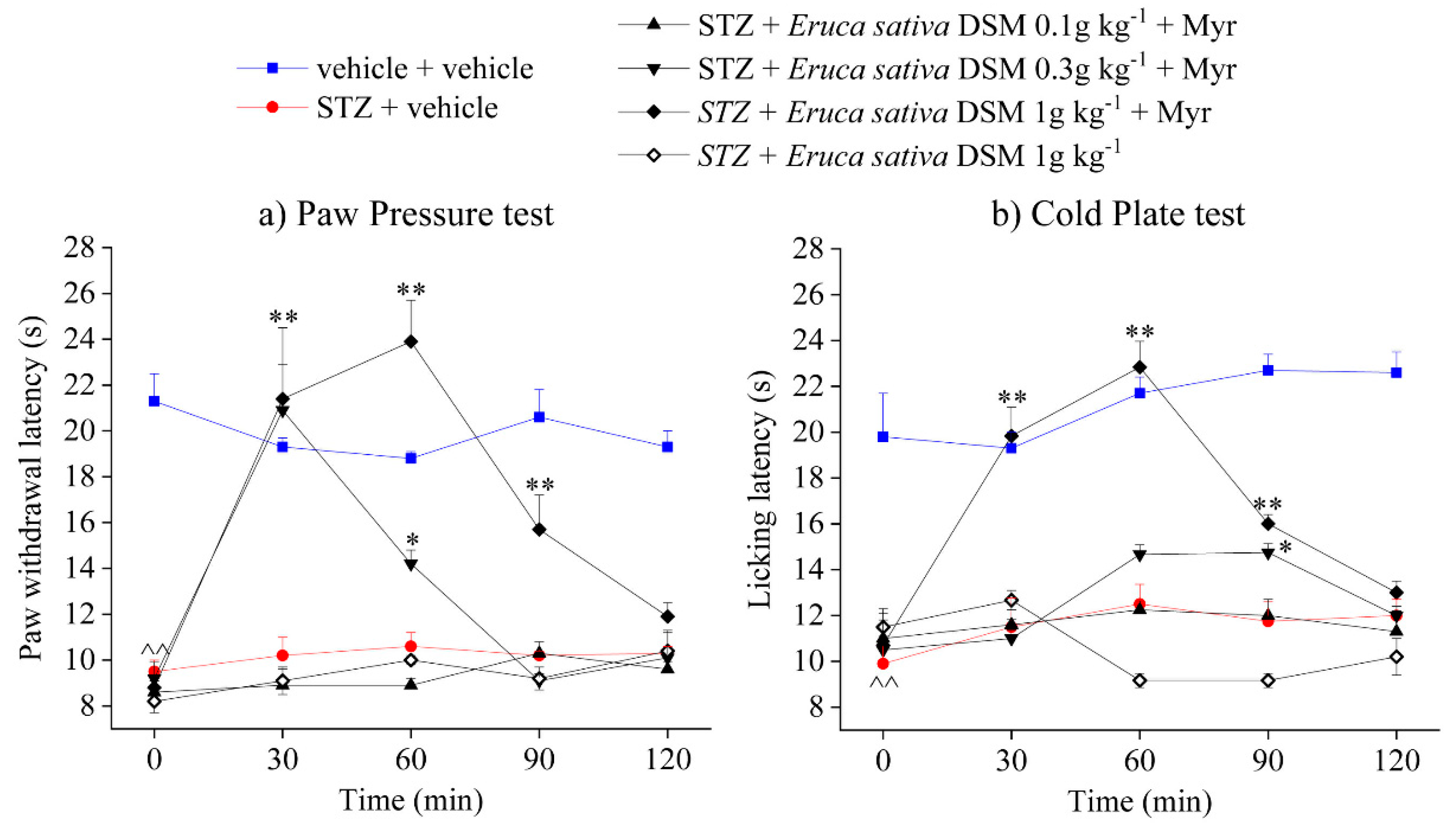{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Eruca sativa Defatted Seed Meal Characterization
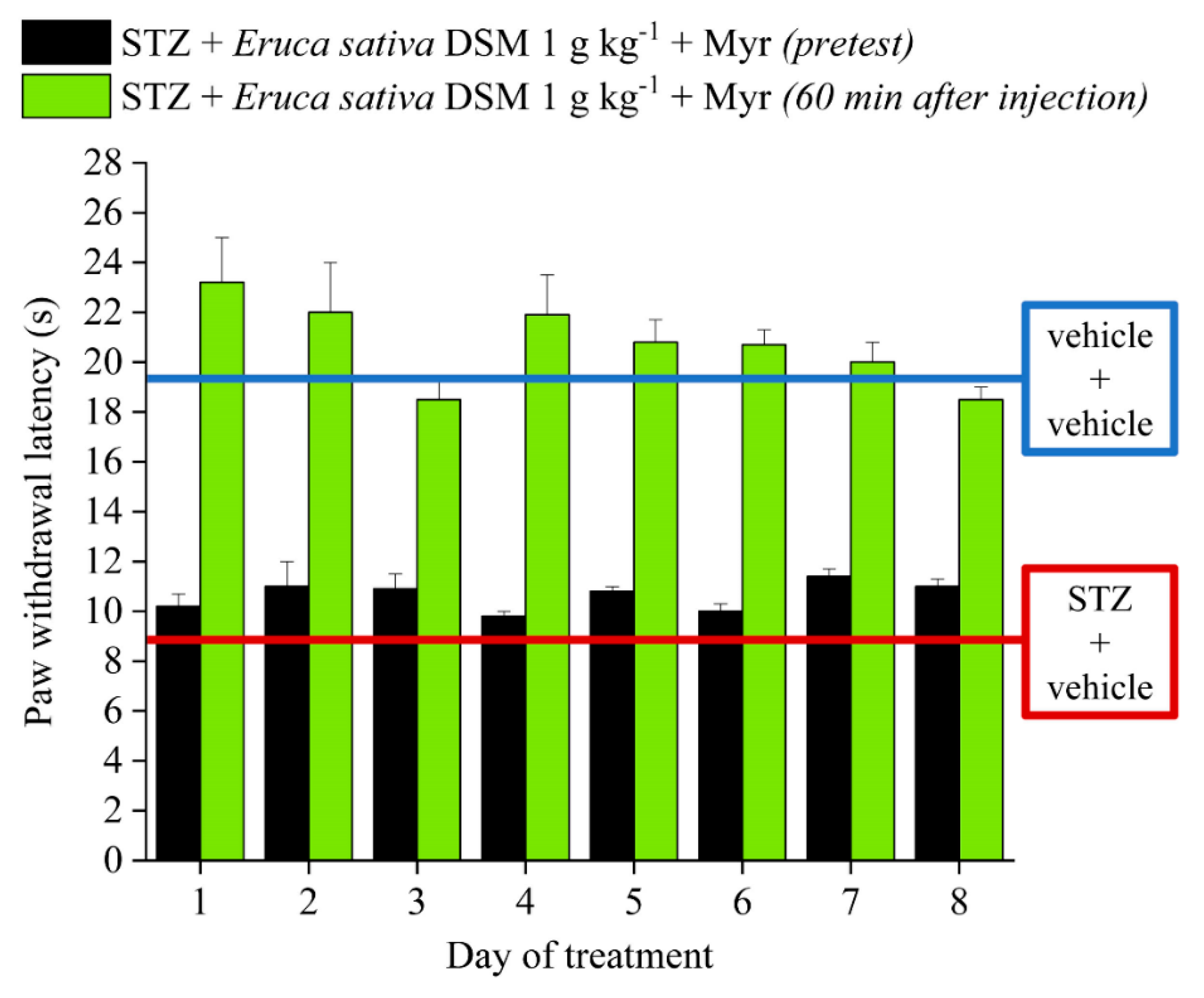
2.2. Effect of Eruca sativa Defatted Seed Meal and Glucoerucin on Diabetes-Induced Neuropathic Pain
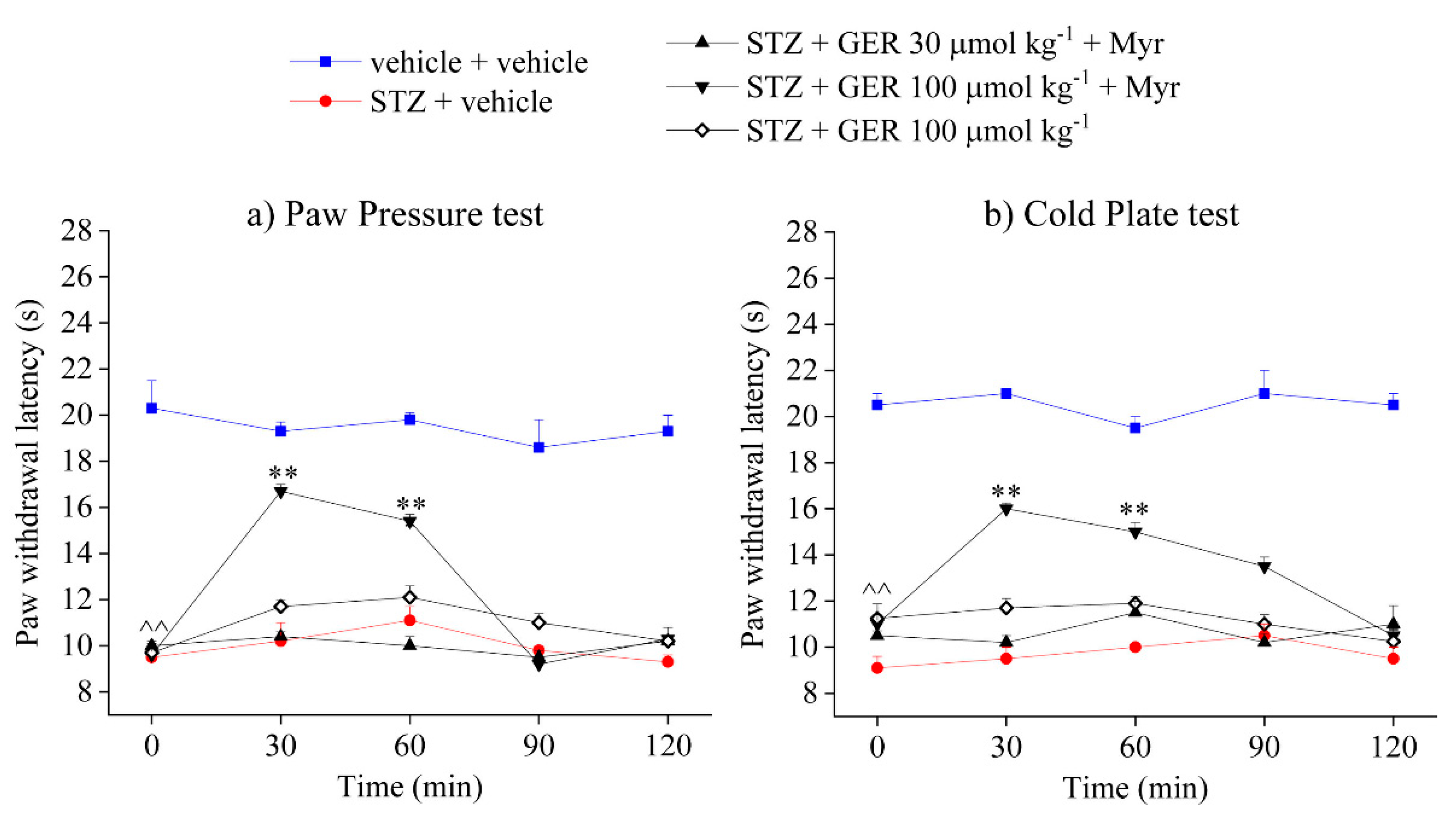
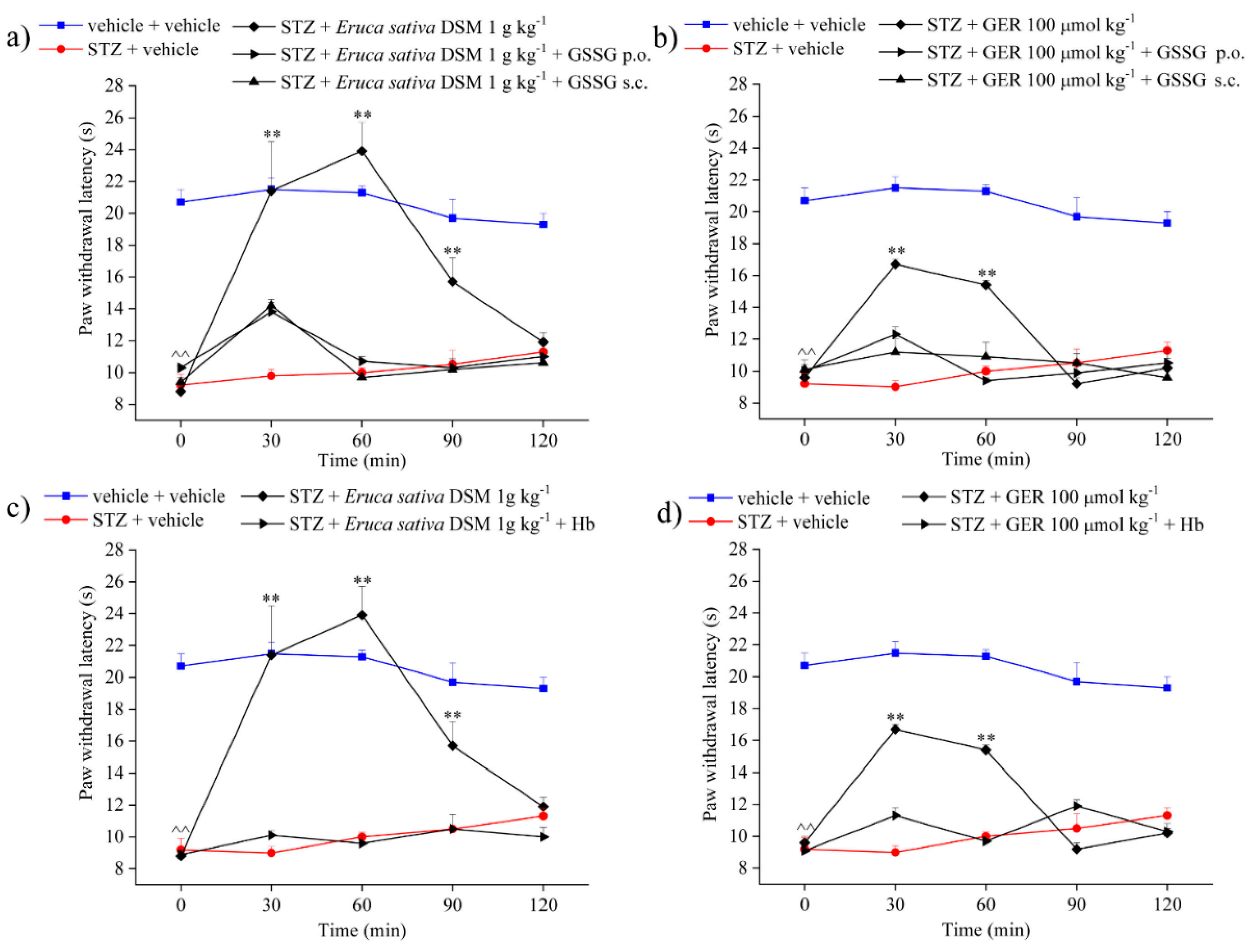
2.3. Role of Isothiocyanates and H2S-Release in the Anti-Hyperalgesic Effect of Eruca sativa Defatted Seed Meal and Glucoerucin
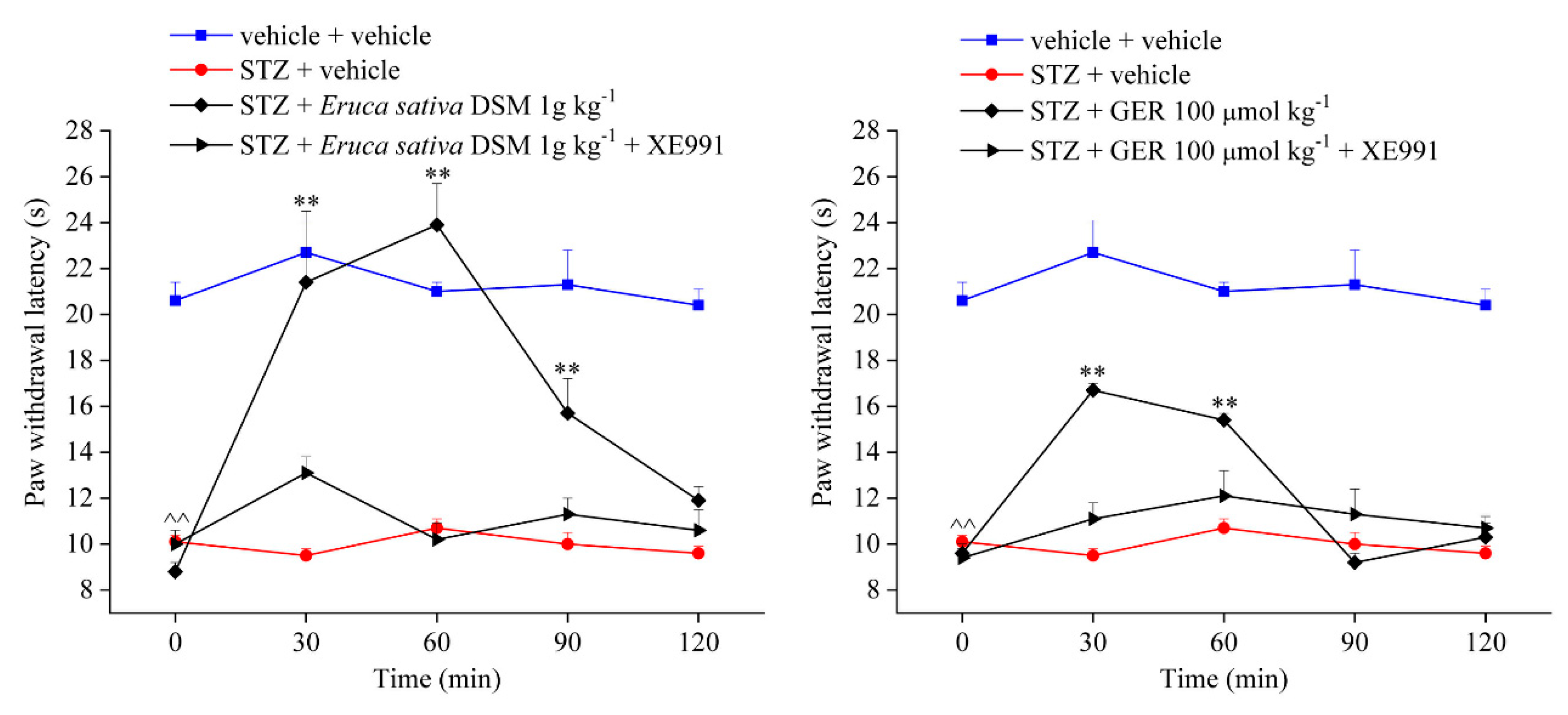
2.4. Involvement of Kv7 Potassium Channels in the Pain-Relieving Effect of Eruca sativa Defatted Seed Meal and Glucoerucin
3. Discussion
4. Materials and Methods
4.1. Eruca sativa Defatted Seed Meal Production and Characterization
- (1)
- Moisture content was determined to evaluate the difference between its weight before and after oven-drying at 105 °C for 12 h.
- (2)
