Recent Advances in the Synthesis of Glycoconjugates for Vaccine Development


Dipartimento di Chimica, Universita’ degli Studi di Milano, via Golgi 19, 20133 Milano, Italy
*
Author to whom correspondence should be addressed.
Molecules 2018, 23(7), 1712; https://doi.org/10.3390/molecules23071712
Submission received: 8 June 2018
/
Revised: 9 July 2018
/
Accepted: 11 July 2018
/
Published: 13 July 2018
(This article belongs to the Special Issue Conjugate Vaccines from Carbohydrate Antigens)
Abstract
:During the last decade there has been a growing interest in glycoimmunology, a relatively new research field dealing with the specific interactions of carbohydrates with the immune system. Pathogens’ cell surfaces are covered by a thick layer of oligo- and polysaccharides that are crucial virulence factors, as they mediate receptors binding on host cells for initial adhesion and organism invasion. Since in most cases these saccharide structures are uniquely exposed on the pathogen surface, they represent attractive targets for vaccine design. Polysaccharides isolated from cell walls of microorganisms and chemically conjugated to immunogenic proteins have been used as antigens for vaccine development for a range of infectious diseases. However, several challenges are associated with carbohydrate antigens purified from natural sources, such as their difficult characterization and heterogeneous composition. Consequently, glycoconjugates with chemically well-defined structures, that are able to confer highly reproducible biological properties and a better safety profile, are at the forefront of vaccine development. Following on from our previous review on the subject, in the present account we specifically focus on the most recent advances in the synthesis and preliminary immunological evaluation of next generation glycoconjugate vaccines designed to target bacterial and fungal infections that have been reported in the literature since 2011.
1. Introduction
Notwithstanding the great advances of modern medicine, infectious diseases still have a strong impact on public health, both in industrialized and developing countries, due to their significant health-related costs for clinical treatment. In particular, the list of the drug-resistant bacteria is increasing continuously, and novel and more efficient means to prevent microbial infections caused by antibiotic-resistant microorganisms are urgently needed. According to the World Health Organization (WHO) [1], vaccination is the most cost-effective strategy for controlling infections caused by pathogenic microorganisms. Actually, vaccines are able to confer long-term protective immunity on the population and have made possible a great revolution in the 20th century, saving millions of lives.
The surface of bacterial pathogens is covered with a dense array of complex glycans, such as lipopolysaccharide of Gram-negative bacteria and the polysaccharide coat (capsular polysaccharides, CPS) of encapsulated bacteria that are crucial protective antigens and major virulence factors. For example, each strain of Streptococcus pneumoniae (the pneumococcus) produces one out of 90 different capsular polysaccharides, which are believed to have been selected as a mechanism to evade the human immune response [2]. All these glycoforms are capable of interacting with the immune system inducing the production of carbohydrate-specific antibodies. They therefore represent attractive targets for vaccine design.
A major drawback of polysaccharide-based vaccines, however, is their limited clinical efficacy. They induce T cell-independent immune responses, featured by poor immunogenicity in children under 5 years of age, in elderly and immunocompromised individuals, and fail to generate conventional B cell-mediated immunological memory. Polysaccharide immunogenicity can be strongly enhanced by conjugation to an immunogenic carrier protein, providing T cell-dependent glycoconjugate antigens able to stimulate B cell maturation to memory cells and induce immunoglobulin class switching from IgM to polysaccharide-specific IgG. The introduction of glycoconjugate vaccines represented one of the keys for success of vaccination, especially for infants and young children who are the most affected population by infectious diseases [3,4,5,6]. Carbohydrate-based antigens needed for inclusion in a glycoconjugate vaccine, however, are not readily available from natural sources. In particular, the isolation and purification of naturally occurring glycans is still a great challenge that may lead to heterogeneous compositions and batch-to-batch variability. A relevant example is the toxic endotoxin lipid A, a major component of the lipopolysaccharide (LPS) of Shigella flexneri 2a. The development of LPS-based conjugate vaccines against Shigella flexneri requires careful LPS-detoxification, a technically demanding and expensive process which also increases the manufacture costs [7,8]. Hence, the development of cost-effective, glycoconjugate vaccines based on fully synthetic saccharide antigens is gaining growing importance, as demonstrated by the outstanding success of the synthetic vaccine Quimi-Hib [9]. Synthetic glycans, indeed, possess well-defined compositions, affording highly reproducible biological properties and a better safety profile. In addition, synthetic oligosaccharides can help to elucidate the minimal structure of the microbial polysaccharide, referred to as epitope or antigenic determinant [10], that can ensure production of a sufficient amount of bactericidal antibodies to confer long term protective immunity of the host. This step is crucial for the design of a new generation of improved and safer vaccines obtained either from chemical synthesis or bacterial source. Consequently, glycoconjugates based on chemically well-defined oligosaccharide structures are now at the forefront of vaccine development.
Over the last years, the synthesis of complex glycans has made significant progress. A variety of synthetic approaches such as automated solid phase synthesis, one-pot programmable synthesis, enzymatic and improved synthetic methods have introduced new and elegant ways to provide oligosaccharide antigens with well-defined chemical structure for immunological studies. Meanwhile, improved methods for structural elucidation, based on X-ray crystallography, NMR, or in silico studies, as well as advanced techniques to study carbohydrate-protein interactions (glycoarray, surface plasmon resonance, isothermal titration calorimetry, competitive ELISA assay) have been extensively applied to predict the minimal structural requirements needed for the immunological activity of the oligosaccharides. Accordingly, a variety of saccharide fragments reproducing or mimicking the surface carbohydrates of pathogens have been synthesized, coupled to carrier proteins or T cell peptides, and tested for their ability to elicit protective antibodies in animal models. In this regard, in the present review we focus on the most significant advances in the synthesis and preliminary immunological evaluation of synthetic antibacterial and antifungal glycoconjugate vaccine candidates, appeared in the literature from 2011 onwards, following our previous account on the subject [11]. For clarity, the diagrammatic representations of the monosaccharide residues [12] illustrated in Figure 1 above are used throughout this review.
2. Shigella
The Shigella family includes four different groups of Gram-negative bacteria—S. dysenteriae, S. sonnei, S. flexneri and S. boydii—each of them comprising different serotypes. Shigella is the causative agent of endemic and epidemic shigellosis or bacillary dysentery, an invasive disease of the lower intestine, highly diffused in developing countries and particularly in pediatric population. The development of a fully synthetic glycoconjugate vaccine against S. dysenteriae type 1 is currently under investigation, using fragments of the O-antigen of the Shigella LPS. Oligomers up to four repeating units of the tetrasaccharide [α-l-Rha-(1→2)-α-d-Gal-(1→3)-α-d-GlcNAc-(1→3)-α-l-Rha] were first synthesized by Pozsgay and covalently linked to human serum albumin (HSA) [13,14]. Preliminary studies showed that the hexadecasaccharide (n = 4) is the most immunogenic fragment able to elicit anti O-SP-specific IgG in mice [14] and that the upstream residue (non-reducing end) of the synthetic fragments is crucial for the immunogenicity of these conjugates [15].
S. flexneri serotype 2a is the most prevalent pathogenic strain in human and a major cause of the endemic form of shigellosis in developing countries. The O-antigen of S. flexneri 2a surface LPS is an essential virulence factor and consists of a branched pentasaccharide repeating unit (Figure 2).
In 2005, the Mulard group reported the synthesis of the monomer, dimer and trimer of the pentasaccharide repeating unit (AB(E)CD, Figure 2) and their conjugation to a universal T cell peptide epitope, the pan HLA DR-binding epitope (PADRE) [16,17]. Subsequently, the synthesis of S. flexneri serotype 2a O-Ag synthetic fragments 1a–18a (Figure 2) was reported [19]. The antigenicity of all synthetic fragments 1a–18a was evaluated by ELISA assays, in order to identify the immunogenic determinants recognized by five protective mIgGs specific to serotype 2a O-Ag. None of the mono- or disaccharides was recognized, while the sequence ECD (trisaccharide 9a) was the only one recognized by one mIgG out of five. Tetrasaccharide B(E)CD 13a was recognized by three of the protective mIgG out of five. The minimal sequences for recognition of all mIgGs were pentasaccharides AB(E)CD 16a and B(E)CDA 14a. Following these encouraging results, some selected synthetic oligosaccharides were conjugated to tetanus toxoid (TT) protein and used for immunization studies in mice, leading to the identification of a hit glycoconjugate, 19b, containing the trimer of the pentasaccharide AB(E)CD. Glycoconjugate 19b induced an efficient serotype 2a-specific anti-O-Ag Ab response and it was found to be a functional mimic of the native polysaccharide [20]. Recently, Mulard et al. established a reproducible bioconjugation method for the synthesis of the pentadecasaccharide−TT conjugate 19b, which allowed complete control of the optimal loading [18]. Alum, an adjuvant used in licensed glycoconjugate vaccines like Prevnar 13 or Synflorix, was added to S. flexneri serotype 2a vaccine candidate 19b, which upon immunization was shown to generate a higher and sustained anti-LPS IgG response compared to their nonadjuvanted form [18]. Importantly, Mulard et al. showed that anti-LPS IgG elicited by their synthetic TT conjugate 19b recognized SF2a bacteria and not only purified SF2a LPS [18]. In addition to these promising findings, the influence of O-acetylation of S. flexneri 2a O-Ag fragments on antigenicity was studied by Mulard group [21]. Polysaccharide O-acetylation has been shown to play a key role for many pathogens in inducing functional Ab responses [22,23,24]. In particular, three diversely O-acetylated S. flexneri 2a O-Ag decasaccharides were synthesized in homogeneous form and their binding to five different protective mAbs was studied, showing some differences in the recognition patterns. Although these data couldn’t provide an exhaustive proof of the role of O-acetylation for S. flexneri 2a O-Ag and of the effect of multiple acetates on the antigen, this work showed that studies using synthetic oligosaccharides may contribute to a better understanding of the antigen-antibody molecular recognition event.
3. Clostridium difficile
Clostridium difficile is a Gram-positive, spore-forming anaerobic bacterium causing Clostridium difficile infection (CDI), a serious diarrhoeal disease and one of the major cause of hospital-acquired infections (also known as nosocomial infections) in Western countries [25]. The epidemiology of CDI has changed dramatically during this millennium, especially in relation to its clinical presentation, response to treatment and antibiotic resistance [25,26]. In general, after antibiotic treatment that leads to the disruption of the gut microbiota, the intestinal epithelium could be colonized by antibiotic-resistant C. difficile spores, which secrete two toxins (toxin A and toxin B) responsible for the clinical symptoms of CDI. Immune-based strategies based on passive administration of monoclonal antibodies against C. difficile toxins and surface proteins to treat or prevent CDI in animal models and in clinical trials have been recently reviewed [27,28]. Concurrently, bacterial surface glycans, such as PS-I and PS-II, have been recently proposed as potential target for vaccine development with the aim of preventing bacterial adhesion and colonization.
3.1. PS-I-Clostridium difficile
Recently, Martin et al. reported the synthesis of the pentasaccharide repeating unit of PS-I cell wall polysaccharide of C. difficile ribotype 027 (Figure 3), of its related substructures (compounds 20a–25a, Figure 3) and their immunological evaluation for the identification of the minimal epitope [29].
The synthetic fragments were synthesized from monosaccharide building blocks 26–30, linearly proceeding from the downstream end to the upstream end. In particular, the use of the non participating benzyl group at C-2 of thioglycoside 29 provided the condition for 1,2-cis stereoselective glycosylation of glucoside 30, bearing the linker at the anomeric position. Thioglycoside 29 was functionalized with the orthogonal protecting groups para-bromobenzyl (PBB) ether at C-3 and levulinoyl (Lev) ester at C-4 for installation of the branching point.
Glycans 20a–25a, immobilized on microarrays, were screened for antibody recognition with samples from C. difficile patients (stools for IgA and serum for IgG). IgA and IgG antibodies specific to all glycan antigens were present in most fecal samples and sera, respectively, of both patients and control groups. Reconvalescent patients showed highly variable antibody levels and statistically higher IgG levels. Pentasaccharide 25a was conjugated to CRM197 (non-toxic mutant of diphtheria toxin) and the resulting glycoconjugate 25b was injected in mice, inducing Ig class switching, affinity maturation and producing self-specific antibodies, without eliciting antibodies against two control oligosaccharides (C. difficile PS-II hexasaccharide and Leishmania lipophosphoglycan capping tetrasaccharide). Interestingly, antibodies raised by glycoconjugate 25b also recognized trisaccharide 22a and disaccharide α-Rha-(1→3)-Glc 21a, which was identified as the minimal epitope. Indeed, α-Rha-(1→3)-Glc disaccharide-CRM197 conjugate 21b was able to induce antibodies recognizing the C. difficile PS-I pentasaccharide 25a. In a following work [30], a multivalent presentation of disaccharide 21a on an oligo(amidoamine) synthetic scaffold [31] was shown to be highly antigenic. In particular, a pentavalent presentation of the disaccharide 31 (Figure 3), built on the oligo(amidoamine) backbone and displaying a T-cell epitope (amino acids 366–383 of the CRM197 protein) showed increased antigenicity compared with monovalent 21b, eliciting antibodies against pentasaccharide 25a. A detailed investigation of the glycan-antibody binding was conducted with a combination of different techniques like glycan microarray, surface plasmon resonance, interaction map, saturation transfer difference (STD)-NMR and isothermal titration calorimetry (ITC). It was demonstrated that the mAbs mainly interacted with the terminal rhamnose and the adjacent glucose of the disaccharide 21a and that in pentasaccharide 25a the linkage connecting the two disaccharides is not directly engaged in antibody binding, although the affinity (KD) increases from micromolar for disaccharide 21a to nanomolar for pentasaccharide 25a. Both glycoconjugates 21b and 25b are currently in preclinical evaluation as novel vaccine candidates against C. difficile.
3.2. PS-II-Clostridium difficile
The chemical synthesis of the hexasaccharide repeating unit (Figure 4) of PS-II cell wall polysaccharide of C. difficile ribotype 027, one of the most virulent strains, with two similar synthetic strategies, was reported simultaneously by two groups in 2011 [32,33].
The synthesis of PS-II oligosaccharide lacking the phosphate group (compound 32a, Figure 4) was carried out by Oberli et al. [32] via a [4+2] glycosylation of tetrasaccharide AB(D)C 33 with disaccharide B’C’ 34, starting from monosaccharide building blocks 35–38. Hexasaccharide conjugated to CRM197 (32b) was used for mice immunization and resulted in the production of IgG antibodies that bound specifically hapten 32a. In addition, IgA antibodies from the stools of patients diagnosed with CDI (the supernatant stools, and not the serum, were chosen because the contact site with C. difficile is the intestinal mucosa) were analyzed. Glycan microarrays containing hexasaccharide antigen 32a were used to screen patient samples. Anti-PS-II IgA antibodies were found in the stools of patients diagnosed with CDI, suggesting that the synthetic hexasaccharide could be used in a glycoconjugate vaccine candidate against CDI.
Danieli et al. [33] reported the synthesis of hexasaccharide 39a and its phosphorylated analogue 40a, starting from disaccharide 41 and tetrasaccharide 42 (or its analogue 43) via a [4+2] convergent approach (Figure 5).
Tetrasaccharide 42 was in turn prepared from monosaccharide building blocks 44–47, by first assembling the linear trisaccharide ABC and then inserting the α-Glc D unit [33]. An alternative route leading to tetrasaccharide 43 starting from building blocks 48, 49 and 50 was also developed [34]. Of note, the glycosylation with 4,6-O-benzylidene-protected ethylthioglycoside 47 allowed the stereoselective introduction of the 1,2-cis linkage for both routes. Tetrasaccharide AB(D)C 51a, obtained from deprotection of 43, was also synthesized to examine the effect of the branching point of the hexaglycosyl unit in determining the immunogenicity. Sera from mice immunized with the PSII-CRM197 conjugate were used to check their capability to bind synthetic fragments 39a, 40a, 51a. Tetrasaccharide 51a showed no binding, while both 39a and phosphorylated fragment 40a bound anti-PSII antibodies. In a second experiment, the three synthetic glycans conjugated to CRM197 (compounds 39b, 40b and 51b) and native PSII-CRM197 were injected in mice and evaluated for their ability to elicit anti PSII antibodies. Sera were analyzed by ELISA for their content of anti PSII IgG, using PSII-HSA for the coating of the plates. Interestingly, only the glycoconjugates obtained from the native polysaccharide and the phosphorylated hexasaccharide 40b were able to induce IgG antibodies that bound PSII and low levels of anti-PSII IgM antibodies. Tetrasaccharide 51b and the nonphosphorylated hexasaccharide 39b elicited self-specific antibodies but did not induce IgG nor IgM anti-PSII titers. A comparison between the studies of Oberli et al. [32] and Adamo et al. [34] reveals that the phosphate group on the hexasaccharide repeating unit of PS-II plays a subtle immunological role. Indeed, the phosphate group is not required to raise IgG antibodies production against hexasaccharide hapten 32a, while it is a prerequisite to elicit antibodies recognizing, besides the phosphorylated and the nonphosporylated hapten, also the native PS-II polysaccharide. These findings suggested that the charged phosphate is crucial to mimic the native PSII polysaccharide [34] and can be used for the design of carbohydrate antigens as vaccine candidates.
4. Burkholderia pseudomallei
Burkholderia pseudomallei is a Gram-negative environmental bacterium which is widespread in the soil and surface water in southeast Asia and northern Australia, causing melioidosis, a serious and often fatal disease presenting acute pulmonary infections, fulminant sepsis and chronic infection mimicking tuberculosis [35]. Antibiotic treatment is usually divided into two phases: a first phase to prevent death from sepsis and a second phase with the aim of preventing recurrence [36]. This protracted treatment is not always successful and mortality rate remains high (from 15% in Australia to 40% in Thailand, approaching 90% with septicaemia) [37]. For this reason, substantial effort has been undertaken to develop vaccine candidates which would protect humans against B. pseudomallei infections [38]. Among the identified virulence factors [39], the capsular polysaccharide of B. pseudomallei, a homopolymer of 2-O-acetyl manno-heptopyranose (Figure 6) has been recently considered for the development of an effective melioidosis vaccine [40].
Of note, B. pseudomallei CPS is expressed as a unique serotype in all reported isolates [41], identical to the CPS of the related bacterium Burkholderia mallei [42]. The synthesis of B. pseudomallei and B. mallei CPS is challenging due to the presence of β-mannoside linkages and of the CH2-extension at C-6 (Figure 6). In 2016, Scott et al. [37] reported the first synthesis of hexasaccharide 52a starting from key disaccharide fragment 53, which was in turn synthesized from disaccharide 54, which was assembled from building blocks 55 and 56, convergently prepared in large scale from common intermediate 57. This compound was obtained in seven steps from glycal intermediate 58, which was synthesized from mannose 59 (Figure 6). The β-mannoside linkages were introduced using an indirect method, based on stereoselective β-glycosylation (ensured by 2-O-acyl participation on the donor) followed by C-2 epimerization. The latter step, leading to the manno-configuration, was performed at the disaccharide level and after each iterative coupling, through a two-step oxidation-reduction with high stereoselectivity. Hexasaccharide 52a was covalently linked to TT and glycoconjugate 52b, upon mice immunization, raised low but detectable levels of IgG/IgM, as determined by ELISA test. Glycoconjugate 52b, however, was shown to stimulate production of antibodies specific for native CPS, with high functional activity correlated with protective efficacy, as observed by protection in mice following a lethal dose administration of B. pseudomallei [37].
5. Brucella
Brucella is one of the world’s major zoonotic pathogens, causing brucellosis, primarily a disease of animals, such as swine, dogs, cattle, sheep, and goats [43]. Humans are infected by close animal contact or consumption of animal products (raw milk, raw milk products, or raw meat) infected by bacteria of the genus [44,45]. The genus Brucella comprises Gram-negative, facultative and intracellular pathogens and the current classification of recognized species is based on phenotypic characteristics, antigenic variation and prevalence of infection in different animal hosts [46,47]. The disease is not spread by human-human contact and the vaccination of animals appears as the only means for disease eradication by vaccination strategies [48]. The O-antigen polysaccharide domain (OPS) of Brucella LPS is a copolymer of two distinct homopolysaccharide sequences containing the rare sugar 4,6-dideoxy-4-formamido-α-d-mannose (-α-d-Rhap4NFo) [6] and simultaneously expresses two antigens, the A and M antigens (Figure 7).
Three Brucella antigenic phenotypes A+M− (A-dominant), A−M+ (M-dominant) and A+M+ have been identified in Brucella strains [50] and antibodies (IgM) against A and M antigens have been used to detect brucellosis [51,52]. The chemical structure of A and M antigens (Figure 7) was definitively elucidated only recently [53]: a longer inner sequence of α(1,2)-linked residues constitutes the A antigen. A shorter sequence, the M antigen, consists of tetrasaccharide repeating units linked as [α(1,2);α(1,3);α(1,2)] and attached to additional copies of this tetrasaccharide or to the A antigen by an α(1,2) linkage [49].
In 2013, the Bundle group [54] reported the synthesis of pentasaccharide 60a (Figure 7) and nonasaccharide 61a, starting from monosaccharides 62, 63 and 64. The synthetic compounds were tested for antigenicity, after conjugation with bovine serum albumin (BSA) [54]. Glycoconjugate 60b was designed to selectively exhibit the M epitope with limited cross reactivity with A-specific antibodies. The nonasaccharide conjugate (compound 61b), containing A and M epitopes, was designed as a possible universal antigen to detect antibodies in animals or humans infected by B. abortus, B. melitensis, and B. suis. An ELISA test was performed with two monoclonal antibodies (YsT9-1 and Bm10) specific for the Brucella A and M antigens, respectively. Interestingly, nonasaccharide antigen 61a bound A- and M-specific antibodies with equivalent avidity, whereas pentasaccharide 60a displays a preference for the M-specific antibody, as expected. However, pentasaccharide 60a, still displaying α(1,2)-linked residues, retained modest to good binding to A-specific mAbs. This initial result paved the way to produce a glycoconjugate vaccine that would not raise antibodies giving false positive results in diagnostic tests for infection. Indeed, the detection of specific anti-M antibodies would indicate infection by Brucella and not by one of the other closely related bacteria that have PS containing 1,2-linked Rha4NFo or Rha4NAc and are known to induce antibodies reactive in the serological test for brucellosis [55]. In a following work, tetrasaccharide 65a, disaccharide 66a and trisaccharides 67a and 68a (Figure 7) were synthesized to assess the largest and smallest M epitopes [56]. International standard B. abortus serum prepared from cattle experimentally infected with an A-dominant strain bound strongly to disaccharide-BSA conjugate 66b and M tetrasaccharide-BSA conjugate 65b [56]. In addition, 65b and 66b also showed strong binding to M-specific mAbs and weak binding with A-specific mAbs. It was also observed that antibodies raised against exclusively α(1,2)-linked Rhap4NFo did not bind well to the 1,3-linked disaccharide [57]. Further improvement of serodiagnosis of brucellosis came when a tether was introduced at the O-4 of the upstream residue (heptasaccharide 69a, Figure 7) [58]. In particular, conjugate 69b (TT) was used for mice immunization and conjugate 68c (with bovine serum albumin, BSA) to monitor antibody responses by ELISA. Mice immunization with glycoconjugate 69b showed that antibodies to the Brucella A antigen could be produced and that these antibodies didn’t react in diagnostic tests based on the M antigen. These findings were confirmed by the results of immunization studies with the OPS of B. abortus strain S99, which contains 98% α-(1,2) and only 2% α(1,3) linkages conjugated to tetanus toxoid. The OPS was subjected to an oxidation reaction using a procedure that concomitantly oxidized all terminal d-Rhap4NFo residue, essentially destroying the M epitope [58]. Immunization studies in mice showed that antibodies against the A epitope dominated. Taken together, all these studies contributed to identify the main elements for a glycoconjugate vaccine candidate for brucellosis and demonstrated that diagnostics based upon the M or A (terminal) epitopes can discriminate infected from vaccinated animals.
6. Haemophilus influenzae Type b (Hib)
Haemophilus influenzae is a Gram-negative bacterium predominantly colonizing the human respiratory tract. H. influenzae strains are divided into two subgroups: unencapsulated strains, also named non-typeable (non-reactive with typing antisera) and encapsulated strains (reactive with typing antisera) comprising six serotypes: a, b, c, d, e and f. In particular, serotype b strains (H. influenzae b, Hib) cause severe diseases including meningitis, pneumonia and septicemia, especially in infants and children [59]. Hib CPS consists of a polymer of β-d-ribose-d-ribitol-5-phosphate (PRP) disaccharide, characterized by the presence of a phosphodiester linkage between repeating units (Figure 8).
The first generation of Hib vaccines, made with purified polyribosyl-ribitol phosphate, induced relatively low titers of serum antibodies, insufficient to protect children from invasive disease [60], and were replaced by Hib PS-conjugate vaccines (PedVaxHIB®, ActHib®, HibTiter®) [61]. Vérez Bencomo et al. developed the first synthetic glycoconjugate vaccine in 2004 [9], QuimiHib® (compound 70, Figure 8) using a one-pot polycondensation strategy starting from synthetic β-d-ribose-(1,1)-d-ribitol-5-H-phosphonate derivative 71 and the phosphodiester-linked compound 72 [9]. Final conjugation to TT gave the fully synthetic glycoconjugate vaccine QuimiHib® [62], which contains a mixture of oligosaccharides with six to eight repeating units on average. Recently, Baek et al. [62] have reported the synthesis of CRM197 glycoconjugates of PRP oligosaccharides up to decamers (compounds 73–76, Figure 9).
Oligosaccharide synthesis was performed via H-phosphonate chemistry starting from tetrasaccharide building block 77 and using a [4+4] iterative elongation strategy. Tetrameric, hexameric, octameric and decameric PRP fragments were obtained using this iterative approach, followed by the introduction of phosphodiester-linked spacer. Tetrasaccharide 77 was synthesized from disaccharide 78 (Figure 9), in turn obtained from the dithioacetal building block 79. After conjugation to CRM197, immunogenicity studies with the synthetic glycoconjugates 73–76 were performed in a rabbit model. After immunization, sera IgG levels towards the PRP oligosaccharides were determined by glycan array analysis. Tetramer conjugate 73 and octamer conjugate 75 exhibited the highest immunogenicity, most likely indicating that four repeating units are sufficient for immunogenicity, while the hexamer conjugate 74 exhibited lower immunogenicity. This result was ascribed to the folding of the structures and to their different interaction with the immune system receptors. The authors concluded that glycoconjugates of synthetic Hib PRP are immunogenic in a rabbit model and, in particular, tetrameric conjugate 73 is a promising candidate for the design of a new glycoconjugate Hib vaccine.
