Nitrate Accumulation and Expression Patterns of Genes Involved in Nitrate Transport and Assimilation in Spinach



Development and Collaborative Innovation Center of Plant Germplasm Resources, Shanghai Engineering Research Center of Plant Germplasm Resources, College of Life and Environment Science, Shanghai Normal University, Shanghai 200234, China
*
Authors to whom correspondence should be addressed.
Molecules 2018, 23(9), 2231; https://doi.org/10.3390/molecules23092231
Submission received: 16 July 2018
/
Revised: 25 August 2018
/
Accepted: 29 August 2018
/
Published: 2 September 2018
Abstract
:Excessive accumulation of nitrate in spinach is not only harmful to human beings, but also limits the efficiency of nitrogen usage. However, the underlying mechanism of nitrate accumulation in plants remains unclear. This study analyzed the physiological and molecular characteristics of nitrate uptake and assimilation in the spinach varieties with high or low nitrate accumulation. Our results showed that the variety of spinach with a high nitrate content (So18) had higher nitrate uptake compared to the variety with a low nitrate content (So10). However, the nitrate reductase activities of both varieties were similar, which suggests that the differential capacity to uptake and transport nitrate may account for the differences in nitrate accumulation. The quantitative PCR analysis showed that there was a higher level of expression of spinach nitrate transporter (SoNRT) genes in So18 compared to those in So10. Based on the function of Arabidopsis homologs AtNRTs, the role of spinach SoNRTs in nitrate accumulation is discussed. It is concluded that further work focusing on the expression of SoNRTs (especially for SoNRT1.4, SoNRT1.5 and SoNRT1.3) may help us to elucidate the molecular mechanism of nitrate accumulation in spinach.
1. Introduction
Nitrate is a common chemical compound in Nature, widely found in soils, waters, and foods. Nitrate itself is relatively non-toxic, but its metabolites may produce a number of health effects, such as infantile methaemoglobinaemia, carcinogenesis and possibly even teratogenesis [1,2]. Vegetables are the major source of the daily intake of nitrate by human beings [3]. Although more recent work has shown that the potential health concerns of nitrate were somewhat exaggerated, nitrate content as an important determinant of the quality of vegetables draws attention around the world. Moreover, it is well known that the majority of the absorbed nitrate is stored in the vacuoles of both roots and shoots. The remobilization and re-utilization of this nitrate is a big challenge for increasing nitrogen use efficiency in vegetables [4,5]. Thus, the suppression of nitrate content in vegetables is a major concern also in terms of potential health hazards and its potential limitations in improving nitrogen use efficiency.
Understanding the mechanism of nitrate accumulation metabolism is a prerequisite for reducing nitrate content in vegetables. It is known that the equilibrium of nitrate concentration in vegetables depends mainly on the nitrate uptake, translocation, assimilation and storage. Factors that influence these process can affect nitrate concentration in vegetables, such as plant genetic factors, environmental factors (photoperiod, CO2, temperature, irrigation, etc.) and agricultural factors (nitrogen doses and chemical forms, availability of other nutrients, etc.) [6]. Many studies have been carried out, which focused on reducing nitrate content in vegetables, such as the optimization of the application of nitrogen fertilizers [7,8,9,10], rational application of nitrification inhibitors (3,4-dimethylpyrazole phosphate and dicyandiamide) [11], exogenous salicylic acid [12] and iodine fertilization [13], growing plants under proper light intensity [14], CO2 concentrations [15] and organic cropping systems [16]. However, up to now, little is known about the molecular mechanisms of nitrate accumulation in plants.
Understanding the plant nitrate transporter and nitrate reductase (NR) enzymes provides us with clues to investigate the mechanism of nitrate accumulation in spinach. Plant nitrate uptake from soil depends on two systems: a low-affinity transporter system and a high-affinity transporter system. These involve two families of proteins, which are named nitrate transporter 1 (NRT1)/peptide transporter (PTR) and nitrate transporter 2 (NRT2), respectively [17,18]. Fifty-three putative NRT1/PTR (also known as NPF) genes were predicted in the genome of Arabidopsis thaliana and 10 were characterized as nitrate transporters [19]. Many NPF family members in other plant species, such as Populus trichocarpa, Cucumis sativus and Oryza sativa, were also identified [20,21,22]. Some of them were found to play an important role in nitrate accumulation in plants. For example, Arabidopsis NRT1.4 is a low-affinity nitrate transporter, which is dominantly expressed in the leaf petioles. The nitrate content of the petiole in nrt1.4 mutants was reduced to 50–64% of the wild-type level [23]. Arabidopsis NPF2.3 is a member of the nitrate excretion transporter (NAXT) sub-group of the NRT1/PTR family (NPF). Both root-to-shoot nitrate translocation and nitrate content in shoots were reduced under saline stress in npf2.3 mutants [24]. Expression of the Cucumis sativus NRT1.7 in Arabidopsis nrt1.7-2 mutants resulted in reduced nitrate content and larger leaf size [25]. Zhao et al. found that the expression of BnNRT2 in root, leaves and stem in the variety with a high nitrate content was higher than that in the variety of Chinese cabbage with low nitrate (Brassica campestris ssp. Chinensis (L.) Makino) [26]. In maize, ZmNrt2.1 may be related to maize nitrate accumulation, as its expression in response to nitrate availability was similar to that of the nitrate influx in roots [27]. All these findings have suggested that NRT genes may play an important role in regulation of the nitrate level in plants. However, up to now, little is known about the spinach NRT genes and their roles in nitrate accumulation remain unclear.
Nitrate reductase (NR) is a key regulatory enzyme in the NO3- assimilation pathway, which reduces NO3− to NO2−. After this, the NO2− was reduced to NH4+, while the NH4+ is incorporated into glutamine via glutamine synthetase (GS). It has been suggested that the high NR activity is beneficial for nitrate reduction, which subsequently decreases nitrate accumulation in plants [28]. The expression of the putative nitrate reductase genes (NIA1 and NIA2) were significantly higher in the genotype of pakchoi with a low nitrate content compared to those that had a higher level of accumulated nitrate [29]. Moreover, overexpression of nitrate reductase genes in lettuce [30] and potato [31] was proven to be helpful in reducing the nitrate content of plants. GS, which is a key enzyme involved in ammonium assimilation, may also function in reducing the nitrate content in plants. However, few reports have focused on the relationship between GS and nitrate accumulation in plants. In summary, although the molecular physiology of nitrate dynamics is well understood in model systems, their attention was mainly devoted to enhancing crop production. Little attention has been paid to the nutritional quality traits determining nitrate accumulation in plants. Thus, the molecular mechanism of nitrate accumulation in plants needs to be investigated.
Spinach (Spinacia oleracea L.) is an edible flowering plant cultivated as one of the most popular vegetables all over the world. It is identified as a good source of vitamins A, C and E, folic acid, minerals (USDA Nutrient Database; http://ndb.nal.usda.gov/ndb/search/list) and other bioactive compounds such as phenolics, carotenoids, glycoglycerol lipids, and lipoic acid. However, high levels of accumulated nitrate have often been found in spinach which may affect human health [2,32,33]. The FAO and WHO food commissions report that the average daily nitrate intake of a 60 kg person should be 220–240 mg. The European Union Food Commission States established an acceptable daily intake (ADI) for nitrate as 0–3.7 mg·kg−1 of body weight and set the maximum limits of nitrogen accumulation in spinach as 3000 and 2500 mg·kg−1 for winter and spring crops, respectively [34]. According to this, most of the spinach nitrate contents reported by various countries, such as Japan [32], France [35], New Zealand [36], Swedish [37], UK [38], Turkey [39], Korea [40] and China [41] were close to or exceeded to this standard. Thus, there is an urgent need to reduce nitrate accumulation in spinach. In this study, we compared the physiological and molecular characteristics of nitrate transport and assimilation between two spinach varieties. We aim to explore the main factors that contribute to nitrate accumulation in spinach and screen the potential genes to be targeted in reducing spinach nitrate content.
2. Results
2.1. Fresh Weight and Nitrate Content
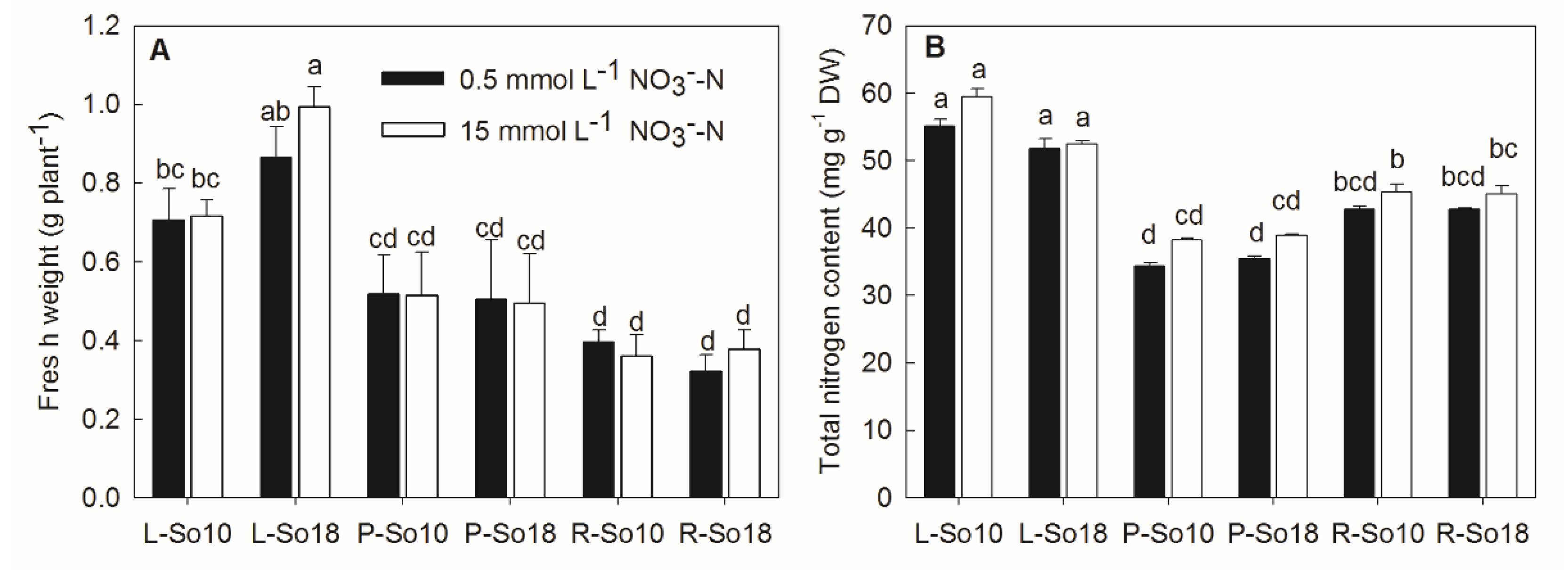
The applied nitrate concentration did not have significant effects on the fresh weight and total nitrogen content of spinach (Figure 1). Significant differences in the tissue fresh weights between two varieties were only observed on leaf blades, as the fresh weight of leaf blade, of So18 was greater than that of So10 (Figure 1A). No significant differences in total nitrogen content were found between the two varieties (Figure 1B).
The nitrate contents generally differed between the two spinach varieties and two nitrate concentration treatments (Figure 2). The nitrate contents in the whole plant of So18 were significantly greater than those in So10 at each treatment time, irrespective of the applied nitrate concentration. Among all treatments, the nitrate contents in leaf petioles were significantly greater than roots and leaf blades, except for the leaf petioles under low nitrate treatment at 1 h. Applying a greater nitrate concentration generally increased the whole plant nitrate contents for both varieties, except for So18 at 7 h and 12 h.
