Exogenous Melatonin Improves Fruit Quality Features, Health Promoting Antioxidant Compounds and Yield Traits in Tomato Fruits under Acid Rain Stress

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
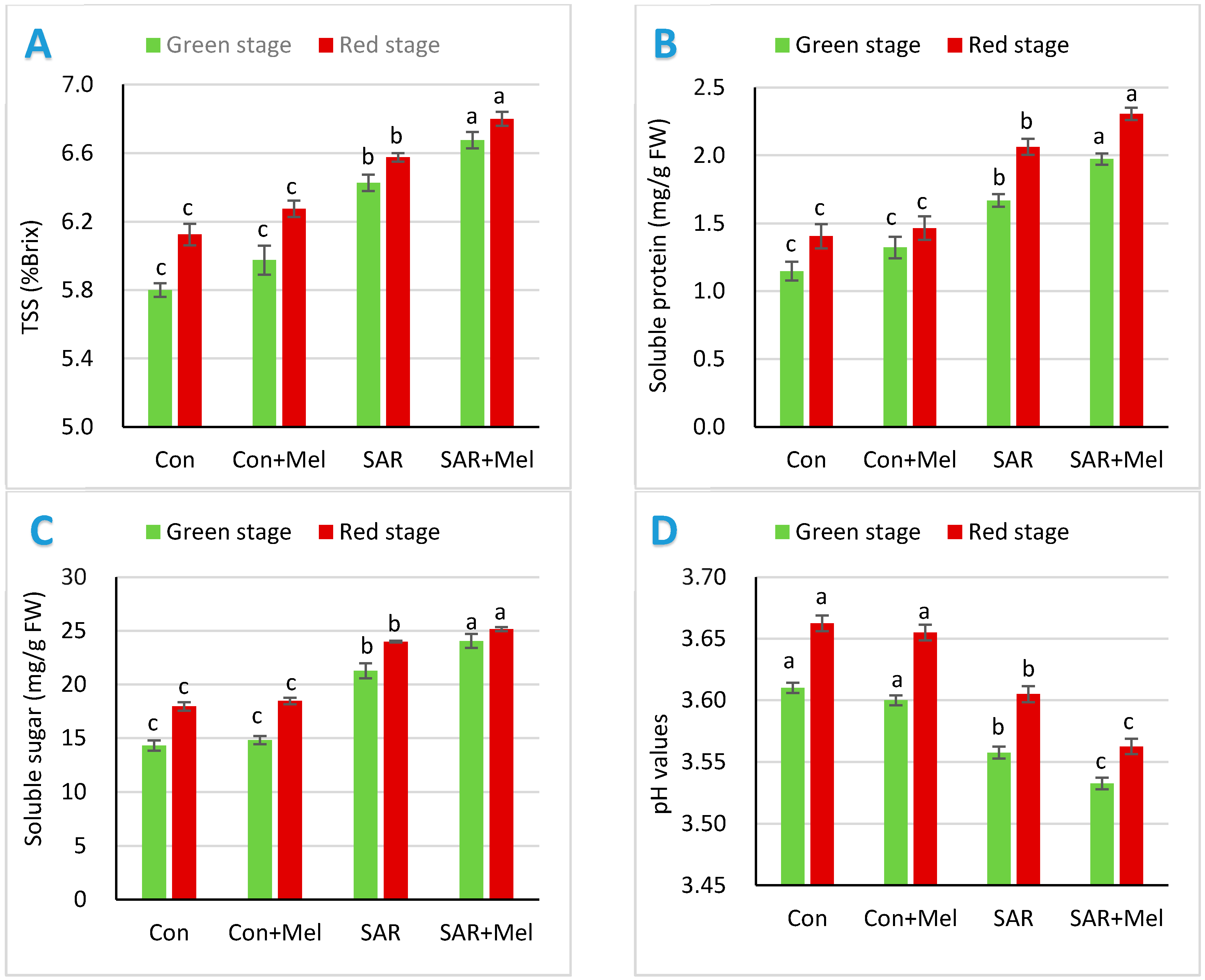
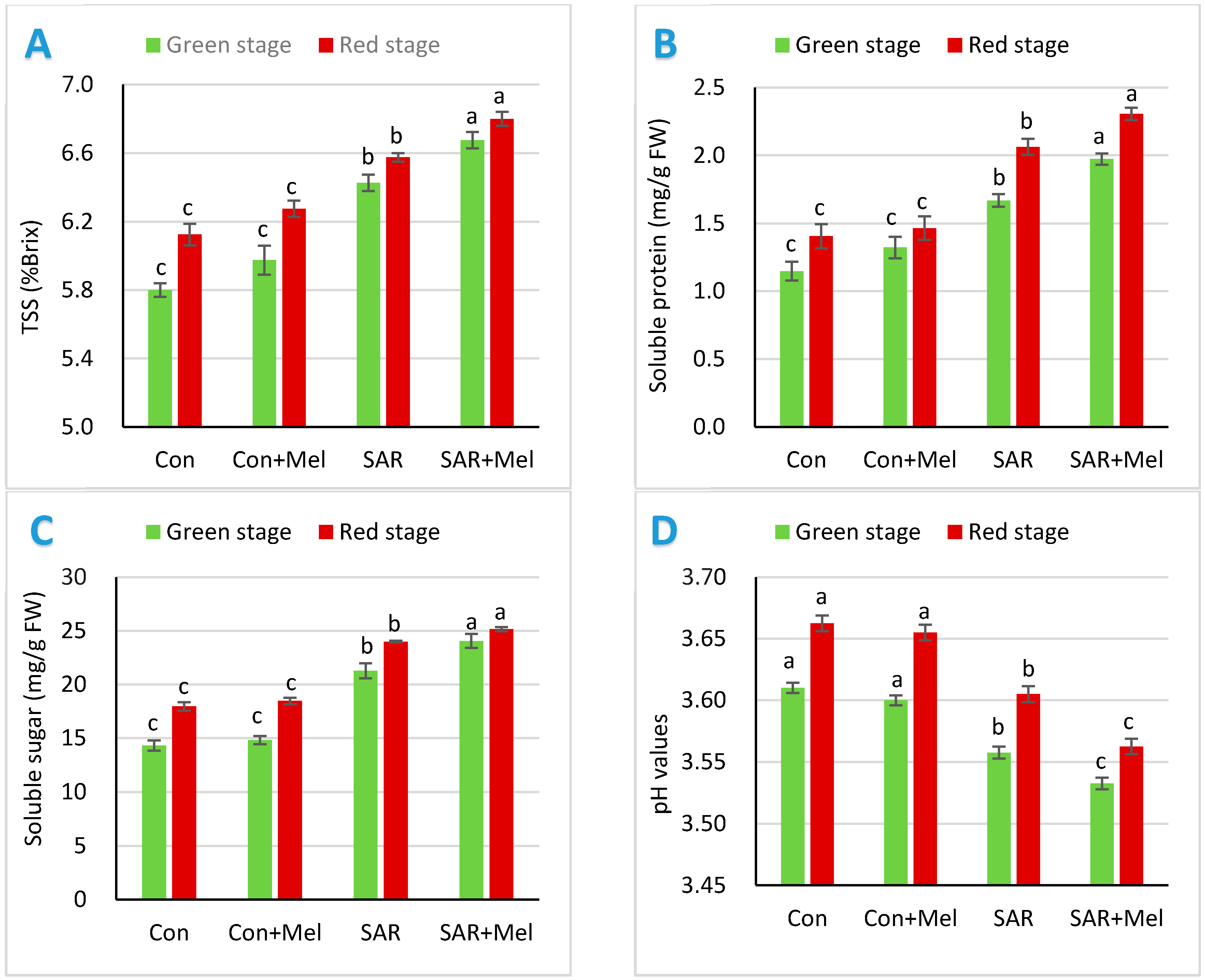
2.1. The Impact of Exogenous Melatonin on Fruit Qualities under SAR-Stress Condition
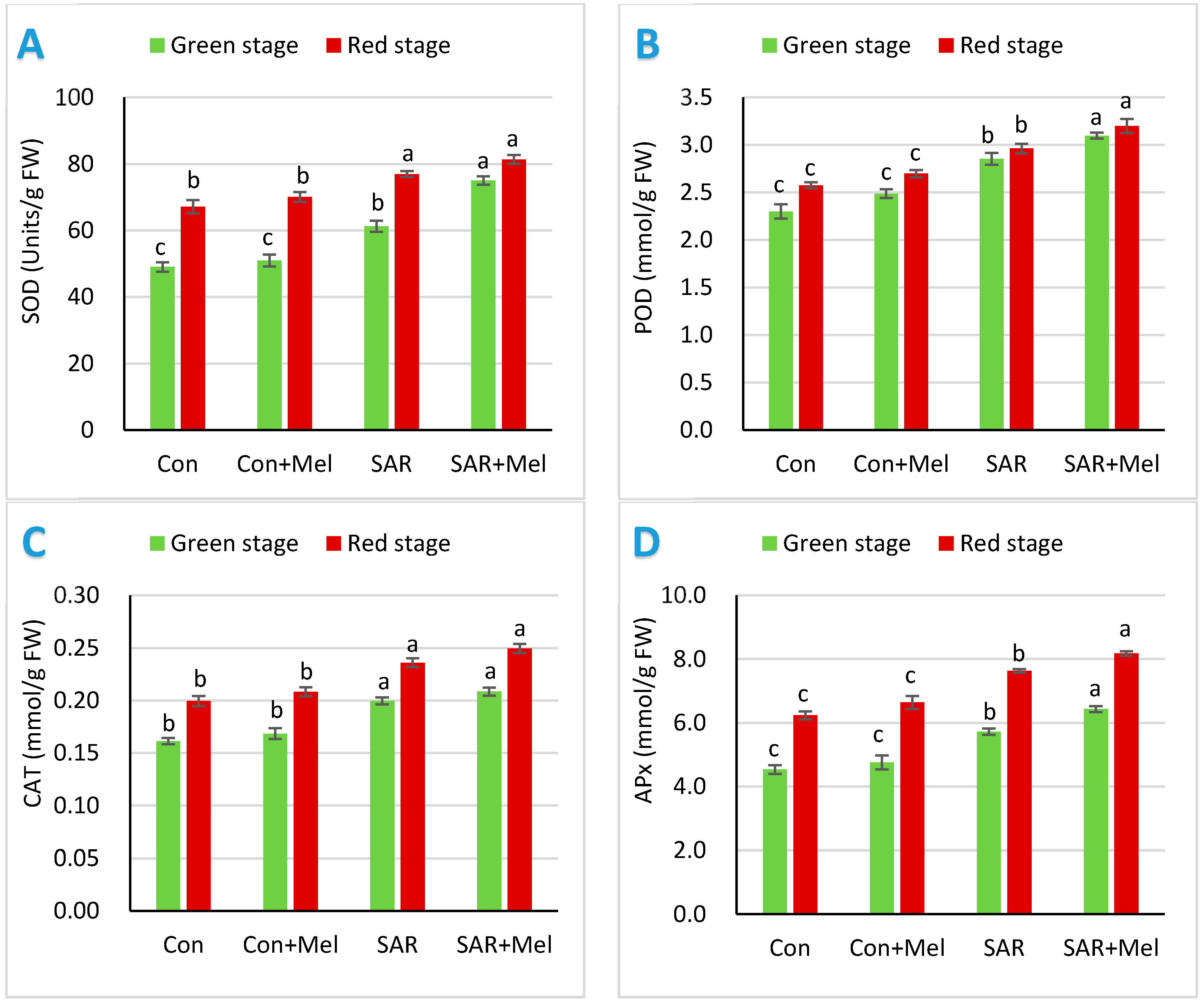
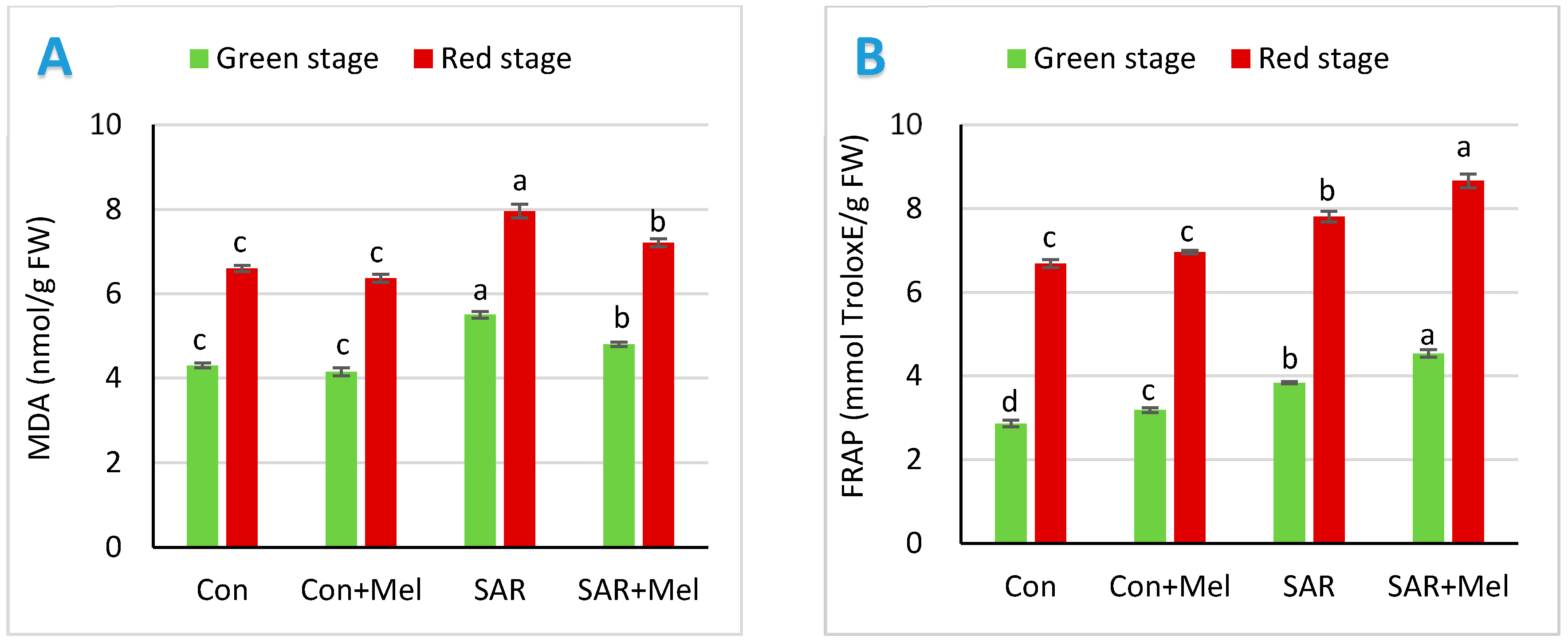
2.2. The Impact of Exogenous Melatonin on Antioxidant Enzymes in Fruits under SAR-Stress Condition
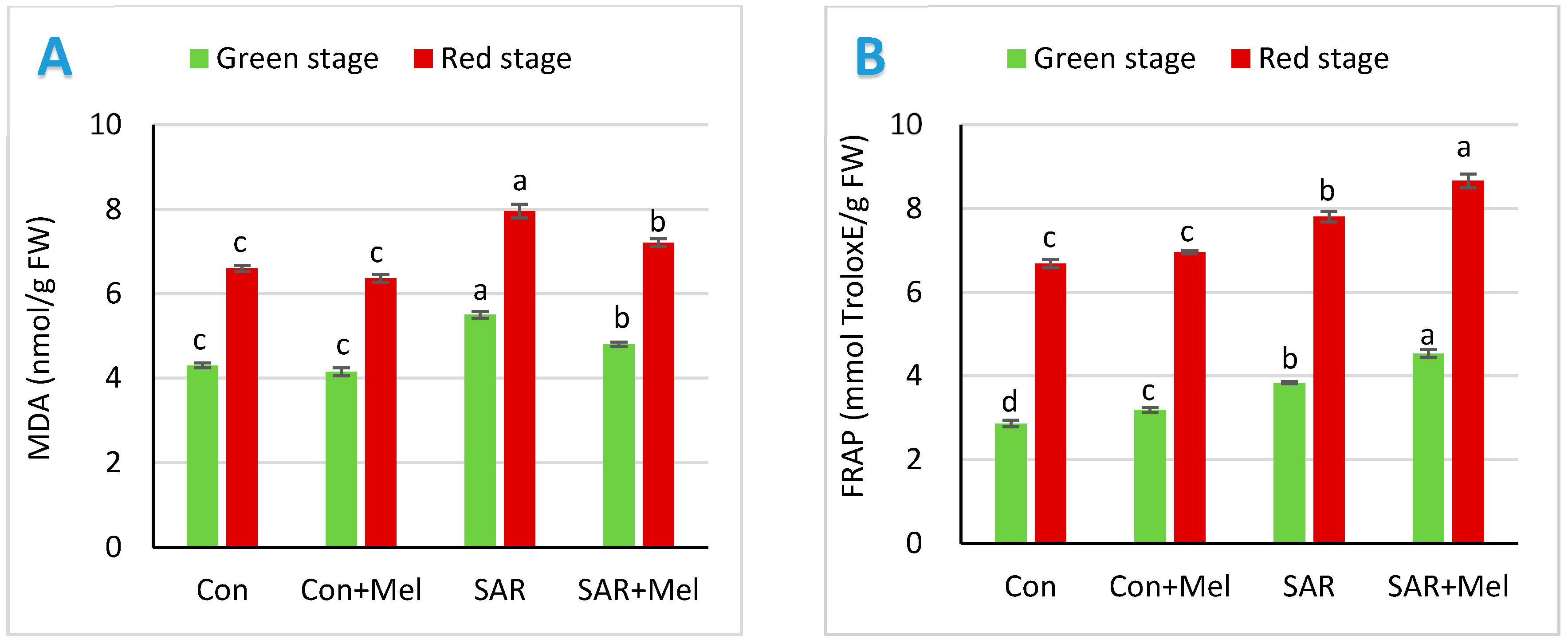
2.3. The Impact of Exogenous Melatonin on Oxidative Damage and Total Antioxidant Potential in Fruits under SAR-Stress Condition
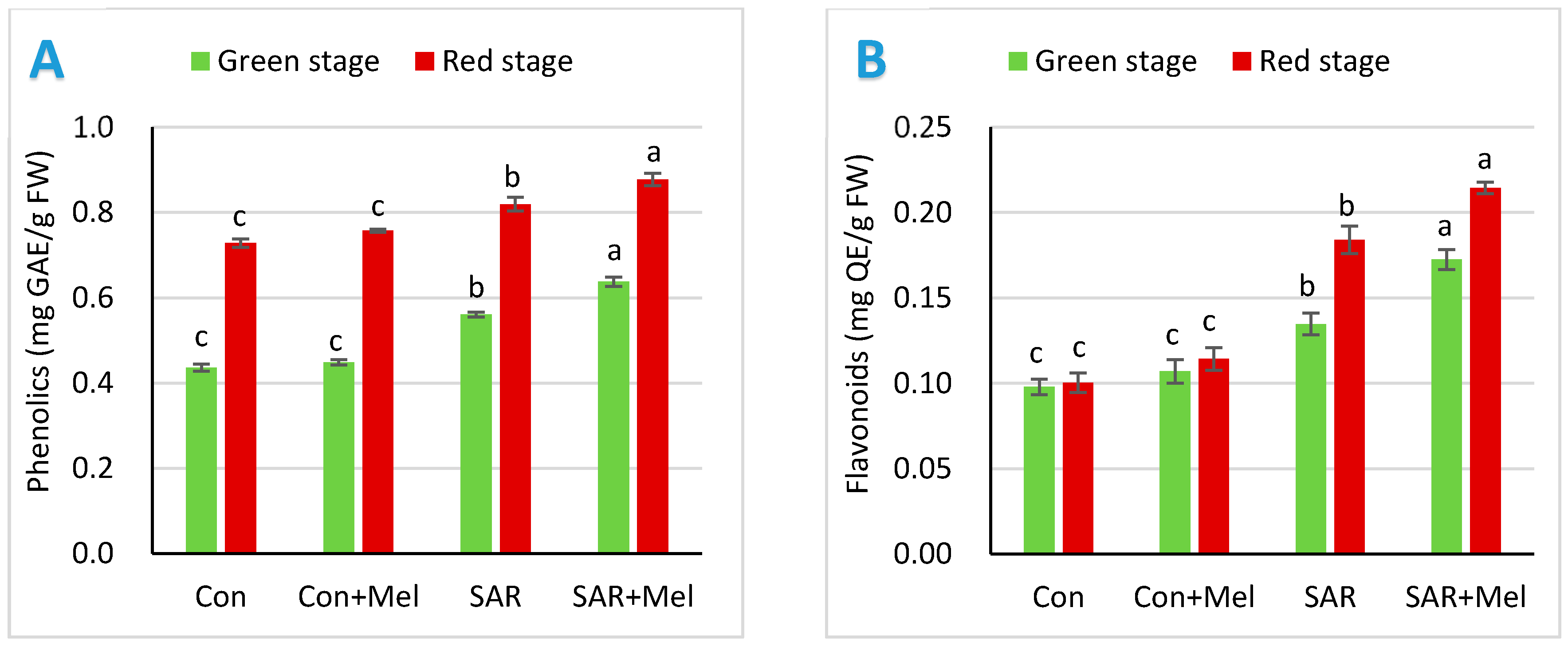
2.4. The Impact of Exogenous Melatonin on Phenolics and Flavonoids in Fruits under SAR-Stress Condition
2.5. The Impact of Exogenous Melatonin on Carotenoids in Fruits under SAR-Stress Condition
2.6. The Impact of Exogenous Melatonin on Yield Attributes of Tomatoes under SAR-Stress Condition
3. Materials and Methods
3.1. Plant Materials and Treatments
3.2. Measurement of Fruit Quality
3.3. Estimation of Enzymatic Antioxidant Activities, Oxidative Damage and Total Antioxidant Power
3.4. Quantification of Phenolics and Flavonoids Content
3.5. Estimation of Carotenoids
3.6. Quantification of Yield