7. Streptococcus pneumoniae
S. pneumoniae, a Gram-positive organism, is a major cause of pneumonia, otitis media, meningitis and septicemia. Various virulence determinants of pneumococci have been identified including the highly variable capsular polysaccharide (CPS), pneumolysin toxin and surface lectins. Bentley and colleagues have determined the DNA sequence of the capsular biosynthesis genes for all 90 serotypes (ST) of S. pneumoniae and found that each serotype has a different CPS composition [2,64]. The first generation carbohydrate-based vaccine PPV23 (Pneumovax®, Merck) containing the 23 most prevalent serotypes is available in the United States and in Europe, although conflicting data about its efficacy have been reported [65]. To improve the immunogenicity, glycoconjugate vaccines like PCV7 (Prevnar®, containing PS from serotypes 4, 6B, 9V, 14, 18C, 19F, and 23F), PCV13 (Prevnar 13™, containing PS from serotypes 4, 6A, 6B, 7F, 9V, 14, 18C, 19A, 19F, 23F, 1, 3 and 5) and PCV10 (GlaxoSmithKline’s Synflorix™, containing PS from serotypes 1, 4, 5, 6B, 7F, 9V, 14, 18C, 19F, and 23F) have been licensed and commercialized. Although S. pneumoniae CPS-based glycoconjugate vaccines are in current routine immunization programs and notwithstanding the increased coverage of strains, diseases caused by serotypes not included in the above vaccines can increase in the long run [64,66]. Recent efforts have been dedicated to the synthesis of antigens from S. pneumoniae serotypes not included in licensed formulations. Of note, glycoconjugates from synthetic fragments of S. pneumoniae serotype 8 have been tested in coformulation with PCV13, as reported in Section 7.6. Recently, the synthesis of the hexasaccharide repeating unit of S. pneumoniae serotype 12F, also not included in marketed formulations, has been reported by Seeberger et al. [67]. Meanwhile, alternative and combined approaches are emerging for vaccine development, based, for instance, on immunization with a combination of bacterial lectins and surface polysaccharides. In a recent study, the surface polysaccharide serotype 6B (PS6B) of S. pneumoniae was conjugated to a recombinant pneumococcal surface protein A (lectin rPspA), a highly immunogenic surface protein produced by all strains of S. pneumoniae, showing the ability of the novel conjugate to induce production of functional anti-rPspA1 and anti-PS6B antibodies [68].
7.1. S. pneumoniae Serotype 1
S. pneumoniae serotype 1 (ST1) CPS (Figure 10) contains the rare monosaccharide 2-acetamido-4-amino-2,4,6-trideoxy-d-galactose (d-AAT) bearing a free amine at C-4. Synthetic fragments of ST1 CPS have been reported by Wu et al. [69], Christina et al. [70] and Schumann et al. [71]. In particular, Schumann et al. also contributed to the identification of the protective epitope of ST1 CPS. ST1 is one of the serotypes difficult to target by vaccination due to the low levels of functional antibodies induced by licensed glycoconjugate vaccines. This was recently [72] ascribed to the concealment of the protective epitope during chemical activation and conjugation to carrier protein. Indeed, conjugation strategies by means of reductive amination (PCV13) or 1-cyano-4-dimethylaminopyridine activation chemistry (PCV10) could lead to partial destruction of the d-AAT moieties by reaction with the free amines on this rare monosaccharide. To confirm this assumption Schumann et al. [72] synthesized and tested fragments of ST1 CPS and of the closely related Bacteroides fragilils PS A1 CPS (Figure 10). Synthetic oligosaccharides 80–85 were then subjected to glycan microarray analysis of ST1- and PS A1-directed antisera. Trisaccharide 80 bound to antibodies contained in ST1 typing serum, while disaccharide 82, missing the d-AAT moiety, was bound in a much lower extent, revealing the importance of d-AAT for immune recognition. Neither the PS A1 repeating unit 81 nor d-AAT alone 83 or galacturonic acid alone 84 were bound. Trisaccharide 80 was then conjugated to CRM197 (glycoconjugate 86) by reaction with the thiol group, thus preserving the amino group of d-AAT.
Immunization studies in rabbit models showed that glycoconjugate 86 elicited a higher immune response against trisaccharide 80, d-AAT 83 as well as ST1 CPS compared to PCV13 or CRM197 alone. The antibacterial properties of sera against glycoconjugate 86 were evaluated in vitro and in vivo. In particular, flow cytometry revealed that antibodies in sera from glycoconjugate 86-immunized rabbits bound better to ST1 bacteria than sera from PCV13-immunized rabbits. Bacterial binding correlated with serum opsonophagocytic killing capacities. Mice were passively immunized with serum of rabbits immunized with glycoconjugate 86 and then transnasally infected with ST1 pneumococci, showing fewer bacterial colonies than mice pretreated with sera from PCV13 or CRM197 alone-immunized rabbits. Given the importance of these findings, glycoconjugate 86 is now advancing in preclinical development for inclusion in semisynthetic vaccines covering multiple pneumococcal serotypes [72]. Interestingly, this work further demonstrates that the use of synthetic oligosaccharide antigens may be crucial to unveil hidden protective epitopes by means of site-selective protein conjugation.
7.2. S. pneumoniae Serotype 2
ST2 is one of “nonvaccine serotype”, i.e., not covered by licensed PCVs based on capsular polysaccharides. It is one of the main cause of invasive pneumococcal diseases (IPD) responsible for pneumonia, septicemia, meningitis, and otitis media in many countries in Asia [73] and Central America [74]. The structure of ST2 CPS is composed of a hexasaccharide repeating unit illustrated in Figure 11 [75].
Emmadi et al. [76] reported the synthesis of the repeating unit of ST2 CPS and of series of synthetic glycans containing portions of the ST2 CPS (compounds 87a–93a), in order to identify the protective oligosaccharide epitope. Hexasaccharide 89a (one repeating unit) was synthesized from disaccharides 94, 95, 96 via a [2+2+2] glycosylation strategy (Figure 11). These disaccharide units were in turn synthesized from l-rhamnose and d-glucose building blocks 97–102 (Figure 11). The β-rhamnosidic linkage in 94 was incorporated by installing a remote C3 picoloyl group on rhamnosyl thioglycoside 97 for hydrogen-bond-mediated aglycon delivery. The 1,2-cis linkage between glucose building blocks 101 and 102 was formed by in situ anomerization, by converting 102 to the corresponding glycosyl bromide and then by reatcion with 101 in the presence of TBAI. Glycan microarrays containing oligosaccharide fragments 87a–93a were used to screen human and rabbit sera specific to serotype 2 CPS and to identify epitope hits. These experiments demonstrated that the α-d-GlcA-(1→6)-α-d-Glc-(1→2) branch is important to have strong specific antibody binding. Hexasaccharide 89a was conjugated to CRM197 and used for mice immunization producing very high titers of CPS-specific opsonizing antibodies that efficiently fix complement and promote killing of pneumococci by phagocytic activity. An in vivo experiment to evaluate the vaccine involved subcutaneous immunization of mice that were infected with highly virulent ST2 strain NCTC7466. Neoglycoconjugate hexasaccharide-CRM197 89b stimulated a T cell-dependent B cell response that induced CPS-specific antibodies resulting in the reduction of the bacterial infection in lung tissues and blood.
7.3. S. pneumoniae Serotype 3
The commercial anti-pneumococcal glycoconjugate vaccine PCV13 includes S. pneumoniae serotype 3 (ST3). However, ST3 glycoconjugate contained in PCV13 has shown an atypical immunogenicity, ascribed to the abundant CPS expression on the capsule and to a weakened booster response leading to hyporesponsiveness (inability of the individual to mount an immune response after booster vaccination) [77,78]. Indeed, the levels of pre-existing ST3-specific antibody were found to be negatively correlated with the B cell memory response to a booster dose of PCV13 containing ST3 glycoconjugate [79]. This behavior has been associated with a lack of protection against acute otitis media [80]. As a consequence, one can assume that pure synthetic antigenic structures, designed on the basis of antibody binding specificities, could improve immunogenic properties of ST3 CPS conjugates. Synthetic oligosaccharides based on ST3 CPS repeating units have been already reported by Benaissa-Trouw et al. in 2001 [80] and they have been proven to protect mice against lethal intraperitoneal challenge with ST3 pneumococci. Recently, Parameswarappa et al. [81] reported the synthesis of a library of oligosaccharides, compounds 103a–110a and their corresponding CRM197 conjugates 103b–110b (Figure 12).
The synthesis of the fragments was achieved from disaccharide 111, in turn obtained from two differentially protected glucose building blocks 112 and 113. Tetrasaccharide 110a was synthesized with a [2+2] strategy in 13% overall yield from 112 and 113. The synthetic ST3 oligosaccharides potentially contained the minimal protective glycan epitope (Figure 12). Glycan arrays containing the different fragments were used to screen human sera for antibodies and to define the recognition site of two protective ST3-specific monoclonal antibodies (mAbs). Tetrasaccharide 110a contains the protective epitope of both mAbs and was selected for further immunogenicity studies. The CRM197 conjugate 110b elicited protective immunity as evidenced by opsonophagocytosis assays and mice immunization experiments against experimental pneumonia caused by transnasal infection with ST3 strain PN36. Formulation of the protective epitope has to be further evaluated to elicit optimal long-term immunity.
The synthesis of ST3 CPS oligosaccharides 114–117 (Figure 13) has been recently reported by Xiong et al. [82] These oligosaccharides were also designed to have different sugar residues, Glc (114 and 116) or GlcA (115 and 117) at the upstream end. As an example, heptasaccharide 116 was synthesized by a 3+[2+2] glycosylation strategy from trisaccharide 118 and disaccharides 119 and 120, all achieved from common precursor 121. Hexasaccharide 115 was synthesized by a [2+2]+2 strategy from disaccharides 122, 123 and 120. The oligosaccharides were designed to expose a free amino group at their downstream ends (114a–117a) to allow conjugation with tetanus toxoid (TT) (114b–117b) and BSA (114c–117c) carrier proteins. [83]. TT conjugates 114b–117b and free oligosaccharides 114a–117a were injected in mice and the obtained antisera were analyzed by ELISA using BSA conjugates 114c–117c as capture antigens. Antisera derived from mice immunized with TT conjugates 114b–117b contained significantly higher specific antibodies compared to 114a–117a. In particular, antibody titers induced by 114b and 115b were significantly higher than those induced by 116b and 117b, showing that the chain length of ST3 CPS oligosaccharides influences the immunological properties and that longer oligosaccharides are not necessarily better haptens.
7.4. S. pneumoniae Serotype 4
The CPS of S. pneumoniae serotype 4 (ST4) contains a rare and labile substituent, the trans-2,3-(S) cyclic pyruvate ketal modified galactose (residue A, Figure 12). The ST4 repeating unit is a tetrasaccharide made of [3)-β-d-ManpNAc-(1→3)-α-l-FucpNAc-(1→3)-α-d-GalpNAc-(1→4)-α-d-Galp-2, 3-(S)-Pyr-(1→] (Figure 14).
Recently, the Seeberger group [84,85] reported the synthesis and immunological evaluation of fragment 124a, corresponding to the repeating unit, and shorter oligomers 125a–131a with and without the pyruvate ketal, demonstrating the importance of the trans-2,3(S)-pyruvate ketal in the ST4 epitope. In particular, the synthetic fragments were obtained with a linear glycosylation approach from building blocks 132–135 (Figure 13). Of note, for the installation of 1,2-cis linkages, glycosylation of galactose 135 with donor 134 occurred with good stereoselectivity of the newly formed glycosidic linkage (α:β = 7:1). On the other hand, installation of the final β-manno linkage in unit D was accomplished using an indirect two-steps method. Indeed, exclusive β-glucosylation was achieved with donor 132 using NIS and TfOH as promoters. The following 2-OH activation and amination established the desired manno configuration at C-2. Glycan arrays showed that ST4-directed antibodies in the human reference serum (serum 007sp) [85] recognized both pyruvate-dependent and pyruvate-independent epitopes. Oligosaccharide 124a showed the highest antibody affinity and cross-reactivity to ST4 CPS in mice and humans immunized with the natural CPS. Human serum 007sp contains antibodies recognizing also non-pyruvalated oligosaccharides 129a and 131a. Thus, it was hypothesized that non-pyruvalated epitopes could be present in the natural CPS, although non-pyruvalated epitopes could be less immunogenic than pyruvalated epitopes, as indicated by lower antibody binding signals to 129a and 131a compared to 124a. To verify this behavior, two selected CRM197 conjugates of non-pyruvalated ST4 oligosaccharides, 129b and 131b, were used for mice immunization. The raised antibodies did not recognize the natural polysaccharide on the surface of ST4 bacteria. This result confirmed that the pyruvate motif on the oligosaccharide is needed for cross-reactivity with the native CPS.
7.5. S. pneumoniae Serotype 5
Serotype 5 (ST5) is the fifth most prevalent serotype of S. pneumoniae and is included in the PCV10 and PCV13 [86]. ST5 CPS repeating unit (Figure 15) [87] contains a branched N-acetyl-l-fucosamine (l-FucpNAc) linked to d-glucose (d-Glc) and d-glucuronic acid (d-GlcA) and two rare deoxyamino sugars: the ketoamino sugar 2-acetamido-2,6-dideoxy-d-xylose-hexos-4-ulose (Sugp) and the N-acetyl-l-pneumosamine (l-PneupNAc), which is α(1→2) linked to d-GlcpA. During CPS isolation and purification for the production of the glycoconjugate vaccine, the keto group of Sugp can be partially or fully reduced to form a mixture of ST5 CPS components with decreased immunogenicity compared with the native ST5 CPS [88].
Recently, Lisboa et al. [89] reported the synthesis of ST5 CPS fragments 136a–138a starting from six differentially protected monosaccharide building blocks 139–144 (Figure 15). In particular, l-fucosamine acceptor 143 and l-pneumosamine donor 144 were both synthesized from l-fucal 145 via an azido-phenylselenation reaction on the double bond. Among the oligomers synthesized, oligomer 136a contains N-acetyl-d-quinovosamine (A’, d-QuiNAc) in place of of Sugp (A), displaying a hydroxyl group at C-4 in place of the labile carbonyl occurring in the native ST5 CPS. Seeberger group [89] uncovered the protective ST5 CPS epitope using a combination of glycan microarray-based mAb generation and immunological evaluation performed in rabbit models. These experiments showed that the rare aminosugar l-PneuNAc, as well as the branching, are essential for antibody recognition and avidity. Interestingly, it was also demonstrated that CRM197 glycoconjugate 136b, containing d-QuiNAc, induced higher antibody titers and opsonic activity compared to native ST5-CRM197 conjugate contained in PCV13 vaccine. Special care should be taken, however, in the interpretation of the results obtained with such different vaccination modalities, i.e., a 13-valent vaccine vs. a monovalent synthetic vaccine. The latter indeed contains only one type of carbohydrate which is administered in much higher amount in comparison to the same carbohydrate contained in PCV13. Nevertheless, this result suggests the possibility for the replacement of labile functional groups, generating manufacture issues, with stable functional groups that do not affect the immunogenic properties of glycoconjugates.
7.6. S. pneumoniae Serotype 8
ST8 is part of the first-generation polysaccharide vaccine PPV23, but it is not included in glycoconjugate vaccines PCV7, PCV13 and PCV10. Many clinical ST8 isolates were found resistant to antibiotics like erythromycin, clindamycin, tetracycline and ciprofloxacin [90]. Furthermore, this multiresistant serotype is a major cause for concern in HIV-infected patients, where its occurrence is significantly more frequent [91]. ST8 CPS consists of a linear tetrasaccharide repeating unit (Figure 16) sharing a common cellobiuronic acid disaccharide [β-d-GlcA-(1→4)-β-d-Glc] with ST3 CPS (BA sequence, Figure 12).
Schumann et al. [92] reported the preparation of tetrasaccharide fragments of ST8 CPS 146a–150a, to identify the minimal protective glycan epitope. The four tetrasaccharides were synthesized by automated glycan assembly, using solid-phase oligosaccharide synthesis, starting from building blocks 151–156. Glycan microarray containing all ST8 CPS frameshifts led to the identification of one tetrasaccharide frameshift (BAEC, 148a) that was preferentially recognized by a protective mAb, a murine immunoglobulin M (IgM) against native ST8 CPS [93]. Conjugation with CRM197 of the tetrasaccharide 148a gave the ST8 glycoconjugate 148b, which was used for immunization of mice and rabbit models. Interestingly, although cellobiuronic acid disaccharide conjugate 103b (BA, Figure 12) conferred protective immunity against ST3 [81], no ST3-directed immune response was found after mice immunization with conjugate 148b (BAEC), probably because of the different presentation of cellobiuronic acid in the ST8 sequence. Conjugation with CRM197 and coformulation with PCV13 of either tetrasaccharide 148a or its congener tetrasaccharide 150a (AAEC), containing a d-Glc residue in place of the naturally occurring d-GlcA, led to a new vaccine which conferred protective immunity in rabbits against all the 14 S. pneumoniae serotypes. This achievement confirms the possibility of adding synthetic oligosaccharide antigens to existing vaccines with the aim of expanding current formulations and replacing serotypes that are not efficiently targeted.
7.7. S. pneumoniae Serotypes 14 and 19F
Among the synthetic glycoconjugate vaccines for S. pneumoniae, gold nanoclusters have been recently explored, highlighting the potential of these carriers for the development of synthetic vaccines [94,95]. In particular, glyconanoparticles bearing the synthetic tetrasaccharide epitope of S. pneumoniae type 14 (ST14) PS have been recently reported [96]. ST14 PS consists of repeating units of the tetrasaccharide (6)-[β-d-Galp-(1→4)-]β-d-GlcpNAc-(1→3)β-d-Galp-(1→4)β-d-Glcp-(1→)n (Figure 17) [97].
The synthetic branched tetrasaccharide Gal-Glc-(Gal-)GlcNAc (157a), synthesized and studied by Mawas et al. [99] was identified as the smallest structure producing protective antibodies against ST14 when conjugated to CRM197 protein (glycoconjugate 157b) [100]. Tetrasaccharide 157c, derivatized with a terminal thiol for nanoparticle functionalization, was conjugated together with the T cell-stimulating ovalbumin peptide (OVA 323–329) and d-glucose fragment 158 to produce small (2 nm) hybrid gold glyconanoparticles 159 (GNPs, Figure 17). Immunogenicity studies in mice showed that 159 induced the production of specific IgG antibodies against ST14 PS. The presence of OVA 323–339 peptide was necessary for the induction of high affinity IgG antibodies, while the T cell epitope did not raise anti-OVA 323–339 peptide antibodies, thus avoiding the risk of epitope suppression. Sera obtained from mice immunized with 159 with a ratio of tetrasaccharide:Glc:OVA 323–339 = 45:50:5 were able to opsonize ST14 bacteria, although less efficiently than sera from mice immunized with native ST14 PS conjugated to CRM197. These results make 159 a promising S. pneumoniae type 14 vaccine candidate.
In another recent study [98], gold glyco-nanoparticles (GNP) were prepared with synthetic oligosaccharide fragments corresponding to the repeating units of S. pneumoniae CPS type 19F and 14. In particular trisaccharide 140a, corresponding to ST19F repeating unit [β-d-ManpNAc-(1→4)-α-d-Glcp-(1→2)-α-l-Rhap-(1→] (Figure 17), was prepared according to procedures described in the literature [101,102] and derivatized as thiol-ending ligand 160b. Tetrasaccharide 157c (fragment of ST14), trisaccharide 160b (fragment of Pn19F), d-glucose fragment 158 and OVA 323–339 peptide were loaded onto GNPs (161, Figure 17) in different ratios. After mice immunization, GNPs 161 enhanced the production of specific IgG antibodies toward ST14 PS, while no IgG antibodies against ST19F PS were elicited. In particular, the titers of specific IgG antibodies towards ST14 polysaccharide raised by 161 were higher than the titers elicited by GNPs exclusively displaying ST14 (159), and comparable with commercially available PCV13. Of note, this work explored the effect on the immunological response of glyconanoparticles displaying two carbohydrate epitopes from different bacterial serotypes.
8. Group A Streptococcus
Group A Streptococcus (GAS) is a Gram-positive microorganism causing post-sequelae autoimmune infections including rheumatic heart disease. The main driver of autoimmunity is the surface-anchored GAS M polymorphic proteins [103]. Indeed, formulated multivalent M protein-based vaccines were tested in animal and human models but they are protective only for the serotypes included in formulation. For this reason, the identification of a common protective antigen is highly desirable. Due to its prominence in the GAS cell wall and its conservation across all GAS strains, the Lancefield group A carbohydrate (GAC) has been considered as a potential antigen for a universal GAS vaccine [103]. The Lancefield group A carbohydrate (GAC) consists of a α-l-Rhap(1→3)-α-l-Rhap(1→2)-[β-d-GlcpNAc]-(1→3) repeating unit (Figure 18).
Increasing concerns regarding autoreactivity of antibodies that recognize the native GAC GlcNAc side chain (anti-GlcNAc monoclonal antibodies) [105], however, have been supported by recent studies [106,107]. Cross-reactivity (especially in heart or brain tissues) of anti-GlcNAc mAb was hypothesized almost twenty years ago [108] and it is still a crucial point of discussion, as well as the role of polyrhamnose on the immunogenicity. More recently, Henningham et al. [109] reported that the relative contribution of GlcNAc side chains to the innate immune resistance of GAS varies among strains and that GlcNAc side chain is not a universal GAS virulence factor in animal models. In 2010, Kabanova et al. [104] reported the synthesis of two sets of hexasaccharide- and dodecasaccharide-CRM197 conjugates 162b–165b (Figure 18) and compared their immunogenicity with the native GAC-CRM197 conjugate. All oligomers 162a–165a were synthesized starting from building blocks 166 and 167. Of note, the GAC isolated from bacterial fermentation was found to be contaminated with polyrhamnose variant species. The synthetic oligosaccharide conjugates 162b–165b showed similar immune response in mice compared to GAC conjugate against two GAS isolates of M1 and M23 serotypes. A saccharide chain length of six (the minimal size of the antigen) was found to be sufficient to elicit protective antibodies.
More recently, Auzanneau et al. [110] reported the synthesis of hexasaccharide 164a and its conjugation to TT carrier protein to give 164c. Previously reported conformational analysis had shown that the branch-point in the trisaccharide repeating motif was important for antibody recognition in the antibody-ligand complex [111]. Epitope mapping of a branched trisaccharide and a doubly-branched hexasaccharide by saturation transfer difference NMR methods [112] confirmed the importance of the branched trisaccharide epitope that was studied in binding experiments with a mouse monoclonal antibody. The immunogenicity of the synthetic hexasaccharide–TT conjugate 164c was confirmed by primary (IgM) and secondary antibody (IgG) responses, with anti-hexasaccharide titers that increased after booster immunizations to mice. These titers were similar to those obtained with the native GAC–TT conjugate.
9. Group B Streptococcus
Streptococcus agalactiae or Group B Streptococcus (GBS) is the leading cause of invasive infections in pregnant women [113], newborns, and elderly people, resulting in pneumonia, sepsis and meningitis [114,115]. GBS is a multiserotype Gram-positive bacterium that expresses Lancefield group B polysaccharide as a major virulence factor. Ten different serotypes of GBS PS have been characterized (Ia, Ib, II, III, IV, V, VI, VII, VIII, IX), but five of them (Ia, Ib, II, III and V) account for the vast majority of the disease [116]. Chemical synthesis of the repeating unit of some of these serotypes (types Ia, II, V) have been reported in recent years by Guo and Gao groups [117,118,119]. These works may be useful for the synthesis of other fragments of GBS PS and for further investigations, like antigenicity and immunological studies. GBS type III repeating unit is composed of →4-β-d-Glcp-(1→6)-β-d-GlcpNAc-[α-NeuNAc-(2→3)-β-d-Galp-β-(1→4)]-(1→3)-β-d-Galp-(1→ (Figure 19).
Baker et al. [113,120] reported that GBS PSIII conjugated to TT carrier protein resulted in high tolerance when administered to pregnant women, raising highly specific IgG Abs titers which were transferred through the placenta to infants. Chemical synthesis of fragments of PSIII and of related desialylated fragments have been reported [121,122,123,124]. Conformational studies and molecular dynamics simulations [125,126] showed high flexibility of GBS PSIII, as it adopts a partial helical conformation thanks to the presence of α-NeuNAc-(2→3)-β-d-Galp-β-(1→4) branch, while without the sialic acid residues a random coil conformation is preferred. In particular, it has been shown that there is a specific interaction between the sialic acid residues and the glucosyl and galactosyl backbone which influences the orientation of the side chain and the backbone conformation [125,126]. These behaviors have been rationalized hypothesizing the existence of an extended conformational epitope. A recent study carried out by Adamo et al. [127] showed that synthetic fragments of GBS PSIII conjugated to CRM197 are recognized by polyclonal PSIII specific serum and that the presence of the branch is a structural relevant motif for the recognition of anti-PSIII antibodies. Even if these neo-glycoconjugates can’t still be considered vaccine candidates, this promising result paves the way to the use of synthetic GBS PSIII oligosaccharide as tools to study their detailed molecular interactions with anti-PSIII mAbs.
10. Salmonella Typhi
Salmonella enterica serovar Typhi, generally termed Salmonella Typhi (S. Typhi) is a highly invasive encapsulated Gram-negative bacterium, responsible for typhoid fever, a systemic infection mostly diffused in less-developed geographic areas, lacking proper sanitary conditions. Infections occur generally via consumption of contaminated food and water. Global estimates of typhoid fever burden range between 11 and 21 million cases and approximately 128,000 to 161,000 deaths annually [128], with a peak incidence in individuals from early childhood to 15 years old [129]. Clinical diagnosis of the infection is difficult due to the often non-specific symptoms of typhoid fever that can be confused with other common febrile illnesses [130] and due to serological tests that often give false-negative and false-positive results [131,132]. S. typhi capsule contains three antigens: the H antigen is a heat sensitive protein of the peritrichous flagellae, while the O or somatic antigen is a cell-wall lipopolysaccharide. The Vi antigen is the capsular polysaccharide which overlies the O antigen. The Vi antigen plays a crucial role in the modulation of early inflammatory responses during S. Typhi infections [133,134] and represents the basis for the formulation of vaccines against this bacterium [134,135,136]. It is called Vi (“Virulence”) antigen due to its ability to enhance S. Typhi virulence [137,138]. It is an anionic polymer composed by α-(1→4)-linked N-acetyl galactosaminuronic acid repeating units predominantly O-acetylated at position 3 (Figure 20). The degree of 3-O-acetylation ranges from 60% to more than 90% in some strains and the immunogenicity of Vi antigen is closely related to its degree of O-acetylation [139]. The carboxylic acids are less exposed and partially shielded by the O-acetyls and this can explain the minor effect upon the immunological properties observed after reduction of the carboxylic acids [139].