2.2. 15NO3−-N Uptake Rate
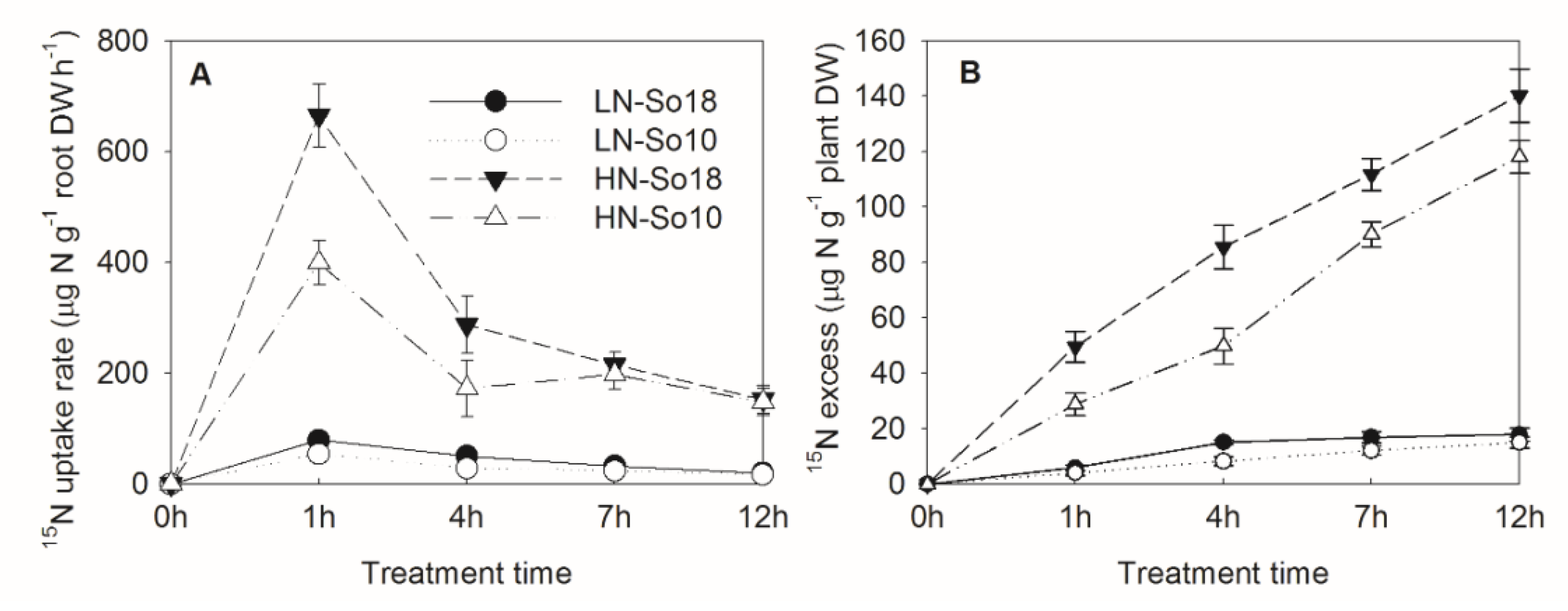
As nitrate uptake by roots is the main source of nitrate accumulation in plants, we further analyzed the 15NO3−-N uptake rate and net 15NO3−-N uptake amount (15NO3−-N excess) and compared these values in the two varieties using isotopic labeling experiment. As shown in Figure 3, the 15NO3−-N uptake rate and 15NO3−-N excess of So18 was generally higher than those of So10 under both low and high nitrate conditions (p < 0.05).
The differences of 15NO3−-N uptake rates and 15NO3−-N excess between two varieties were significant at 1–4 h and 1–12 h (except those treated with a low nitrate concentration for 1 h), respectively. The So18 15NO3−-N uptake amount was higher by 18.9% (low nitrate treatment) and 18.7% (high nitrate treatment) compared to So10 within 12 h, which suggested that So18 has a greater capacity for nitrate uptake from nutrient solution.
2.3. N Assimilation Related-Enzymes Activities
NR and GS are key enzymes involved in nitrate assimilation pathway, which play important roles in nitrate accumulation in spinach. The activities of two N assimilation related enzymes were analyzed in two spinach varieties under two different nitrate treatments. As shown in Figure 4, under low nitrate concentration treatment, the shoot NR activities of both two varieties decreased, while there was only a small change in the root NR activities. Under high nitrate concentration treatment, the dynamics of NR activities in petioles and roots were generally similar to those under low nitrate treatment. The NR activities in the leaf blade were the only exception as they first increased at 1 h and then decreased at 12 h. Under both low and high nitrate treatments, NR activities did not differ significantly between two varieties among the three tissues.
The dynamics of GS activities were more complex than those of NR. Generally, leaf blade GS activities of So18 were higher than those of So10 under both nitrate treatments. In petioles, the GS activities of two spinach varieties showed opposite trends in proportion to treatment time under both nitrate treatments. The petiole GS activities of So10 first increased at 1 h and were higher than those of So18 at 1 h, before decreasing at 12 h and becoming lower than those of So18. Under low nitrate conditions, root GS activities first increased then decreased at 12 h for both varieties, while this was reversed under high nitrate conditions. The So10 GS activities were generally higher than those of So18 under low nitrate conditions, while under high nitrate treatment, they did not differ from those of So18 at 1 h and were lower than those of So18 at 12 h.
2.4. Expression Analysis of Nitrate Transport and Assimilation Related Genes
To demonstrate a correlation between the gene expression level and N concentration as well as its subsequent effect on nitrate accumulation, a qRT-PCR assay was performed on three spinach tissues (leaf blade, leaf petiole and root) of 12 SoNRTs, two SoGLNs and one SoNIA in two spinach varieties under two nitrate concentration treatments (Table S1). They identified 12 SoNRTs, which included: two homologs of AtNRT1.1 (SoNRT1.1a and SoNRT1.1b), one homolog of AtNRT 1.2 (SoNRT1.2), one homolog of AtNRT 1.3 (SoNRT1.3), one homolog of AtNRT1.4 (SoNRT1.4), one homolog of AtNRT1.5 (SoNRT1.5), one putative spinach homolog of AtNRT1.6 and AtNRT1.7 (SoNRT1.6), two homologs of AtNRT1.8 (SoNRT1.8a and SoNRT1.8b), two homologs of AtNRT1.9 (SoNRT1.9a and SoNRT1.9b) and one homolog of AtNRT2.1 (SoNRT2). All the tested genes were shown to be expressed in all spinach tissues.
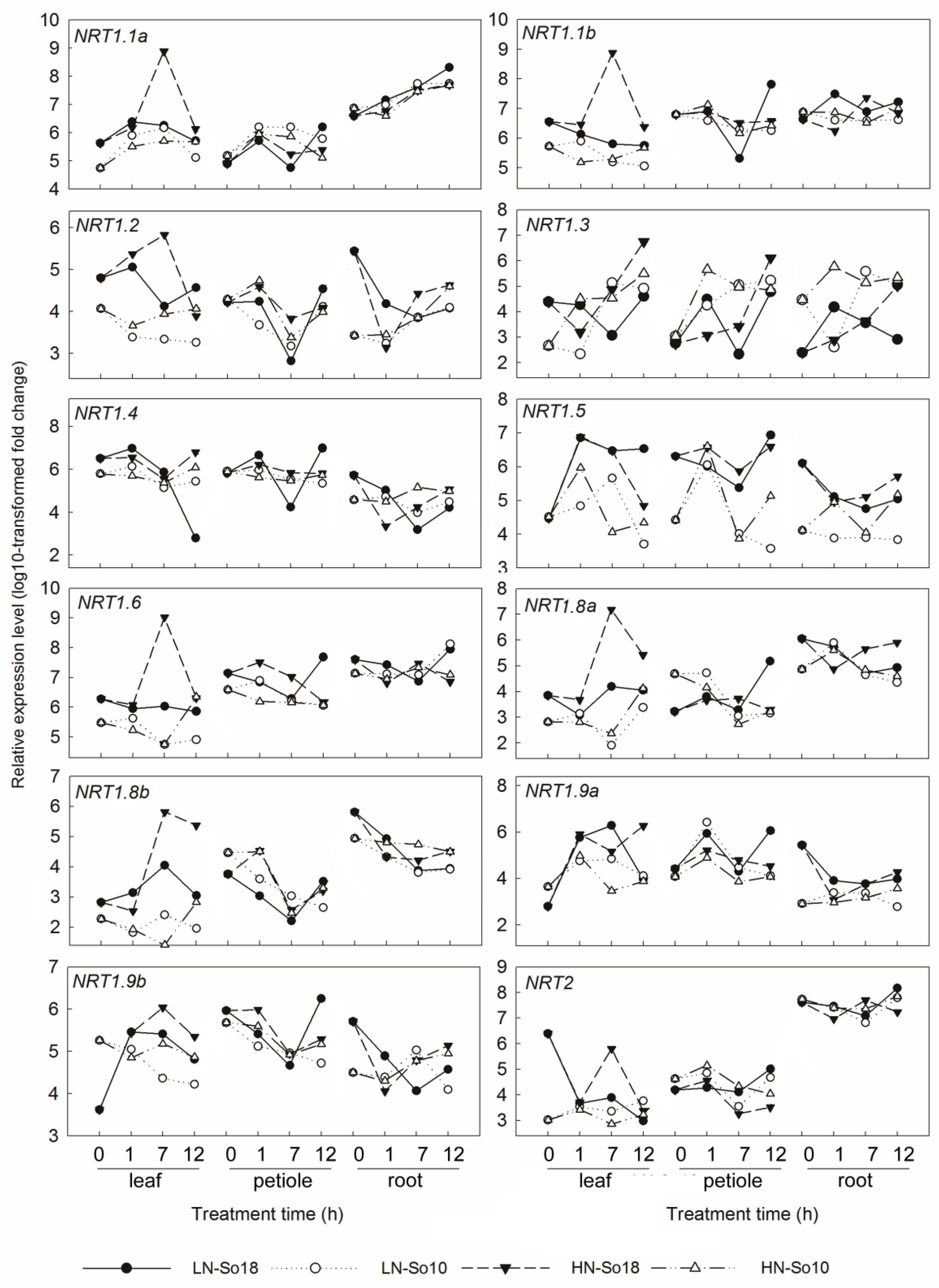
The tested genes displayed distinct expression patterns among different nitrate concentrations, tissues, treatment times and varieties (Figure 5, Figure 6 and Figure 7). Most of the tested gene transcripts were induced by high nitrate although their increases depended on treatment times and varieties (Figure S1). The relative expression levels of SoNRT1.1a, SoNRT1.1b, SoNRT1.3, SoNRT1.4, SoNRT1.6, SoNRT1.8a, SoNRT1.8b and SoNRT1.9a were at most 100–1000 times greater in varieties under high nitrate conditions than those under low nitrate conditions. SoNRT1.1a, SoNRT1.1b, SoNRT1.6, SoNRT1.8a, SoNRT1.8b, SoNRT2, SoNIA and SoGLN1 were mainly expressed in roots, while SoNRT1.4, SoNRT1.5, SoNRT1.9a, SoNRT1.9b and SoGLN2 were more abundant in leaf blades and petioles compared to roots (Figure 5). The expression level of all the tested genes varied along with the treatment time (Figure 6 and Figure 7).
As shown by repeated measures analysis of variance, significant differences in gene expression were found between two spinach varieties (Table 1). Under low nitrate treatment conditions, the transcripts of all genes in leaf blades except SoNRT 1.4 of So18 were generally greater than those of So10 across all time points. The expression levels of SoNRT1.1b, SoNRT1.4, SoNRT1.5, SoNRT1.6, SoNRT1.9a, SoNIA, SoGLN1 and SoGLN2 in petioles and SoNRT1.1b, SoNRT1.2, SoNRT1.4, SoNRT1.5, SoNRT1.8a, SoNRT1.8b, SoNRT1.9a and SoGLN1 in roots was greater in low nitrate treated So18 than those of So10. However, a significant increase of expression of five genes in petioles (SoNRT1.1a, SoNRT1.3, SoNRT1.8a, SoNRT1.8b and SoNRT1.9b) and one gene in root (SoNRT1.3) was found in So10, as there was a greater transcription level compared to those of So18. Under high nitrate treatment conditions, all of the tested genes showed higher expression levels in leaf blades of So18 than So10. The transcription level of six genes in petioles (SoNRT1.4, SoNRT1.5, SoNRT1.6, SoNRT1.8a, SoNRT1.9a, SoNIA and SoGLN1) and six genes in roots (SoNRT1.2, SoNRT1.5, SoNRT1.8a, SoNRT1.9a, SoGLN1 and SoGLN2) was increased in So18 than those in So10. In contrast, five genes in petioles (SoNRT1.1a, SoNRT1.3, SoNRT1.8a, SoNRT1.9b and SoNRT2) and three genes in roots (SoNRT1.3, SoNRT1.9b and SoNRT2) had higher expression levels in So10 than in So18. Other genes showed no significant differences in expression level between two varieties.