- Proteins were determined from the total content of nitrogen determined using the elemental analyzer LECO CHN TruSpec according to the American Society for Testing Materials (ASTM D5373).
- (3)
- Residual oil content was determined by the standard Soxhlet extraction method using hexane as a solvent and characterized for its fatty acid composition by the UNI EN ISO 5508 method (1998) [67]. Fatty acid composition of residual oil was analyzed after trans-methylation in 2N KOH methanol solution. Fatty acid methyl esters (FAMEs) were evaluated by gas chromatography and the internal normalization method [68] was used for determining the fatty acid profile.
- (4)
- Glucosinolate content was determined following the ISO 9167-1 method with some minor modifications [69]. Briefly, 250 mg DSM were extracted in 70% ethanol at 80 °C. One milliliter of crude extract was loaded onto a DEAE Sephadex A-25 (GE Healthcare, Freiburg, Germany) mini-column. After washing with 25 mM acetate buffer (pH 5.6), GSLs were desulfated by adding purified sulfatase (200 µL, 0.35 U/mL). The desulfo-GSLs were eluted in water (HPLC grade) and detected in HPLC-UV [69] monitoring their absorbance at 229 nm. They were identified with respect to their UV spectra and retention time, according to our library [70], and their amounts were estimated using sinigrin as an internal standard. Each extraction and analysis was performed in triplicate.
- (5)
- Total free phenolic content was assayed with the Folin–Ciocalteu method according to [71]. Values are the mean ± SD of three independent extractions by four replicates for each measurement. Eruca sativa DSM extracts were obtained in acidified ethanol, ethanol/1 N HCl (85:15; v/v), after 30 min at 21 °C in 40 kHz ultrasonic bath (Sonica Sweep System, Soltec). The supernatants of a triple extraction procedure were collected and maintained at −20 °C in the dark for 48 h to facilitate macromolecule precipitation. Five serial dilutions of the filtered extracts were assayed at 765 nm, 20 °C, in an Infinite M200 NanoQuant Plate reader (Tecan, Switzerland). The slope of each calibration curve was compared to a standard gallic acid calibration curve (range 0.3–27 µg ml−1, r2 = 0.9972). The slope ratio of sample/standard curves was calculated, and results were expressed as mg of GAE per g of DSM.
- (6)
- Myrosinase activity was determined by the pH-stat technique according to [28]. Briefly, 300 mg of E. sativa DSM were loaded in 15 mL of 1% NaCl into a reaction cell at 37 °C in a DL50 pH-Stat titrator (Mettler Toledo, Switzerland). The reaction started by adding 0.5 mL of 0.5 M sinigrin solution in distilled water, after 8–10 min of conditioning, and was monitored following NaOH additions used to maintain pH constant at 6.5, versus time in minutes. The assay was carried out in triplicate. One enzyme unit (U) corresponded to 1 μmol/g DSM of sinigrin transformed in 1 min.