Although pure polysaccharide vaccines based on the purified Vi antigen have been proven effective in adults, they have been so far ineffective in infants (especially children younger than 5 years of age), in the elderly and immunocompromised individuals [143]. The coupling of S. Typhi CPS fragments to carrier proteins (rEPA, TT, CRM197) produced glycoconjugate vaccines able to elicit a T cell dependent immune response [144,145]. In particular, the conjugate vaccine Vi-TT was recently found effective in the prevention of typhoid fever in a phase 2b trial and proven to be safe and highly immunogenic [146]. The injectable Vi-TT conjugate vaccine (TCV) is currently licensed and recommended by WHO for children from 6 months of age and adults up to 45 years of age. Synthetic oligomers of Vi antigen were first reported in the literature by Sinaÿ and coworkers [140]. In particular, Vi oligosaccharides up to hexasaccharide 168–172 (Figure 20) bearing an unnatural O-methyl group both at the C4 position of the upstream residue and at C-1 position of the downstream residue have been synthesized, thus precluding protein conjugation [140]. Recently, Ye and co-workers [141] reported the synthesis of Vi oligomers 173–176 as methyl glycosides containing an unnatural acetyl group at C-4 of the upstream residue (Figure 20). In particular, N-acetyloxazolidinone-containing glycosyl donor 177 and acceptor 178 were used to direct alpha stereoselectivity during glycosylation reactions. ELISA competitive assays showed that synthetic tri- and tetra-saccharides 174 and 175 had improved antigenic activities in comparison to Sinaÿ fragments [140]. The authors speculated that improved affinities could be ascribed to the presence of an acetyl group at C-4 of the upstream residue in place of the methyl ether present in Sinaÿ’s structures. More recently, Ye et al. [142] synthesized a series of Vi pseudo-oligosaccharides 179–183 (Figure 20) conjugated by carbon chain spacers through olefin cross metathesis or by the 1,2,3-triazole moiety through Huisgen cycloaddition reaction. The binding affinities to anti-Vi antibodies of proposed mimics 179–183 were investigated, showing that the affinity of divalent compounds was generally comparable to the monovalents of the same length. For example, the affinity of 181, containing the butylene linker did not increase significantly when compared with that of monovalent tetrasaccharide. Heterodimer 182, which mimics the native Vi antigen with the similar chain-elongation direction did not result in improved antigenicity, perhaps suggesting that longer Vi oligomers are needed for higher affinity.
Recently, the synthesis of di- and trisaccharide fragments of S. Typhi Vi capsular polysaccharide analogues and their zwitterionic counterparts has been accomplished by the Lay group (Figure 21) [147]. These fragments were composed of N-acetylgalactosaminuronic acid repeating units non-acetylated at position 3 (Figure 21). The synthetic strategy was designed in order to obtain the two distinct series of oligomers 184–187 (2-acetamido derivatives and their zwitterionic analogues) from common building blocks, donor 188 and glycosyl acceptor 189, in turn synthesized from commercially available d-galactosamine hydrochloride. Glycosylation reaction of acceptor 189 with donor 188 gave exclusively the desired α (1,4) disaccharide and the same 1,2-cis stereoselective outcome was observed for the trisaccharides.
ELISA tests showed that oligosaccharides 184–187 were recognized by specific anti-Vi polyclonal antibodies in a concentration-dependent manner with similar efficacies, lower than the natural Vi polysaccharide. This might be related to the short chain length of the synthetic fragments and to the lack of the 3-O-acetyl group, which has been reported as being important for the immunogenicity [139].
11. Pseudomonas aeruginosa
Pseudomonas aeruginosa is an opportunistic Gram-negative bacterium that can cause hospital-associated infections, often life-threatening in critically ill patients [148]. Cystic fibrosis patients often become infected with P. aeruginosa in chronic lung infections [149]. P. aeruginosa encodes several multidrug efflux pump genes [150] and has acquired multiple resistance mechanisms to most antibiotic classes, selected by years of antibiotic treatments [151]. In the past few years, new approaches such as the administration of anti-bacterial monoclonal antibodies are being investigated for the prevention or treatment of P. aeruginosa infections [152]. In a recent study, P. aeruginosa bloodstream infection isolates from patients with acute P. aeruginosa infections were analyzed for the ability to express PcrV, a type 3 secretion protein, and Psl exopolysaccharide, an important component of the microbial biofilm extracellular matrix [153]. The study showed that the majority of isolates expresses PcrV and Psl. However, most of the patient’s sera lacked IgG and functionally active responses to these targets. These findings suggest that Psl can shield the bacterium from the host immune response, allowing the survival of the bacterium [153]. In particular, Psl is a serotype-independent antigen anchored to the cell surface in a helical pattern, an organization that can be crucial for cell–cell interactions and to engage in interaction with other biofilm-initiating components [154,155].
Di Giandomenico et al. [156] reported the identification of mAbs, classified in class I, II, and III antibodies, binding three different epitopes of Psl, as suggested using competition antibody binding assays. The mAbs possessed opsonophagocytic killing activity and anti–cell attachment activity. In particular, class I mAb were shown to be the most functionally active and protective anti-Psl antibodies against P. aeruginosa [156]. The repeating unit of PsI of P. aeruginosa is the pentasaccharide shown in Figure 22, as determined by Kocharova et al. [157]. The chemical synthesis of different fragments of PsI, tetra-, penta-, hexa- and decasaccharide 190a–193a was reported starting from building blocks 194–199 (Figure 22) [158].
The synthetic strategy dealt with the stereoselective glycosylation of mannosides and the formation of two 1,2-cis mannosides, one of which is also extended at C-1, C-2, and C-3 in a crowded 1,2,3-cis configuration. In particular, 4,6-O-benzylidene protected mannosyl donors [159] 194 and 195, modified by a C-3 Nap ether and a C-2 silyl ether, respectively, provided optimal 1,2-cis stereoselectivity in the glycosylation reactions. On the other hand, mannosyl donor 196, functionalized with a participating acetyl ester at C-2, resulted suitable for the preparation of 1,2-trans mannosides. Compounds 190a–193a were used to identify the epitope requirements of monoclonal antibodies of class I, II, and III, showing some new insights about immune recognition of P. aeruginosa Psl exopolysaccharide [158]. Oligosaccharides 190a–193a were conjugated to BSA (190b–193b) to facilitate coating of ELISA plates followed by testing reactivity with an antibody that bound each epitope class. The class II mAb reacted potently with all oligosaccharides, suggesting that the epitope for this class resides within tetrasaccharide 190b and does not require the 1,2-cis mannoside of compound 191b. The class III antibody did not bind tetra- (190b) or pentasaccharide BSA-conjugate (191b). On the contrary, it showed weak affinity to glycoconjugate 193b and strong affinity to glycoconjugate 192b, suggesting that the terminal glucoside contained in glycoconjugate 192b is required for optimal binding. The class I antibody did not bind any of the oligosaccharides, suggesting the possibility that the class I mAb binds to a conformational epitope of PsI or to a substructure yet to be determined. The identification of this epitope could provide an attractive lead compound for the development of a synthetic Psl-based vaccine for P. aeruginosa.
12. Neisseria meningitidis
Neisseria meningitidis is a Gram-negative bacterium that colonizes the mucous membranes of humans. Meningococcal meningitis and sepsis are severe diseases that kill children and young adults within hours despite the availability of effective antibiotics. Mortality rates and permanent disability, like amputation, hearing loss and neurologic deficiency associated with N. meningitidis infections are high, even in countries where optimal health care practices are in place [160]. Among the 12 serogroups of N. meningitidis [161], serogroups B, C, Y and W cause approximately 90% of invasive meningococcal infections. Group A, however, is the only meningococcal serotype capable of causing of meningitis epidemics. Serogroups B and C express α-(2,8)- and α-(2,9)-linked polysialic acid, respectively. Alternating sequences of d-galactose or d-glucose and sialic acid are expressed by serogroups W and Y [162,163]. The serogroup A capsule is composed of α-(1,6)-linked N-acetyl-d-mannosamine-1-phosphate repeating units, partially acetylated at 3-OH (about 70%) and 4-OH (10–30%) [164]. First generation polysaccharide-based vaccines against N. meningitidis comprise the bivalent (groups A and C), the trivalent (groups A, C and W), and the tetravalent (groups A, C, Y and W) forms. Among second generation meningococcal glycoconjugate vaccines, three monovalent group C conjugate vaccines and one tetravalent meningococcal conjugate vaccine against groups A, C, Y and W are currently available. Of note, serogroup B (Men B) is not included in current formulations and remains a major cause of endemic meningitis in both developed and developing countries. The main obstacle for group B polysaccharides vaccine development is that the group B polysaccharide, composed of α-(2,8)-sialic acid polymers, is expressed in a number of human neurologic tissues since early fetal development. Men B CPS is therefore perceived as self-antigen by the innate immune system and it induces immune tolerance. Structural modification of this “self” antigen replacing the N-acetyl group of sialic acid units with an N-propanonyl group [165] induced high levels of bactericidal IgG antibodies without detection of autoantibodies [166]. However, its development has been suspended due to the poor performance of the vaccine in a limited human trial and to the high perceived risk of autoimmunity [61]. A new type of group B vaccine, Bexsero® (GlaxoSmithKline) developed by conjugation of three recombinant surface antigens (PorA, NadA and fHbp) and outer membrane vesicles from group B strain NZ98/254, is now licensed in more than 35 countries worldwide, including the EU, Australia, Brazil, Canada, Chile, Uruguay and the USA [167,168,169]. More recently, a new anti-MenB vaccine based on two recombinant lipidated factor H binding protein (Trumenba®, Pfizer) has been licensed by FDA and approved for use in EU countries in 2017.
12.1. N. meningitidis Serogroup A (MenA)
N. meningitidis serogroup A is most often implicated in seasonal epidemic diseases, especially in sub-Saharan Africa and asian developing countries [170,171]. The MenA CPS structure consists of (1→6)-linked 2-acetamido-2-deoxy-α-d-mannopyranosyl phosphate repeating units, with about 70% of O-acetylation at 3-OH (Figure 23).
The synthesis of MenA CPS fragments was reported in 2002 by Pozgay [173] and by Oscarson in 2005 [174], and upon conjugation to HSA the synthetic fragments were found to be immunogenic. MenA CPS, however, suffers from poor stability in water, due to the chemical lability of the phosphodiester linkages involving the anomeric position of each repeating unit. This issue stimulated the design of novel and hydrolytically stable analogues of MenA CPS repeating unit, like carbocyclic analogues (Figure 23) [172] and 1-C-phosphono analogues (Figure 24) [175,176], where a methylene group replaces the pyranose oxygen atom or the anomeric oxygen, respectively. The conformational behaviour of these analogues was investigated through DFT calculation and NMR spectroscopy [177], with a particular focus on the orientation of the phosphate or phosphonate aglycone and on the possibility of pyranose ring inversion [178]. The comparison between mimics and natural fragment showed the preservation of the 4C1 geometry in both classes of analogues. The synthesis of carbocyclic stabilized analogues of MenA CPS fragments was reported by the Lay group with the obtainment of monomer 200a, dimer 201a and trimer 202a of carba-N-acetylmannosamine-1-O-phosphate. The formation of the phosphodiester bridges was achieved through the use of the H-phosphonate methodology [179], followed by functionalization with a phosphodiester-linked aminopropyl spacer to allow protein conjugation. Oligomer synthesis was achieved starting from carbasugar 203, derived from compound 204. Carbocycle formation was carried out by Claisen rearrangement of glucal 205, in turn obtained from commercially available glucal 206 [172,177]. The inhibition abilities of the synthetic molecules were investigated by a competitive ELISA assay, showing that carba-disaccharide 201a is recognized by a polyclonal anti-MenA serum with an affinity similar to a native MenA oligosaccharide with average polymerization degree of 3 [172]. The conjugation of carbocyclic analogues 200a–202a to the protein carrier CRM197 gave glycoconjugates 200b–202b (Figure 23) that were tested for immunogenicity [180]. MenA fragments, produced by mild acid hydrolysis of native MenA polysaccharide (average degree of polymerization from 6 to 15) and conjugated to CRM197 were used to compare the activity of 200b–202b. Upon mice immunization, all glycoconjugates elicited antibodies that recognized the respective structures, although only conjugated trimer 202b was able to induce specific anti-MenA IgG antibodies with detectable in vitro bactericidal activity. Compound 202b, however, elicited antibodies to a lesser extent than hexamer and pentadecamer conjugated oligomers 207 and 208 obtained from hydrolysis of the native polysaccharide, suggesting that hydrolytically stable analogues of MenA CPS can be used for the development of vaccine and that conjugates with longer carbocyclic oligomers could further increase the induced immune response. In addition, a strategy for the multivalent presentation of carba analogues was developed [181] allowing conjugation of monomer 200a and dimer 201a to the metallic surface of superparamagnetic iron oxide nanoparticles (SPION) to generate 200c and 201c (Figure 23). SPIONs can act as multivalent carriers and as a contrast agent for magnetic resonance imaging (MRI) [182]. Functionalized SPIONs dispersed in aqueous media can aggregate into clusters inducing a reduction of T2 [183] and this event can be monitored as a decrease in brightness of a T2-weighted MR image [183,184]. This property has been widely used for ligand detection in biological media [185]. SPIONs 200c and 201c were produced as approximately spherical nanoparticles, with a size dispersion of 13 ± 3 nm and an average particle coating of 320 unities per nanoparticle for 200c and of 160 ligands per nanoparticle for 201c, as determined by transmission electron microscopy (TEM). Both 200c and 201c were able to bind the polyclonal anti-MenA antibody, as evaluated by MRI analysis, exploiting the magnetic peculiarity of SPIONs.
The synthesis of C-phosphono analogues of N. meningitidis group A CPS oligomers was reported by the Oscarson [176] and Lay [186] groups. In particular, an improved strategy for the synthesis of monosaccharides 209a–210a and phosphonoester-bridged fragments 211a–213a was recently reported [175] starting from compound 214, 215 and 216. The introduction of the phosphonate moiety was accomplished on alcohol 217, obtained from α-C-allenyl derivative 218 which, in turn, was prepared in six steps from orthoester intermediate 219 (Figure 24).
Competitive ELISA assay showed that monosaccharides 209a–210a and synthetic fragments 211a–213a containing the unnatural phosphonoester linkage were recognized by a human polyclonal anti-MenA serum [175]. The comparison with the inhibition of either MenA (positive control) or MenY (negative control) indicated that the chain lengths of the saccharide molecules is important for the efficacy, while the presence of the phosphonate residue (comparison between compounds 211a–213a and glycosides 209a–210a) and the orientation of the anomeric linker (comparison between compounds 211a and 212a) did not affect the affinity. Multivalent presentation on gold nanoparticles of monomer 209a, dimer 211a and trimer 213a were obtained (GNPs 220, 221 and 222 respectively, Figure 24) [187]. Interestingly, nanoparticles 220, 221 and 222 showed a more than three order of magnitude higher binding affinity than their counterparts not bound to the gold cluster 209a, 211a and 213a, at the same nominal concentration of saccharides. Fallarini et al. [188] used functionalized gold nanoparticles to test their ability to induce immune cell responses as a consequence of multivalency. In particular, monodisperse gold nanoparticles (2 and 5 nm) coated with mono- and disaccharides (220 and 221) were synthesized. Conjugation to gold nanoparticles conferred to the saccharides the ability to activate macrophages and this property is dependent on the size of the nanoparticles, with 5 nm nanoparticles giving comparable results to those obtained with the polysaccharide bacterium capsule (MenA) used as a natural antigen. Activation of macrophages occurred, independently of the saccharide oligomerization (or charge) on the nanoparticle surface. However, only nanoparticles 220, exposing a phosphonodisaccharide-functionalized monolayer, induced T cells proliferation and the increase of released interleukin-2 levels, the latter being a typical marker of T cell activation. Recently, HSA conjugates 209b, 211b and 213b [189] were shown to induce both T cell proliferation and interleukin-2 release in vitro, and to stimulate moderate specific IgG antibody production in vivo. All HSA-conjugated compounds 209b, 211b and 213b induced T cell proliferation (40% of proliferation at 102 μM), whereas only phosphonodisaccharide 211a was effective (28% of proliferation at 102 μM) among the unconjugated forms, showing the unusual behavior of triggering T cell proliferation in vitro and causing interleukin-2 release.
12.2. N. meningitidis Serogroup C (MenC)
N. meningitidis group C CSP is a α-(2,9)-polysialic acid with sporadic 7/8-O-acetylation (Figure 25). Non-acetylated fragments have been shown to be immunogenic and to elicit an immune response that is effective in recognizing and killing the bacterium [190]. A series of non-acetylated α-2,9-oligosialic acids of different length 223a–233a were prepared by a convergent synthetic route employing 5N,4O-oxazolidinone-protected phosphate-based building blocks 234 and acceptor 235 [191]. The dodecamer was synthesized with a [4+8] strategy that allowed to retain the α-selectivity even when the size of donor and acceptor increased.
In a further study, di-, tri-, tetra-, and penta-sialic acids 223a–226a were coupled with KLH carrier protein and studied for immunogenicity [192]. The glycoconjugates elicited robust T cell-mediated immunity in mice, in particular the immunogenicity of the tested oligo sialic acids increased in the order tri(224b) > di(223b) > tetra(225b) > penta(226b). The antibodies elicited were tested for efficacy and specificity, verifying if they could recognize and target group C N. meningitidis. All of the antisera obtained with the oligosaccharide-conjugates 223b–226b had strong binding to the N. meningitidis cell and no significant binding to cells not expressing α-2,9-poly and oligosialic acids (sialoglycans sTn, GM3, GM2, α-2,8-linked polysialic). The α-2,9-trisialic acid was identified as a promising antigen for developing glycoconjugate vaccines against group C Neisseria meningitidis. Recently, α-2,9-oligosialic acid fragments (di-, tri-, tetra-, and penta, 223a–226a) were conjugated with monophosphoryl lipid A (MPLA), a known immune potentiator (molecule with adjuvant activity) [193]. Immunological studies of the conjugates 223c–226c (Figure 25) in mice revealed that they elicited robust immune responses, mainly IgG2b and IgG2c, consistent with T cell dependent immunities. In particular, the immune response was comparable to the corresponding KLH-conjugates 223b–226b plus adjuvant. The immunogenicity of oligosialic acids decreased with elongated sugar chain, although all tested MPLA conjugates exhibited strong immune responses. The dimer 223c, actually, induced the highest titers of antigen-specific total and IgG2b antibodies, but some of the produced antibodies did not bind to oligosialic acids or bacterial CPS. In contrast, the tri- and tetra-sialic acid−MPLA conjugates 224c–225c, were fully effective showing the highest binding to bacterial cells and were identified as promising vaccine candidates worthy of further investigation.
12.3. N. meningitidis Serogroup W (MenW)
N. meningitidis serogroup W CPS consists of a glycan repeating unit of [→6)-α-d-Galp-(1→4)-α-d-Neup5Ac(7/9OAc)-(2→] (Figure 26). The synthesis of MenW CPS oligosaccharides of various lengths was performed by Wu group in 2013 [194], with the aim of studying the relationship between oligosaccharide length and immunogenicity.
Oligomers up to decasaccharides 236a–240a were obtained starting from protected disaccharides 241 and 242, bearing a N-acetyl-5-N,4-O-oxazolidinone (Figure 26), in turn synthesized from sialyl phosphate donor 243, functionalized with an oxazolidinone and a phosphate leaving group to increase the α-selectivity, and galactosides 244 and 245. Compound 243 was prepared from compound 246. Oligosaccharide elongation was accomplished by iterative glycosylation and deprotections, using disaccharide 241 for the [2+n] glycosylation reactions. After conjugation with CRM197, immunization of mice and glycan array analysis of produced antisera showed that antibodies induced by conjugated disaccharide 236b recognized only the disaccharide itself and did not cross react with longer oligomers. In contrast, antibodies induced by glycoconjugates 237b, 238b, 239b and 240b all recognized tetra- to decasaccharides, but not disaccharides. A serum bactericidal assay was used to study the bactericidal activity of the antibodies and showed that 236b did not induce antibodies with bactericidal activity. On the contrary, longer oligomers could and in particular, the tetramer 239b elicited antibodies with the highest bactericidal effect. Taken together, these data suggested that the tetrasaccharide is the minimum saccharide length required to induce bactericidal antibodies.
12.4. N. meningitidis Serogroup X (MenX)
Serotype X of N. meningitidis (MenX) emerged as a substantial threat to public health, especially in the “meningitis belt” area, after the introduction of MenA vaccine (MenAfriVac) and other conjugate vaccines (consisting of MenA, C, Y and W serotypes) that do not provide coverage for MenX serotype [195,196]. As a consequence, there is the need to extend the protection of current vaccines, developing more comprehensive conjugate vaccines [197]. The MenX CPS is a homopolymer of 2-acetamido-2-deoxy-α-d-glucopyranosyl phosphate moiety (Figure 27) [198,199].
In 2013 Morelli et al. reported the synthesis of monomer 247a, dimer 248a and trimer 249a of N. meningitidis X CPS [200], starting from intermediates 250–252, in turn derived from d-GlcNAc.
In 2014 an improvement of the synthesis of MenX fragments, their conjugation to CRM197 and the immunological evaluation of the resulting conjugates 247b–249b was reported [203]. Upon mice immunization, the conjugated trimer (249b) was found as the minimal fragment possessing immunogenic activity, although significantly lower than pentadecasaccharide-conjugate 253 obtained from the native polymer and used in the same study. This finding suggests that oligomers longer than three repeating units were possibly needed to mimic the activity of the native polysaccharide. The following year, Harale et al. reported an alternative synthesis of the tetrameric fragment of MenX CPS 254a, lacking the phosphate at the downstream end, starting from intermediates 251 and 225 in turn synthesized from monosaccharide 256 (Figure 27) [201]. Competitive ELISA experiment, using MenX bacterial CPS as a control, gave a concentration-dependent inhibition of anti-MenX antibodies from both compound 254a (unconjugated synthetic MenX tetramer) and tetramer-TT conjugate 254b. Lower inhibition for unconjugated fragment 254a (up to 68% inhibition) was observed compared to its conjugate form 254b (up to 89% inhibition) at all antigen concentrations tested. Also, bacterial MenX CPS showed higher inhibition than synthetic compounds at all respective concentrations used [201]. Very recently, an alternative strategy for MenX oligomer synthesis (Figure 27) was developed [202], based on an enzyme-catalyzed one-pot elongation of synthetic trimer 257a. Oligomers with predefined average length (compound 258a for general formula of avDP = 12) were synthesized and conjugated to CRM197. Mice immunized with 258b elicited functional antibodies comparable to controls immunized with the current MenX glycoconjugates prepared from the natural CPS or from its fragments enzymatically produced.
13. Mycobacterium tuberculosis
Tuberculosis (TB), caused by Mycobacterium tuberculosis (Mtb), remains one of the leading causes of death in the world. In 2016, some 1.7 million people died from the disease, with 64% cases of the total occurring in India, Indonesia, China, Philippines, Pakistan, Nigeria, and South Africa [204]. Moreover, it is estimated that one-third of the human population is latently infected with M. tuberculosis and is highly vulnerable if immunocompromised (in 2016, 40% of HIV deaths were due to TB). The mycobacterial cell wall is a highly complex structure largely composed of carbohydrates and lipids. Major components are lipidated polysaccharides, like the mycolyl−arabinogalactan complex, essentially composed of galactofuranose (Galf) and arabinofuranose (Araf) [205]. In recent years, numerous efforts have been done to develop inhibitors of UDP-galactopyranose mutase (UGM), an enzyme essential for the growth and survival of this mycobacterium [206,207,208,209,210,211,212]. Among the vital cell envelope components, phosphatidylinositol mannosides (PIMs) and their corresponding hyper-mannosylated derivatives (lipomannans and lipoarabinomannans) are noncovalently anchored to the plasma membrane and the outer capsule via their lipid chains. PIMs have a crucial role in the intracellular life of the bacterium, by binding to macrophages, to Toll-like receptors and C-type lectins [213,214], expressed on antigen presenting cell surfaces. PIMs also activate natural killer T cells for the production of interferon-γ [215]. Structurally, PIMs consist of a myo-inositol moiety linked with a diacylated glycerophospholipid unit and two α-mannosylation sites at O2 and O6 (Figure 28, compounds 259–261). When additional lipid chains are linked to the mannosyl-and myo-inositol-moieties, triacylated PIMs (AcPIMs) and tetraacylated PIMs (Ac2PIMs) are formed. Higher PIMs (for example, AcnPIM3 AcnPIM6) are formed by elongation at the mannose residue. In the course of the past few years, several synthesis of these structures have been reported [216,217,218,219,220,221]. In addition, heterocyclic analogues of PIMs in which the inositol ring is replaced by a piperidine or a tetrahydropyran moiety have been prepared and shown to retain the biological activity of the parent PIM structures [222].
Recently, the synthesis of a tetraacylated phosphatidylinositol hexamannoside (Ac2PIM6, 262) and its immunological evaluation have been reported [223]. Oligomer 262 was synthesized starting from pseudotrisaccharide 263, tetramannoside donor 264 and hydrogen phosphonate 265 (Figure 28). Compounds 263 and 264 were in turn obtained from monosaccharides 266, 267, 268 and 269, 270, respectively through a [1+1] strategy. The immunological evaluation was performed by observing the induction of antigen-specific antibodies in mice immunized with ovalbumin or tetanus toxoid adjuvanted with compound 262. The adjuvant effects of Alum or various PIMs isolated from M. tuberculosis strain H37Rv (iPIM1,2 and iPIM6) were also examined in parallel for comparison. Mice exposed to the synthesized Ac2PIM6 262 exhibited increased production of interleukin-4 and interferon-γ, suggesting proper activation of the innate immune system. Interestingly, 262 induced an approximately two to four-fold increase in the level of antigen specific antibodies, similarly to bacteria-derived PIMs and slightly lower than Alum.
Among M. tuberculosis cell wall polysaccharide and lipid components, some complex structures of arabinogalactan and lipoarabinomannan (LAM) have been synthesized in recent years [224,225]. In particular, Wang et al. [83] synthesized LAM oligosaccharides 271a–273a (Figure 29), that were conjugated to BSA (271b–273b) and KLH (271c–273c) and evaluated with immunological studies.
LAM tetra-, hepta-, and undecasaccharides 271a–273a, containing the α-1,5-, α-1,3-, and β-1,2-linked arabinan domain with the 5-OH of the upstream residues capped with the α-1,2-linked dimannose motif, were synthesized in good overall yields from d-arabinose in 10, 15, and 14 linear steps, respectively [83]. KLH conjugates 271c–273c and free oligosaccharides 271a–273a were injected in mice and the obtained antisera were analyzed by ELISA using BSA conjugates 271b–273b as capture antigens. Antisera derived from mice immunized with oligosaccharides 271a–273a did not contain carbohydrate antigen-specific antibodies, while all KLH conjugates 271c–273c elicited antigen-specific immune responses in mice. In particular, antibody titers induced by 271c and 273c were slightly higher than those induced by 272c. In a further work [226], the LAM tetrasaccharide was coupled to a monophosphoryl lipid A (MPLA) derivative generating a MPLA-based synthetic glycoconjugate 274 (Figure 29). In 274, the carbohydrate antigen was attached to the MPLA C-6′-position via an amide bond. Retrosynthetic analysis of 274 gives LAM tetrasaccharide 275, conveniently synthesized with a [2+2] strategy from 276 and 277 as reported before by the same authors [83], linker 278, and MPLA derivative 279. In turn, 279 was synthesized from fatty acids 280 and 281 and disaccharide 282, assembled from glycosyl donor 283 and acceptor 284. Immunological activity of conjugate 274 was evaluated in mice, affording robust IgG antibody responses. Since intraperitoneal injection elicited responses significantly stronger than those from subcutaneous injection, it was hypothesized that MPLA conjugates may stimulate B1 lymphocytes in the intrapleural and peritoneal cavities. These results revealed also the self-adjuvant properties of MPLA conjugates, paving the way to further investigation of these compounds as antituberculosis vaccine candidates.