To understand the gene expression and their impact on plant nitrate content, we analyzed the correlations between nitrate contents and gene expression levels using Pearson correlation statistics. As shown by Table 2, the root nitrate content was positively correlated with the expression levels of SoNRT1.2 (r = 0.84, p < 0.05) and SoNRT1.8 (r = 0.82, p < 0.05) when under low nitrate treatment conditions. Under high nitrate treatment conditions, significant correlations were only observed in petioles. The petiole nitrate contents were positively correlated to the expression levels of SoNRT1.4 (r = 0.84, p < 0.05) and SoNRT1.5 (r = 0.81, p < 0.05), but negatively correlated to SoNRT1.3 (r = −0.93, p < 0.01).
3. Discussion
The main factors that influence nitrate content can differ in different plant species, varieties or tissues. To investigate the mechanism of nitrate accumulation in spinach, we analyzed the physiological and molecular characteristics of nitrate transport and assimilation. Our results showed that the plant nitrate contents differed between two spinach varieties, with higher nitrate contents in S18 and lower contents in S10, which occurred irrespectively of applied nitrate concentration. The differences in nitrate content of plants may be due to their differential capacities to absorb, reduce and assimilate nitrate. The fact that the nitrate uptake far exceeds its assimilation may account for the higher nitrate content in some cultivars [42]. In this study, the 15NO3−-N uptake rates and 15NO3−-N uptake amount of S18 were significantly greater than those of S10, while the NR and GS activities of So18 were similar to (NR activities among three tissues and leaf blade GS activities) or sometimes even lower than those of So10 (GS activities in petioles and roots). These data suggest that the nitrate accumulation differences between two tested spinach varieties is mainly due to their differential capacities to uptake and transport nitrate from the root to shoot.
Considering that nitrate uptake and translocation played a significantly more prominent role in nitrate accumulation in spinach, research on SoNRTs at the transcription level will help us to better understand the pattern of nitrate absorption in spinach and will be of great significance in further studies focusing on reducing nitrate accumulation in spinach. qRT-PCR analysis showed that the mRNA levels of the most SoNRTs in the high nitrate genotype So18 were greater than those in low nitrate variety So10, which is consistent with the greater nitrate uptake rates found in So18 compared to So10. These differences in NRT level may result from genetic variance of NRT or its regulation, which accounts for the higher nitrate uptake. A high nitrate uptake often requires high nitrate transportation and thus, we proposed that the higher NRT expression in So18 may have also indicated higher NO3− transportation in So18. Thus, these differentially expressed SoNRTs genes may be involved in nitrate uptake or nitrate translocation in spinach, which provided candidate gene resources for further investigating nitrate accumulation in spinach.
Since nitrate was mainly accumulated in petioles, we paid more attention to the differentially expressed genes in petioles. Among them, four differentially expressed genes, SoNRT1.4, SoNRT1.5, SoNRT1.6 and SoNRT1.9a were found to be more abundant in the leaf petioles of So18 than those of So10. In Arabidopsis, NRT1.4 is expressed predominantly in the shoots and roots, which has been shown to be involved in nitrate storage in the leaf petiole [23]. The mutation of AtNRT1.4 gene could greatly reduce petiole nitrate content [23]. In spinach, the homologous gene of AtNRT1.4, SoNRT1.4 showed an AtNRT1.4-like tissue expression pattern, and its mRNA level was significantly increased under high nitrate provision. This suggests that SoNRT1.4 may be a putative functional homolog of AtNRT1.4. Moreover, our study revealed a strong positive correlation of SoNRT1.4 gene expression with nitrate content in petioles. This suggested that SoNRT1.4 expression in petioles plays an important role in controlling the dynamic level of nitrate in petioles, which is a potential target or reducing nitrate accumulation.
Similar to SoNRT1.4, SoNRT1.5 expression level was also positively correlated to nitrate content in petioles. In Arabidopsis, AtNRT1.5 mediates the xylem-loading of nitrate and enhances the root-to-shoot transport of nitrate [15,43]. Similar to the expression pattern of its Arabidopsis homolog, SoNRT1.5 mRNA level was also greatly increased by nitrate provision. We suggested that SoNRT1.5 encoded protein has a similar function to AtNRT1.5 in enhancing shoot-to-root transport, and contributes to the greater nitrate accumulation in shoots of the So18 variety. Thus, SoNRT1.5 may also have a potential to be used for reducing nitrate accumulation. However, SoNRT1.5 expression was also strongly correlated with nitrate content in low nitrate treated roots. Sufficient nitrate is important for plant growth, especially when only a low concentration of nitrate available. Thus, we should consider whether the regulation of SoNRTs could reduce plant nitrate content while simultaneously not greatly affect plant growth.
As for SoNRT1.6, its Arabidopsis ortholog AtNRT1.7 was mainly expressed in the phloem tissue of older leaves, which mediates the phloem loading of nitrate in source leaves to remobilize nitrate from older leaves to nitrogen-demanding tissues [44]. In contrast, its ortholog AtNRT1.6 was only expressed in the vascular bundles of the siliques and the funiculi, which are involved in delivering nitrate to developing seeds [45]. It seems that the SoNRT1.6 encoded protein may fulfill similar physiological functions as its Arabidopsis homolog AtNRT1.7 but not AtNRT1.6. However, in contrast to AtNRT1.7 which mainly expressed in the shoot, the spinach SoNRT1.6 showed reasonable expression in both the shoot and root. If the orthologous genes of spinach NRT1.6 are also expressed in the phloem, the function of SoNRT1.6 in nitrate loading and/or unloading to or from the shoot/root is possible. SoNRT1.9a, which is another highly expressed SoNRT gene in So18, was preferentially expressed in the shoots and is strongly induced by a high concentration of nitrate. In contrast, AtNRT1.9 is expressed predominantly in roots where it is probably involved in the loading of nitrate into the root phloem to enhance downward nitrate transport in roots [43]. In addition, AtNRT1.9 expression is not rapidly induced by nitrate [46]. This suggests that the SoNRT1.9a encoded protein may fulfill different physiological functions compared to its Arabidopsis homolog. The different orthologous NRT1.9 expression in relation to tissue specificity and N-supply have also been found in other plant species. The Cucumis sativus orthologous NRT1.9 gene expression is highly up-regulated by high nitrate supply in all vegetative organs except for roots [22]. While none of the wheat homologous TaNRT1.9 genes showed dominance for root expression [47]. This emphasizes the complexity and variation in gene regulation in relation to nutrient uptake in crop plants in comparison to model plant. Being different with SoNRT1.4 and SoNRT1.5, both SoNRT1.6 and SoNRT1.9a expression showed no statistically significant correlation with the petiole nitrate content. Considering their strongly increased transcripts under high nitrate, their detailed roles in nitrate response still need to be further investigated.
Four SoNRTs (SoNRT1.1a, SoNRT1.3, SoNRT1.8a and SoNRT1.9b) have high levels of expression in the petioles of So10. Even though these SoNRTs might be involved in nitrate accumulation, their contributions to nitrate accumulation in plant might be not as high as their contributions in So18, especially considering that more SoNRTs were abundant in So18 than in So10. Interestingly, among these genes, SoNRT1.3 expression was negatively correlated with the nitrate content in petioles, which indicated that the SoNRT1.3 encoded protein may negatively regulate the dynamic level of nitrate in petioles. As far as we know, the functional role of its Arabidopsis homolog, AtNRT1.3, remains unclear although it has been proposed that it has a possible role in polyamine resistance [48]. We proposed that SoNRT1.3 encoded protein fulfills different physiological function from AtNRT1.3 and may play an important role in reducing nitrate accumulation in spinach.