4.2. Isolation of Glucoerucin
4.3. Animals
4.4. Induction of Diabetic Neuropathy in Mice
4.5. Assessment of Mechanical Hyperalgesia
4.6. Assessment of Thermal Allodynia
4.7. Compounds Administration
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Abbott, C.A.; Malik, R.A.; Van Ross, E.R.; Kulkarni, J.; Boulton, A.J. Prevalence and characteristics of painful diabetic neuropathy in a large community based diabetic population in the U.K. Diabetes Care 2011, 34, 2220–2224. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, S.; Boulton, A.J.; Dickenson, A.H. Mechanisms and management of diabetic painful distal symmetrical polyneuropathy. Diabetes Care 2013, 36, 2456–2465. [Google Scholar] [CrossRef] [PubMed]
- Boyle, J.; Eriksson, M.E.; Gribble, L.; Gouni, R.; Johnsen, S.; Coppini, D.V.; Kerr, D. Randomized, placebo-controlled comparison of amitriptyline, duloxetine, and pregabalin in patients with chronic diabetic peripheral neuropathic pain: Impact on pain, polysomnographic sleep, daytime functioning, and quality of life. Diabetes Care 2012, 35, 2451–2458. [Google Scholar] [CrossRef]
- Di Cesare Mannelli, L.; Lucarini, E.; Micheli, L.; Mosca, I.; Ambrosino, P.; Soldovieri, M.V.; Martelli, A.; Testai, L.; Taglialatela, M.; Calderone, V.; et al. Effects of natural and synthetic isothiocyanate-based H2S-releasers against chemotherapy-induced neuropathic pain: Role of Kv7 potassium channels. Neuropharmacology 2017, 121, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Lucarini, E.; Micheli, L.; Martelli, A.; Testai, L.; Calderone, V.; Ghelardini, C.; Di Cesare Mannelli, L. Efficacy of isothiocyanate-based compounds on different forms of persistent pain. J. Pain Res. 2018, 11, 2905–2913. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Citi, V.; Martelli, A.; Testai, L.; Marino, A.; Breschi, M.C.; Calderone, V. Hydrogen sulfide releasing capacity of natural isothiocyanates: Is it a reliable explanation for the multiple biological effects of Brassicaceae? Planta Medica 2014, 80, 610–613. [Google Scholar] [CrossRef] [PubMed]
- Mithen, R.; Ho, E. Isothiocyanates for Human Health. Mol. Nutr. Food Res. 2018, 62, 1870079. [Google Scholar] [CrossRef] [PubMed]
- Ishida, M.; Hara, M.; Fukino, N.; Kakizaki, T.; Morimitsu, Y. Glucosinolate metabolism, functionality and breeding for the improvement of Brassicaceae vegetables. Breed. Sci. 2014, 64, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Fahey, J.W.; Wade, K.L.; Wehage, S.L.; Holtzclaw, W.D.; Liu, H.; Talalay, P.; Fuchs, E.; Stephenson, K.K. Stabilized sulforaphane for clinical use: Phytochemical delivery efficiency. Mol. Nutr. Food Res 2017, 61, 1600766. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Beltran, C.E.; Calderon-Oliver, M.; Pedraza-Chaverri, J.; Chirino, Y.I. Protective effect of sulforaphane against oxidative stress: Recent advances. Exp. Toxicol. Pathol. 2012, 64, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kensler, T.W.; Cho, C.G.; Posner, G.H.; Talalay, P. Anticarcinogenicactivities of sulforaphane and structurally related synthetic norbornylisothiocyanates. Proc. Natl. Acad. Sci. USA 1994, 91, 3147–3150. [Google Scholar] [CrossRef]
- Pu, D.; Zhao, Y.; Chen, J.; Sun, Y.; Lv, A.; Zhu, S.; Luo, C.; Zhao, K.; Xiao, Q. Protective effects of sulforaphane on cognitive impairments and ad-like lesions in diabetic mice are associated with the upregulation of Nrf2 transcription activity. Neuroscience 2018, 381, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Silva Rodrigues, J.F.; Silva, E.; Silva, C.; França Muniz, T.; De Aquino, A.F.; Neuza da Silva Nina, L.; Fialho Sousa, N.C.; Nascimento da Silva, L.C.; De Souza, B.G.G.F.; Da Penha, T.A.; et al. Sulforaphane Modulates Joint Inflammation in a Murine Model of Complete Freund’s Adjuvant-Induced Mono-Arthritis. Molecules 2018, 23, 988. [Google Scholar] [CrossRef] [PubMed]
- Tarozzi, A.; Angeloni, C.; Malaguti, M.; Morroni, F.; Hrelia, S.; Hrelia, P. Sulforaphane as a Potential Protective Phytochemical against Neurodegenerative Diseases. Oxidative Med. Cell Longev. 2013, 2013, 415078. [Google Scholar] [CrossRef] [PubMed]
- Yanaka, A. Daily intake of broccoli sprouts normalizes bowel habits in human healthy subjects. J. Clin. Biochem. Nutr. 2018, 62, 75–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axelsson, A.S.; Tubbs, E.; Mecham, B.; Chacko, S.; Nenonen, H.A.; Tang, Y.; Fahey, J.W.; Derry, J.M.J.; Wollheim, C.B.; Wierup, N.; et al. Sulforaphane reduces hepatic glucose production and improves glucose control in patients with type 2 diabetes. Sci. Transl. Med. 2017, 9, 394. [Google Scholar] [CrossRef] [PubMed]
- Lucarini, E.; Micheli, L.; Trallori, E.; Citi, V.; Martelli, A.; Testai, L.; De Nicola, G.R.; Iori, R.; Calderone, V.; Ghelardini, C.; et al. Effect of glucoraphanin and sulforaphane against chemotherapy-induced neuropathic pain: Kv7 potassium channels modulation by H2S release in vivo. Phytother. Res. 2018, 32, 2226–2234. [Google Scholar] [CrossRef]
- McDonnell, C.; Leánez, S.; Pol, O. The induction of the transcription factor Nrf2 enhances the antinociceptive effects of delta-opioid receptors in diabetic mice. PLoS ONE 2017, 12, e0180998. [Google Scholar] [CrossRef]