14. Fungal Infections
Fungal infections can occur in healthy people, although immunosuppressed or immunocompromised patients are the major risk group for invasive fungal infections, as well as patients who use antibiotics able to modify the human microbiota [227]. Antifungal drugs are commercially available but they are limited compared to antibacterial drugs. In recent years, several strategies have been developed for the identification of new anti-fungal compounds, including components of plants, animals and microorganism [227]. In particular, vaccination offers promising alternative solutions for the treatment of fungal infections that are resistant to antibiotics. Recognition of fungi by the innate immune system depends on several Pathogen-Associated Molecular Patterns (PAMPs) in the fungal cell wall [228]. Specific receptors, exposed on antigen presenting cells surface, are involved in the recognition of polysaccharide cell wall components, like the mannose receptor (MR) and DC-SIGN for recognition of branched N-linked mannan [229], Toll-like receptor 4 (TLR4) for recognition of linear O-linked mannan [230], galectin 3 for β-mannosides, complement receptor 3 (CR3) for β-(1,6)-glucan, and dectin 1 and TLR2 for β-glucan and phospholipomannan [231]. Mannans consist of differently linked oligomannoside, and phospholipomannans are composed of phospholipids and mannans. The common linkages between mannose units in fungal mannans are α-(1,6), α-(1,2)-, α-(1,3) and β-(1,2), as shown in Figure 30. β-glucans are composed of β-d-glucose with β-(1,3) linkages and sporadic β-(1,6) branched points. Galactomannans consist of structurally diverse heteropolysaccharides composed of a poly-d-mannose backbone linked to galactofuranoside (Galf) units (Figure 30). Of note, the galactomannan of Aspergillus fumigatus is a specific carbohydrate antigen used for clinical detection of fungal infection.
In the course of the past few years, several synthetic strategies [232,233,234] have been developed to prepare these oligosaccharides expressed on the cell wall of various fungi, like Candida albicans and Apergillus fumigatus, as detailed in Section 14.1 and Section 14.2.
14.1. Candida albicans
The yeast Candida albicans is an opportunistic pathogenic microorganism, found in the normal microflora, skin and the mucosal surfaces of most healthy individuals. However, it is able to cause severe infections in immunocompromised individuals and patients undergoing immunosuppressive therapy [235]. The three major glycans expressed on the cell wall of C. albicans are phosphomannan-based glycoproteins in the outermost part of the cell wall and β-glucans and chitin, especially at the level of the bud scars (Figure 31) [228].
Being part of the phosphomannan glycoproteins, the acid-labile β-mannan is thought to be a major epitope of all C. Albicans serotypes and thus represents an ideal target for vaccine development. On this subject, an extensive work has been carried out by the Bundle group to develop a conjugate vaccine against C. albicans, starting from binding studies of β-mannans with two monoclonal protective antibodies, the mAb C3.1 and its immunoglobulin M (IgM) counterpart B6.1 [236]. By means of STD-NMR, computational analysis [237] and hydroxyl group replacement [238], the identification of key recognition elements was used for the development of antigens that elicit polyclonal protective antibodies, a process referred to as “reverse engineering” [236]. Initially, a set of β-(1-2)-mannan oligosaccharide propyl glycosides 285a–290a (Figure 32) ranging in size from di-to heptasaccharides were evaluated against mAbs C3.1 and B6.1. Interestingly, di and trisaccharides 285a and 286a had maximum binding capacity, while larger oligosaccharides were bound progressively more weakly [239]. This unusual pattern of inhibition was consistent with a binding site that could accommodate the trisaccharide, even though the primary polar contacts are located within the disaccharide. The synthesis of complementary mono-deoxy and mono-O-methyl analogues of dimers 285b, 291b–304b and trimers 286b and 305b (Figure 32) led to epitope mapping of anti-Candida albicans antibodies [240]. The strategy for the construction of these β-mannosides is based on the formation of β-glucosidic linkages, followed by epimerization at C-2 via an oxidation−reduction sequence.
Dimer 285c and trimer 286c (R = (CH2)3S(CH2)2NH2) were conjugated to chicken serum albumin (CSA) and the resulting glycoconjugates 285d and 286d (Figure 32) were able to generate highly specific IgG Abs titers in mice and rabbit [241]. The trisaccharide conjugated to CSA (286d) raised protective antibodies in rabbits [242]. Serum from rabbits immunized by 286d contained antibodies that stained C. albicans cells in fluorescent labeling studies more intensely than antibodies from rabbits immunized with 285d [105,241]. Both glycoconjugates 285d and 286d reduced C. albicans counts in vital organs but they were insufficient to provide 100% protection. In a further work, trisaccharide–TT conjugate 286e was found to be a stronger immunogen in rabbits, but it was poorly immunogenic in mice [241]. However, when the same trisaccharide was conjugated to different T cell peptides, found in cell wall proteins expressed during pathogenesis of human candidiasis, the resulting glycopeptides (286f) elicited a peptide- and carbohydrate-specific response that gave protection against challenge by C. albicans infection in mouse models [243]. All glycoconjugates showed immunogenicity with higher Abs titers compared to unconjugated 286c. When hyphal wall protein-1 (Hwp1), fructose-bisphosphate aldolase (Fba) and methyltetrahydropteroyltriglutamate (Met6) were used as T cell peptides for conjugation, the survival rate in mice challenge experiments was 80–100% against 40–80% survival with other glycopeptides.
The C. albicans cell wall consists of approximately 60% β-glucan. Although initially thought to be hidden underneath the mannoprotein layer, recent evidence suggest that β-glucans are exposed on the cell surface, possibly restricted to specific regions, such as bud scars [228]. β-glucans have also been investigated for their binding capacity to dectin-1 [244,245], a dendritic cell receptor that mediates phagocytosis and mediator production during inflammation caused by fungal pathogens [231]. Among β-glucans, Laminarin (Lam) is a polysaccharide extracted from Laminaria digitate plant. In 2010, Bromuro et al. [246] investigated the potential of laminarin as antigen for C. albicans, aimed at mediating antifungal protection. The authors showed that a synthetic linear structure composed of pentadecamer of Lam β-(1-3) repeating units (15mer) conjugated to CRM197 conferred protection against C. albicans. In the same study, a synthetic β-(1-6) branched 17mer conjugated to CRM197 was immunogenic but not protective [246], suggesting that the linear β-(1-3) fragment contains the protective epitope. To further investigate this aspect, Adamo et al. [247] reported the synthesis of short linear and β-(1-6) branched Lam fragments 306a–309a (Figure 33) and their immunological evaluation.
Compounds 306a–315a were synthesized starting from common building blocks 310, 311, 312 and 313 (Figure 33). Preliminary ELISA test with fragments 306a–309a showed that compound 309a was the best inhibitor of the binding between Lam and anti-Lam Abs elicited in mice by Lam-CRM197 conjugate, confirming the hypothesis that the linear β-(1-3) fragments contain the dominant epitope. Hexasaccharide 314b, prepared from building block 315 (Figure 33), was studied in order to identify the protective epitope. ELISA assays on compounds 306a–309a and 314b showed that hexasaccharide 314b provided 95% of inhibition at 4 mM concentration, while trisaccharide 309a and tetrasaccharide 307a provided 85% and 73% of inhibition at the same concentration, respectively. Based on these results, compound 314b was conjugated to CRM197 and employed for mice immunization. Sera analysis showed that glycoconjugate 314c was able to induce specific anti-Lam IgG Abs titers significantly higher and homogeneous compared to Lam-CRM197 conjugate. This result suggested that the linear hexasaccharide β-(1-3) glucan 314b is long enough to cover the dominant epitope of C. albicans.
In 2015, Liao et al. [248] reported the synthesis of longer β-(1-3) glucan chains such as fragments 316a–319a (Figure 34). Hexasaccharide 316a was assembled with a [4+2] glycosylation reaction from donor 320 and compound 321 after cleavage of 2-Naphthylmethyl ether (Nap) group. Longer oligomers 317a, 318a and 319a were obtained via [4+4] (from 320), [8+2] (from 322 and 321) and [8+4] (from 322 and 320) glycosylation strategies, respectively (Figure 34). Oligomers 316a–319a were conjugated to KLH and KLH conjugates (316b–319b) were injected in mice, raising high IgG1 titers with significant statistical difference between hexasaccharide 316b and longer oligomers 317b–319b. Octasaccharide 317b resulted the most immunogenic and protective compared to decasaccharide 318b and dodecasaccharide 319b. This result encouraged the authors to perform fungal challenge experiment in mice with glycoconjugate 317b using C. albicans fungus strain SC5314. In this experiment, 11 mice were immunized with 317b and, after challenge, 4 mice (about 34%) were unaffected, suggesting their protection from C. albicans.
More recently, Liao et al. [249] reported similar immunological studies of fragments 323a–325a containing the dominant linear octasaccharide and β-(1-6) and β-(1-3) branches with different chain length (Figure 35). KLH conjugates 323b and 324b were more immunogenic than 325b but the majority of elicited Abs were against the common β-glucan motif. In addition, conjugates 323b and 325b confer similar long-term protection against C. albicans infection (survival rate about 37%, 30 days after challenge for both conjugates). These data confirmed the hypothesis that linear β-(1-3) glucans of 6–8 units without branches can cover the dominant epitope of C. albicans.
14.2. Aspergillus fumigatus
Aspergillus fumigatus causes severe and usually fatal invasive aspergillosis infections in immunosuppressed hospitalized patients. More than 90% of the cell wall of A. fumigatus comprises polysaccharides, among which α-(1,3)-glucan is present in percentages variable from 20 to 40%. This virulence factor suffers of poor solubility in water, making difficult the study of its immunological properties. The use of shorter synthetic fragments could overcome the solubility issue. Indeed, Komarova et al. [250] reported the synthesis of pentasaccharide fragment 326a (Figure 36), the BSA-glycoconjugate (326b) and its evaluation as a vaccine candidate. Pentasaccharide 326a was prepared via a [3+2] glycosylation between donor 327 and acceptor 328. These protected fragments were in turn synthesized from monomers 329 and 330. The use of benzoyl group at 6-OH as remote partecipating group during glycosylation reactions facilitated the formation of α-linkages. Glycoconjugate 326b was immunized in mice, eliciting highly specific and protective polyclonal antibodies.
Galactomannan is a specific polysaccharide produced by A. fumigatus composed of a linear mannan core partially branched with β-(1,5)-galactofuranoside (Galf) units via β-(1,6) or β-(1,3) linkages (Figure 37) [251]. The length of β-(1,5)-Galf branches differs among different cultures, making difficult the identification of a common repeating unit for galactomannans. Recently, Kudoh et al. [252] demonstrated the presence of significant structural differences in both the O-linked and N-linked oligosaccharide of A. fumigatus galactomannans, depending on growth conditions in different culture media. Moreover, the authors revealed new structural elements of A. fumigatus galactomannan, the like the presence of β-(1,6)-linked Galf residues in addition to the β-(1,5)-linked Galf residues (Figure 37).
The galactofuranosides (Galf) are not presents in mammalian cells and they are therefore used for diagnosis of human infections. In addition, they can be advantageously used as antigen candidates for an anti A. fumigatus vaccine.
Recently, the Nifantiev group reported the synthesis of pentasaccharide fragments of the galactomannan containing the β-(1,5)-linked galactofuranoside chain attached to O-3 or O-6 of a mannopyranoside residue (GM-1 331a, GM-2 332a and GM-3 333a) [253] and fragments 334a–343a (Figure 38) [254]. In particular, pentasaccharide 331a was achieved via [2+3] glycosylation between disaccharide donor 344 and trisaccharide acceptor 345 (Figure 38), in turn prepared from donor 344 and mannosyl acceptor 346. Disaccharide 344 was synthesized from monosaccharides 347 and 348, both derived from allyl galactoside 350 through furanoside 349 (Figure 38).
The authors showed [255] that two mAbs (7B8 and 8G4), recognizing galactomannan in A. fumigatus, were elicited in mice after immunization with glycoconjugate 331b. Glycoarray analysis performed with synthetic biotinylated oligosaccharides 331c–343c and mAbs 7B8 and 8G4 showed high affinity towards pentasaccharide 331c and heptasaccharide 343c while no recognition was observed for mono- and disaccharides 334c–338c. Finally, confocal microscopy was used to evaluate the binding between the two mAbs and A. fumigatus and a series of other fungi and bacteria. mAbs 7B8 and 8G4 were selective for A. fumigatus, suggesting that they can be used as reagents for immune diagnostics. Recent advances in galactofuranoside synthesis [205,232,233] should facilitate the access to galactomannan fragments, thus opening the way to anti A. fumigatus vaccines based on synthetic Galf oligomers.
15. Conclusions
Pathogen surface glycans are promising vaccine targets due to their crucial role in adhesion to host tissues. Firstly, pathogen polysaccharides act as a protection barrier either by delaying the host’s immune response or by mimicking the host self-glycans. They are therefore essential for the survival of microorganisms in the blood and play a major role in their virulence. Secondly, most glycans are recognized by antigen presenting cells and triggers the innate immune response, starting the inflammatory process and eventually the antigen-specific adaptive immunity. Based on these considerations, the highly conserved polysaccharides (PS) exposed on pathogens cells surface are important virulence factors and have been used as antigens for vaccines development in a range of infectious diseases, including meningitis, pneumonia, otitis media, sepsis, infectious diarrheas, etc... The first-generation polysaccharide vaccines made of purified T cell-independent polysaccharide antigens have limited efficacy in infants, in the elderly and immunocompromised individuals and, in general, they don’t result in B cell-mediated immunological memory. The second-generation polysaccharide vaccines consist of native PS, produced and purified from natural sources and chemically conjugated to immunogenic carrier proteins able to elicit a T cell-dependent immune response (glycoconjugate vaccines). The purified native PS displays heterogeneity of chain length and may contain copurified endotoxin or other polysaccharides, introducing complications into the manufacturing process. Recent advances in carbohydrate chemistry has opened the way to a third-generation of polysaccharide vaccines, based on the use of synthetic oligosaccharides conjugated to carrier protein. The breakthrough in synthetic vaccines was carried out for H. influenzae type b in 2004, when the Vérez Bencomo research team in Cuba synthesized the first synthetic conjugate vaccine currently available under the trade name Quimi-Hib [9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62]. This approach is based on the hypothesis that protein conjugates of synthetic oligosaccharide fragments, smaller than the native PS, can elicit PS-specific and protective antibodies. The main challenge for the development of effective glycoconjugate vaccines is the determination of the carbohydrate antigen chain length and the sequence of the minimal protective immunogenic fragment of the antigen, a process often referred to as epitope mapping. The optimal carbohydrate antigen chain length needed for inclusion in a glycoconjugate vaccine, for example, cannot be predicted a priori for any given bacterial species. Despite the old paradigm established by Kabat [256], stating that immunogenic glycan epitopes usually comprise structures not longer than six-eight sugar units, the effect of the carbohydrate chain length on the immunogenicity of the corresponding protein conjugate is strictly case-dependent. There are many reported cases where both long chain glycans and oligosaccharides as short as tetrasaccharides have been shown to possess the minimal structural requirements for raising protective immunity, and several examples are illustrated throughout this review. In this regard, synthetic chemistry is a formidable tool to define the role of this crucial parameter, and more generally speaking of each structural variable affecting the immunogenicity and efficacy of vaccine candidates. Over the last years, much progress has been made. The rational design of carbohydrate antigens and the use of increasingly sophisticated synthetic strategies has made the preparation of even highly complex microbial glycans possible. In addition, the application of modern site-selective protein conjugation strategies [6,257,258] (a subject not addressed in the present review), has enabled the preparation of chemically defined glycoconjugate vaccine candidates featured by robust structure-immunogenicity relationship, which are expected to display improved safety and efficacy profiles [259]. These new achievements have certainly opened up new perspectives and hope for the prevention of severe bacterial and fungal infections still affecting the pediatric population worldwide. We truly wish that these promising results translate into new and efficient vaccines in the next few years.
Author Contributions
C.C., O.P., and L.L. wrote the paper.
Funding
We gratefully acknowledge funding from the H2020-MSCA-ITN-2015 “Glycovax” under Grant agreement No. 675671.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vaccination Greatly Reduces Disease, Disability, Death and Inequity Worldwide. Available online: http://www.who.int/bulletin/volumes/86/2/07-040089/en/ (accessed on 28 February 2018).
- Bentley, S.D.; Aanensen, D.M.; Mavroidi, A.; Saunders, D.; Rabbinowitsch, E.; Collins, M.; Donohoe, K.; Harris, D.; Murphy, L.; Quail, M.A.; et al. Genetic analysis of the capsular biosynthetic locus from all 90 pneumococcal serotypes. PLoS Genet. 2006, 2, e31. [Google Scholar] [CrossRef] [PubMed]
- Avci, F.Y.; Li, X.; Tsuji, M.; Kasper, D.L. A novel mechanism for glycoconjugate vaccine activation of the adaptive immune system. Nat. Med. 2011, 17, 1602–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avci, F.Y.; Kasper, D.L. How bacterial carbohydrates influence the adaptive immune system. Annu. Rev. Immunol. 2010, 28, 107–130. [Google Scholar] [CrossRef] [PubMed]
- Berti, F.; Adamo, R. Recent mechanistic insights on glycoconjugate vaccines and future perspectives. ACS Chem. Biol. 2013, 8, 1653–1663. [Google Scholar] [CrossRef] [PubMed]
- Adamo, R.; Nilo, A.; Castagner, B.; Boutureira, O.; Berti, F.; Bernardes, G.J.L. Synthetically defined glycoprotein vaccines: Current status and future directions. Chem. Sci. 2013, 4, 2995–3008. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, R.W.; Oaks, E.V. Inactivated and subunit vaccines to prevent shigellosis. Expert Rev. Vaccines 2009, 8, 1693–1704. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.; Wierzba, T.; Walker, R.I. Status of vaccine research and development for shigella. Vaccine 2016, 34, 2887–2894. [Google Scholar] [CrossRef] [PubMed]
- Verez-Bencomo, V.; Fernández-Santana, V.; Hardy, E.; Toledo, M.E.; Rodríguez, M.C.; Heynngnezz, L.; Rodriguez, A.; Baly, A.; Herrera, L.; Izquierdo, M.; et al. A synthetic conjugate polysaccharide vaccine against haemophilus influenzae type b. Science 2004, 305, 522–525. [Google Scholar] [CrossRef] [PubMed]
- Astronomo, R.D.; Burton, D.R. Carbohydrate vaccines: Developing sweet solutions to sticky situations? Nat. Rev. Drug Discov. 2010, 9, 308–324. [Google Scholar] [CrossRef] [PubMed]
- Morelli, L.; Poletti, L.; Lay, L. Carbohydrates and immunology: Synthetic oligosaccharide antigens for vaccine formulation. Eur. J. Org. Chem. 2011, 2011, 5723–5777. [Google Scholar] [CrossRef]
- The Symbolic Representation of Monosaccharides in the Age of Glycobiology. Available online: https://glycopedia.eu/References-102 (accessed on 4 April 2018).