It should be noted that the expression patterns of these SoNRTs in this study are mainly related to an early nitrate response (0–12 h), considering the NRT expression is spatiotemporally regulated and may change with short- and long-term nitrate treatment [49,50]). Moreover, allelic differences and/or different gene regulation efficiencies between varieties may also affect NRT expression [51] and thus, the screening of these SoNRT genes is only the first step in creating a comprehensive understanding of the nitrate accumulation mechanisms in spinach. More functional analyses of these candidate SoNRTs in nitrate accumulation in spinach need to be conducted, especially under long-term nitrate treatment conditions.
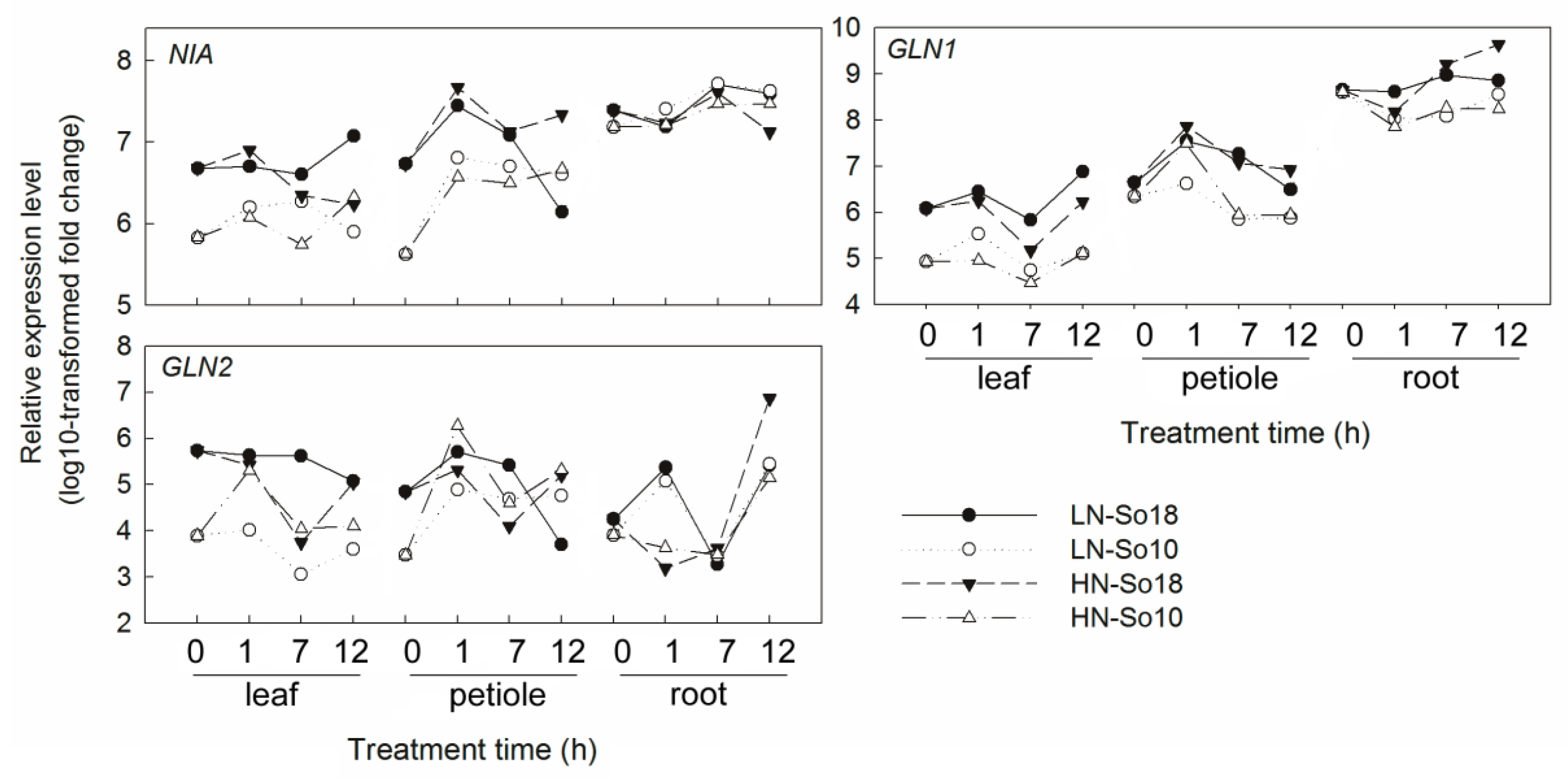
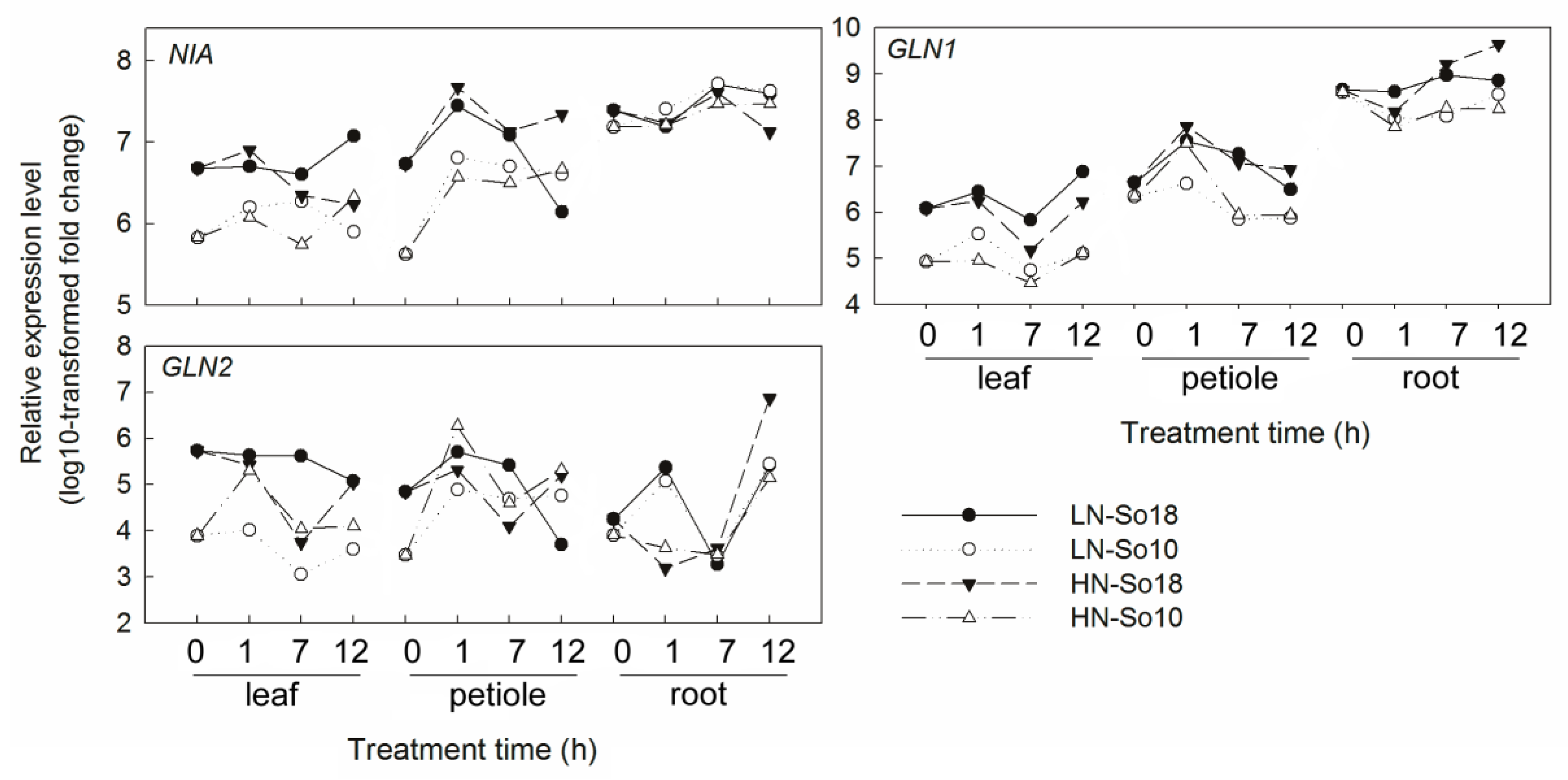
As for the nitrate assimilation related genes, it seems suggest that the spinach NRT expression is compatible with the expression of NIA and GLNs, for their expression levels were all higher in So18 than in So10. It was reported that initial exposure to nitrate causes an increase of NR activity and NR-mRNA, while the NR induction is inhibited by GS and/or its reduced N-compounds (e.g., ammonium or amino acids) after long time exposition [50]. Similar to NR, a rapid and transient increase in GS activity was also found under nitrate treatment [52]. Based on these findings, we suppose that the higher nitrate uptake and higher NRT expression in So18 cause a higher level of nitrate content in some cells, and thus stimulate an increased nitrate reduction, meaning increasing NIA and GLNs expression in So18 under early nitrate treatment conditions. With the prolonging of nitrate treatment, and the long-distance transport of N, the nitrate content varies in different tissues and subcellular fractions, which may accounts for the dynamic regulation of NR and GS by nitrate supply. Meanwhile, the net nitrate uptake may also be inhibited due to a reduced demand for nitrate or feed-back control by various C/N metabolites, which further regulated the NRT expression and also the nitrate assimilation related genes expression. The interaction between NRT, NIA and GLNs in response to nitrate condition and their roles in nitrate accumulation are promising targets for further research. During the experiment period, despite the relatively higher SoNIA and SoGLNs transcription levels in So18, the abundances of SoNRTs increased greatly compared to that of SoNIA and SoGLNs in response to a high nitrate concentration. As a result, the SoNIA and SoGLNs abundances were not high compared to the super-high nitrate concentration in So18.
It should be note that, the expression patterns of SoNIA, SoGLN1 and SoGLN2 in response to nitrate provision were not consistent with changes in their enzyme activities. Generally, the mRNA levels of these genes were higher in So18 than in So10 irrespectively of nitrate concentration, while NR activities did not differ between two varieties and GS activities fluctuated greatly between all treatments. The absence of consistency between the transcripts and enzyme activities may be due to the post-transcriptional and post-translational regulation, while the highly regulated NR enzyme not only can be regulated by many internal (e.g., internal nitrate content, reduced N-compounds) and external (e.g., nitrate application, light, CO2) factors [53,54,55,56]. Another explanation for this phenomenon may be related to the study is mainly for early expression response of homologous genes, while the enzyme activity tested here is a mixture of the homologous products. The detailed roles of NIA and GLNs in nitrate accumulation still need to be further investigated.
4. Materials and Methods
4.1. Plant Sample Preparation
Spinach seeds (So10 and So18) were sterilized in 70% ethanol for 5 min and 10% (v/v) H2O2 for 30 min. After this, they were rinsed with distilled water, and germinated in a plant growth chamber under a 10 h day/14 h night cycle (23 °C /16 °C day/night temperature cycle) with 60–70% relative humidity. The uniform seedlings with four leaves were transferred into a nutrition solution. The nutrient solution which was used to support plant growth contained: 0.2 g·L−1 KNO3, NaH2PO4 2H2O 0.09 g·L−1, Na2HPO4 12H2O 0.15 g·L−1, CaCl2 2H2O 0.29 g·L−1, MgSO4 7H2O 0.49 g·L−1, NaFeEDTA 0.02 g L−1, H3BO3 2.86 mg·L−1, MnCl2 4H2O 1.81 mg·L−1, ZnSO4 7H2O 0.22 mg·L−1, CuSO4 5H2O 0.08 mg·L−1 and NaMoO3 2H2O 0.09 mg·L−1. The pH was adjusted using 0.1 mol·L−1 NaOH and 0.1 mol·L−1 HCl to be within a pH range of 5.0–7.0. After two weeks, seedlings of uniform size and growth were picked randomly and transferred to −N (N free) solution for 4 d (other nutrients remained as before), before being immersed in 0.5 mmol·L−1 KNO3 or 15 mmol·L−1 KNO3 (other nutrients remained as before) from 0 to 12 h. The high nitrate concentration (15 mmol·L−1) represents the maximum amount of nitrate for hydroponic spinach growth. Our pre-experiment indicated that the tested spinach varieties could grow well under 15 mmol·L−1 nitrate when compared with 20 and 30 mmol·L−1 concentrations (data not shown). Plants were harvested and three independent replicates (three plants for each replicate) were collected for each sample. Plant materials were frozen in liquid nitrogen immediately and stored at −80 °C until subsequent analyses.
4.2. 15NO3−-N Uptake Rate
Nitrate uptake was determined by the 15N isotopic tracer method as previously described [57]. Briefly, the plants were exposed to 0.1 mmol·L−1 CaSO4 for 1 min, before being immersed in a complete nutrient solution containing 0.5 or 15 mmol·L−1 15NO3− for 0 h, 1 h, 4 h, 7 h or 12 h. After this, they were rinsed in 0.1 mmol·L−1 CaSO4 for 1 min before washing with deionized water. The plant shoots and roots were freeze-dried, weighed to determine the dry weight, and subsequently milled to fine powder. The total contents of N and 15N were determined using a Vario ELIII elemental analyzer (Elementar, Langenselbold, Germany). The rates of 15N uptake were calculated by dividing total plant 15N amount at time by the root dry weight in grams (µg N·g−1 root DW·h−1). The net 15N uptake amount by plant was represented as 15NO3−-N excess (µg N·g-1 plant DW).
4.3. Measurements of Nitrate Content and Enzymes Activity
Plant samples were sampled and washed with flowing water to obtain the whole plants. They were then divided into roots, petioles, and leaf blades to determine the nitrate contents in different organs. Nitrate content determination followed the method described by Tang et al. [58]. The method of determining NR and GS activity was described by Tang et al. [58] and Wang et al. [59]. One unit of NR activity is defined as the production of 1 µg nitrite per hour per fresh weight (μg NO2−·g−1 FW·h−1). One unit of GS activity is defined as 1 A540·g−1 FW·h−1.
4.4. Identification of Nitrogen-Responsive Genes and Gene Expression Analysis
The Blast software was used to search the spinach genes against the spinach genomics database (http://www.spinachbase.org/cgi-bin/spinach/index.cgi) [60] and National Center for Biotechnology Information (NCBI) with the corresponding 10 functionally characterized Arabidopsis gene sequences (AtNRT1.1, AtNRT1.2, AtNRT1.3, AtNRT1.4, AtNRT1.5, AtNRT1.6, AtNRT1.7, AtNRT1.8, AtNRT1.9 and AtNRT2.1) as the initial query. The information of the identified genes is listed in Table 3. To improve the quality of primers and prevent mismatch and DNA contamination, the specific primers were obtained from the non-conserved regions and span an exon-exon junction. Gene specific primers designed by Prime 5 are listed in Table 4.
Total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Two μg of DNase-treated RNA was used to synthesize cDNA using the PrimeScript™ RT Master Mix (TAKARA, Tokyo, Japan). qRT-PCR was performed using a SYBR® Premix Ex Taq™ II (TAKARA, Tokyo, Japan) on a ABI 7500 Real-Time PCR System (Applied Biosystems, Foster, CA, USA). The initial denaturing time was 10 min, which was followed by 35 PCR cycles consisting of 94 °C for 0 s, 63 °C for 5 s and 72 °C for 10 s. A melting curve was run after the PCR cycles, which was followed by a cooling step. The relative quantitation of gene expression of qRT-PCR was measured via the 2−ΔΔCt method [61], with 18s rRNA used as the endogenous reference gene. Twice the ΔCT value of SoNRT1.1a in leaves of So10 under low nitrate condition was used as the control group. Data were presented as log10 transformation.
4.5. Statistical Analysis
Significant differences of physiological indices between different samples were calculated using a multiple factor analysis of variance with a LSD test (at a significance level of p < 0.05). Repeated measures analysis of variance was used to test for the effects of spinach genotype on gene expression (p < 0.05). The data are expressed as the mean and the standard deviation (mean ± SD, n = 3). The relationship between nitrate content and gene expression was determined using the Pearson’s correlation analysis. Statistical calculations were performed using SPSS software version 13.0 (IBM, New York, NY, USA).