- Redondo, A.; Chamorro, P.A.F.; Riego, G.; Leánez, S.; Pol, O. Treatment with sulforaphane produces antinociception and improves morphine effects during inflammatory pain in mice. J. Pharmacol. Exp. Ther. 2017, 363, 293–302. [Google Scholar] [CrossRef]
- Martelli, A.; Testai, L.; Breschi, M.C.; Lawson, K.; McKay, N.G.; Miceli, F.; Taglialatela, M.; Calderone, V. Vasorelaxation by hydrogen sulphide involves activation of Kv7 potassium channels. Pharmacol. Res. 2013, 70, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Hedegaard, E.R.; Gouliaev, A.; Winther, A.K.; Arcanjo, D.D.; Aalling, M.; Renaltan, N.S.; Wood, M.E.; Whiteman, M.; Skovgaard, N.; Simonsen, U. Involvement of Potassium Channels and Calcium-Independent Mechanisms in Hydrogen Sulfide-Induced Relaxation of Rat Mesenteric Small Arteries. J. Pharmacol. Exp. Ther. 2016, 356, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.; Testai, L.; Citi, V.; Marino, A.; Bellagambi, F.G.; Ghimenti, S.; Breschi, M.C.; Calderone, V. Pharmacological characterization of the vascular effects of aryl isothiocyanates: Is hydrogen sulfide the real player? Vasc. Pharmacol. 2014, 60, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Platz, S.; Piberger, A.L.; Budnowski, J.; Herz, C.; Schreiner, M.; Blaut, M.; Hartwig, A.; Lamy, E.; Hanske, L.; Rohn, S. Bioavailability and biotransformation of sulforaphane and erucin metabolites in different biological matrices determined by LC-MS-MS. Anal. Bioanal. Chem. 2015, 407, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Valgimigli, L.; Iori, R. Antioxidant and pro-oxidant capacities of ITCs. Environ. Mol. Mutagen. 2009, 50, 222–237. [Google Scholar] [CrossRef] [PubMed]
- Lazzeri, L.; Errani, O.; Leoni, M.; Venturi, G. Eruca sativa spp. oleifera: A new non-food crop. Ind. Crops Prod. 2004, 20, 67–73. [Google Scholar] [CrossRef]
- Fuentes, E.; Alarcón, M.; Fuentes, M.; Carrasco, G.; Palomo, I. A novel role of Eruca sativa Mill. (Rocket) extract: Antiplatelet (NF-kB inhibition) and antithrombotic activities. Nutrients 2014, 6, 5839–5852. [Google Scholar] [CrossRef] [PubMed]
- Sadiq, A.; Hayat, M.Q.; Mall, S.M. Qualitative and quantitative determination of secondary metabolites and antioxidant potential of Eruca sativa. Nat. Prod. Chem. Res. 2014, 2, 1000137. [Google Scholar] [CrossRef]
- Sarwar Alam, M.; Kaur, G.; Jabbar, Z.; Javed, K.; Athar, M. Eruca sativa seeds possess antioxidant activity and exert a protective effect on mercuric chloride induced renal toxicity. Food Chem. Toxicol. 2007, 45, 910–920. [Google Scholar] [CrossRef]
- Martelli, A.; Piragine, E.; Citi, V.; Testai, L.; Pagnotta, E.; Ugolini, L.; Lazzeri, L.; Di Cesare Mannelli, L.; Manzo, O.L.; Bucci, M.; et al. Erucin exhibits vasorelaxing effects and antihypertensive activity by H2 S-releasing properties. Br. J. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Citi, V.; Piragine, E.; Pagnotta, E.; Ugolini, L.; Di Cesare Mannelli, L.; Testai, L.; Ghelardini, C.; Lazzeri, L.; Calderone, V.; Martelli, A. Anticancer properties of erucin, an H2 S-releasing isothiocyanate, on human pancreatic adenocarcinoma cells (AsPC-1). Phytother. Res. 2019, 33, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.D.; Hsu, A.; Riedl, K.; Bella, D.; Schwartz, S.J.; Stevens, J.F.; Ho, E. Bioavailability and inter-conversion of sulforaphane and erucin in human subjects consuming broccoli sprouts or broccoli supplement in a cross-over study design. Pharmacol. Res. 2011, 64, 456–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, P.; Spinozzi, S.; Pagnotta, E.; Lazzeri, L.; Ugolini, L.; Camborata, C.; Roda, A. Development of a liquid chromatography-electrospray ionization-tandem mass spectrometry method for the simultaneous analysis of intact glucosinolates and isothiocyanates in Brassicaceae seeds and functional foods. J. Chromatogr. A 2016, 1428, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Zaka, M.; Abbasi, B.H. Effects of bimetallic nanoparticles on seed germination frequency and biochemical characterisation of Eruca sativa. IET Nanobiotechnol. 2017, 11, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Oliviero, T.; Verkerk, R.; Dekker, M. Isothiocyanates from Brassica Vegetables—Effects of Processing, Cooking, Mastication, and Digestion. Mol. Nutr. Food Res. 2018, 62, 1701069. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Fahey, J.W.; Kostov, R.V.; Kensler, T.W. KEAP1 and done? Targeting the NRF2 pathway with sulforaphane. Trends Food Sci. Technol. 2017, 69, 257–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohwerder, T.; Sand, W. The sulfane sulfur of persulfides is the actual substrate of the sulfur-oxidizing enzymes from Acidithiobacillus and Acidiphilium spp. Microbiology 2003, 149, 1699–1710. [Google Scholar] [CrossRef]
- Mishanina, T.V.; Libiad, M.; Banerjee, R. Biogenesis of reactive sulfur species for signaling by hydrogen sulfide oxidation pathways. Nat. Chem. Biol. 2015, 11, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, A.K.; Nones, C.F.; Reis, R.C.; Chichorro, J.G.; Joice, M.; Cunha, J.M. Diabetic neuropathic pain: Physiopathology and treatment. World J. Diabetes 2015, 6, 432–444. [Google Scholar] [CrossRef]