- Pozsgay, V. Synthesis of glycoconjugate vaccines against shigella dysenteriae type 1. J. Org. Chem. 1998, 63, 5983–5999. [Google Scholar] [CrossRef] [PubMed]
- Pozsgay, V. Synthetic shigella vaccines: A carbohydrate–protein conjugate with totally synthetic hexadecasaccharide haptens. Angew. Chem. Int. Ed. 1998, 37, 138–142. [Google Scholar] [CrossRef]
- Pozsgay, V.; Kubler-Kielb, J.; Schneerson, R.; Robbins, J.B. Effect of the nonreducing end of shigella dysenteriae type 1 o-specific oligosaccharides on their immunogenicity as conjugates in mice. Proc. Natl. Acad. Sci. USA 2007, 104, 14478–14482. [Google Scholar] [CrossRef] [PubMed]
- Bélot, F.; Wright, K.; Costachel, C.; Phalipon, A.; Mulard, L.A. Blockwise approach to fragments of the o-specific polysaccharide of shigella flexneri serotype 2a: Convergent synthesis of a decasaccharide representative of a dimer of the branched repeating unit1. J. Org. Chem. 2004, 69, 1060–1074. [Google Scholar] [CrossRef] [PubMed]
- Belot, F.; Guerreiro, C.; Baleux, F.; Mulard, L.A. Synthesis of two linear padre conjugates bearing a deca- or pentadecasaccharide b epitope as potential synthetic vaccines against shigella flexneri serotype 2a infection. Chem. Eur. J. 2005, 11, 1625–1635. [Google Scholar] [CrossRef] [PubMed]
- Van der Put, R.M.; Kim, T.H.; Guerreiro, C.; Thouron, F.; Hoogerhout, P.; Sansonetti, P.J.; Westdijk, J.; Stork, M.; Phalipon, A.; Mulard, L.A. A synthetic carbohydrate conjugate vaccine candidate against shigellosis: Improved bioconjugation and impact of alum on immunogenicity. Bioconjug. Chem. 2016, 27, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Phalipon, A.; Costachel, C.; Grandjean, C.; Thuizat, A.; Guerreiro, C.; Tanguy, M.; Nato, F.; Vulliez-Le Normand, B.; Belot, F.; Wright, K.; et al. Characterization of functional oligosaccharide mimics of the shigella flexneri serotype 2a o-antigen: Implications for the development of a chemically defined glycoconjugate vaccine. J. Immunol. 2006, 176, 1686–1694. [Google Scholar] [CrossRef] [PubMed]
- Phalipon, A.; Tanguy, M.; Grandjean, C.; Guerreiro, C.; Belot, F.; Cohen, D.; Sansonetti, P.J.; Mulard, L.A. A synthetic carbohydrate-protein conjugate vaccine candidate against shigella flexneri 2a infection. J. Immunol. 2009, 182, 2241–2247. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, C.; Chassagne, P.; Theillet, F.X.; Guerreiro, C.; Thouron, F.; Nato, F.; Delepierre, M.; Sansonetti, P.J.; Phalipon, A.; Mulard, L.A. Non-stoichiometric o-acetylation of shigella flexneri 2a o-specific polysaccharide: Synthesis and antigenicity. Org. Biomol. Chem. 2014, 12, 4218–4232. [Google Scholar] [CrossRef] [PubMed]
- Berti, F.; De Ricco, R.; Rappuoli, R. Role of o-acetylation in the immunogenicity of bacterial polysaccharide vaccines. Molecules 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Hlozek, J.; Kuttel, M.M.; Ravenscroft, N. Conformations of neisseria meningitidis serogroup a and x polysaccharides: The effects of chain length and o-acetylation. Carbohydr. Res. 2018, 465, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Mulard, L.A. Bacterial polysaccharides as major surface antigens: Interest in o-acetyl substitutions. In Carbohydrate Chemistry: Chemical and Biological Approaches Volume 43; RSC Publishing: London, UK, 2018; Volume 43, pp. 71–103. [Google Scholar]
- Rebeaud, F.; Bachmann, M.F. Immunization strategies for clostridium difficile infections. Expert Rev. Vaccines 2012, 11, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.; Bauer, M.P.; Baines, S.D.; Corver, J.; Fawley, W.N.; Goorhuis, B.; Kuijper, E.J.; Wilcox, M.H. The changing epidemiology of clostridium difficile infections. Clin. Microbiol. Rev. 2010, 23, 529–549. [Google Scholar] [CrossRef] [PubMed]
- Péchiné, S.; Janoir, C.; Collignon, A. Emerging monoclonal antibodies against clostridium difficile infection. Expert Opin. Biol. Ther. 2017, 17, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Posteraro, B.; Pea, F.; Masucci, L.; Posteraro, P.; Sanguinetti, M. Actoxumab + bezlotoxumab combination: What promise for clostridium difficile treatment? Expert Opin. Biol. Ther. 2018, 18, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.E.; Broecker, F.; Oberli, M.A.; Komor, J.; Mattner, J.; Anish, C.; Seeberger, P.H. Immunological evaluation of a synthetic clostridium difficile oligosaccharide conjugate vaccine candidate and identification of a minimal epitope. J. Am. Chem. Soc. 2013, 135, 9713–9722. [Google Scholar] [CrossRef] [PubMed]
- Broecker, F.; Hanske, J.; Martin, C.E.; Baek, J.Y.; Wahlbrink, A.; Wojcik, F.; Hartmann, L.; Rademacher, C.; Anish, C.; Seeberger, P.H. Multivalent display of minimal clostridium difficile glycan epitopes mimics antigenic properties of larger glycans. Nat. Commun. 2016, 7, 11224. [Google Scholar] [CrossRef] [PubMed]
- Wojcik, F.; O’Brien Alexander, G.; Götze, S.; Seeberger Peter, H.; Hartmann, L. Synthesis of carbohydrate-functionalised sequence-defined oligo(amidoamine)s by photochemical thiolene coupling in a continuous flow reactor. Chem. Eur. J. 2013, 19, 3090–3098. [Google Scholar] [CrossRef] [PubMed]
- Oberli, M.A.; Hecht, M.-L.; Bindschädler, P.; Adibekian, A.; Adam, T.; Seeberger, P.H. A possible oligosaccharide-conjugate vaccine candidate for clostridium difficile is antigenic and immunogenic. Chem. Biol. 2011, 18, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Danieli, E.; Lay, L.; Proietti, D.; Berti, F.; Costantino, P.; Adamo, R. First synthesis of c. Difficile ps-ii cell wall polysaccharide repeating unit. Org. Lett. 2011, 13, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Adamo, R.; Romano, M.R.; Berti, F.; Leuzzi, R.; Tontini, M.; Danieli, E.; Cappelletti, E.; Cakici, O.S.; Swennen, E.; Pinto, V.; et al. Phosphorylation of the synthetic hexasaccharide repeating unit is essential for the induction of antibodies to clostridium difficile psii cell wall polysaccharide. ACS Chem. Biol. 2012, 7, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Meumann, E.M.; Cheng, A.C.; Ward, L.; Currie, B.J. Clinical features and epidemiology of melioidosis pneumonia: Results from a 21-year study and review of the literature. Clin. Infect. Dis. 2012, 54, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Dance, D. Treatment and prophylaxis of melioidosis. Int. J. Antimicrob. Agents 2014, 43, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.E.; Christ, W.J.; George, A.J.; Stokes, M.G.; Lohman, G.J.; Guo, Y.; Jones, M.; Titball, R.W.; Atkins, T.P.; Campbell, A.S.; et al. Protection against experimental melioidosis with a synthetic manno-heptopyranose hexasaccharide glycoconjugate. Bioconjug. Chem. 2016, 27, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Conejero, L.; De Reynal, M.; Easton, A.; Bancroft, G.J.; Titball, R.W. Development of vaccines against burkholderia pseudomallei. Front. Microbiol. 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.B.; Dow, S.W. Development of burkholderia mallei and pseudomallei vaccines. Front. Cell. Infect. Microbiol. 2013, 3, 10. [Google Scholar] [CrossRef] [PubMed]
- Burtnick, M.N.; Shaffer, T.L.; Ross, B.N.; Muruato, L.A.; Sbrana, E.; DeShazer, D.; Torres, A.G.; Brett, P.J. Development of subunit vaccines that provide high-level protection and sterilizing immunity against acute inhalational melioidosis. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [PubMed]
- Reckseidler, S.L.; DeShazer, D.; Sokol, P.A.; Woods, D.E. Detection of bacterial virulence genes by subtractive hybridization: Identification of capsular polysaccharide of burkholderia pseudomallei as a major virulence determinant. Infect. Immun. 2001, 69, 34–44. [Google Scholar] [CrossRef] [PubMed]
- DeShazer, D.; Waag, D.M.; Fritz, D.L.; Woods, D.E. Identification of a burkholderia mallei polysaccharide gene cluster by subtractive hybridization and demonstration that the encoded capsule is an essential virulence determinant. Microb. Pathog. 2001, 30, 253–269. [Google Scholar] [CrossRef] [PubMed]
- Boschiroli, M.-L.; Foulongne, V.; O’Callaghan, D. Brucellosis: A worldwide zoonosis. Curr. Opin. Microbiol. 2001, 4, 58–64. [Google Scholar] [CrossRef]
- Mantur, B.; Akki, A.; Mangalgi, S.S.; Patil, S.; Gobbur, R.; Peerapur, B. Childhood brucellosis—A microbiological, epidemiological and clinical study. J. Trop. Pediatr. 2004, 50, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Mantur, B.G.; Biradar, M.S.; Bidri, R.C.; Mulimani, M.S.; Veerappa, K.; Kariholu, P.; Patil, S.B.; Mangalgi, S.S. Protean clinical manifestations and diagnostic challenges of human brucellosis in adults: 16 years’ experience in an endemic area. J. Med. Microbiol. 2006, 55, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Moreno, E.; Cloeckaert, A.; Moriyón, I. Brucella evolution and taxonomy. Vet. Microbiol. 2002, 90, 209–227. [Google Scholar] [CrossRef]
- Cloeckaert, A.; Grayon, M.; Grépinet, O.; Boumedine, K.S. Classification of brucella strains isolated from marine mammals by infrequent restriction site-PCR and development of specific PCR identification tests. Microb. Infect. 2003, 5, 593–602. [Google Scholar] [CrossRef]
- Roth, F.; Zinsstag, J.; Orkhon, D.; Chimed-Ochir, G.; Hutton, G.; Cosivi, O.; Carrin, G.; Otte, J. Human health benefits from livestock vaccination for brucellosis: Case study. Bull. World Health Organ. 2003, 81, 867–876. [Google Scholar] [PubMed]
- Bundle, D.R.; McGiven, J. Brucellosis: Improved diagnostics and vaccine insights from synthetic glycans. Acc. Chem. Res. 2017, 50, 2958–2967. [Google Scholar] [CrossRef] [PubMed]
- Zygmunt, M.S.; Bundle, D.R.; Ganesh, N.V.; Guiard, J.; Cloeckaert, A. Monoclonal antibody-defined specific c epitope of brucella o-polysaccharide revisited. Clin. Vaccine Immunol. 2015, 22, 979–982. [Google Scholar] [CrossRef] [PubMed]
- Smits, H.L.; Basahi, M.A.; Díaz, R.; Marrodan, T.; Douglas, J.T.; Rocha, A.; Veerman, J.; Zheludkov, M.M.; Witte, O.W.M.; de Jong, J.; et al. Development and evaluation of a rapid dipstick assay for serodiagnosis of acute human brucellosis. J. Clin. Microbiol. 1999, 37, 4179–4182. [Google Scholar] [PubMed]
- Clavijo, E.; Díaz, R.; Anguita, A.; García, A.; Pinedo, A.; Smits, H.L. Comparison of a dipstick assay for detection of brucella-specific immunoglobulin m antibodies with other tests for serodiagnosis of human brucellosis. Clin. Diagn. Lab. Immunol. 2003, 10, 612–615. [Google Scholar] [CrossRef] [PubMed]
- Kubler-Kielb, J.; Vinogradov, E. Reinvestigation of the structure of brucella o-antigens. Carbohydr. Res. 2013, 378, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Guiard, J.; Paszkiewicz, E.; Sadowska, J.; Bundle, D.R. Design and synthesis of a universal antigen to detect brucellosis. Angew. Chem. Int. Ed. Engl. 2013, 52, 7181–7185. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K. Diagnosis of brucellosis by serology. Vet. Microbiol. 2002, 90, 447–459. [Google Scholar] [CrossRef]
- Ganesh, N.V.; Sadowska, J.M.; Sarkar, S.; Howells, L.; McGiven, J.; Bundle, D.R. Molecular recognition of brucella a and m antigens dissected by synthetic oligosaccharide glycoconjugates leads to a disaccharide diagnostic for brucellosis. J. Am. Chem. Soc. 2014, 136, 16260–16269. [Google Scholar] [CrossRef] [PubMed]
- McGiven, J.; Howells, L.; Duncombe, L.; Stack, J.; Ganesh, N.V.; Guiard, J.; Bundle, D.R. Improved serodiagnosis of bovine brucellosis by novel synthetic oligosaccharide antigens representing the capping m epitope elements of brucella o-polysaccharide. J. Clin. Microbiol. 2015, 53, 1204–1210. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.S.; Duncombe, L.; Ganesh, N.V.; Sarkar, S.; Howells, L.; Hogarth, P.J.; Bundle, D.R.; McGiven, J. Novel solutions for vaccines and diagnostics to combat brucellosis. ACS Cent. Sci. 2017, 3, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Murphy, T.F. Haemophilus influenzae infections in the h. Influenzae type b conjugate vaccine era. J. Clin. Microbiol. 2011, 49, 3728–3732. [Google Scholar] [CrossRef] [PubMed]
- Peltola, H.; Käythy, H.; Sivonen, A.; Mäkelä, P.H. Haemophilus influenzae type b capsular polysaccharide vaccine in children: A double-blind field study of 100,000 vaccinees 3 months to 5 years of age in finland. Pediatrics 1977, 60, 730–737. [Google Scholar] [PubMed]
- Shivatare, S.S.; Wu, C.-Y. Carbohydrate-based antibacterial vaccines: Current progress and future outlook. In Carbohydrate Chemistry: State of the Art and Challenges for Drug Development: An Overview on Structure, Biological Roles, Synthetic Methods and Application as Therapeutics; World Scientific Publishers: Singapore, 2015; pp. 321–356. [Google Scholar]
- Fernández-Santana, V.; Cardoso, F.; Rodriguez, A.; Carmenate, T.; Peña, L.; Valdés, Y.; Hardy, E.; Mawas, F.; Heynngnezz, L.; Rodríguez, M.C.; et al. Antigenicity and immunogenicity of a synthetic oligosaccharide-protein conjugate vaccine against haemophilus influenzae type b. Infect. Immun. 2004, 72, 7115–7123. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.Y.; Geissner, A.; Rathwell, D.C.K.; Meierhofer, D.; Pereira, C.L.; Seeberger, P.H. A modular synthetic route to size-defined immunogenic haemophilus influenzae b antigens is key to the identification of an octasaccharide lead vaccine candidate. Chem. Sci. 2018, 9, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- Miyaji, E.N.; Oliveira, M.L.; Carvalho, E.; Ho, P.L. Serotype-independent pneumococcal vaccines. Cell. Mol. Life Sci. 2013, 70, 3303–3326. [Google Scholar] [CrossRef] [PubMed]
- Trotter, C.L.; Waight, P.; Andrews, N.J.; Slack, M.; Efstratiou, A.; George, R.; Miller, E. Epidemiology of invasive pneumococcal disease in the pre-conjugate vaccine era: England and wales, 1996–2006. J. Infect. 2010, 60, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Pelton, S.I.; Loughlin, A.M.; Marchant, C.D. Seven valent pneumococcal conjugate vaccine immunization in two boston communities: Changes in serotypes and antimicrobial susceptibility among streptococcus pneumoniae isolates. Pediatr. Infect. Dis. J. 2004, 23, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Seeberger, P.H.; Pereira, C.L.; Govindan, S. Total synthesis of a streptococcus pneumoniae serotype 12f cps repeating unit hexasaccharide. Beilstein J. Org. Chem. 2017, 13, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Perciani, C.T.; Barazzone, G.C.; Goulart, C.; Carvalho, E.; Cabrera-Crespo, J.; Gonçalves, V.M.; Leite, L.C.C.; Tanizaki, M.M. Conjugation of polysaccharide 6b from streptococcus pneumoniae with pneumococcal surface protein a: Pspa conformation and its effect on the immune response. Clin. Vaccine Immunol. 2013, 20, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cui, L.; Lipinski, T.; Bundle, D.R. Synthesis of monomeric and dimeric repeating units of the zwitterionic type 1 capsular polysaccharide from streptococcus pneumoniae. Chem. Eur. J. 2010, 16, 3476–3488. [Google Scholar] [CrossRef] [PubMed]
- Christina, A.E.; van den Bos, L.J.; Overkleeft, H.S.; van der Marel, G.A.; Codée, J.D.C. Galacturonic acid lactones in the synthesis of all trisaccharide repeating units of the zwitterionic polysaccharide sp1. J. Org. Chem. 2011, 76, 1692–1706. [Google Scholar] [CrossRef] [PubMed]
- Schumann, B.; Pragani, R.; Anish, C.; Pereira, C.L.; Seeberger, P.H. Synthesis of conjugation-ready zwitterionic oligosaccharides by chemoselective thioglycoside activation. Chem. Sci. 2014, 5, 1992–2002. [Google Scholar] [CrossRef]
- Schumann, B.; Reppe, K.; Kaplonek, P.; Wahlbrink, A.; Anish, C.; Witzenrath, M.; Pereira, C.L.; Seeberger, P.H. Development of an efficacious, semisynthetic glycoconjugate vaccine candidate against streptococcus pneumoniae serotype 1. ACS Cent. Sci. 2018, 4, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, N.; Singh, M.; Das, R.R.; Jindal, I.; Agarwal, A.; Thumburu, K.K.; Kumar, A.; Chauhan, A. Distribution of serotypes, vaccine coverage, and antimicrobial susceptibility pattern of streptococcus pneumoniae in children living in saarc countries: A systematic review. PLoS ONE 2014, 9, e108617. [Google Scholar] [CrossRef] [PubMed]
- Gaensbauer, J.T.; Asturias, E.J.; Soto, M.; Holt, E.; Olson, D.; Halsey, N.A.; Guatemala Pediatric Bacterial Surveillance Working Group. Pediatric invasive pneumococcal disease in guatemala city: Importance of serotype 2. Pediatr. Infect. Dis. J. 2016, 35, e139–e143. [Google Scholar] [CrossRef] [PubMed]
- Jansson, P.-E.; Lindberg, B.; Andersson, M.; Lindquist, U.; Henrichsen, J. Structural studies of the capsular polysaccharide from streptococcus pneumoniae type 2, a reinvestigation. Carbohydr. Res. 1988, 182, 111–117. [Google Scholar] [CrossRef]
- Emmadi, M.; Khan, N.; Lykke, L.; Reppe, K.; Parameswarappa, S.G.; Lisboa, M.P.; Wienhold, S.M.; Witzenrath, M.; Pereira, C.L.; Seeberger, P.H. A streptococcus pneumoniae type 2 oligosaccharide glycoconjugate elicits opsonic antibodies and is protective in an animal model of invasive pneumococcal disease. J. Am. Chem. Soc. 2017, 139, 14783–14791. [Google Scholar] [CrossRef] [PubMed]
- Poolman, J.; Kriz, P.; Feron, C.; Di-Paolo, E.; Henckaerts, I.; Miseur, A.; Wauters, D.; Prymula, R.; Schuerman, L. Pneumococcal serotype 3 otitis media, limited effect of polysaccharide conjugate immunisation and strain characteristics. Vaccine 2009, 27, 3213–3222. [Google Scholar] [CrossRef] [PubMed]
- Poolman, J.; Borrow, R. Hyporesponsiveness and its clinical implications after vaccination with polysaccharide or glycoconjugate vaccines. Expert Rev. Vaccines 2011, 10, 307–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truck, J.; Thompson, A.; Morales-Aza, B.; Clutterbuck, E.A.; Voysey, M.; Clarke, E.; Snape, M.D.; Kelly, D.F.; Finn, A.; Pollard, A.J. Memory b cell response to a PCV-13 booster in 3.5year old children primed with either PCV-7 or PCV-13. Vaccine 2017, 35, 2701–2708. [Google Scholar] [CrossRef] [PubMed]
- Benaissa-Trouw, B.; Lefeber, D.J.; Kamerling, J.P.; Vliegenthart, J.F.; Kraaijeveld, K.; Snippe, H. Synthetic polysaccharide type 3-related di-, tri-, and tetrasaccharide-CRM(197) conjugates induce protection against streptococcus pneumoniae type 3 in mice. Infect. Immun. 2001, 69, 4698–4701. [Google Scholar] [CrossRef] [PubMed]
- Parameswarappa, S.G.; Reppe, K.; Geissner, A.; Menova, P.; Govindan, S.; Calow, A.D.J.; Wahlbrink, A.; Weishaupt, M.W.; Monnanda, B.P.; Bell, R.L.; et al. A semi-synthetic oligosaccharide conjugate vaccine candidate confers protection against streptococcus pneumoniae serotype 3 infection. Cell Chem. Biol. 2016, 23, 1407–1416. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Feng, S.; Qiao, Y.; Guo, Z.; Gu, G. Synthesis and immunological studies of oligosaccharides that consist of the repeating unit of streptococcus pneumoniae serotype 3 capsular polysaccharide. Chem. Eur. J. 2018, 24, 8205–8216. [Google Scholar] [CrossRef]
- Wang, L.; Feng, S.; An, L.; Gu, G.; Guo, Z. Synthetic and immunological studies of mycobacterial lipoarabinomannan oligosaccharides and their protein conjugates. J. Org. Chem. 2015, 80, 10060–10075. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.L.; Geissner, A.; Anish, C.; Seeberger, P.H. Chemical synthesis elucidates the immunological importance of a pyruvate modification in the capsular polysaccharide of streptococcus pneumoniae serotype 4. Angew. Chem. Int. Ed. 2015, 54, 10016–10019. [Google Scholar] [CrossRef] [PubMed]
- Geissner, A.; Pereira, C.L.; Leddermann, M.; Anish, C.; Seeberger, P.H. Deciphering antigenic determinants of streptococcus pneumoniae serotype 4 capsular polysaccharide using synthetic oligosaccharides. ACS Chem. Biol. 2016, 11, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Sucher, A.J.; Chahine, E.B.; Nelson, M.; Sucher, B.J. Prevnar 13, the new 13-valent pneumococcal conjugate vaccine. Ann. Pharmacother. 2011, 45, 1516–1524. [Google Scholar] [CrossRef] [PubMed]
- Jansson, P.-E.; Lindberg, B.; Lindquist, U. Structural studies of the capsular polysaccharide from streptococcus pneumoniae type 5. Carbohydr. Res. 1985, 140, 101–110. [Google Scholar] [CrossRef]
- Mistretta, N.; Danve, E.; Moreau, M. Conjugates Obtained by Reductive Amination of the Pneumococcus Serotype 5 Capsular Polysaccharide. US Patent 7812006B2, 12 October 2010. [Google Scholar]
- Lisboa, M.P.; Khan, N.; Martin, C.; Xu, F.-F.; Reppe, K.; Geissner, A.; Govindan, S.; Witzenrath, M.; Pereira, C.L.; Seeberger, P.H. Semisynthetic glycoconjugate vaccine candidate against streptococcus pneumoniae serotype 5. Proc. Natl. Acad. Sci. USA 2017, 114, 11063–11068. [Google Scholar] [CrossRef] [PubMed]
- Ardanuy, C.; de la Campa, A.G.; García, E.; Fenoll, A.; Calatayud, L.; Cercenado, E.; Pérez-Trallero, E.; Bouza, E.; Liñares, J. Spread of streptococcus pneumoniae serotype 8-ST63 multidrug-resistant recombinant clone, spain. Emerg. Infect. Dis. 2014, 20, 1848–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz, J.C.; Cercenado, E.; Marín, M.; Ramos, B.; Ardanuy, C.; Rodríguez-Avial, I.; Bouza, E. Multidrug-resistant pneumococci (serotype 8) causing invasive disease in hiv+ patients. Clin. Microbiol. Infect. 2011, 17, 1094–1098. [Google Scholar] [CrossRef] [PubMed]
- Schumann, B.; Hahm, H.S.; Parameswarappa, S.G.; Reppe, K.; Wahlbrink, A.; Govindan, S.; Kaplonek, P.; Pirofski, L.-A.; Witzenrath, M.; Anish, C.; et al. A semisynthetic streptococcus pneumoniae serotype 8 glycoconjugate vaccine. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Yano, M.; Pirofski, L.-A. Characterization of gene use and efficacy of mouse monoclonal antibodies to streptococcus pneumoniae serotype 8. Clin. Vaccine Immunol. 2011, 18, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Hao, T.; Ou, S.; Hu, C.; Chen, L. Applications and perspectives of nanomaterials in novel vaccine development. MedChemComm 2018, 9, 226–238. [Google Scholar] [CrossRef]
- Compostella, F.; Pitirollo, O.; Silvestri, A.; Polito, L. Glyco-gold nanoparticles: Synthesis and applications. Beilstein J. Org. Chem. 2017, 13, 1008–1021. [Google Scholar] [CrossRef] [PubMed]
- Safari, D.; Marradi, M.; Chiodo, F.; Th Dekker, H.A.; Shan, Y.; Adamo, R.; Oscarson, S.; Rijkers, G.T.; Lahmann, M.; Kamerling, J.P. Gold nanoparticles as carriers for a synthetic streptococcus pneumoniae type 14 conjugate vaccine. Nanomedicine 2012, 7, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, B.; Lonngren, J.; Powell, D.A. Structural studies on the specific type-14 pneumococcal polysaccharide. Carbohydr. Res. 1977, 58, 177–186. [Google Scholar] [CrossRef]
- Vetro, M.; Safari, D.; Fallarini, S.; Salsabila, K.; Lahmann, M.; Penadés, S.; Lay, L.; Marradi, M.; Compostella, F. Preparation and immunogenicity of gold glyco-nanoparticles as antipneumococcal vaccine model. Nanomedicine 2017, 12, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Mawas, F.; Niggemann, J.; Jones, C.; Corbel, M.J.; Kamerling, J.P.; Vliegenthart, J.F.G. Immunogenicity in a mouse model of a conjugate vaccine made with a synthetic single repeating unit of type 14 pneumococcal polysaccharide coupled to CRM197. Infect. Immun. 2002, 70, 5107–5114. [Google Scholar] [CrossRef] [PubMed]
- Safari, D.; Dekker, H.A.; Joosten, J.A.; Michalik, D.; de Souza, A.C.; Adamo, R.; Lahmann, M.; Sundgren, A.; Oscarson, S.; Kamerling, J.P. Identification of the smallest structure capable of evoking opsonophagocytic antibodies against streptococcus pneumoniae type 14. Infect. Immun. 2008, 76, 4615–4623. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, E.; Khitri, M.; Lay, L.; Nicotra, F.; Panza, L.; Russo, G. Capsular polysaccharide of streptococcus pneumoniae type 19f: Synthesis of the repeating unit. Carbohydr. Res. 1998, 311, 171–181. [Google Scholar] [CrossRef]
- Legnani, L.; Ronchi, S.; Fallarini, S.; Lombardi, G.; Campo, F.; Panza, L.; Lay, L.; Poletti, L.; Toma, L.; Ronchetti, F.; et al. Synthesis, molecular dynamics simulations, and biology of a carba-analogue of the trisaccharide repeating unit of streptococcus pneumoniae19f capsular polysaccharide. Org. Biomol. Chem. 2009, 7, 4428–4436. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.J.; Barnett, T.C.; McArthur, J.D.; Cole, J.N.; Gillen, C.M.; Henningham, A.; Sriprakash, K.S.; Sanderson-Smith, M.L.; Nizet, V. Disease manifestations and pathogenic mechanisms of group a streptococcus. Clin. Microbiol. Rev. 2014, 27, 264–301. [Google Scholar] [CrossRef] [PubMed]
- Kabanova, A.; Margarit, I.; Berti, F.; Romano, M.R.; Grandi, G.; Bensi, G.; Chiarot, E.; Proietti, D.; Swennen, E.; Cappelletti, E.; et al. Evaluation of a group a streptococcus synthetic oligosaccharide as vaccine candidate. Vaccine 2010, 29, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Van Sorge, N.M.; Cole, J.N.; Kuipers, K.; Henningham, A.; Aziz, R.K.; Kasirer-Friede, A.; Lin, L.; Berends, E.T.M.; Davies, M.R.; Dougan, G.; et al. The classical lancefield antigen of group a streptococcus is a virulence determinant with implications for vaccine design. Cell Host Microbe 2014, 15, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Galvin, J.E.; Hemric, M.E.; Ward, K.; Cunningham, M.W. Cytotoxic mab from rheumatic carditis recognizes heart valves and laminin. J. Clin. Investig. 2000, 106, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Kirvan, C.A.; Swedo, S.E.; Heuser, J.S.; Cunningham, M.W. Mimicry and autoantibody-mediated neuronal cell signaling in sydenham chorea. Nat. Med. 2003, 9, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Malkiel, S.; Liao, L.; Cunningham, M.W.; Diamond, B. T-cell-dependent antibody response to the dominant epitope of streptococcal polysaccharide, n-acetyl-glucosamine, is cross-reactive with cardiac myosin. Infect. Immun. 2000, 68, 5803–5808. [Google Scholar] [CrossRef] [PubMed]
- Henningham, A.; Davies, M.R.; Uchiyama, S.; van Sorge, N.M.; Lund, S.; Chen, K.T.; Walker, M.J.; Cole, J.N.; Nizet, V. Virulence role of the glcnac side chain of the lancefield cell wall carbohydrate antigen in non-m1-serotype group a streptococcus. MBio 2018, 9. [Google Scholar] [CrossRef]
- Auzanneau, F.I.; Borrelli, S.; Pinto, B.M. Synthesis and immunological activity of an oligosaccharide-conjugate as a vaccine candidate against group a streptococcus. Bioorg. Med. Chem. Lett. 2013, 23, 6038–6042. [Google Scholar] [CrossRef] [PubMed]
- Reimer, K.B.; Gidney, M.A.J.; Bundle, D.R.; Pinto, B.M. Immunochemical characterization of polyclonal and monoclonal streptococcus group a antibodies by chemically defined glycoconjugates and synthetic oligosaccharides. Carbohydr. Res. 1992, 232, 131–142. [Google Scholar] [CrossRef]
- Johnson, M.A.; Pinto, B.M. Saturation transfer difference 1d-tocsy experiments to map the topography of oligosaccharides recognized by a monoclonal antibody directed against the cell-wall polysaccharide of group a streptococcus. J. Am. Chem. Soc. 2002, 124, 15368–15374. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.J.; Rench, M.A.; Edwards, M.S.; Carpenter, R.J.; Hays, B.M.; Kasper, D.L. Immunization of pregnant women with a polysaccharide vaccine of group b streptococcus. N. Engl. J. Med. 1988, 319, 1180–1185. [Google Scholar] [CrossRef] [PubMed]
- Heath, P.T.; Schuchat, A. Perinatal group b streptococcal disease. Best Pract. Res. Clin. Obstet. Gynaecol. 2007, 21, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Skoff, T.H.; Farley, M.M.; Petit, S.; Craig, A.S.; Schaffner, W.; Gershman, K.; Harrison, L.H.; Lynfield, R.; Mohle-Boetani, J.; Zansky, S. Increasing burden of invasive group b streptococcal disease in nonpregnant adults, 1990–2007. Clin. Infect. Dis. 2009, 49, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Nuccitelli, A.; Rinaudo, C.D.; Maione, D. Group b streptococcus vaccine: State of the art. Ther. Adv. Vaccines 2015, 3, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Mondal, P.K.; Liao, G.; Mondal, M.A.; Guo, Z. Chemical synthesis of the repeating unit of type ia group b streptococcus capsular polysaccharide. Org. Lett. 2015, 17, 1102–1105. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Guo, Z. Chemical synthesis of the repeating unit of type v group b streptococcus capsular polysaccharide. Org. Lett. 2016, 18, 5552–5555. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Zhang, H.; Li, Y.; Gu, G.; Cai, F.; Guo, Z.; Gao, J. Chemical synthesis of the repeating unit of type ii group b streptococcus capsular polysaccharide. J. Org. Chem. 2018, 83, 5920–5930. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.J.; Rench, M.A.; McInnes, P. Immunization of pregnant women with group b streptococcal type iii capsular polysaccharide-tetanus toxoid conjugate vaccine. Vaccine 2003, 21, 3468–3472. [Google Scholar] [CrossRef]
- Demchenko, A.V.; Boons, G.-J. A highly convergent synthesis of a complex oligosaccharide derived from group b type iii streptococcus. J. Org. Chem. 2001, 66, 2547–2554. [Google Scholar] [CrossRef] [PubMed]
- Pozsgay, V.; Brisson, J.-R.; Jennings, H.J. Synthesis of a tri-and a tetra-saccharide fragment of the capsular polysaccharide of type iii group b streptococcus. Carbohydr. Res. 1990, 205, 133–146. [Google Scholar] [CrossRef]
- Demchenko, A.V.; Boons, G.-J. A novel and versatile glycosyl donor for the preparation of glycosides of n-acetylneuraminic acid. Tetrahedron Lett. 1998, 39, 3065–3068. [Google Scholar] [CrossRef]
- Demchenko, A.; Boons, G.-J. A highly convergent synthesis of a hexasaccharide derived from the oligosaccharide of group b type iii streptococcus. Tetrahedron Lett. 1997, 38, 1629–1632. [Google Scholar] [CrossRef]
- Jennings, H.J.; Lugowski, C.; Kasper, D.L. Conformational aspects critical to the immunospecificity of the type iii group b streptococcal polysaccharide. Biochemistry 1981, 20, 4511–4518. [Google Scholar] [CrossRef] [PubMed]
- González-Outeiriño, J.; Kadirvelraj, R.; Woods, R.J. Structural elucidation of type iii group b streptococcus capsular polysaccharide using molecular dynamics simulations: The role of sialic acid. Carbohydr. Res. 2005, 340, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, V.; Carboni, F.; Oldrini, D.; De Ricco, R.; Donadio, N.; Ros, I.M.Y.; Berti, F.; Adamo, R. Synthesis of group b streptococcus type iii polysaccharide fragments for evaluation of their interactions with monoclonal antibodies. Pure Appl. Chem. 2017, 89, 855–875. [Google Scholar] [CrossRef]
- Weekly Epidemiological Record. 2018, 93, 153–172. Available online: http://www.who.int/wer/2018/wer9313/en/ (accessed on 27 May 2018).