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Mirmiran, P.; Noori, N.; Zavareh, M.B.; Azizi, F. Fruit and vegetable consumption and risk factors for cardiovascular disease. Metabolism 2009, 58, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Salas, P.; Morales-Soto, A.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Phenolic-compound-extraction systems for fruit and vegetable samples. Molecules 2010, 15, 8813–8826. [Google Scholar] [CrossRef] [PubMed]
- Buer, C.S.; Imin, N.; Djordjevic, M.A. Flavonoids: New roles for old molecules. J. Integr. Plant Biol. 2010, 52, 98–111. [Google Scholar] [CrossRef] [PubMed]
- De Pascual-Teresa, S.; Moreno, D.A.; García-Viguera, C. Flavanols and anthocyanins in cardiovascular health: A review of current evidence. Int. J. Mol. Sci. 2010, 11, 1679–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, J.P. The impact of fruit flavonoids on memory and cognition. Br. J. Nutr. 2010, 104, S40–S47. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Gu, L.; Prior, R.L.; McKay, S. Characterization of anthocyanins and proanthocyanidins in some cultivars of ribes, aronia, and sambucus and their antioxidant capacity. J. Agr. Food Chem. 2004, 52, 7846–7856. [Google Scholar] [CrossRef] [PubMed]
- Torres, C.A.; Davies, N.M.; YANez, J.A.; Andrews, P.K. Disposition of selected flavonoids in fruit tissues of various tomato (Lycopersicon esculentum Mill.) genotypes. J. Agr. Food Chem. 2005, 53, 9536–9543. [Google Scholar] [CrossRef] [PubMed]
- Erba, D.; Casiraghi, M.C.; Ribas-Agustí, A.; Cáceres, R.; Marfà, O.; Castellari, M. Nutritional value of tomatoes (Solanum lycopersicum L.) grown in greenhouse by different agronomic techniques. J. Food Compos. Anal. 2013, 31, 245–251. [Google Scholar] [CrossRef]