5. Conclusions
The nitrate accumulation differences between two tested spinach genotypes may be due to their differential capacities to uptake and transport nitrate from the root to shoot. The highly expressed SoNRT genes found in So18, especially SoNRT1.4 and SoNRT1.5, may facilitate nitrate transport to petioles and contribute to excessive nitrate accumulation in So18. The SoNRT1.3 seems to play a negative role in shoot nitrate accumulation and thus, may be a potential target for reducing spinach nitrate content. Other SoNRTs, which may fulfill different physiological functions from their Arabidopsis homologs, may also be involved in the early nitrate response in spinach, although more evidence is needed to prove their roles in nitrate accumulation. The identification of these SoNRT genes is only the first step in providing comprehensive insights into nitrate accumulation mechanisms in spinach. As a first attempt, the present analysis will be useful for further investigations for understanding the roles of SoNRTs in nitrate accumulation in spinach.
Supplementary Materials
The following are available online. Table S1: The relative expression levels of nitrate accumulation related genes under 0.5 and 15 mmol·L−1 nitrate treatment conditions; Figure S1: The ratios of the relative gene expression under high to low nitrate treatments.
Author Contributions
X.W. and Q.W. designed this work. X.W., C.X. and X.C. performed the experiments; X.W. wrote the manuscript. S.W. and S.D. analyzed the data and helped edit the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China [31601744], Natural Science Foundation of Shanghai [15ZR1431300], Shanghai Engineering Research Center of Plant Germplasm Resources [17DZ2252700] and Development and Collaborative Innovation Center of Shanghai [ZF1205].
Acknowledgments
The authors wish to thank Jianjian Zhou and Tianli Song for helping with the hydropnics.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Walker, R. Nitrates, nitrites and N-nitrosocompounds: A review of the occurrence in food and diet and the toxicological implications. Food Addit. Contam. 1990, 7, 717–768. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. J. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- Anjana, U.; Iqbal, M. Factors responsible for nitrate accumulation: A review. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 533–549. ISBN 978-90-481-2665-1. [Google Scholar]
- Granstedt, R.C.; Huffaker, R.C. Identification of the leaf vacuole as a major nitrate storage pool. Plant Physiol. 1982, 70, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Liu, Q.; Gu, J.; Gong, J.; Guan, C.; Lepo, J.E.; Rong, X.; Song, H.; Zhang, Z. V-ATPase and V-PPase at the tonoplast affect NO3- content in Brassica napus by controlling distribution of NO3− between the cytoplasm and vacuole. J. Plant Growth Regul. 2015, 34, 22–34. [Google Scholar] [CrossRef]
- Anjana, S.U.; Iqbal, M. Nitrate accumulation in plants, factors affecting the and human health implications. A review. Agron. Sustain. Dev. 2007, 27, 45–57. [Google Scholar]
- Mobini, M.; Khoshgoftarmanesh, A.H.; Ghasemi, S. The effect of partial replacement of nitrate with arginine, histidine, and a mixture of amino acids extracted from blood powder on yield and nitrate accumulation in onion bulb. Sci. Hortic. Amsterdam 2014, 176, 232–237. [Google Scholar] [CrossRef]
- Marsic, N.K.; Osvald, J. The influence of different concentration of nitrogen in nutrient solution on plant growth and nitrate accumulation in aeroponically grown lettuce (Lactuca sativa L.). Agrochimica 2002, 46, 56–65. [Google Scholar]
- Onyango, C.M.; Harbinson, J.; Imungi, J.K.; Shibairo, S.S.; van Kooten, O. Influence of organic and mineral fertilization on germination, leaf nitrogen, nitrate accumulation and yield of vegetable amaranth. J. Plant Nutr. 2012, 35, 342–365. [Google Scholar] [CrossRef]
- Inal, A.; Tarakcioglu, C. Effects of nitrogen forms on growth, nitrate accumulation, membrane permeability, and nitrogen use efficiency of hydroponically grown bunch onion under boron deficiency and toxicity. J. Plant Nutr. 2001, 24, 1521–1534. [Google Scholar] [CrossRef]
- Irigoyen, I.; Lamsfus, C.; Aparicio-Tejo, P.; Muro, J. The influence of 3,4-dimethylpyrazole phosphate and dicyandiamide on reducing nitrate accumulation in spinach under Mediterranean conditions. J. Agric. Sci. 2006, 144, 555–562. [Google Scholar] [CrossRef]
- Ma, C.; Feng, S.; Huang, L.; Li, N.; Xu, X.; Zhou, B.; Jiao, K.; Xu, Q.; Li, R.; Herbert, S.J. Exogenous salicylic acid prevents nitrogen dioxide-induced oxidative injury and nitrate accumulation in Brassica campestris L. ssp. chinensis seedlings. J. Hortic. Sci. Biotechnol. 2010, 85, 215–218. [Google Scholar] [CrossRef]
- Strzetelski, P.; Smolen, S.; Rozek, S.; Sady, W. The effect of diverse iodine fertilization on nitrate accumulation and content of selected compounds in radish plants (Raphanus sativus L.). Acta Sci. Pol. Hortic. 2010, 9, 65–73. [Google Scholar]
- Liu, W.K.; Yang, Q.C. Effects of short-term treatment with various light intensities and hydroponic solutions on nitrate concentration of lettuce. Acta Agric. Scand. B-SP 2012, 62, 109–113. [Google Scholar] [CrossRef]
- Agüera, E.; Ruano, D.; Cabello, P.; de la Haba, P. Impact of atmospheric CO2 on growth, photosynthesis and nitrogen metabolism in cucumber (Cucumis sativus L.) plants. J. Plant Physiol. 2006, 163, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.; Charoenprasert, S.; Mitchell, A.E. Effect of organic and conventional cropping systems on ascorbic acid, vitamin C, flavonoids, nitrate, and oxalate in 27 varieties of spinach (Spinacia oleracea L.). J. Agric. Food Chem. 2012, 60, 3144–3150. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Hsu, P.K.; Tsay, Y.F. Uptake, allocation and signaling of nitrate. Trends Plant Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen acquisition by roots: physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Tsay, Y.F.; Chiu, C.C.; Tsai, C.B.; Ho, C.H.; Hsu, P.K. Nitrate transporters and peptide transporters. FEBS Lett. 2007, 581, 2290–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Wittgenstein, N.J.; Le, C.H.; Hawkins, B.J.; Ehlting, J. Evolutionary classification of ammonium, nitrate, and peptide transporters in land plants. BMC Evol. Biol. 2014, 14, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, H.; Euring, D.; Volmer, K.; Janz, D.; Polle, A. The nitrate transporter (NRT) gene family in poplar. PLoS ONE 2013, 8, e72126. [Google Scholar]
- Migocka, M.; Warzybok, A.; Kobus, G. The genomic organization and transcriptional pattern of genes encoding nitrate transporters 1 (NRT1) in cucumber. Plant Soil 2013, 364, 245–260. [Google Scholar] [CrossRef]
- Chiu, C.; Lin, C.; Hsia, A.; Su, R.; Lin, H.; Tsay, Y. Mutation of a nitrate transporter, AtNRT1: 4, results in a reduced petiole nitrate content and altered leaf development. Plant Cell Physiol. 2004, 45, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Taochy, C.; Gaillard, I.; Ipotesi, E.; Oomen, R.; Leonhardt, N.; Zimmermann, S.; Peltier, J.-B.; Szponarski, W.; Simonneau, T.; Sentenac, H.; et al. The Arabidopsis root stele transporter NPF2.3 contributes to nitrate translocation to shoots under salt stress. Plant J. 2015, 83, 466–479. [Google Scholar] [PubMed] [Green Version]
- Wu, T.; Qin, Z.; Fan, L.; Xue, C.; Zhou, X.; Xin, M.; Du, Y. Involvement of CsNRT1.7 in nitrate recycling during senescence in cucumber. J. Plant Nutr. Soil Sci. 2014, 177, 714–721. [Google Scholar]
- Zhao, S.; Ye, X.; Zhang, Y.; Zheng, J. The contribution of bnnrt1 and bnnrt2 to nitrate accumulation varied according to genotypes in Chinese cabbage. Afr. J. Biotechnol. 2010, 9, 4910–4917. [Google Scholar]
- Quaggiotti, S.; Ruperti, B.; Borsa, P.; Destro, T.; Malagoli, M. Expression of a putative high-affinity NO3− transporter and of an H+-ATPase in relation to whole plant nitrate transport physiology in two maize genotypes differently responsive to low nitrogen availability. J. Exp. Bot. 2003, 54, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- North, K.A.; Ehlting, B.; Koprivova, A.; Rennenberg, H.; Kopriva, S. Natural variation in Arabidopsis adaptation to growth at low nitrogen conditions. Plant Physiol. Biochem. 2009, 47, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Sun, S.; Jia, L.; Chen, W.; Shen, Q. The mechanism of nitrate accumulation in pakchoi [Brassica campestris L. ssp. Chinensis (L.)]. Plant Soil 2006, 282, 291–300. [Google Scholar] [CrossRef]
- Curtis, I.S.; Power, J.B.; de Laat, A.M.M.; Caboche, M.; Davey, M.R. Expression of a chimeric nitrate reductase gene in transgenic lettuce reduces nitrate in leaves. Plant Cell Rep. 1999, 18, 889–896. [Google Scholar] [CrossRef]
- Djennane, S.; Chauvin, J.-E.; Quilleré, I.; Meyer, C.; Chupeau, Y. Introduction and expression of a deregulated tobacco nitrate reductase gene in potato lead to highly reduced nitrate levels in transgenic tubers. Transgen. Res. 2002, 11, 175–184. [Google Scholar] [CrossRef]
- Kaminishi, A.; Kita, N. Seasonal change of nitrate and oxalate concentration in relation to the growth rate of spinach cultivars. Hortscience 2006, 41, 1589–1595. [Google Scholar]
- Gangolli, S.D.; van den Brandt, P.A.; Feron, V.J.; Janzowsky, C.; Koeman, J.H.; Speijers, G.J.; Spiegelhalder, B.; Walker, R.; Wisnok, J.S. Nitrate, nitrite and N-nitroso compounds. Eur. J. Pharmacol. 1994, 292, 1–38. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs. 2006. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32006R1881&rid=1 (accessed on 01.07.2018).