- Britto, C.; Pollard, A.J.; Voysey, M.; Blohmke, C.J. An appraisal of the clinical features of pediatric enteric fever: Systematic review and meta-analysis of the age-stratified disease occurrence. Clin. Infect. Dis. 2017, 64, 1604–1611. [Google Scholar] [CrossRef] [PubMed]
- Wijedoru, L.; Mallett, S.; Parry, C.M. Rapid diagnostic tests for typhoid and paratyphoid (enteric) fever. Cochrane Database Syst. Rev. 2017, CD008892. [Google Scholar] [CrossRef] [PubMed]
- Olopoenia, L.A.; King, A.L. Widal agglutination test—100 years later: Still plagued by controversy. Postgrad. Med. J. 2000, 76, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Crump, J.A.; Sjölund-Karlsson, M.; Gordon, M.A.; Parry, C.M. Epidemiology, clinical presentation, laboratory diagnosis, antimicrobial resistance, and antimicrobial management of invasive salmonella infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Qadri, A. Vi polysaccharide of salmonella typhi targets the prohibitin family of molecules in intestinal epithelial cells and suppresses early inflammatory responses. Proc. Natl. Acad. Sci. USA 2004, 101, 17492–17497. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, S.K.; Dutta, D.; Parween, F.; Qadri, A. The virulence polysaccharide vi released by salmonella typhi targets membrane prohibitin to inhibit t-cell activation. J. Infect. Dis. 2014, 210, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Virlogeux-Payant, I.; Popoff, M.Y. The vi antigen of salmonella typhi. Bull. Inst. Pasteur 1996, 94, 237–250. [Google Scholar] [CrossRef]
- Felix, A.; Pitt, R.M. The vi antigens of various salmonella types. Br. J. Exp. Pathol. 1936, 17, 81–86. [Google Scholar]
- Hu, X.; Chen, Z.; Xiong, K.; Wang, J.; Rao, X.; Cong, Y. Vi capsular polysaccharide: Synthesis, virulence, and application. Crit. Rev. Microbiol. 2017, 43, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Ermin, S.; Petr, D.; Marian, H. Role of salmonella typhi vi antigen and secretory systems on immune response. Curr. Pharm. Des. 2016, 22, 6251–6260. [Google Scholar] [CrossRef]
- Szu, S.C.; Li, X.R.; Stone, A.L.; Robbins, J.B. Relation between structure and immunologic properties of the vi capsular polysaccharide. Infect. Immun. 1991, 59, 4555–4561. [Google Scholar] [PubMed]
- Kiow Shi-Shun, L.; Mallet, J.-M.; Moreau, M.; Sinaÿ, P. Synthèse d’oligomères du polysaccharide capsulaire de salmonella typhi, bactérie à l’origine de la fièvre typhoïde. Tetrahedron 1999, 55, 14043–14068. [Google Scholar] [CrossRef]
- Yang, L.; Zhu, J.; Zheng, X.J.; Tai, G.; Ye, X.S. A highly alpha-stereoselective synthesis of oligosaccharide fragments of the vi antigen from salmonella typhi and their antigenic activities. Chem. Eur. J. 2011, 17, 14518–14526. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.-L.; Yang, L.; Zhu, J.; Wei, M.; Yan, W.; Xiong, D.-C.; Ye, X.-S. Synthesis and antigenic evaluation of oligosaccharide mimics of vi antigen from salmonella typhi. Chem. Eur. J. 2017, 23, 10670–10677. [Google Scholar] [CrossRef]
- Khan, M.I.; Soofi, S.B.; Ochiai, R.L.; Habib, M.A.; Sahito, S.M.; Nizami, S.Q.; Acosta, C.J.; Clemens, J.D.; Bhutta, Z.A.; DOMI Typhoid Karachi Vi Effectiveness Study Group. Effectiveness of vi capsular polysaccharide typhoid vaccine among children: A cluster randomized trial in karachi, pakistan. Vaccine 2012, 30, 5389–5395. [Google Scholar] [CrossRef] [PubMed]
- Szu, S.C. Development of vi conjugate—A new generation of typhoid vaccine. Expert Rev. Vaccines 2013, 12, 1273–1286. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Rondini, S.; Pisoni, I.; Proietti, D.; Berti, F.; Costantino, P.; Rappuoli, R.; Szu, S.; Saul, A.; Martin, L.B. Vi-CRM197 as a new conjugate vaccine against salmonella typhi. Vaccine 2011, 29, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Gibani, M.M.; Moore, M.; Juel, H.B.; Jones, E.; Meiring, J.; Harris, V.; Gardner, J.; Nebykova, A.; Kerridge, S.A.; et al. Efficacy and immunogenicity of a vi-tetanus toxoid conjugate vaccine in the prevention of typhoid fever using a controlled human infection model of salmonella typhi: A randomised controlled, phase 2b trial. Lancet 2017, 390, 2472–2480. [Google Scholar] [CrossRef]
- Fusari, M.; Fallarini, S.; Lombardi, G.; Lay, L. Synthesis of di- and tri-saccharide fragments of salmonella typhi vi capsular polysaccharide and their zwitterionic analogues. Bioorg. Med. Chem. 2015, 23, 7439–7447. [Google Scholar] [CrossRef] [PubMed]
- Kerr, K.G.; Snelling, A.M. Pseudomonas aeruginosa: A formidable and ever-present adversary. J. Hosp. Infect. 2009, 73, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Pier, G.B. The challenges and promises of new therapies for cystic fibrosis. J. Exp. Med. 2012, 209, 1235–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Deng, Z.; Yan, A. Bacterial multidrug efflux pumps: Mechanisms, physiology and pharmacological exploitations. Biochem. Biophys. Res. Commun. 2014, 453, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Kunz, A.N.; Brook, I. Emerging resistant gram-negative aerobic bacilli in hospital-acquired infections. Chemotherapy 2010, 56, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Oleksiewicz, M.B.; Nagy, G.; Nagy, E. Anti-bacterial monoclonal antibodies: Back to the future? Arch. Biochem. Biophys. 2012, 526, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Thaden, J.T.; Keller, A.E.; Shire, N.J.; Camara, M.M.; Otterson, L.; Huband, M.; Guenther, C.M.; Zhao, W.; Warrener, P.; Stover, C.K.; et al. Pseudomonas aeruginosa bacteremic patients exhibit nonprotective antibody titers against therapeutic antibody targets pcrv and psl exopolysaccharide. J. Infect. Dis. 2016, 213, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Conover, M.; Lu, H.; Parsek, M.R.; Bayles, K.; Wozniak, D.J. Assembly and development of the pseudomonas aeruginosa biofilm matrix. PLoS Pathog. 2009, 5, e1000354. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wang, S.; Wang, D.; Parsek, M.R.; Wozniak, D.J. The roles of biofilm matrix polysaccharide psl in mucoid pseudomonas aeruginosa biofilms. FEMS Immunol. Med. Microbiol. 2012, 65, 377–380. [Google Scholar] [CrossRef] [PubMed]
- DiGiandomenico, A.; Warrener, P.; Hamilton, M.; Guillard, S.; Ravn, P.; Minter, R.; Camara, M.M.; Venkatraman, V.; MacGill, R.S.; Lin, J.; et al. Identification of broadly protective human antibodies to pseudomonas aeruginosa exopolysaccharide psl by phenotypic screening. J. Exp. Med. 2012, 209, 1273–1287. [Google Scholar] [CrossRef] [PubMed]
- Kocharova, N.A.; Knirel, Y.A.; Shashkov, A.S.; Kochetkov, N.K.; Pier, G.B. Structure of an extracellular cross-reactive polysaccharide from pseudomonas aeruginosa immunotype 4. J. Biol. Chem. 1988, 263, 11291–11295. [Google Scholar] [PubMed]
- Li, H.; Mo, K.F.; Wang, Q.; Stover, C.K.; DiGiandomenico, A.; Boons, G.J. Epitope mapping of monoclonal antibodies using synthetic oligosaccharides uncovers novel aspects of immune recognition of the psl exopolysaccharide of pseudomonas aeruginosa. Chem. Eur. J. 2013, 19, 17425–17431. [Google Scholar] [CrossRef] [PubMed]
- Crich, D.; Sun, S. Direct chemical synthesis of β-mannopyranosides and other glycosides via glycosyl triflates. Tetrahedron 1998, 54, 8321–8348. [Google Scholar] [CrossRef]
- Rosenstein, N.E.; Perkins, B.A.; Stephens, D.S.; Popovic, T.; Hughes, J.M. Meningococcal disease. N. Engl. J. Med. 2001, 344, 1378–1388. [Google Scholar] [CrossRef] [PubMed]
- Harrison, O.B.; Claus, H.; Jiang, Y.; Bennett, J.S.; Bratcher, H.B.; Jolley, K.A.; Corton, C.; Care, R.; Poolman, J.T.; Zollinger, W.D.; et al. Description and nomenclature of neisseria meningitidis capsule locus. Emerg. Infect. Dis. 2013, 19, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.Y.; Gotschlich, E.C.; Dunne, F.T.; Jonssen, E.K. Studies on the meningococcal polysaccharides: Ii. Composition and chemical properties of the group b and group c polysaccharide. J. Biol. Chem. 1971, 246, 4703–4712. [Google Scholar] [PubMed]
- Bhattacharjee, A.K.; Jennings, H.J.; Kenny, C.P.; Martin, A.; Smith, I.C. Structural determination of the polysaccharide antigens of neisseria meningitidis serogroups Y, W-135, and BO. Can. J. Biochem. 1976, 54, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.-Y.; Gotschlich, E.C.; Jonssen, E.K.; Wysocki, J.R. Studies on the meningococcal polysaccharides: I. Composition and chemical properties of the group a polysaccharide. J. Biol. Chem. 1971, 246, 2849–2858. [Google Scholar] [PubMed]
- Pon, R.A.; Lussier, M.; Yang, Q.-L.; Jennings, H.J. N-propionylated group b meningococcal polysaccharide mimics a unique bactericidal capsular epitope in group b neisseria meningitidis. J. Exp. Med. 1997, 185, 1929–1938. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.; Farley, E.K.; Fusco, P.C.; Michon, F. Specificity of the immune response to a modified group b meningococcal polysaccharide conjugate vaccine. Clin. Vaccine Immunol. 2007, 14, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Tirani, M.; Meregaglia, M.; Melegaro, A. Health and economic outcomes of introducing the new menb vaccine (bexsero) into the italian routine infant immunisation programme. PLoS ONE 2015, 10, e0123383. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Findlow, J.; Borrow, R. Recombinant protein meningococcal serogroup b vaccine combined with outer membrane vesicles. Expert Opin Biol Ther 2011, 11, 969–985. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, M.M.; Adu-Bobie, J.; Comanducci, M.; Aricò, B.; Savino, S.; Santini, L.; Brunelli, B.; Bambini, S.; Biolchi, A.; Capecchi, B.; et al. A universal vaccine for serogroup b meningococcus. Proc. Natl. Acad. Sci. USA 2006, 103, 10834–10839. [Google Scholar] [CrossRef] [PubMed]
- Coulson, G.B.; von Gottberg, A.; du Plessis, M.; Smith, A.M.; de Gouveia, L.; Klugman, K.P.; for the Group for Enteric, Respiratory; Meningeal Disease Surveillance in South Africa; Germs, S.A. Meningococcal disease in Siouth Afrca, 1999–2002. Emerg. Infect. Dis. 2007, 13, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Borrow, R.; Lee, J.-S.; Vázquez, J.A.; Enwere, G.; Taha, M.-K.; Kamiya, H.; Kim, H.M.; Jo, D.S. Meningococcal disease in the asia-pacific region: Findings and recommendations from the global meningococcal initiative. Vaccine 2016, 34, 5855–5862. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Zaccaria, C.; Tontini, M.; Poletti, L.; Costantino, P.; Lay, L. Synthesis and preliminary biological evaluation of carba analogues from neisseria meningitidis a capsular polysaccharide. Org. Biomol. Chem. 2012, 10, 6673–6681. [Google Scholar] [CrossRef] [PubMed]
- Berkin, A.; Coxon, B.; Pozsgay, V. Towards a synthetic glycoconjugate vaccine against neisseria meningitidis a. Chem. Eur. J. 2002, 8, 4424–4433. [Google Scholar] [CrossRef]
- Slattegard, R.; Teodorovic, P.; Kinfe, H.H.; Ravenscroft, N.; Gammon, D.W.; Oscarson, S. Synthesis of structures corresponding to the capsular polysaccharide of neisseria meningitidis group a. Org. Biomol. Chem. 2005, 3, 3782–3787. [Google Scholar] [CrossRef] [PubMed]
- Torres-Sanchez, M.I.; Zaccaria, C.; Buzzi, B.; Miglio, G.; Lombardi, G.; Polito, L.; Russo, G.; Lay, L. Synthesis and biological evaluation of phosphono analogues of capsular polysaccharide fragments from neisseria meningitidis a. Chem. Eur. J. 2007, 13, 6623–6635. [Google Scholar] [CrossRef] [PubMed]
- Teodorovic, P.; Slattegard, R.; Oscarson, S. Synthesis of stable c-phosphonate analogues of neisseria meningitidis group a capsular polysaccharide structures using modified mitsunobu reaction conditions. Org. Biomol. Chem. 2006, 4, 4485–4490. [Google Scholar] [CrossRef] [PubMed]
- Toma, L.; Legnani, L.; Rencurosi, A.; Poletti, L.; Lay, L.; Russo, G. Modeling of synthetic phosphono and carba analogues of N-acetyl-α-D-mannosamine 1-phosphate, the repeating unit of the capsular polysaccharide from neisseria meningitidis serovar a. Org. Biomol. Chem. 2009, 7, 3734–3740. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Barbero, J.; Calloni, I.; Unione, L.; Jimenez-Oses, G.; Corzana, F.; Del Bino, L.; Corrado, A.; Pitirollo, O.; Colombo, C.; Lay, L.; et al. The conformation of the mannopyranosyl phosphate repeating unit of the capsular polysaccharide of Neisseria meningitides serogroup A and its carba-mimetic. Eur. J. Org. Chem. 2018, in press. [Google Scholar] [CrossRef]
- Nikolaev, A.V.; Botvinko, I.V.; Ross, A.J. Natural phosphoglycans containing glycosyl phosphate units: Structural diversity and chemical synthesis. Carbohydr. Res. 2007, 342, 297–344. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Tontini, M.; Brogioni, G.; Nilo, A.; Filippini, S.; Harfouche, C.; Polito, L.; Romano, M.R.; Costantino, P.; Berti, F.; et al. Immunoactivity of protein conjugates of carba analogues from neisseria meningitidis a capsular polysaccharide. ACS Chem. Biol. 2013, 8, 2561–2567. [Google Scholar] [CrossRef] [PubMed]
- Ramella, D.; Polito, L.; Mazzini, S.; Ronchi, S.; Scaglioni, L.; Marelli, M.; Lay, L. A strategy for multivalent presentation of carba analogues fromn. Meningitidisa capsular polysaccharide. Eur. J. Org. Chem. 2014, 2014, 5915–5924. [Google Scholar] [CrossRef]
- Neuberger, T.; Schöpf, B.; Hofmann, H.; Hofmann, M.; von Rechenberg, B. Superparamagnetic nanoparticles for biomedical applications: Possibilities and limitations of a new drug delivery system. J. Magn. Magn. Mater. 2005, 293, 483–496. [Google Scholar] [CrossRef]
- Chavhan, G.B.; Babyn, P.S.; Thomas, B.; Shroff, M.M.; Haacke, E.M. Principles, techniques, and applications of t2*-based mr imaging and its special applications. Radiographics 2009, 29, 1433–1449. [Google Scholar] [CrossRef] [PubMed]
- Bitar, R.; Leung, G.; Perng, R.; Tadros, S.; Moody, A.R.; Sarrazin, J.; McGregor, C.; Christakis, M.; Symons, S.; Nelson, A.; et al. MR pulse sequences: What every radiologist wants to know but is afraid to ask. RadioGraphics 2006, 26, 513–537. [Google Scholar] [CrossRef] [PubMed]
- Jedlovszky-Hajdú, A.; Bombelli, F.B.; Monopoli, M.P.; Tombácz, E.; Dawson, K.A. Surface coatings shape the protein corona of spions with relevance to their application in vivo. Langmuir 2012, 28, 14983–14991. [Google Scholar] [CrossRef] [PubMed]
- Torres-Sánchez, M.I.; Draghetti, V.; Panza, L.; Lay, L.; Russo, G. Synthesis of the phosphono analogue of the dimeric subunit of neisseria meningitidis type a capsular polysaccharide. Synlett 2005, 2005, 1147–1151. [Google Scholar] [CrossRef]
- Manea, F.; Bindoli, C.; Fallarini, S.; Lombardi, G.; Polito, L.; Lay, L.; Bonomi, R.; Mancin, F.; Scrimin, P. Multivalent, saccharide-functionalized gold nanoparticles as fully synthetic analogs of type a neisseria meningitidis antigens. Adv. Mater. 2008, 20, 4348–4352. [Google Scholar] [CrossRef]
- Fallarini, S.; Paoletti, T.; Battaglini, C.O.; Ronchi, P.; Lay, L.; Bonomi, R.; Jha, S.; Mancin, F.; Scrimin, P.; Lombardi, G. Factors affecting t cell responses induced by fully synthetic glyco-gold-nanoparticles. Nanoscale 2013, 5, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Fallarini, S.; Buzzi, B.; Giovarruscio, S.; Polito, L.; Brogioni, G.; Tontini, M.; Berti, F.; Adamo, R.; Lay, L.; Lombardi, G. A synthetic disaccharide analogue from neisseria meningitidis a capsular polysaccharide stimulates immune cell responses and induces immunoglobulin g production in mice when protein-conjugated. ACS Infect. Dis. 2015, 1, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Richmond, P.; Borrow, R.; Findlow, J.; Martin, S.; Thornton, C.; Cartwright, K.; Miller, E. Evaluation of de-o-acetylated meningococcal c polysaccharide-tetanus toxoid conjugate vaccine in infancy: Reactogenicity, immunogenicity, immunologic priming, and bactericidal activity against o-acetylated and de-o-acetylated serogroup c strains. Infect. Immun. 2001, 69, 2378–2382. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.C.; Ren, C.T.; Lu, C.P.; Hsu, C.H.; Sun, T.H.; Han, J.L.; Pal, B.; Chao, T.A.; Lin, Y.F.; Wu, S.H.; et al. Efficient and stereoselective synthesis of alpha(2→9) oligosialic acids: From monomers to dodecamers. Angew. Chem. Int. Ed. Engl. 2011, 50, 9391–9395. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Zhou, Z.; Guo, Z. Synthesis and immunological study of alpha-2,9-oligosialic acid conjugates as anti-group c meningitis vaccines. Chem. Commun. (Camb.) 2015, 51, 9647–9650. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Zhou, Z.; Suryawanshi, S.; Mondal, M.A.; Guo, Z. Fully synthetic self-adjuvanting alpha-2,9-oligosialic acid based conjugate vaccines against group c meningitis. ACS Cent. Sci. 2016, 2, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Li, S.T.; Lin, T.L.; Cheng, Y.Y.; Sun, T.H.; Wang, J.T.; Cheng, T.J.R.; Mong, K.K.T.; Wong, C.H.; Wu, C.Y. Synthesis of neisseria meningitidis serogroup W135 capsular oligosaccharides for immunogenicity comparison and vaccine development. Angew. Chem. Int. Ed. 2013, 52, 9157–9161. [Google Scholar] [CrossRef] [PubMed]
- Delrieu, I.; Yaro, S.; Tamekloé, T.A.S.; Njanpop-Lafourcade, B.-M.; Tall, H.; Jaillard, P.; Ouedraogo, M.S.; Badziklou, K.; Sanou, O.; Drabo, A.; et al. Emergence of epidemic neisseria meningitidis serogroup x meningitis in togo and burkina faso. PLoS ONE 2011, 6, e19513. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Yao, P.-P.; Zhang, L.-Y.; Li, Y.; Xu, F.; Mei, L.-L.; Zhu, S.-R.; Zhang, Y.-J.; Zhu, H.-P.; van der Veen, S. Capsule switching of neisseria meningitidis sequence type 7 serogroup a to serogroup x. J. Infect. 2017, 75, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Romano, M.R.; Tontini, M.; Cappelletti, E.; Gavini, M.; Proietti, D.; Rondini, S.; Swennen, E.; Santini, L.; Filippini, S.; et al. Development of a glycoconjugate vaccine to prevent meningitis in africa caused by meningococcal serogroup x. Proc. Natl. Acad. Sci. USA 2013, 110, 19077–19082. [Google Scholar] [CrossRef] [PubMed]
- Bundle, D.R.; Smith, I.C.P.; Jennings, H.J. Determination of the structure and conformation of bacterial polysaccharides by carbon 13 nuclear magnetic resonance: Studies on the group-specific antigens of neisseria meningitidis serogroups a and x. J. Biol. Chem. 1974, 249, 2275–2281. [Google Scholar] [PubMed]
- Xie, O.; Bolgiano, B.; Gao, F.; Lockyer, K.; Swann, C.; Jones, C.; Delrieu, I.; Njanpop-Lafourcade, B.-M.; Tamekloe, T.A.; Pollard, A.J.; et al. Characterization of size, structure and purity of serogroup x neisseria meningitidis polysaccharide, and development of an assay for quantification of human antibodies. Vaccine 2012, 30, 5812–5823. [Google Scholar] [CrossRef] [PubMed]
- Morelli, L.; Lay, L. Synthesis of neisseria meningitidis x capsular polysaccharide fragments. ARKIVOC Online J. Org. Chem. 2013, 166–184. [Google Scholar]
- Harale, K.R.; Dumare, N.B.; Singh, D.; Misra, A.K.; Chhikara, M.K. Synthesis of a tetrasaccharide and its glycoconjugate corresponding to the capsular polysaccharide of neisseria meningitidis serogroup x and its immunochemical studies. RSC Adv. 2015, 5, 41332–41340. [Google Scholar] [CrossRef]
- Oldrini, D.; Fiebig, T.; Romano, M.R.; Proietti, D.; Berger, M.; Tontini, M.; De Ricco, R.; Santini, L.; Morelli, L.; Lay, L.; et al. Combined chemical synthesis and tailored enzymatic elongation provide fully synthetic and conjugation-ready neisseria meningitidis serogroup x vaccine antigens. ACS Chem. Biol. 2018, 13, 984–994. [Google Scholar] [CrossRef] [PubMed]
- Morelli, L.; Cancogni, D.; Tontini, M.; Nilo, A.; Filippini, S.; Costantino, P.; Romano, M.R.; Berti, F.; Adamo, R.; Lay, L. Synthesis and immunological evaluation of protein conjugates of neisseria meningitidis x capsular polysaccharide fragments. Beilstein J. Org. Chem. 2014, 10, 2367–2376. [Google Scholar] [CrossRef] [PubMed]
- Tuberculosis. Available online: http://www.who.int/en/news-room/fact-sheets/detail/tuberculosis (accessed on 28 Febraury 2018).