- Callaghan, B.C.; Cheng, H.T.; Stables, C.L.; Smith, A.L.; Feldman, E.L. Diabetic neuropathy: Clinical manifestations and current treatments. Lancet Neurol. 2012, 11, 521–534. [Google Scholar] [CrossRef]
- Galer, B.S.; Gianas, A.; Jensen, M.P. Painful diabetic polyneuropathy: Epidemiology, pain description, and quality of life. Diabetes Res. Clin. Pract. 2000, 47, 123–128. [Google Scholar] [CrossRef]
- Themistocleous, A.C.; Ramirez, J.D.; Shillo, P.R.; Lees, J.G.; Selvarajah, D.; Orengo, C.; Tesfaye, S.; Rice, A.S.; Bennett, D.L. The Pain in Neuropathy Study (PiNS): A cross-sectional observational study determining the somatosensory phenotype of painful and painless diabetic neuropathy. Pain 2016, 157, 1132–1145. [Google Scholar] [CrossRef] [PubMed]
- Gore, M.; Brandenburg, N.A.; Dukes, E.; Hoffman, D.L.; Tai, K.S.; Stacey, B. Pain severity in diabetic peripheral neuropathy is associated with patient functioning, symptom levels of anxiety and depression, and sleep. J. Pain Symptom Manag. 2005, 30, 374–385. [Google Scholar] [CrossRef] [PubMed]
- Malcangio, M.; Tomlinson, D.R. A pharmacologic analysis of mechanical hyperalgesia in streptozotocin/diabetic rats. Pain 1998, 76, 151–157. [Google Scholar] [CrossRef]
- Murakami, T.; Iwanaga, T.; Ogawa, Y.; Fujita, Y.; Sato, E.; Yoshitomi, H.; Sunada, Y.; Nakamura, A. Development of sensory neuropathy in streptozotocin-induced diabetic mice. Brain Behav. 2013, 3, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, S. Recent advances in the management of diabetic distal symmetrical polyneuropathy. J. Diabetes Investig. 2011, 2, 33–42. [Google Scholar] [CrossRef]
- Pop-Busui, R.; Boulton, A.J.; Feldman, E.L.; Bril, V.; Freeman, R.; Malik, R.A.; Sosenko, J.M.; Ziegler, D. Diabetic Neuropathy: A Position Statement by the American Diabetes Association. Diabetes Care 2017, 40, 136–154. [Google Scholar] [CrossRef]
- Tesfaye, S.; Vileikyte, L.; Rayman, G.; Sindrup, S.H.; Perkins, B.A.; Baconja, M.; Vinik, A.I.; Boulton, A.J. The Toronto expert Panel on Diabetic Neuropathy. Painful diabetic peripheral neuropathy: Consensus recommendations on diagnosis, assessment and management. Diabetes Metab. Res. Rev. 2011, 27, 629–638. [Google Scholar] [CrossRef]
- Tölle, T.; Xu, X.; Sadosky, A.B. Painful diabetic neuropathy: A cross-sectional survey of health state impairment and treatment patterns. J. Diabetes Complicat. 2006, 20, 26–33. [Google Scholar] [CrossRef]
- Wang, A.; Leong, D.J.; Cardoso, L.; Sun, H.B. Nutraceuticals and osteoarthritis pain. Pharmacol. Ther. 2018, 187, 167–179. [Google Scholar] [CrossRef]
- Crawford, C.; Boyd, C.; Berry, K.; Deuster, P. HERB Working Group. Dietary Ingredients Requiring Further Research Before Evidence-Based Recommendations Can Be Made for Their Use as an Approach to Mitigating Pain. Pain Med. 2019. [Google Scholar] [CrossRef]
- Brain, K.; Burrows, T.L.; Rollo, M.E.; Chai, L.K.; Clarke, E.D.; Hayes, C.; Hodson, F.J.; Collins, C.E. A systematic review and meta-analysis of nutrition interventions for chronic noncancer pain. J. Hum. Nutr. Diet. 2019, 32, 198–225. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ballesta, M.C.; Moreno, D.A.; Carvajal, M. The Physiological Importance of Glucosinolates on Plant Response to Abiotic Stress in Brassica. Int. J. Mol. Sci. 2013, 14, 11607–11625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetta, M.H.; Owis, A.I.; Haddad, P.S.; Eid, H.M. The fatty acid-rich fraction of Eruca sativa (rocket salad) leaf extract exerts antidiabetic effects in cultured skeletal muscle, adipocytes and liver cells. Pharm. Biol. 2017, 55, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Khoobchandani, M.; Ojeswi, B.K.; Ganesh, N.; Srivastava, M.M.; Gabbanini, S.; Matera, R.; Iori, R.; Valgimigli, L. Antimicrobial properties and analytical profile of 850 traditional Eruca sativa seed oil: Comparison with various aerial and root plant extracts. Food Chem. 2010, 120, 217–224. [Google Scholar] [CrossRef]
- Bennett, R.N.; Rosa, E.A.; Mellon, F.A.; Kroon, P.A. Ontogenic profiling of glucosinolates, flavonoids, and other secondary metabolites in Eruca sativa (salad rocket), Diplotaxis erucoides (wall rocket), Diplotaxis tenuifolia (wild rocket), and Bunias orientalis (Turkish rocket). J. Agric. Food Chem. 2006, 54, 4005–4015. [Google Scholar] [CrossRef] [PubMed]
- Lazzeri, L.; Leoni, O.; Manici, L.M.; Palmieri, S.; Patalano, G. Use of Seed Flour as Soil Pesticide. U.S. Patent No. 7749549, 6 July 2010. [Google Scholar]
- Rajanandh, M.G.; Kosey, S.; Prathiksha, G. Assessment of antioxidant supplementation on the neuropathic pain score and quality of life in diabetic neuropathy patients—A randomized controlled study. Pharmacol. Rep. 2014, 66, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Stepanović-Petrović, R.; Micov, A.; Tomić, M.; Pecikoza, U. Levetiracetam synergizes with gabapentin, pregabalin, duloxetine and selected antioxidants in a mouse diabetic painful neuropathy model. Psychopharmacology (Berl.) 2017, 234, 1781–1794. [Google Scholar] [CrossRef] [PubMed]