- FAO. Food and Agricultural Commodities Production for, Production of Tomato by Countries. 2014. Available online: http://faostat3.fao.org/home/E (accessed on 15 October 2017).
- Mu, L.; Fang, L. Changes in tomato fruit quality and antioxidant enzyme activities under deficit irrigation and fertilizer application in a solar greenhouse in Northwest China. Commun. Soil Sci. Plant Anal. 2016, 47, 1329–1341. [Google Scholar] [CrossRef]
- Duan, L.; Yu, Q.; Zhang, Q.; Wang, Z.; Pan, Y.; Larssen, T.; Tang, J.; Mulder, J. Acid deposition in asia: Emissions, deposition, and ecosystem effects. Atmos. Environ. 2016, 146, 55–69. [Google Scholar] [CrossRef]
- Bouwman, A.F.; Vuuren, D.P.V.; Derwent, R.G.; Posch, M. A global analysis of acidification and eutrophication of terrestrial ecosystems. Water Air Soil Pollut. 2002, 141, 349–382. [Google Scholar] [CrossRef]
- Kita, I.; Sato, T.; Kase, Y.; Mitropoulos, P. Neutral rains at athens, greece: A natural safeguard against acidification of rains. Sci. Total Environ. 2004, 327, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, W.; Gao, F. Biogeochemical effects of forest vegetation on acid precipitation-related water chemistry: A case study in Southwest China. J. Environ. Monit. 2010, 12, 1799–1806. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.; Liang, C.; Wang, L.; Hu, G.; Zhou, Q. Combined effects of lanthanumion and acid rain on growth, photosynthesis and chloroplast ultrastructure in soybean seedlings. Chemosphere 2011, 84, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Du, Y.; Wang, L.; Zhou, Q.; Huang, X.; Sun, Z. Combined effects of lanthanum (III) and acid rain on antioxidant enzyme system in soybean roots. PLoS ONE 2015, 10, e0134546. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, W.-H.; Liu, T.-W.; Wu, F.-H.; Zheng, H.-L. Photosynthetic and antioxidant responses of Liquidambar formosana and Schima superba seedlings to sulfuric-rich and nitric-rich simulated acid rain. Plant Physiol. Biochem. 2013, 64, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Kováčik, J.; Klejdus, B.; Bačkor, M.; Stork, F.; Hedbavny, J. Physiological responses of root-less epiphytic plants to acid rain. Ecotoxicology 2011, 20, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Dolatabadian, A.; Sanavy, S.A.M.M.; Gholamhoseini, M.; Joghan, A.K.; Majdi, M.; Kashkooli, A.B. The role of calcium in improving photosynthesis and related physiological and biochemical attributes of spring wheat subjected to simulated acid rain. Physiol. Mol. Biol. Plants 2013, 19, 189–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debnath, B.; Irshad, M.; Mitra, S.; Li, M.; Liu, S.; Rizwan, H.M.; Pan, T.; Qiu, D. Acid rain deposition modulates photosynthesis, enzymatic and non-enzymatic antioxidant activities in tomato. Int. J. Environ. Res. 2018, 12, 203–214. [Google Scholar] [CrossRef]
- Dursun, A.; Yildirim, E.; Güvenc, I.; Kumlay, A.M. Effects of Simulated Acid Rain on Plant Growth and Yield of Tomato (Lycopersicon Esculentum); International Society for Horticultural Science (ISHS): Leuven, Belgium, 2002; pp. 245–248. [Google Scholar]