- Lowary, T.L. Twenty years of mycobacterial glycans: Furanosides and beyond. Acc. Chem. Res. 2016, 49, 1379–1388. [Google Scholar] [CrossRef] [PubMed]
- Veerapen, N.; Yuan, Y.; Sanders, D.A.; Pinto, B.M. Synthesis of novel ammonium and selenonium ions and their evaluation as inhibitors of udp-galactopyranose mutase. Carbohydr. Res. 2004, 339, 2205–2217. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Huang, Z.; Liu, H.-W. Synthesis and analysis of substrate analogues for udp-galactopyranose mutase: Implication for an oxocarbenium ion intermediate in the catalytic mechanism. Org. Lett. 2007, 9, 879–882. [Google Scholar] [CrossRef] [PubMed]
- Kuppala, R.; Borrelli, S.; Slowski, K.; Sanders, D.A.; Kartha, K.R.; Pinto, B.M. Synthesis and biological evaluation of nonionic substrate mimics of UDP-Galp as candidate inhibitors of udp galactopyranose mutase (UGM). Bioorg. Med. Chem. 2015, 25, 1995–1997. [Google Scholar] [CrossRef] [PubMed]
- Soltero-Higgin, M.; Carlson, E.E.; Phillips, J.H.; Kiessling, L.L. Identification of inhibitors for udp-galactopyranose mutase. J. Am. Chem. Soc. 2004, 126, 10532–10533. [Google Scholar] [CrossRef] [PubMed]
- Castagnolo, D.; De Logu, A.; Radi, M.; Bechi, B.; Manetti, F.; Magnani, M.; Supino, S.; Meleddu, R.; Chisu, L.; Botta, M. Synthesis, biological evaluation and sar study of novel pyrazole analogues as inhibitors of mycobacterium tuberculosis. Bioorg. Med. Chem. 2008, 16, 8587–8591. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Colombo, C.; Kuttiyatveetil, J.R.; Zalatar, N.; van Straaten, K.E.; Mohan, S.; Sanders, D.A.; Pinto, B.M. A second, druggable binding site in udp-galactopyranose mutase from mycobacterium tuberculosis? ChemBioChem 2016, 17, 2264–2273. [Google Scholar] [CrossRef] [PubMed]
- Dykhuizen, E.C.; May, J.F.; Tongpenyai, A.; Kiessling, L.L. Inhibitors of udp-galactopyranose mutase thwart mycobacterial growth. J. Am. Chem. Soc. 2008, 130, 6706–6707. [Google Scholar] [CrossRef] [PubMed]
- Gilleron, M.; Ronet, C.; Mempel, M.; Monsarrat, B.; Gachelin, G.; Puzo, G. Acylation state of the phosphatidylinositol mannosides from mycobacterium bovis bacillus calmette guérin and ability to induce granuloma and recruit natural killer t cells. J. Biol. Chem. 2001, 276, 34896–34904. [Google Scholar] [CrossRef] [PubMed]
- Torrelles, J.B.; Azad, A.K.; Schlesinger, L.S. Fine discrimination in the recognition of individual species of phosphatidyl-myo-inositol mannosides from mycobacterium tuberculosis by c-type lectin pattern recognition receptors. J. Immunol. 2006, 177, 1805–1816. [Google Scholar] [CrossRef] [PubMed]
- Parlane, N.A.; Compton, B.J.; Hayman, C.M.; Painter, G.F.; Basaraba, R.J.; Heiser, A.; Buddle, B.M. Phosphatidylinositol di-mannoside and derivates modulate the immune response to and efficacy of a tuberculosis protein vaccine against mycobacterium bovis infection. Vaccine 2012, 30, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Stocker, B.L.; Seeberger, P.H. Total synthesis of phosphatidylinositol mannosides of mycobacterium tuberculosis. J. Am. Chem. Soc. 2006, 128, 3638–3648. [Google Scholar] [CrossRef] [PubMed]
- Boonyarattanakalin, S.; Liu, X.; Michieletti, M.; Lepenies, B.; Seeberger, P.H. Chemical synthesis of all phosphatidylinositol mannoside (PIM) glycans from mycobacterium tuberculosis. J. Am. Chem. Soc. 2008, 130, 16791–16799. [Google Scholar] [CrossRef] [PubMed]
- Fraser-Reid, B.; Lu, J.; Jayaprakash, K.; Lopez, J.C. Synthesis of a 28-mer oligosaccharide core of mycobacterial lipoarabinomannan (LAM) requires only two n-pentenyl orthoester progenitors. Tetrahedron Asymmetry 2006, 17, 2449–2463. [Google Scholar] [CrossRef]
- Ali, A.; Wenk, M.R.; Lear, M.J. Total synthesis of a fully lipidated form of phosphatidyl-myo-inositol dimannoside (PIM-2) of mycobacterium tuberculosis. Tetrahedron Lett. 2009, 50, 5664–5666. [Google Scholar] [CrossRef]
- Patil, P.S.; Hung, S.-C. Synthesis of mycobacterial triacylated phosphatidylinositol dimannoside containing an acyl lipid chain at 3-o of inositol. Org. Lett. 2010, 12, 2618–2621. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; White, J.M.; Williams, S.J. Synthesis of glycoconjugate fragments of mycobacterial phosphatidylinositol mannosides and lipomannan. Beilstein J. Org. Chem. 2011, 7, 369. [Google Scholar] [CrossRef] [PubMed]
- Front, S.; Bourigault, M.-L.; Rose, S.; Noria, S.; Quesniaux, V.F.; Martin, O.R. Synthesis and biological investigation of pim mimics carrying biotin or a fluorescent label for cellular imaging. Bioconjug. Chem. 2012, 24, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.S.; Cheng, T.J.; Zulueta, M.M.; Yang, S.T.; Lico, L.S.; Hung, S.C. Total synthesis of tetraacylated phosphatidylinositol hexamannoside and evaluation of its immunomodulatory activity. Nat. Commun. 2015, 6, 7239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Xiong, D.-C.; Chen, S.-C.; Wang, Y.-S.; Ye, X.-S. Total synthesis of mycobacterial arabinogalactan containing 92 monosaccharide units. Nat. Commun. 2017, 8, 14851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahloul, K.; Lowary, T.L. Development of an orthogonal protection strategy for the synthesis of mycobacterial arabinomannan fragments. J. Org. Chem. 2015, 80, 11417–11434. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, S.; Wang, S.; Li, H.; Guo, Z.; Gu, G. Synthesis and immunological comparison of differently linked lipoarabinomannan oligosaccharide–monophosphoryl lipid a conjugates as antituberculosis vaccines. J. Org. Chem. 2017, 82, 12085–12096. [Google Scholar] [CrossRef] [PubMed]
- Parente-Rocha, J.A.; Bailão, A.M.; Amaral, A.C.; Taborda, C.P.; Paccez, J.D.; Borges, C.L. Antifungal resistance, metabolic routes as drug targets, and new antifungal agents: An overview about endemic dimorphic fungi. Mediat. Inflamm. 2017, 2017, 16. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Brown, G.D.; Kullberg, B.J.; Gow, N.A.R. An integrated model of the recognition of candida albicans by the innate immune system. Nat. Rev. Microbiol. 2008, 6, 67. [Google Scholar] [CrossRef] [PubMed]
- Cambi, A.; Gijzen, K.; de Vries, I.J.; Torensma, R.; Joosten, B.; Adema, G.J.; Netea, M.G.; Kullberg, B.J.; Romani, L.; Figdor, C.G. The c-type lectin dc-sign (CD209) is an antigen-uptake receptor for candida albicans on dendritic cells. Eur. J. Immunol. 2003, 33, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Gow, N.A.R.; Munro, C.A.; Bates, S.; Collins, C.; Ferwerda, G.; Hobson, R.P.; Bertram, G.; Hughes, H.B.; Jansen, T.; et al. Immune sensing of candida albicans requires cooperative recognition of mannans and glucans by lectin and toll-like receptors. J. Clin. Investig. 2006, 116, 1642–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, G.D.; Gordon, S. Immune recognition: A new receptor for β-glucans. Nature 2001, 413, 36. [Google Scholar] [CrossRef] [PubMed]
- Krylov, V.B.; Paulovičová, L.; Paulovičová, E.; Tsvetkov, Y.E.; Nifantiev, N.E. Recent advances in the synthesis of fungal antigenic oligosaccharides. Pure Appl. Chem. 2017, 89. [Google Scholar] [CrossRef]
- Richards, M.R.; Lowary, T.L. Chemistry and biology of galactofuranose-containing polysaccharides. ChemBioChem 2009, 10, 1920–1938. [Google Scholar] [CrossRef] [PubMed]
- Krylov, V.B.; Argunov, D.A.; Solovev, A.S.; Petruk, M.I.; Gerbst, A.G.; Dmitrenok, A.S.; Shashkov, A.S.; Latge, J.P.; Nifantiev, N.E. Synthesis of oligosaccharides related to galactomannans from aspergillus fumigatus and their nmr spectral data. Org. Biomol. Chem. 2018, 16, 1188–1199. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Rex, J.H.; Lee, J.; Hamill, R.J.; Larsen, R.A.; Powderly, W.; Kauffman, C.A.; Hyslop, N.; Mangino, J.E.; Chapman, S.; et al. A prospective observational study of candidemia: Epidemiology, therapy, and influences on mortality in hospitalized adult and pediatric patients. Clin. Infect. Dis. 2003, 37, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Bundle, D.R. The evolution of a glycoconjugate vaccine for candida albicans. In Carbohydrates as Drugs; Seeberger, P.H., Rademacher, C., Eds.; Springer International Publishing: Cham, Switzerland, 2014; pp. 187–234. [Google Scholar]
- Johnson, M.A.; Cartmell, J.; Weisser, N.E.; Woods, R.J.; Bundle, D.R. Molecular recognition of candida albicans (1→2)-β-mannan oligosaccharides by a protective monoclonal antibody reveals the immunodominance of internal saccharide residues. J. Biol. Chem. 2012, 287, 18078–18090. [Google Scholar] [CrossRef] [PubMed]
- Nycholat, C.M.; Bundle, D.R. Synthesis of monodeoxy and mono-O-methyl congeners of methyl β-D-mannopyranosyl-(1→2)-β-D-mannopyranoside for epitope mapping of anti-candida albicans antibodies. Carbohydr. Res. 2009, 344, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Nitz, M.; Ling, C.-C.; Otter, A.; Cutler, J.E.; Bundle, D.R. The unique solution structure and immunochemistry of thecandida albicans β-1,2-mannopyranan cell wall antigens. J. Biol. Chem. 2002, 277, 3440–3446. [Google Scholar] [CrossRef] [PubMed]
- Nikrad, P.V.; Beierbeck, H.; Lemieux, R.U. Molecular recognition x. A novel procedure for the detection of the intermolecular hydrogen bonds present in a protein oligosaccharide complex. Can. J. Chem. 1992, 70, 241–253. [Google Scholar] [CrossRef]
- Bundle, D.R.; Nycholat, C.; Costello, C.; Rennie, R.; Lipinski, T. Design of a candida albicans disaccharide conjugate vaccine by reverse engineering a protective monoclonal antibody. ACS Chem. Biol. 2012, 7, 1754–1763. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, T.; Wu, X.; Sadowska, J.; Kreiter, E.; Yasui, Y.; Cheriaparambil, S.; Rennie, R.; Bundle, D.R. A β-mannan trisaccharide conjugate vaccine aids clearance of candida albicans in immunocompromised rabbits. Vaccine 2012, 30, 6263–6269. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Dziadek, S.; Bundle, D.R.; Cutler, J.E. Synthetic glycopeptide vaccines combining beta-mannan and peptide epitopes induce protection against candidiasis. Proc. Natl. Acad. Sci. USA 2008, 105, 13526–13531. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Kawai, T.; Adachi, Y.; Ohno, N.; Takahashi, T. [small beta](1,3) branched heptadeca- and linear hexadeca-saccharides possessing an aminoalkyl group as a strong ligand to dectin-1. Chem. Commun. 2010, 46, 8249–8251. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Kawai, T.; Adachi, Y.; Hanashima, S.; Yamaguchi, Y.; Ohno, N.; Takahashi, T. Synthesis of beta(1,3) oligoglucans exhibiting a dectin-1 binding affinity and their biological evaluation. Bioorg. Med. Chem. 2012, 20, 3898–3914. [Google Scholar] [CrossRef] [PubMed]
- Bromuro, C.; Romano, M.; Chiani, P.; Berti, F.; Tontini, M.; Proietti, D.; Mori, E.; Torosantucci, A.; Costantino, P.; Rappuoli, R.; et al. Beta-glucan-CRM197 conjugates as candidates antifungal vaccines. Vaccine 2010, 28, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Adamo, R.; Tontini, M.; Brogioni, G.; Romano, M.R.; Costantini, G.; Danieli, E.; Proietti, D.; Berti, F.; Costantino, P. Synthesis of laminarin fragments and evaluation of a β-(1,3) glucan hexasaccaride-crm197 conjugate as vaccine candidate against candida albicans. J. Carbohydr. Chem. 2011, 30, 249–280. [Google Scholar] [CrossRef]
- Liao, G.; Zhou, Z.; Burgula, S.; Liao, J.; Yuan, C.; Wu, Q.; Guo, Z. Synthesis and immunological studies of linear oligosaccharides of beta-glucan as antigens for antifungal vaccine development. Bioconjug. Chem. 2015, 26, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Zhou, Z.; Liao, J.; Zu, L.; Wu, Q.; Guo, Z. 6-O-branched oligo-β-glucan-based antifungal glycoconjugate vaccines. ACS Infect. Dis. 2016, 2, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Komarova, B.S.; Orekhova, M.V.; Tsvetkov, Y.E.; Beau, R.; Aimanianda, V.; Latge, J.P.; Nifantiev, N.E. Synthesis of a pentasaccharide and neoglycoconjugates related to fungal alpha-(1→3)-glucan and their use in the generation of antibodies to trace aspergillus fumigatus cell wall. Chem. Eur. J. 2015, 21, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Latgé, J.P.; Kobayashi, H.; Debeaupuis, J.P.; Diaquin, M.; Sarfati, J.; Wieruszeski, J.M.; Parra, E.; Bouchara, J.P.; Fournet, B. Chemical and immunological characterization of the extracellular galactomannan of aspergillus fumigatus. Infect. Immun. 1994, 62, 5424–5433. [Google Scholar] [PubMed]
- Kudoh, A.; Okawa, Y.; Shibata, N. Significant structural change in both O- and N-linked carbohydrate moieties of the antigenic galactomannan from aspergillus fumigatus grown under different culture conditions. Glycobiology 2015, 25, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Argunov, D.A.; Krylov, V.B.; Nifantiev, N.E. Convergent synthesis of isomeric heterosaccharides related to the fragments of galactomannan from aspergillus fumigatus. Org. Biomol. Chem. 2015, 13, 3255–3267. [Google Scholar] [CrossRef] [PubMed]
- Argunov, D.A.; Krylov, V.B.; Nifantiev, N.E. The use of pyranoside-into-furanoside rearrangement and controlled O(5)→O(6) benzoyl migration as the basis of a synthetic strategy to assemble (1→5)- and (1→6)-linked galactofuranosyl chains. Org. Lett. 2016, 18, 5504–5507. [Google Scholar] [CrossRef] [PubMed]
- Matveev, A.L.; Krylov, V.B.; Emelyanova, L.A.; Solovev, A.S.; Khlusevich, Y.A.; Baykov, I.K.; Fontaine, T.; Latgé, J.-P.; Tikunova, N.V.; Nifantiev, N.E. Novel mouse monoclonal antibodies specifically recognize aspergillus fumigatus galactomannan. PLoS ONE 2018, 13, e0193938. [Google Scholar] [CrossRef] [PubMed]
- Kabat, E.A. The upper limit for the size of the human antidextran combining site. J. Immunol. 1960, 84, 82–85. [Google Scholar] [PubMed]
- Nilo, A.; Morelli, L.; Passalacqua, I.; Brogioni, B.; Allan, M.; Carboni, F.; Pezzicoli, A.; Zerbini, F.; Maione, D.; Fabbrini, M.; et al. Anti-group b streptococcus glycan-conjugate vaccines using pilus protein gbs80 as carrier and antigen: Comparing lysine and tyrosine-directed conjugation. ACS Chem. Biol. 2015, 10, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- Stefanetti, G.; Hu, Q.-Y.; Usera, A.; Robinson, Z.; Allan, M.; Singh, A.; Imase, H.; Cobb, J.; Zhai, H.; Quinn, D.; et al. Sugar-protein connectivity impacts on the immunogenicity of site-selective salmonella o-antigen glycoconjugate vaccines. Angew. Chem. Int. Ed. 2015, 54, 13198–13203. [Google Scholar] [CrossRef]
- Fernandez-Tejada, A.; Canada, F.J.; Jimenez-Barbero, J. Recent developments in synthetic carbohydrate-based diagnostics, vaccines, and therapeutics. Chem. Eur. J. 2015, 21, 10616–10628. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Representation of the monosaccharide building blocks contained in the glycans of pathogen cell wall PS accounted in this manuscript.
Figure 1.
Representation of the monosaccharide building blocks contained in the glycans of pathogen cell wall PS accounted in this manuscript.
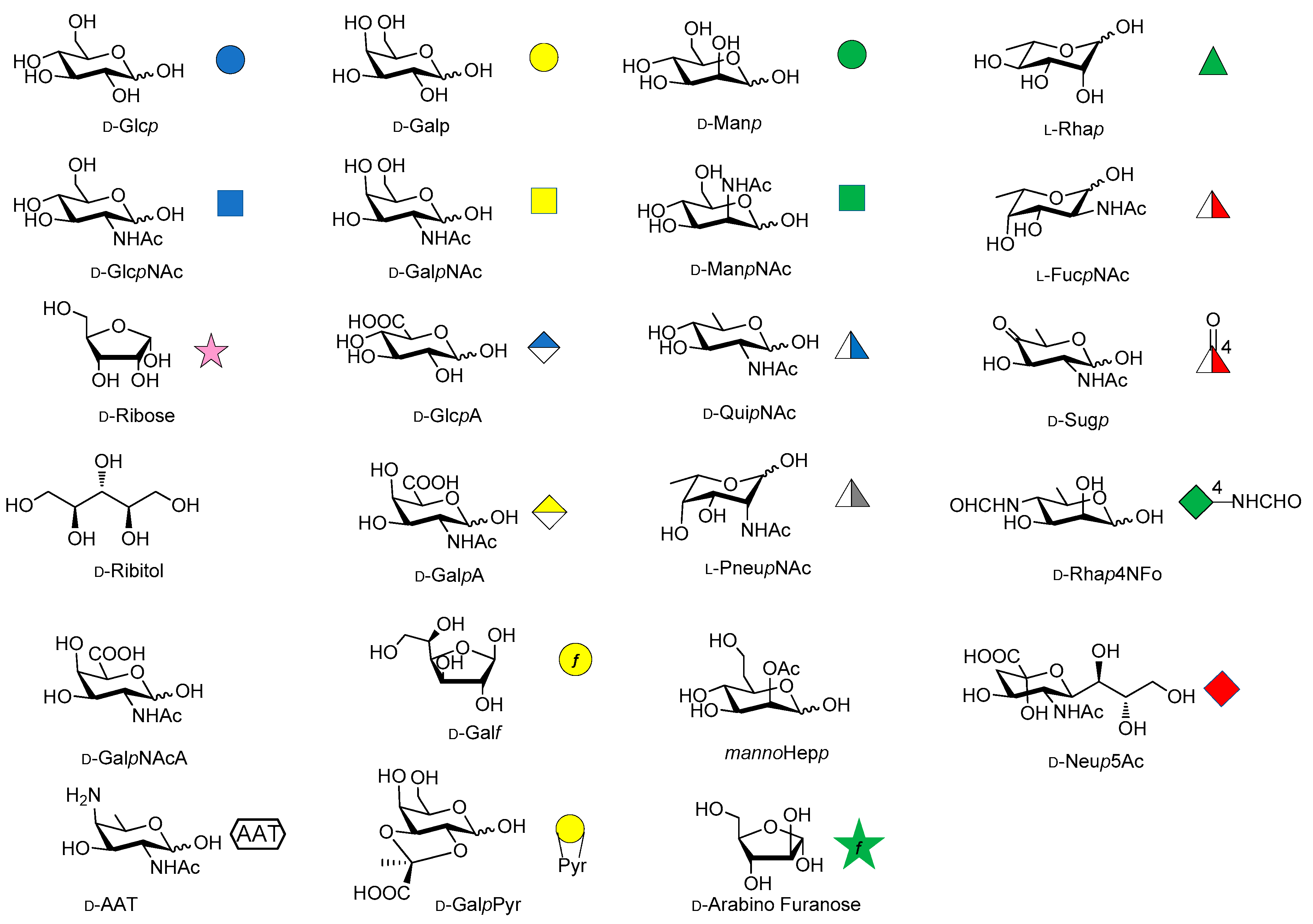
Figure 2.
Repeating unit of the O-antigen of S. flexneri 2a LPS and synthetic fragments reported in [16,17,18].
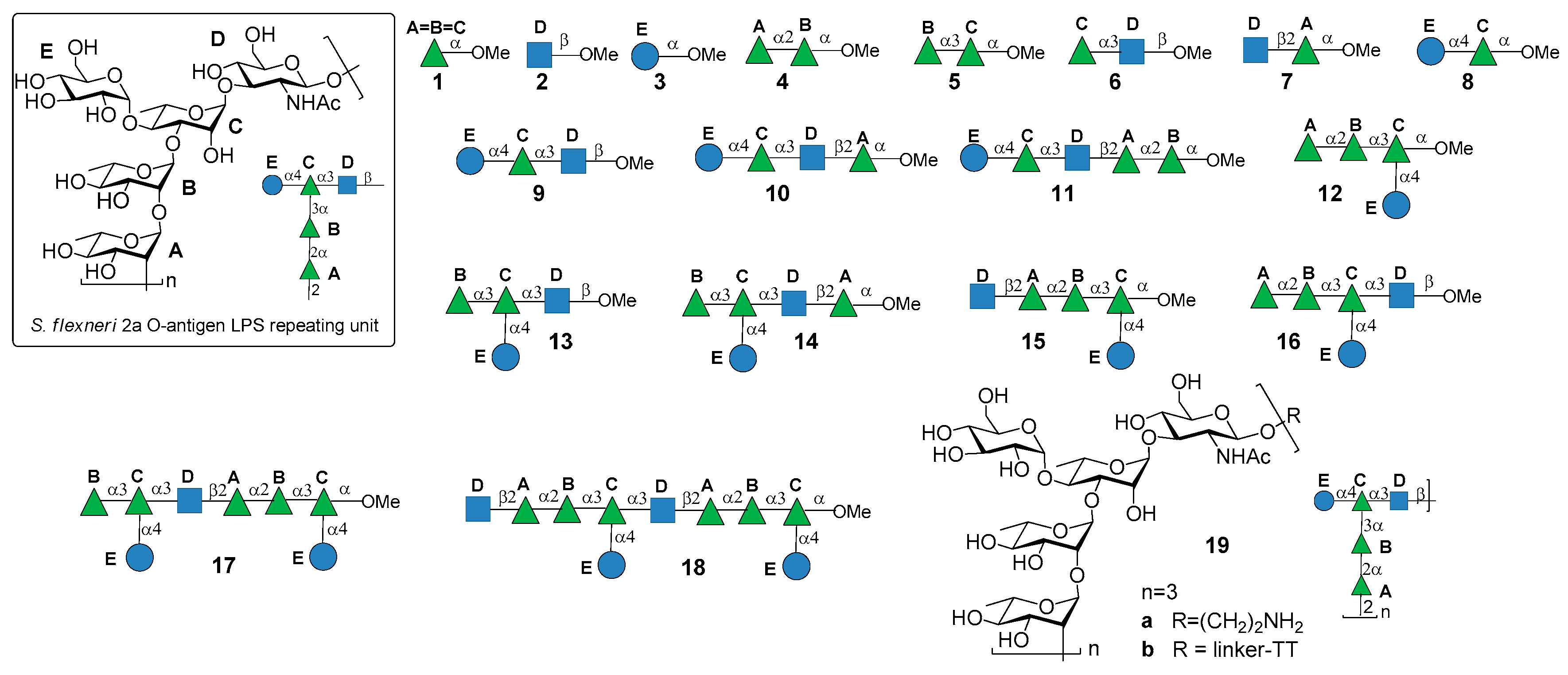
Figure 3.
C. difficile PS-I repeating unit, synthetic fragments 20–25 and retrosynthetic analysis reported in [29].
Figure 3.
C. difficile PS-I repeating unit, synthetic fragments 20–25 and retrosynthetic analysis reported in [29].
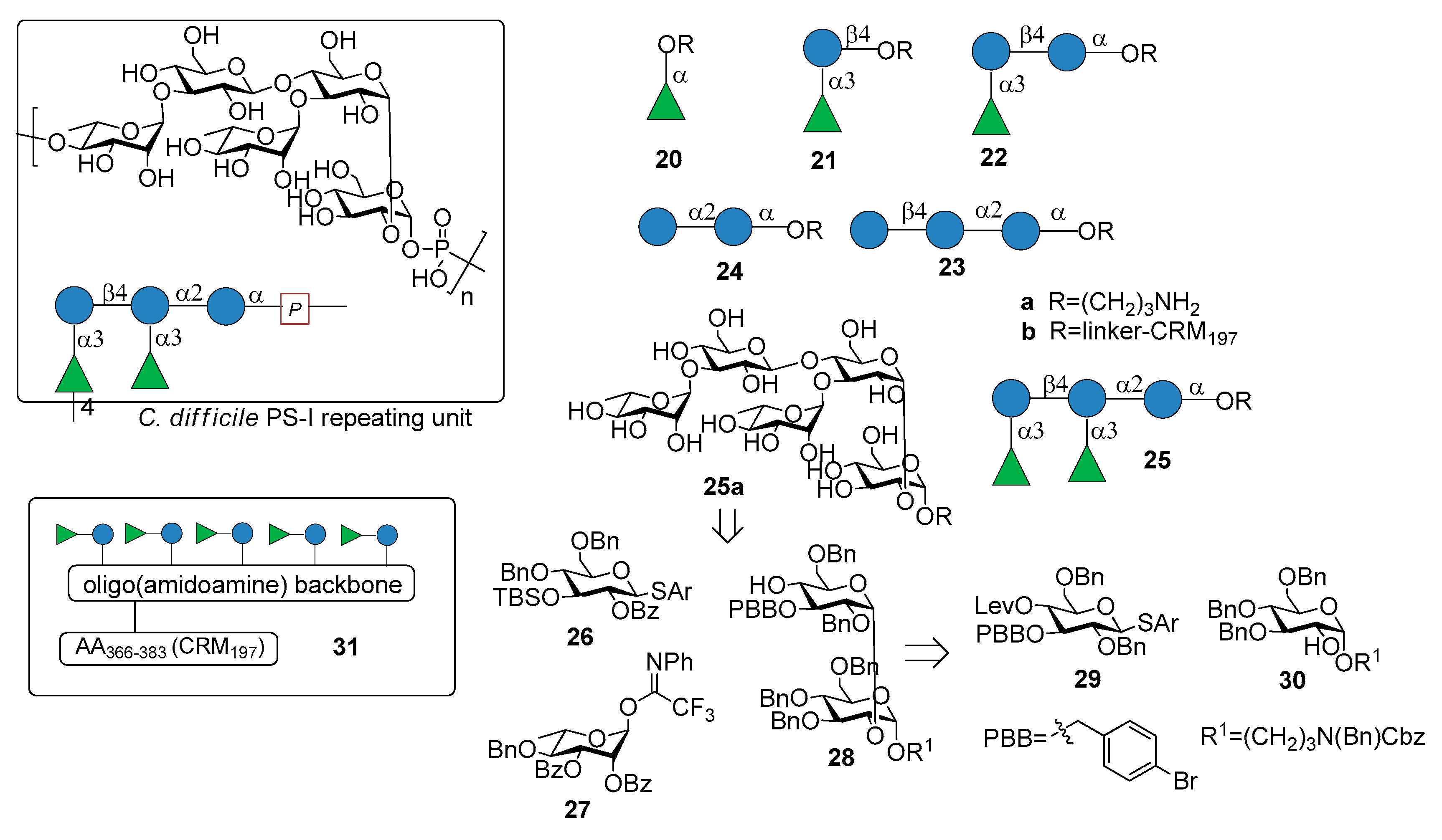
Figure 4.
C. difficile PS-II repeating unit and retrosynthetic analysis of fragment 32 reported in [32].
Figure 4.
C. difficile PS-II repeating unit and retrosynthetic analysis of fragment 32 reported in [32].

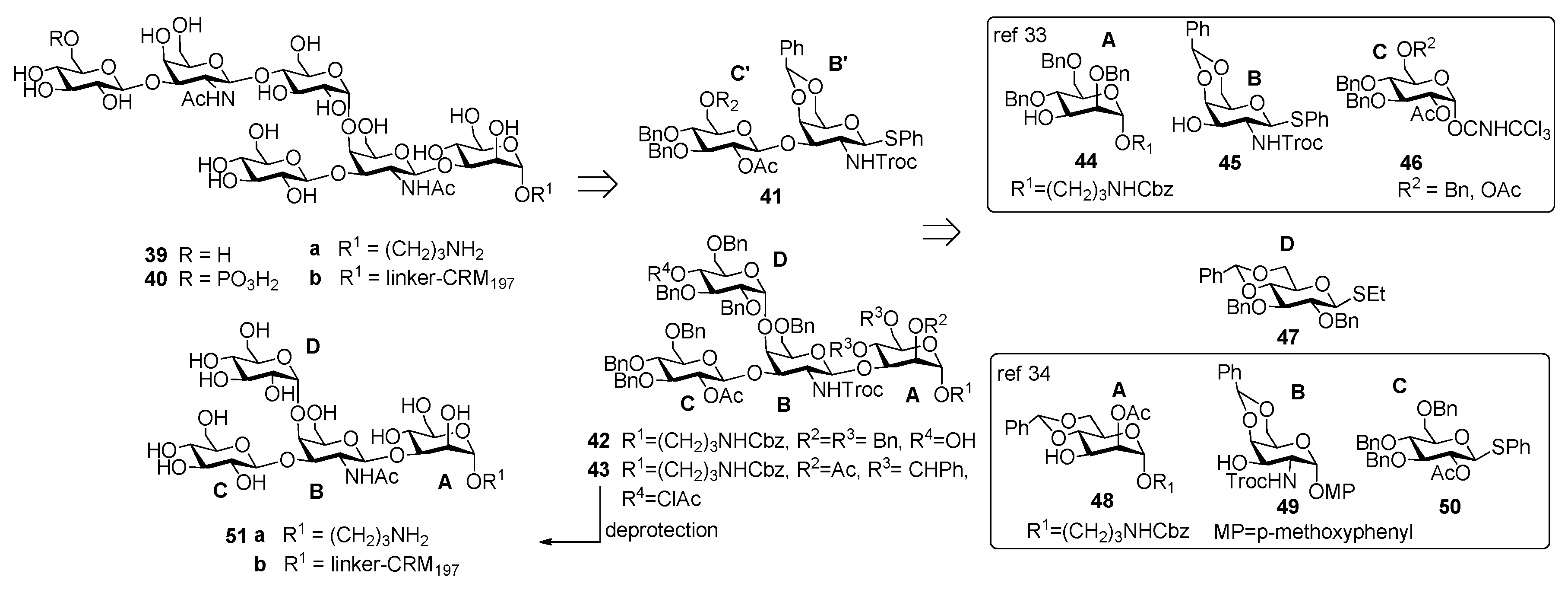
Figure 6.