- Menard, C.; Heraud, F.; Volatier, J.L.; Leblanc, J.C. Assessment of dietary exposure of nitrate and nitrite in France. Food Addit. Contam. 2008, 25, 971–988. [Google Scholar] [Green Version]
- Thomson, B.M.; Nokes, C.J.; Cressey, P.J. Intake and risk assessment of nitrate and nitrite from New Zealand foods and drinking water. Food Addit. Contam. 2007, 24, 113–121. [Google Scholar]
- Merino, L.; Darnerud, P.O.; Edberg, U.; Åman, P.; Castillo, M.D.P. Levels of nitrate in Swedish lettuce and spinach over the past 10 years. Food Addit. Contam. 2006, 23, 1283–1289. [Google Scholar]
- Ysart, G.; Clifford, R.; Harrison, N. Monitoring for nitrate in UK-grown lettuce and spinach. Food Addit. Contam. 1999, 16, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Mor, F.; Sahindokuyucu, F.; Erdogan, N. Nitrate and nitrite contents of some vegetables consumed in south province of Turkey. J. Anim. Vet. Adv. 2010, 9, 2013–2016. [Google Scholar]
- Chung, S.Y.; Kim, J.S.; Kim, M.; Hong, M.K.; Lee, J.O.; Kim, C.M.; Song, I.S. Survey of nitrate and nitrite contents of vegetables grown in Korea. Food Addit. Contam. 2003, 20, 621–628. [Google Scholar]
- Zhong, W.; Hu, C.; Wang, M. Nitrate and nitrite in vegetables from north China: Content and intake. Food Addit. Contam. 2002, 19, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Olday, F.C.; Barker, A.V.; Maynard, D.N. A physiological basis for different patterns of nitrate accumulation in two spinach cultivars. J. Am. Soc. Hortic. Sci. 1976, 101, 217–219. [Google Scholar]
- Lin, S.H.; Kuo, H.F.; Canivenc, G.; Lin, C.S.; Lepetit, M.; Hsu, P.K.; Tillard, P.; Lin, H.L.; Wang, Y.Y.; Tsai, C.B.; et al. Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root-to-shoot nitrate transport. Plant Cell 2008, 20, 2514–2528. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.C.; Lin, C.S.; Hsu, P.K.; Lin, S.H.; Tsay, Y.F. The Arabidopsis nitrate transporter NRT1.7, expressed in phloem, is responsible for source-to-sink remobilization of nitrate. Plant Cell 2009, 21, 2750–2761. [Google Scholar] [CrossRef] [PubMed]
- Almagro, A.; Lin, S.H.; Tsay, Y.F. Characterization of the Arabidopsis nitrate transporter NRT1. 6 reveals a role of nitrate in early embryo development. Plant Cell 2008, 20, 3289–3299. [Google Scholar] [PubMed]
- Wang, Y.Y.; Tsay, Y.F. Arabidopsis nitrate transporter NRT1.9 is important in phloem nitrate transport. Plant Cell 2011, 23, 1945–1957. [Google Scholar] [PubMed]
- Buchner, P.; Hawkesford, M.J. Complex phylogeny and gene expression patterns of members of the nitrate transporter 1/peptide transporter family (NPF) in wheat. J. Exp. Bot. 2014, 65, 5697–5710. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.R.N.; Imai, A.; Tabata, R.; Shigenobu, S.; Yamaguchi, K.; Yamada, M.; Hasebe, M.; Sawa, S.; Motose, H.; Takahashi, T. Polyamine resistance is increased by mutations in a nitrate transporter gene NRT1.3 (AtNPF6.4) in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 834. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Vidmar, J.J.; Glass, A.D.M. Regulation of NRT1 and NRT2 gene families of Arabidopsis thaliana: Responses to nitrate provision. Plant Cell Physiol. 2003, 44, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Tischner, R. Nitrate uptake and reduction in higher and lower plants. Plant Cell Environ. 2000, 23, 1005–1024. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Xiang, F.; Zhong, M.; Zhou, L.; Liu, H.; Li, S.; Wang, X. Transcriptome and metabolite analysis identifies nitrogen utilization genes in tea plant (Camellia sinensis). Sci. Rep. UK 2017, 7, e1693. [Google Scholar] [CrossRef] [PubMed]
- Garnica, M.; Houdusse, F.; Zamarreno, A.M.; Garcia-Mina, J.M. Nitrate modifies the assimilation pattern of ammonium and urea in wheat seedlings. J. Sci. Food Agric. 2010, 90, 357–369. [Google Scholar]
- Wu, Y.; Zhang, W.; Xu, L.; Wang, Y.; Zhu, X.; Li, C.; Liu, L. Isolation and molecular characterization of nitrite reductase (RsNiR) gene under nitrate treatments in radish. Sci Hortic-Amsterdam. 2015, 193, 276–285. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, W.; Yang, Y.; Li, Z.; Li, N.; Qi, S.; Crawford, N.M.; Wang, Y. The Arabidopsis NLP7 gene regulates nitrate signaling via NRT1.1-dependent pathway in the presence of ammonium. Sci. Rep. UK 2018, 8, 1487. [Google Scholar] [CrossRef]
- Castaings, L.; Marchive, C.; Meyer, C.; Krapp, A. Nitrogen signalling in Arabidopsis: how to obtain insights into a complex signalling network. J. Exp. Bot. 2011, 62, 1391–1397. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Gordonweeks, R.; Shen, Q.; Miller, A.J. Glutamine transport and feedback regulation of nitrate reductase activity in barley roots leads to changes in cytosolic nitrate pools. J. Exp. Bot. 2006, 57, 1333–1340. [Google Scholar] [Green Version]
- Orsel, M.; Eulenburg, K.; Krapp, A.; Daniel-Vedele, F. Disruption of the nitrate transporter genes AtNRT2.1 and AtNRT2.2 restricts growth at low external nitrate concentration. Planta 2004, 219, 714–721. [Google Scholar] [PubMed]
- Tang, Y.; Sun, X.; Hu, C.; Tan, Q.; Zhao, X. Genotypic differences in nitrate uptake, translocation and assimilation of two Chinese cabbage cultivars [Brassica campestris L. ssp. Chinensis (L.)]. Plant Physiol. Biochem. 2013, 70, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S. Principles and Techniques of Plant Pysiological Biochemical Experiment, 3rd ed.; Higher Education Press: Beijing, China, 2015; pp. 129–130. ISBN 978-7-04-039646-1. [Google Scholar]
- Xu, C.; Jiao, C.; Sun, H.; Cai, X.; Wang, X.; Ge, C.; Zheng, Y.; Liu, W.; Sun, X.; Xu, Y. Draft genome of spinach and transcriptome diversity of 120 Spinacia accessions. Nat. Commun. 2017, 8, e15275. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available |
Figure 1.
Fresh weights (A) and total nitrogen contents (B) of two spinach varieties (S10 and S18) after 12 h of nitrate treatment. L-So10, the leaf blades of S10; L-So18, the leaf blades of So18; P-So10, the petioles of So10; P-So18, the petioles of So18; R-So10, the roots of So10; and R-So18, the roots of So18. Different letters in columns indicate significant difference between the mean values at p < 0.05 by the LSD test (n = 3). Values are provided as mean ± SD.
Figure 1.
Fresh weights (A) and total nitrogen contents (B) of two spinach varieties (S10 and S18) after 12 h of nitrate treatment. L-So10, the leaf blades of S10; L-So18, the leaf blades of So18; P-So10, the petioles of So10; P-So18, the petioles of So18; R-So10, the roots of So10; and R-So18, the roots of So18. Different letters in columns indicate significant difference between the mean values at p < 0.05 by the LSD test (n = 3). Values are provided as mean ± SD.
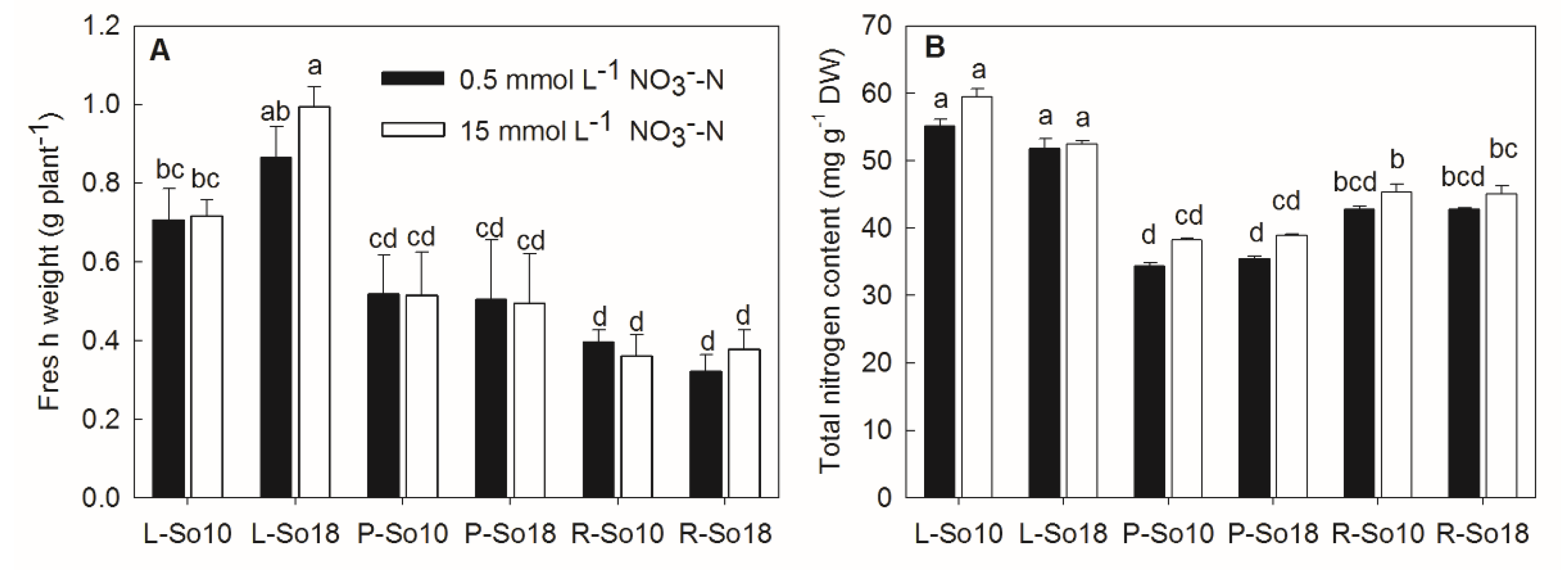
Figure 2.
Nitrate contents in tissues of two spinach varieties under low (0.5 mmol·L−1) and high (15 mmol·L−1) nitrate treatments. LN-So18, So18 treated with low nitrate concentration; LN-So10, So10 treated with low nitrate concentration; HN-So18, So18 treated with high nitrate concentration; and HN-So10, So10 treated with high nitrate concentration.
Figure 2.
Nitrate contents in tissues of two spinach varieties under low (0.5 mmol·L−1) and high (15 mmol·L−1) nitrate treatments. LN-So18, So18 treated with low nitrate concentration; LN-So10, So10 treated with low nitrate concentration; HN-So18, So18 treated with high nitrate concentration; and HN-So10, So10 treated with high nitrate concentration.
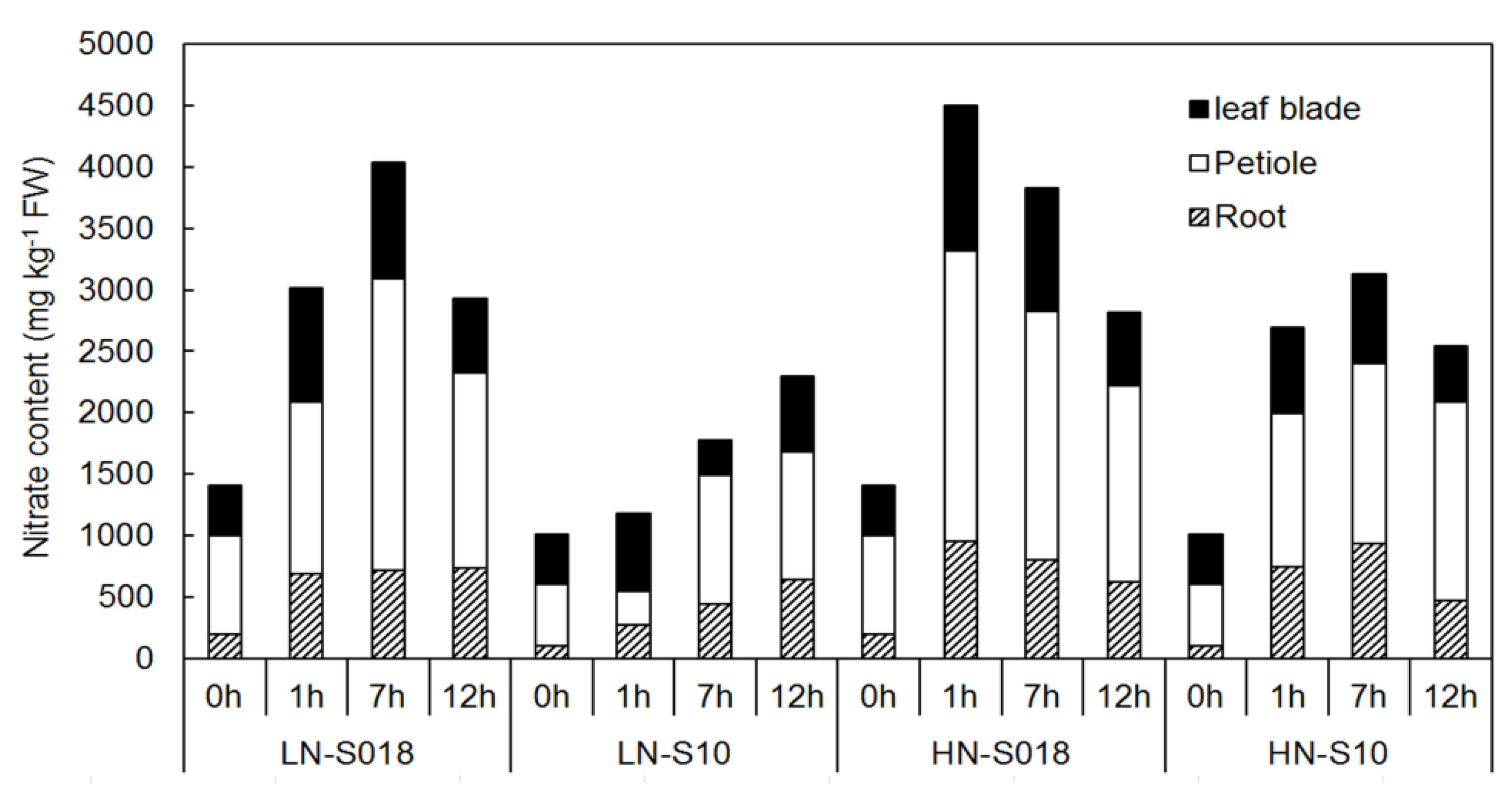
Figure 3.