- Zhai, Y.; Yang, Q.; Hou, M. The effects of saline water drip irrigation on tomato yield, quality, and blossom-end rot incidence—A 3a case study in the south of China. PLoS ONE 2015, 10, e0142204. [Google Scholar] [CrossRef] [PubMed]
- Zushi, K.; Ono, M.; Matsuzoe, N. Light intensity modulates antioxidant systems in salt-stressed tomato (Solanum lycopersicum L. Cv. Micro-tom) fruits. Sci. Hortic. 2014, 165, 384–391. [Google Scholar] [CrossRef]
- Murshed, R.; Lopez-Lauri, F.; Sallanon, H. Effect of water stress on antioxidant systems and oxidative parameters in fruits of tomato (Solanum lycopersicon L, cv. Micro-tom). Physiol. Mol. Biol. Plants 2013, 19, 363–378. [Google Scholar] [CrossRef] [PubMed]
- Moyle, L.C. Ecological and evolutionary genomics in the wild tomatoes (Solanum sect. Lycopersicon). Evolution 2008, 62, 2995–3013. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Hardeland, R.; Manchester, L.C.; Korkmaz, A.; Ma, S.; Rosales-Corral, S.; Reiter, R.J. Functional roles of melatonin in plants, and perspectives in nutritional and agricultural science. J. Exp. Bot. 2011, 63, 577–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of melatonin in plants: A review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: A physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Debnath, B.; Hussain, M.; Irshad, M.; Mitra, S.; Li, M.; Liu, S.; Qiu, D. Exogenous melatonin mitigates acid rain stress to tomato plants through modulation of leaf ultrastructure, photosynthesis and antioxidant potential. Molecules 2018, 23, 388. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, M.; Ezura, H. Profiling of melatonin in the model tomato (Solanum lycopersicum L.) cultivar micro-tom. J. Pineal Res. 2009, 46, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Zhang, N.; Wang, J.; Zhang, H.; Li, D.; Shi, J.; Li, R.; Weeda, S.; Zhao, B.; Ren, S. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life. J. Exp. Bot. 2014, 66, 657–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Wang, M.; Zhao, Y.; Han, P.; Dai, Y. Melatonin from different fruit sources, functional roles, and analytical methods. Trends Food Sci. Technol. 2014, 37, 21–31. [Google Scholar] [CrossRef]
- Keutgen, A.J.; Pawelzik, E. Modifications of strawberry fruit antioxidant pools and fruit quality under nacl stress. J. Agr. Food Chem. 2007, 55, 4066–4072. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Zhao, X.; Liu, S.; Sun, F.; Zhang, C.; Xi, Y. Beneficial effects of melatonin in overcoming drought stress in wheat seedlings. Plant Physiol. Biochem. 2017, 118, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Luthria, D.L.; Mukhopadhyay, S.; Krizek, D.T. Content of total phenolics and phenolic acids in tomato (Lycopersicon esculentum Mill.) fruits as influenced by cultivar and solar uv radiation. J. Food Compos. Anal. 2006, 19, 771–777. [Google Scholar] [CrossRef]
- Vinha, A.F.; Alves, R.C.; Barreira, S.V.; Castro, A.; Costa, A.S.; Oliveira, M.B.P. Effect of peel and seed removal on the nutritional value and antioxidant activity of tomato (Lycopersicon esculentum L.) fruits. LWT Food Sci. Technol. 2014, 55, 197–202. [Google Scholar] [CrossRef]