- Aswar, M.; Patil, V. Ferulic acid ameliorates chronic constriction injury induced painful neuropathy in rats. Inflammopharmacology 2016, 24, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Hasanein, P.; Mohammad Zaheri, L. Effects of rosmarinic acid on an experimental model of painful diabetic neuropathy in rats. Pharm. Biol. 2014, 52, 1398–1402. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, F.; Braun, C.; Zhou, Y.Q.; Rittner, H.; Tian, Y.K.; Cai, X.Y.; Ye, D.W. Role of curcumin in the management of pathological pain. Phytomedicine 2018, 48, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; Wang, R. Hydrogen sulfide-based therapeutics: Exploiting a unique but ubiquitous gasotransmitter. Nat. Rev. Drug Discov. 2015, 14, 329–345. [Google Scholar] [PubMed]
- Angelino, D.; Dosz, E.B.; Sun, J.; Hoeflinger, J.L.; van Tassell, M.L.; Chen, P.; Harnly, J.M.; Miller, M.J.; Jeffery, E.H. Myrosinase-dependent and–independent formation and control of isothiocyanate products of glucosinolate hydrolysis. Front. Plant Sci. 2015, 6, 1345. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.M.; Russell, J.W.; Low, P.; Feldman, E.L. Oxidative stress in the pathogenesis of diabetic neuropathy. Endocr. Rev. 2004, 25, 612–628. [Google Scholar] [PubMed]
- Di Cesare Mannelli, L.; Zanardelli, M.; Failli, P.; Ghelardini, C. Oxaliplatin-induced neuropathy: Oxidative stress as pathological mechanism. Protective effect of silibinin. J. Pain. 2012, 13, 276–284. [Google Scholar] [CrossRef] [PubMed]
- UNI EN ISO. Animal and Vegetable Fats 536 and Oils—Analysis by Gaschromatography of Methyl Esters of Fatty Acids, Oli E Grassi Animali E Vegetali—Analisi Gascromatografica Degli Esteri Metilici Degli Acidi Grassi; Ente Nazionale Italiano di Unificazione: Milan, Italy, 1998. [Google Scholar]
- ISO. Animal and Vegetable Fats and Oils—Gas Chromatography of Fatty acid Methyl Esters—Part 4: Determination by Capillary Gas Chromatography; International Organization for Standardization: Geneva, Switzerland, 2015. [Google Scholar]
- Pagnotta, E.; Agerbirk, N.; Olsen, C.E.; Ugolini, L.; Cinti, S.; Lazzeri, L. Hydroxyl and Methoxyl Derivatives of Benzylglucosinolate in Lepidium densiflorum with Hydrolysis to Isothiocyanates and non-Isothiocyanate Products: Substitution Governs Product Type and Mass Spectral Fragmentation. J. Agric. Food Chem. 2017, 65, 3167–3178. [Google Scholar] [CrossRef] [PubMed]
- Abdull Razis, A.F.; Bagatta, M.; De Nicola, G.R.; Iori, R.; Plant, N.; Ioannides, C. Characterization of the temporal induction of hepatic xenobiotic- metabolizing enzymes by glucosinolates and isothiocyanates: Requirement for at least a 6 h exposure to elicit complete induction profile. J. Agric. Food Chem. 2012, 60, 5556–5564. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidant substrates and antioxidants by mean of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Pessina, A.; Thomas, R.M.; Palmieri, S.; Luisi, P.L. An improved method for the purification of myrosinase and its physicochemical characterization. Arch. Biochem Biophys. 1990, 280, 383–389. [Google Scholar] [CrossRef]
- Obrosov, A.; Shevalye, H.; Coppey, L.J.; Yorek, M.A. Effect of tempol on peripheral neuropathy in diet-induced obese and high-fat fed/low-dose streptozotocin-treated C57Bl6/J mice. Free Radic Res. 2017, 51, 360–367. [Google Scholar] [CrossRef] [Green Version]
- Rakieten, N.; Rakieten, M.L.; Nadkarni, M.V. Studies on the diabetogenic action of streptozotocin. Cancer Chemother. Rep. 1963, 29, 91–98. [Google Scholar] [PubMed]
- Russo, R.; D’Agostino, G.; Mattace Raso, G.; Avagliano, C.; Cristiano, C.; Meli, R.; Calignano, A. Central administration of oxytocin reduces hyperalgesia in mice: Implication for cannabinoid and opioid systems. Peptides 2012, 38, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare Mannelli, L.; Maresca, M.; Farina, C.; Scherz, M.W.; Ghelardini, C. A model of neuropathic pain induced by sorafenib in the rat: Effect of dimiracetam. Neurotoxicology 2015, 50, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Blackburn-Munro, G.; Jensen, B.S. The anticonvulsant retigabine attenuates nociceptive behaviours in rat models of persistent and neuropathic pain. Eur. J. Pharmacol. 2003, 460, 109–116. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucarini, E.; Pagnotta, E.; Micheli, L.; Parisio, C.; Testai, L.; Martelli, A.; Calderone, V.; Matteo, R.; Lazzeri, L.; Di Cesare Mannelli, L.; et al. Eruca sativa Meal against Diabetic Neuropathic Pain: An H2S-Mediated Effect of Glucoerucin. Molecules 2019, 24, 3006. https://doi.org/10.3390/molecules24163006
Lucarini E, Pagnotta E, Micheli L, Parisio C, Testai L, Martelli A, Calderone V, Matteo R, Lazzeri L, Di Cesare Mannelli L, et al. Eruca sativa Meal against Diabetic Neuropathic Pain: An H2S-Mediated Effect of Glucoerucin. Molecules. 2019; 24(16):3006. https://doi.org/10.3390/molecules24163006
Chicago/Turabian StyleLucarini, Elena, Eleonora Pagnotta, Laura Micheli, Carmen Parisio, Lara Testai, Alma Martelli, Vincenzo Calderone, Roberto Matteo, Luca Lazzeri, Lorenzo Di Cesare Mannelli, and et al. 2019. "Eruca sativa Meal against Diabetic Neuropathic Pain: An H2S-Mediated Effect of Glucoerucin" Molecules 24, no. 16: 3006. https://doi.org/10.3390/molecules24163006