15NO3−-N uptake rate (A) and 15NO3−-N excess (net NO3−-N uptake amount) (B) of two spinach varieties under low (0.5 mmol·L−1) and high (15 mmol·L−1) nitrate treatments. Values are provided as mean ± SD, n = 3. LN-So18, So18 treated with low nitrate concentration; LN-So10, So10 treated with low nitrate concentration; HN-So18, So18 treated with high nitrate concentration; and HN-So10, So10 treated with high nitrate concentration.
Figure 3.
15NO3−-N uptake rate (A) and 15NO3−-N excess (net NO3−-N uptake amount) (B) of two spinach varieties under low (0.5 mmol·L−1) and high (15 mmol·L−1) nitrate treatments. Values are provided as mean ± SD, n = 3. LN-So18, So18 treated with low nitrate concentration; LN-So10, So10 treated with low nitrate concentration; HN-So18, So18 treated with high nitrate concentration; and HN-So10, So10 treated with high nitrate concentration.

Figure 4.
The activities of nitrate reductase (NR) (A) and glutamine synthetase (GS) (B) in shoots of two spinach varieties under low (0.5 mmol·L−1) and high nitrate (15 mmol·L−1) treatments. Values are represented as mean ± SD, n = 3. LN-So18, So18 treated with low nitrate concentration; LN-So10, So10 treated with low nitrate concentration; HN-So18, So18 treated with high nitrate concentration; and HN-So10, So10 treated with high nitrate concentration.
Figure 4.
The activities of nitrate reductase (NR) (A) and glutamine synthetase (GS) (B) in shoots of two spinach varieties under low (0.5 mmol·L−1) and high nitrate (15 mmol·L−1) treatments. Values are represented as mean ± SD, n = 3. LN-So18, So18 treated with low nitrate concentration; LN-So10, So10 treated with low nitrate concentration; HN-So18, So18 treated with high nitrate concentration; and HN-So10, So10 treated with high nitrate concentration.
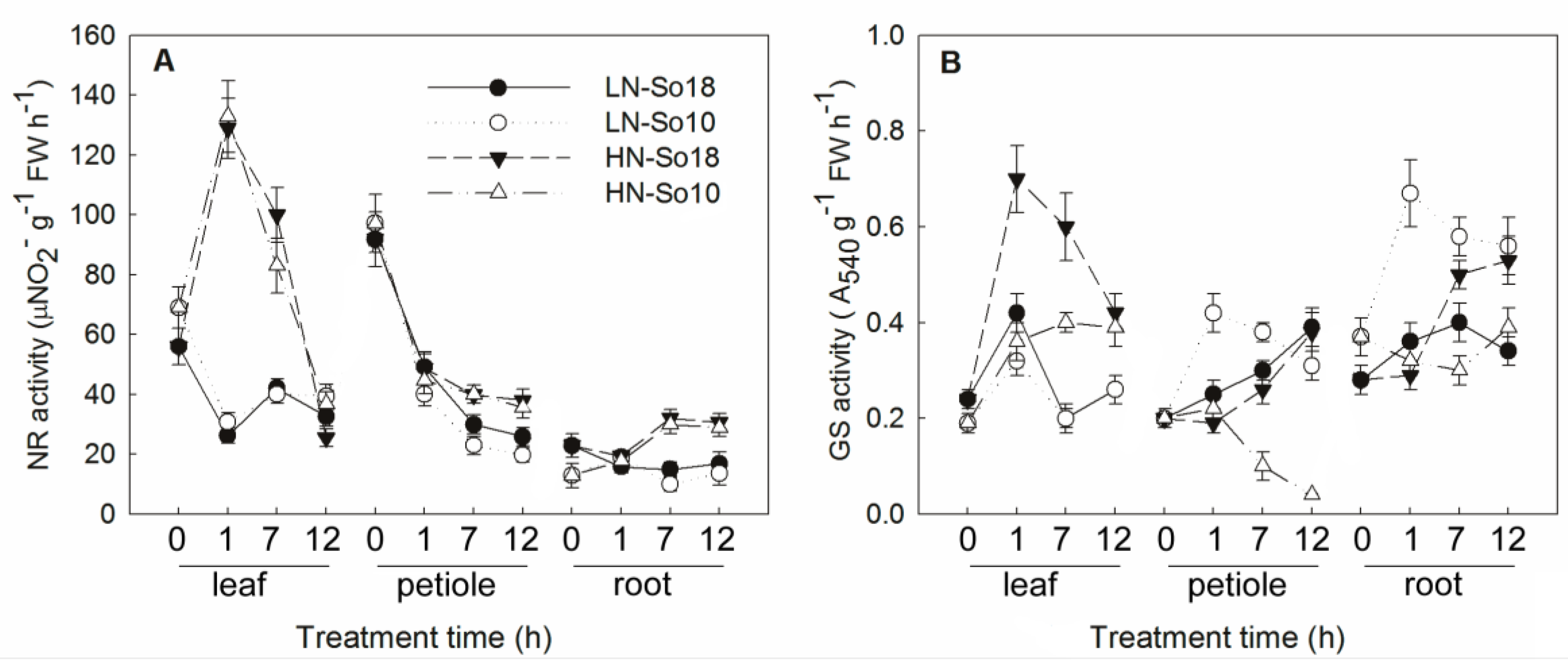
Figure 5.
Expression profiles of NRTs genes in tissues of two spinach varieties under two nitrate concentration treatments. S10, the spinach So10 variety; S18, the spinach So18 variety; B, leaf blade; P, petiole; R, root; L, low nitrate (0.5 mmol·L−1) treatment; H, high nitrate (15 mmol·L−1) treatment; 1, 1 h; 7, 7 h; and 12, 12 h.
Figure 5.
Expression profiles of NRTs genes in tissues of two spinach varieties under two nitrate concentration treatments. S10, the spinach So10 variety; S18, the spinach So18 variety; B, leaf blade; P, petiole; R, root; L, low nitrate (0.5 mmol·L−1) treatment; H, high nitrate (15 mmol·L−1) treatment; 1, 1 h; 7, 7 h; and 12, 12 h.
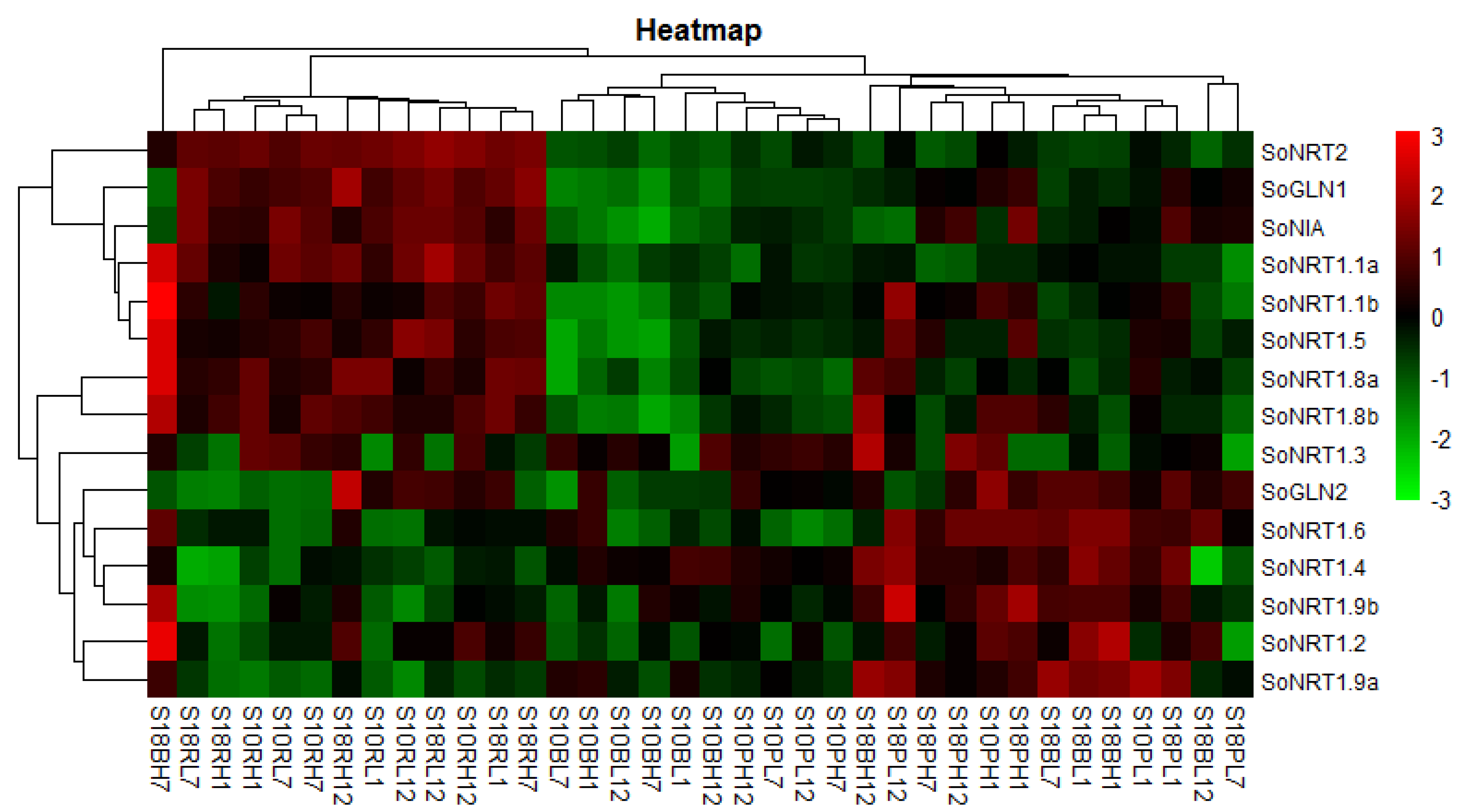
Figure 6.
Expression analysis of genes encoding nitrate transporters in leaf blades, petioles and roots of two spinach varieties under two concentrations (0.5 and 15 mmol·L−1) of nitrate treatments by qRT-PCR. All values represent the averages of three replicates (see Supplementary Table S1 for details). LN-So18, So18 treated with low nitrate concentration; LN-So10, So10 treated with low nitrate concentration; HN-So18, So18 treated with high nitrate concentration; and HN-So10, So10 treated with high nitrate concentration.
Figure 6.
Expression analysis of genes encoding nitrate transporters in leaf blades, petioles and roots of two spinach varieties under two concentrations (0.5 and 15 mmol·L−1) of nitrate treatments by qRT-PCR. All values represent the averages of three replicates (see Supplementary Table S1 for details). LN-So18, So18 treated with low nitrate concentration; LN-So10, So10 treated with low nitrate concentration; HN-So18, So18 treated with high nitrate concentration; and HN-So10, So10 treated with high nitrate concentration.

Figure 7.
Expression analysis of nitrogen assimilation genes in leaf blades, petioles and roots of two spinach varieties under two concentrations (0.5 and 15 mmol·L−1) of nitrate treatments by qRT-PCR. All values represent the averages of three replicates (see Supplementary Table S1 for details). LN-So18, So18 treated with low nitrate concentration; LN-So10, So10 treated with low nitrate concentration; HN-So18, So18 treated with high nitrate concentration; and HN-So10, So10 treated with high nitrate concentration.
Figure 7.