- Saleem, A.; Ashraf, M.; Akram, N.; Al-Qurainy, F. Salinity-induced changes in key anti-oxidant enzyme activities and in the levels of some anti-oxidants, osmo-protectants, inorganic ions, and chlorophyll pigments in okra fruit (Abelmoschus esculentus L.). J. Hortic. Sci. Biotech. 2012, 87, 271–277. [Google Scholar] [CrossRef]
- Turk, H.; Erdal, S.; Genisel, M.; Atici, O.; Demir, Y.; Yanmis, D. The regulatory effect of melatonin on physiological, biochemical and molecular parameters in cold-stressed wheat seedlings. Plant Growth Regul. 2014, 74, 139–152. [Google Scholar] [CrossRef]
- Borghesi, E.; González-Miret, M.L.; Escudero-Gilete, M.L.; Malorgio, F.; Heredia, F.J.; Meléndez-Martínez, A.J. Effects of salinity stress on carotenoids, anthocyanins, and color of diverse tomato genotypes. J. Agr. Food Chem. 2011, 59, 11676–11682. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Yemm, E.; Willis, A. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.M.; Cao, F.; Zhang, M.; Chen, X.; Zhang, G.; Wu, F. Difference in yield and physiological features in response to drought and salinity combined stress during anthesis in tibetan wild and cultivated barleys. PLoS ONE 2013, 8, e77869. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (frap) as a measure of “antioxidant power”: The frap assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | β-Carotene (mg/100 g FW) | Lycopene (mg/100 g FW) |
|---|---|---|
| Con | 31.61 ± 1.19 c | 35.53 ± 1.04 c |
| Con + Mel | 34.61 ± 0.46 c | 36.67 ± 0.32 c |
| SAR | 41.96 ± 0.45 b | 42.83 ± 0.44 b |
| SAR + Mel | 45.87 ± 0.87 a | 47.32 ± 0.46 a |
| Treatments | Diameter of Fruit (mm) | Length of Fruit (mm) | Number of Fruit | Average Weight g/Fruit | Dry Matter of Fruit (%) |
|---|---|---|---|---|---|
| Con | 21.99 ± 0.34 a | 21.44 ± 0.18 a | 48.25 ± 1.75 a | 6.29 ± 0.15 a | 5.16 ± 0.06 c |
| Con + Mel | 22.73 ± 0.41 a | 22.13 ± 0.24 a | 51.25 ± 1.65 a | 6.50 ± 0.16 a | 5.24 ± 0.05 c |
| SAR | 16.79 ± 0.11 c | 16.35 ± 0.19 c | 22.75 ± 1.49 c | 3.95 ± 0.11 c | 5.87 ± 0.03 b |
| SAR + Mel | 19.35 ± 0.23 b | 19.12 ± 0.14 b | 38.25 ± 1.25 b | 5.03 ± 0.11 b | 6.11 ± 0.04 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Debnath, B.; Hussain, M.; Li, M.; Lu, X.; Sun, Y.; Qiu, D. Exogenous Melatonin Improves Fruit Quality Features, Health Promoting Antioxidant Compounds and Yield Traits in Tomato Fruits under Acid Rain Stress. Molecules 2018, 23, 1868. https://doi.org/10.3390/molecules23081868
Debnath B, Hussain M, Li M, Lu X, Sun Y, Qiu D. Exogenous Melatonin Improves Fruit Quality Features, Health Promoting Antioxidant Compounds and Yield Traits in Tomato Fruits under Acid Rain Stress. Molecules. 2018; 23(8):1868. https://doi.org/10.3390/molecules23081868
Chicago/Turabian StyleDebnath, Biswojit, Mubasher Hussain, Min Li, Xiaocao Lu, Yueting Sun, and Dongliang Qiu. 2018. "Exogenous Melatonin Improves Fruit Quality Features, Health Promoting Antioxidant Compounds and Yield Traits in Tomato Fruits under Acid Rain Stress" Molecules 23, no. 8: 1868. https://doi.org/10.3390/molecules23081868