Expression analysis of nitrogen assimilation genes in leaf blades, petioles and roots of two spinach varieties under two concentrations (0.5 and 15 mmol·L−1) of nitrate treatments by qRT-PCR. All values represent the averages of three replicates (see Supplementary Table S1 for details). LN-So18, So18 treated with low nitrate concentration; LN-So10, So10 treated with low nitrate concentration; HN-So18, So18 treated with high nitrate concentration; and HN-So10, So10 treated with high nitrate concentration.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of repeated measures ANOVA for the effects of varieties on gene expression.
| Nitrate Concentration Treatment | Gene Symbol | Leaf blade | Petiole | Root | |||
|---|---|---|---|---|---|---|---|
| F Value | P Value | F Value | P Value | F Value | P Value | ||
| 0.5 mmol L−1 | SoNRT1.1a | 65.03 | 0.001 | 70.66 | 0.001 | 1.172 | 0.340 |
| SoNRT1.1b | 165.68 | 0.000 | 17.38 | 0.014 | 32.42 | 0.005 | |
| SoNRT1.2 | 218.12 | 0.000 | 4.42 | 0.103 | 96.52 | 0.001 | |
| SoNRT1.3 | 34.63 | 0.004 | 676.55 | 0.000 | 345.14 | 0.000 | |
| SoNRT1.4 | 1.96 | 0.235 | 35.10 | 0.004 | 1.43 | 0.298 | |
| SoNRT 1.5 | 308.94 | 0.000 | 529.18 | 0.000 | 305.17 | 0.000 | |
| SoNRT1.6 | 1389.78 | 0.000 | 182.38 | 0.000 | 1.87 | 0.243 | |
| SoNRT1.8a | 393.56 | 0.000 | 0.17 | 0.702 | 25.67 | 0.007 | |
| SoNRT1.8b | 291.00 | 0.000 | 175.47 | 0.000 | 38.39 | 0.003 | |
| SoNRT1.9a | 18.23 | 0.013 | 296.94 | 0.000 | 176.73 | 0.000 | |
| SoNRT1.9b | 75.74 | 0.001 | 96.22 | 0.001 | 10.19 | 0.033 | |
| SoNRT2 | 1097.97 | 0.000 | 0.26 | 0.637 | 7.44 | 0.053 | |
| SoNIA | 182.52 | 0.000 | 71.58 | 0.001 | 0.03 | 0.879 | |
| SoGLN1 | 526.29 | 0.000 | 484.55 | 0.000 | 153.67 | 0.000 | |
| SoGLN2 | 1479.40 | 0.000 | 213.74 | 0.000 | 2.07 | 0.223 | |
| 15 mmol·L−1 | SoNRT1.1a | 1684.78 | 0.000 | 40.11 | 0.00 | 0.09 | 0.775 |
| SoNRT1.1b | 915.06 | 0.000 | 0.99 | 0.375 | 1.58 | 0.277 | |
| SoNRT1.2 | 351.99 | 0.000 | 1.33 | 0.313 | 69.87 | 0.001 | |
| SoNRT1.3 | 166.19 | 0.000 | 190.68 | 0.000 | 396.17 | 0.000 | |
| SoNRT1.4 | 191.83 | 0.000 | 39.81 | 0.003 | 7.76 | 0.050 | |
| SoNRT 1.5 | 4024.10 | 0.000 | 255.51 | 0.000 | 357.70 | 0.000 | |
| SoNRT1.6 | 2276.30 | 0.000 | 267.66 | 0.000 | 1.09 | 0.356 | |
| SoNRT1.8a | 3253.03 | 0.000 | 9.88 | 0.035 | 88.67 | 0.001 | |
| SoNRT1.8b | 1447.21 | 0.000 | 7.51 | 0.052 | 0.23 | 0.657 | |
| SoNRT1.9a | 210.99 | 0.000 | 106.84 | 0.000 | 85.43 | 0.001 | |
| SoNRT1.9b | 33.88 | 0.004 | 16.43 | 0.015 | 156.65 | 0.000 | |
| SoNRT2 | 1431.02 | 0.000 | 136.43 | 0.000 | 27.52 | 0.006 | |
| SoNIA | 452.08 | 0.000 | 219.11 | 0.000 | 0.01 | 0.921 | |
| SoGLN1 | 493.49 | 0.000 | 78.36 | 0.001 | 475.91 | 0.000 | |
| SoGLN2 | 556.74 | 0.000 | 0.65 | 0.467 | 3329.69 | 0.000 | |
The statistically significant values in italics and regular font indicate that the corresponding transcripts were significantly more abundant in So10 and So18, respectively. All results were considered significant at the p < 0.05 level, n = 3.
Table 2.
Pearson correlation between nitrate contents and gene expressions.
| Low Nitrate Treatment | High Nitrate Treatment | |||||
|---|---|---|---|---|---|---|
| Leaf Blade | Petiole | Root | Leaf Blade | Petiole | Root | |
| SoNRT1.1a | −0.24 | −0.40 | 0.51 | 0.53 | −0.10 | −0.49 |
| SoNRT1.1b | 0.40 | 0.01 | 0.56 | 0.53 | 0.00 | −0.51 |
| SoNRT1.2 | 0.39 | 0.06 | 0.84 * | 0.78 | 0.08 | −0.69 |
| SoNRT1.3 | 0.00 | −0.15 | 0.33 | −0.75 | −0.93 ** | −0.54 |
| SoNRT1.4 | 0.06 | −0.05 | 0.43 | −0.07 | 0.84 * | −0.51 |
| SoNRT1.5 | 0.29 | 0.00 | 0.82 * | 0.28 | 0.91 * | 0.17 |
| SoNRT1.8a | 0.24 | −0.29 | −0.21 | 0.13 | 0.11 | 0.06 |
| SoNRT1.8b | −0.29 | −0.47 | 0.17 | 0.07 | 0.05 | −0.05 |
| SoNRT1.6 | −0.09 | −0.03 | 0.39 | 0.79 | 0.21 | −0.59 |
| SoNRT1.9a | −0.28 | −0.38 | 0.05 | 0.42 | 0.47 | −0.51 |
| SoNRT1.9b | 0.19 | 0.20 | 0.32 | 0.69 | 0.28 | −0.59 |
| SoNRT2 | −0.11 | −0.26 | 0.61 | 0.56 | −0.32 | −0.62 |
| SoNIA | 0.10 | −0.02 | −0.21 | 0.47 | 0.76 | 0.02 |
| SoGLN1 | 0.51 | 0.18 | 0.36 | 0.26 | 0.37 | −0.17 |
| SoGLN2 | 0.29 | −0.08 | 0.58 | 0.16 | −0.45 | −0.77 |
* indicates correlation is significant at the 0.05 level; ** indicates correlation is significant at the 0.01 level.
Table 3.
Genes involved in nitrate transport and assimilation identified in spinach.
| Gene | Ortholog Locus | Spinach Source Gene | Spinach Gene Symbol | Position of Predicted Genes | Length of ORF | Length of Protein |
|---|---|---|---|---|---|---|
| AtNRT1.1 | AT1G12110.1 | Spo26413 | SoNRT1.1a | 24047-30641 | 1782 | 593 aa |
| AT1G12110.1 | Spo11551 | SoNRT1.1b | 27689-32901 | 1230 | 409 aa | |
| AtNRT1.2 | AT1G69850.1 | Spo24381 | SoNRT1.2 | 9278773-9283346 | 1767 | 588 aa |
| AtNRT1.3 | AT3G21670.1 | Spo18484 | SoNRT1.3 | 32544817-32547940 | 1737 | 578 aa |
| AtNRT1.4 | AT2G26690.1 | Spo05548 | SoNRT1.4 | 370979-375454 | 1755 | 584 aa |
| AtNRT1.5 | AT4G21680.1 | Spo23405 | SoNRT 1.8 | 43590162-43598583 | 1800 | 599 aa |
| AtNRT1.6/1.71 | AT1G27080.1 | Spo25744 | SoNRT1.6 | 39077918-39086543 | 1875 | 624 aa |
| AtNRT1.8 | AT1G32450.1 | Spo00021 | SoNRT1.5a | 111335-115169 | 1752 | 583 aa |
| AT1G32450.1 | Spo00022 | SoNRT1.5b | 135693-137789 | 1773 | 590 aa | |
| AtNRT1.9 | AT1G18880.1 | Spo20247 | SoNRT1.9a | 9835695-9840007 | 1764 | 587 aa |
| AT1G18880.1 | Spo20248 | SoNRT1.9b | 9830221-9833091 | 1776 | 591 aa | |
| AtNRT2.1 | AT1G08090 | Spo09966 | SoNRT2 | 44925216-44928954 | 1584 | 527 aa |
| AtNIA | AT1G77760 | Spo23607 | SoNIA | 51343436-51348754 | 867 | 288 aa |
| AtGLN1 | AT5G37600 | Spo17102 | SoGLN1 | 9174181-9181171 | 1785 | 594 aa |
| AtGLN2 | AT5G35630 | Spo06798 | SoGLN2 | 19807870-19834770 | 2664 | 887 aa |
1 Spo25744 is the ortholog of two AtNRTs (AtNRT1.6 and AtNRT1.7).
Table 4.
Specific primers for NRT, NIA and GLN in spinach.
| Symbol | Gene ID | Primer Sequence (5′→3′) | |
|---|---|---|---|
| Forward | Reverse | ||
| SoNRT1.1a | Spo26413 | TAAGACTGGCGGTTGG | GACAGAGCATGAAGGAGG |
| SoNRT1.1b | Spo11551 | GTGAGGCATGTGAAAGATTA | CCACCAAGTAGGCAGAGC |
| SoNRT1.2 | Spo24381 | TTGTGGAGGTGCTGGAGA | ACGGCTGATTTAGATGGTGAGA |
| SoNRT1.3 | Spo18484 | TTTGGAGGATTTGTCG | ATGTAAAGAGCAGCGTAT |
| SoNRT1.4 | Spo05548 | GATAGCCGTGAACCTTG | GCGAAAATTGCGACA |
| SoNRT1.5 | Spo23405 | CAATCAGGAAATGGAGGAC | TGTTATGGGACAAAGACG |
| SoNRT1.6 | Spo25744 | CCGTTTCCTCGTTCCAGC | ACCCTTCAGCAAGCCCTA |
| SoNRT1.8a | Spo00021 | CATCTATCGCCGTGTT | GCTTAGACTACTTGCCTCT |
| SoNRT1.8b | Spo00022 | TTTTCGGCGTAGGAGT | TGTGGCGATGTTTGGT |
| SoNRT1. 9a | Spo20247 | GGAGTCCCCTTACGAGTG | TCCTTCTGGGTTTATTTTG |
| SoNRT1.9b | Spo20248 | GTGAGTTGGAGCATCGG | CGTCAGGGGTTATTATCG |
| SoNRT2 | Spo09966 | TTTGCCTTCTCGGTCTC | CGCCGCGAATGTGGATAT |
| SoNIA | Spo23607 | CTTGCCAATTCTGAAGCT | TAGTCCAGGCGTTGATAG |
| SoGLN1 | Spo17102 | GGATATTTCGAGGACAGG | TGGTTTCCAAAGAAGGGT |
| SoGLN2 | Spo06798 | CGGAGAAGGGAATGAAA | CACGGATTGAGCAACCAC |
| 18SrRNA | Spo24625 | CCATAAACGATGCCGACCAG | AGCCTTGCGACCATACTCCC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, X.; Cai, X.; Xu, C.; Wang, S.; Dai, S.; Wang, Q. Nitrate Accumulation and Expression Patterns of Genes Involved in Nitrate Transport and Assimilation in Spinach. Molecules 2018, 23, 2231. https://doi.org/10.3390/molecules23092231
AMA Style
Wang X, Cai X, Xu C, Wang S, Dai S, Wang Q. Nitrate Accumulation and Expression Patterns of Genes Involved in Nitrate Transport and Assimilation in Spinach. Molecules. 2018; 23(9):2231. https://doi.org/10.3390/molecules23092231
Chicago/Turabian StyleWang, Xiaoli, Xiaofeng Cai, Chenxi Xu, Shui Wang, Shaojun Dai, and Quanhua Wang. 2018. "Nitrate Accumulation and Expression Patterns of Genes Involved in Nitrate Transport and Assimilation in Spinach" Molecules 23, no. 9: 2231. https://doi.org/10.3390/molecules23092231