B. pseudomallei CPS repeating unit and retrosynthetic analysis of compound 52 reported in [37].
Figure 6.
B. pseudomallei CPS repeating unit and retrosynthetic analysis of compound 52 reported in [37].
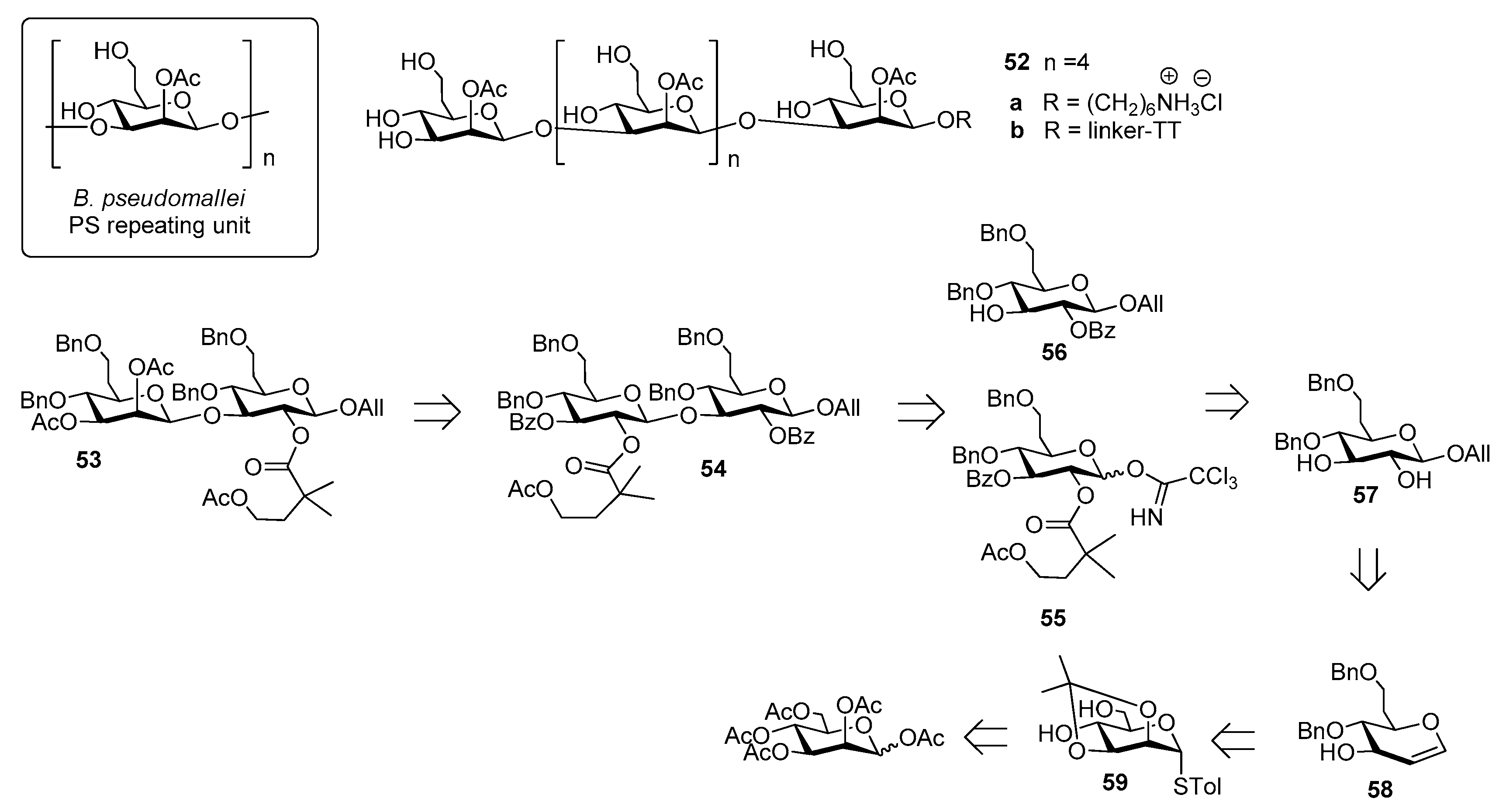
Figure 7.
A and M antigenic determinants in Brucella OPS and oligomers synthesized by Bundle et al. [49].
Figure 7.
A and M antigenic determinants in Brucella OPS and oligomers synthesized by Bundle et al. [49].
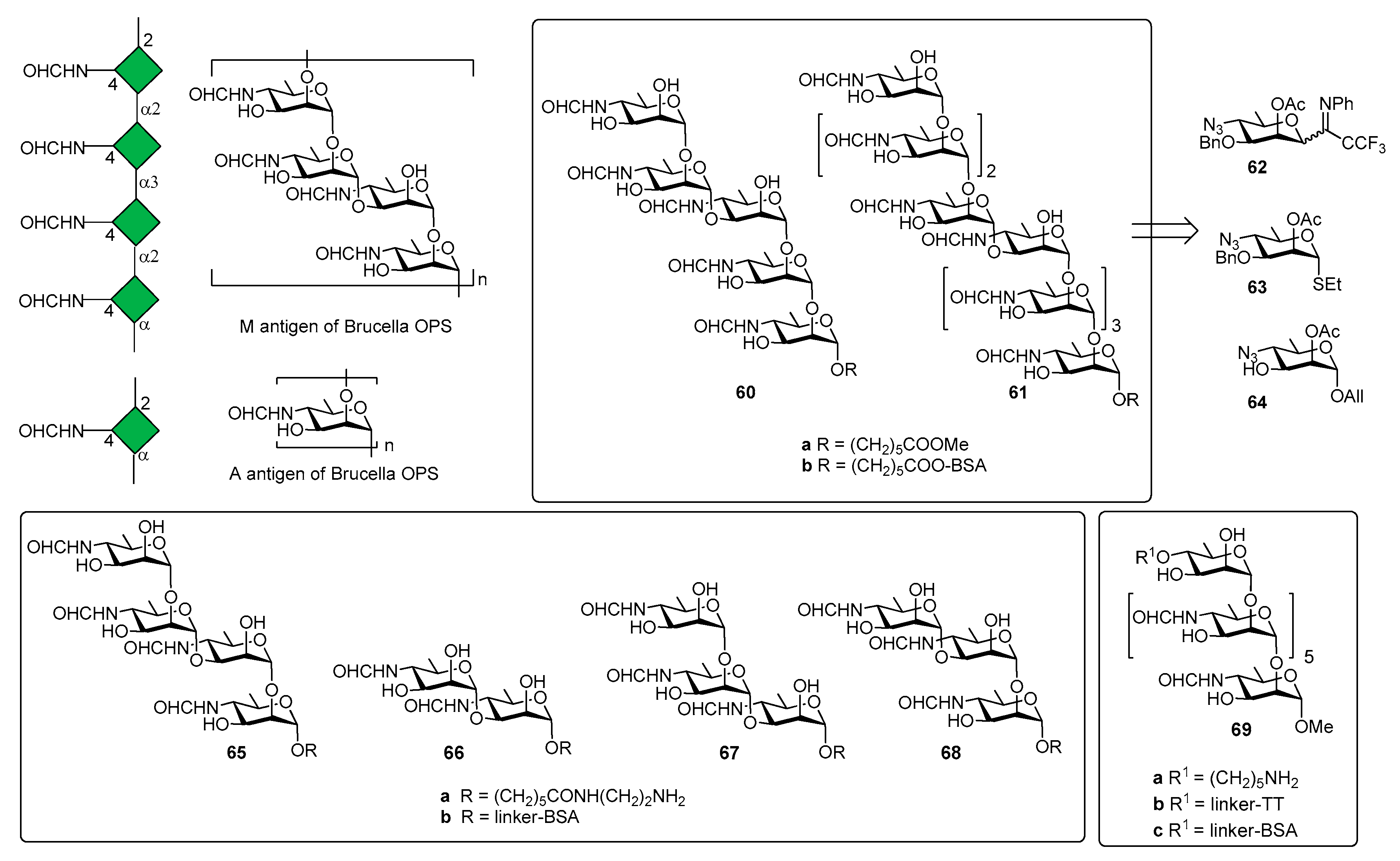
Figure 8.
Synthesis of QuimiHib.
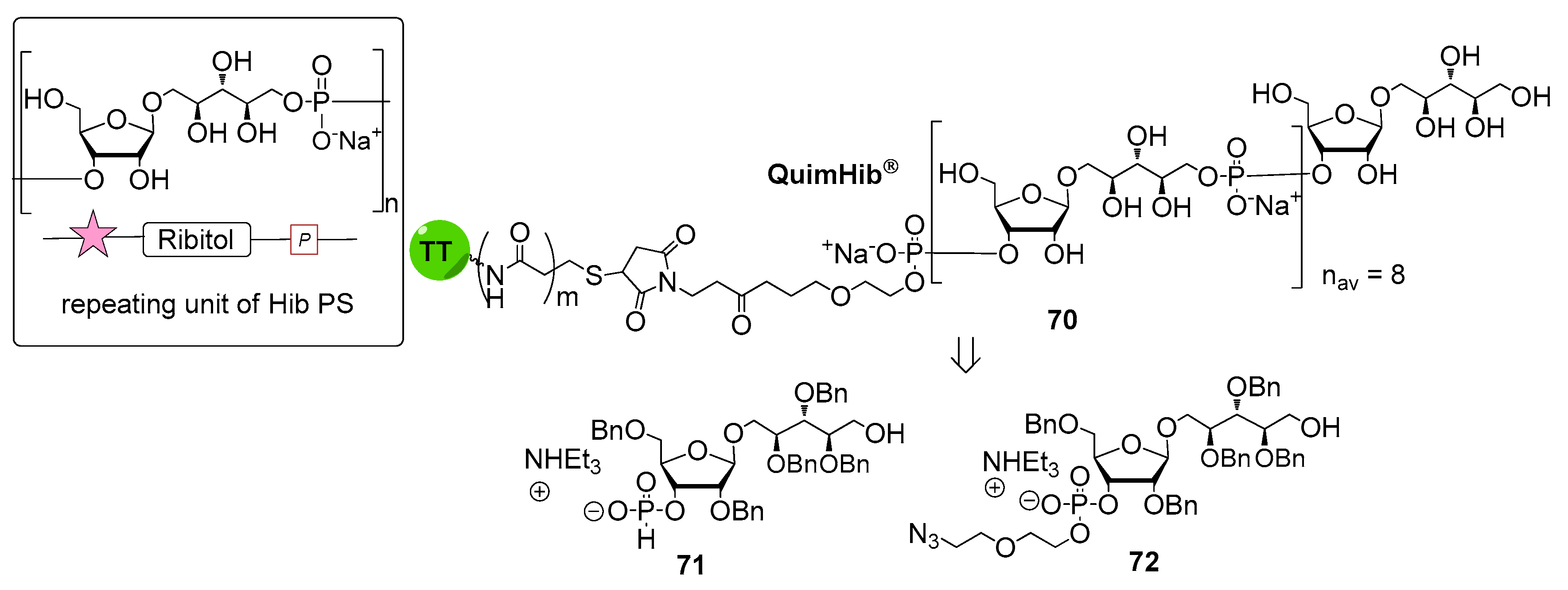
Figure 9.
Haemophilus influenzae type b PRP oligosaccharides reported in [63].
Figure 9.
Haemophilus influenzae type b PRP oligosaccharides reported in [63].
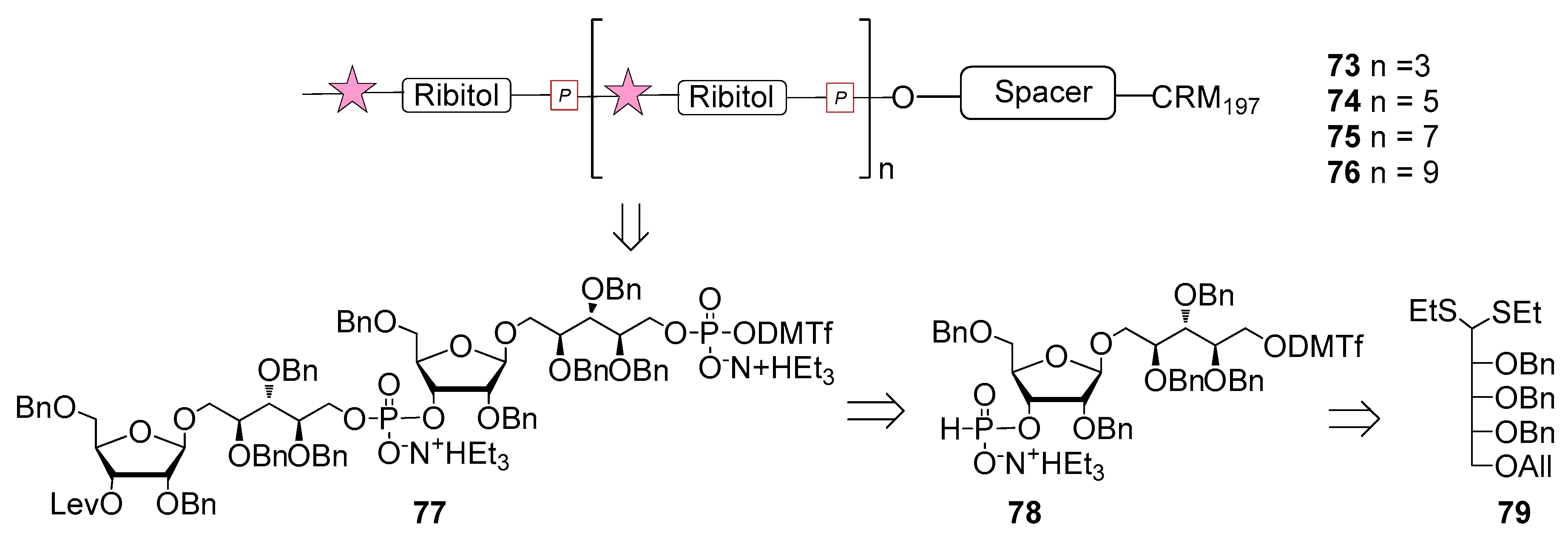
Figure 10.
Repeating units of Streptococcus pneumoniae 1 CPS and Bacteroides fragilils PS A1 CPS and synthetic glycan fragments reported in [72].
Figure 10.
Repeating units of Streptococcus pneumoniae 1 CPS and Bacteroides fragilils PS A1 CPS and synthetic glycan fragments reported in [72].
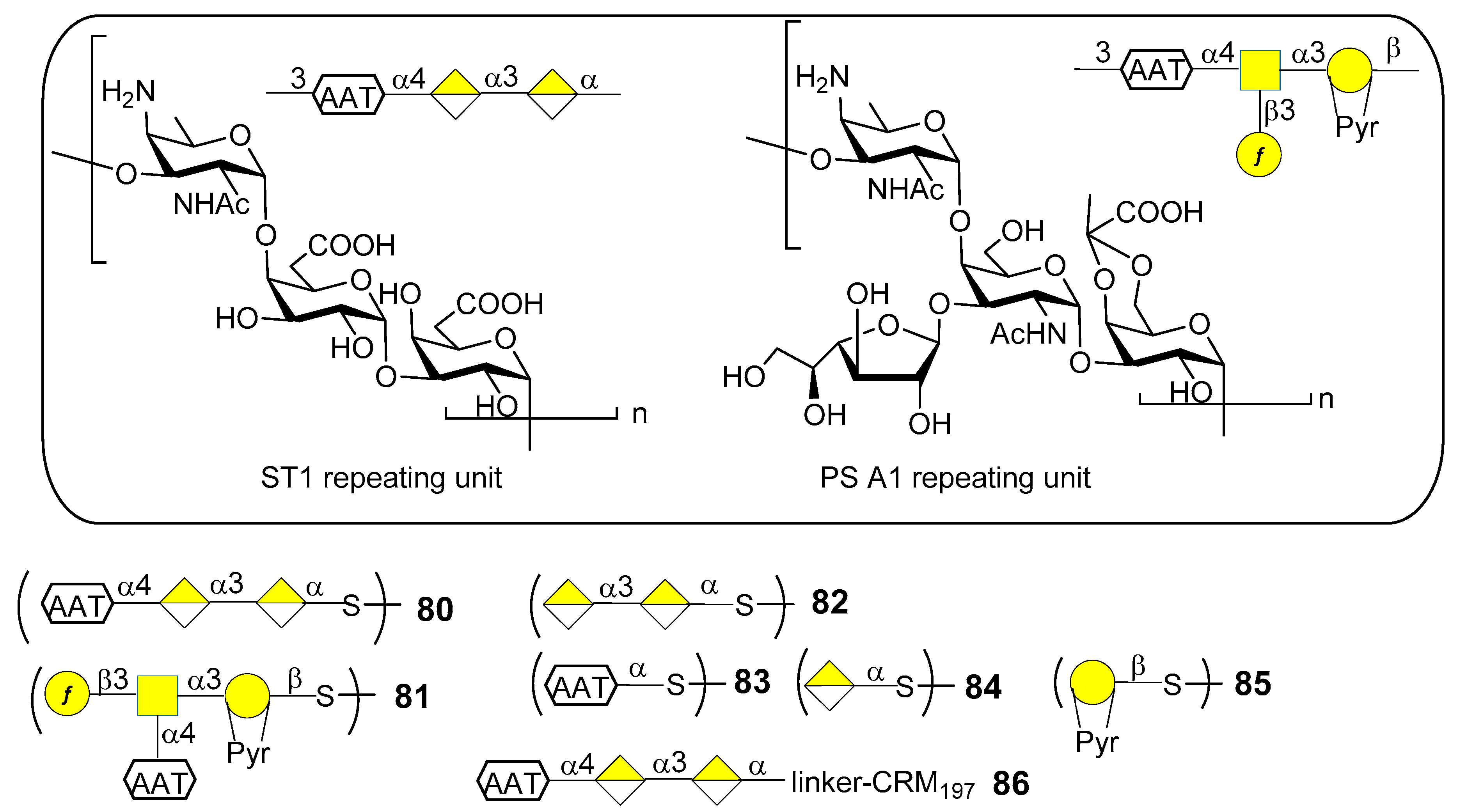
Figure 11.
Repeating unit of Streptococcus pneumoniae 2 CPS and synthetic ST2 glycan fragments reported in [76].
Figure 11.
Repeating unit of Streptococcus pneumoniae 2 CPS and synthetic ST2 glycan fragments reported in [76].
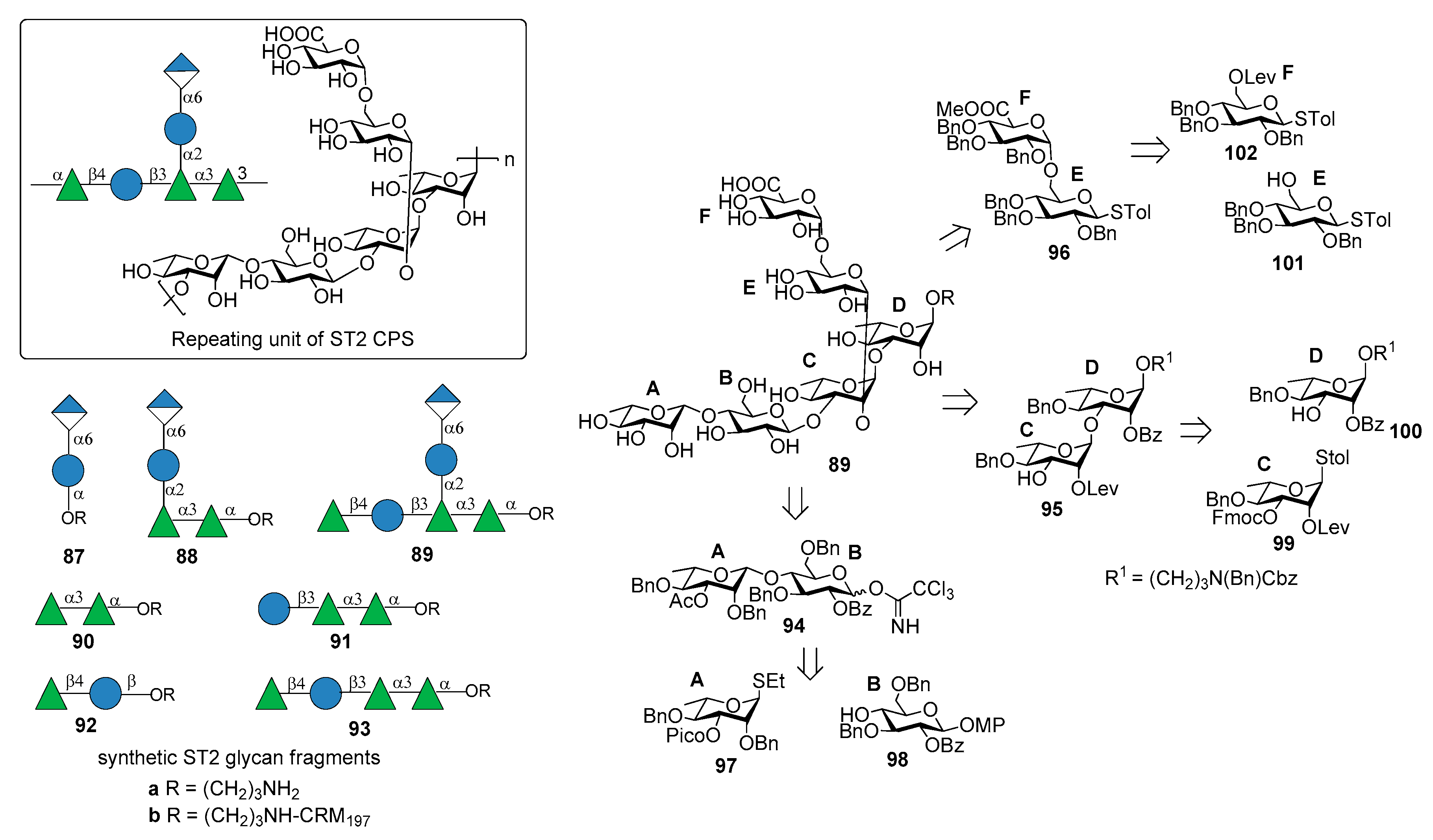
Figure 12.
Repeating unit of Streptococcus pneumoniae 3 CPS and synthetic ST3 glycan fragments reported in [81].
Figure 12.
Repeating unit of Streptococcus pneumoniae 3 CPS and synthetic ST3 glycan fragments reported in [81].
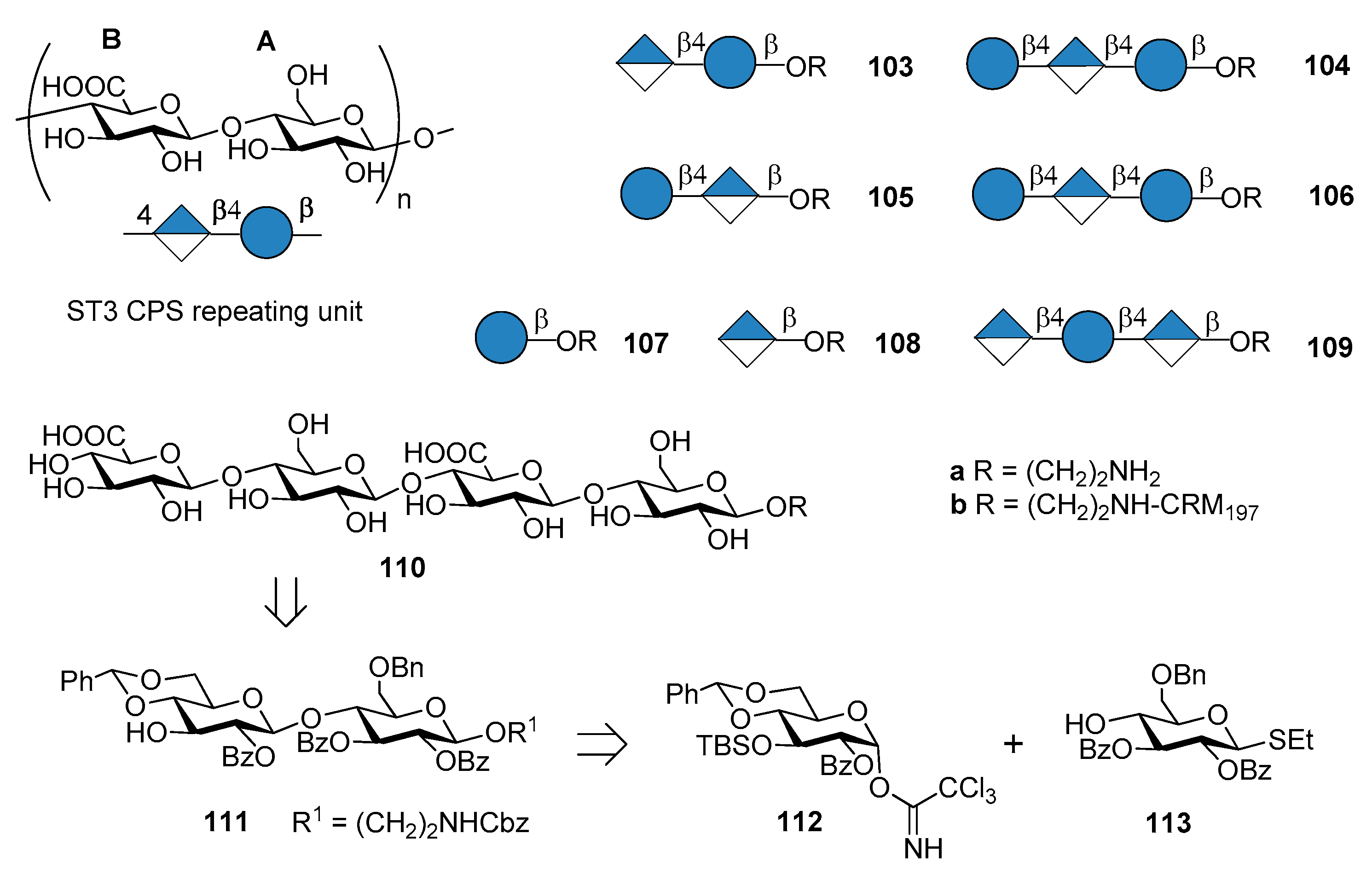
Figure 13.
Synthesis of ST3 oligosaccharides and glycoconjugates from [82].
Figure 13.
Synthesis of ST3 oligosaccharides and glycoconjugates from [82].
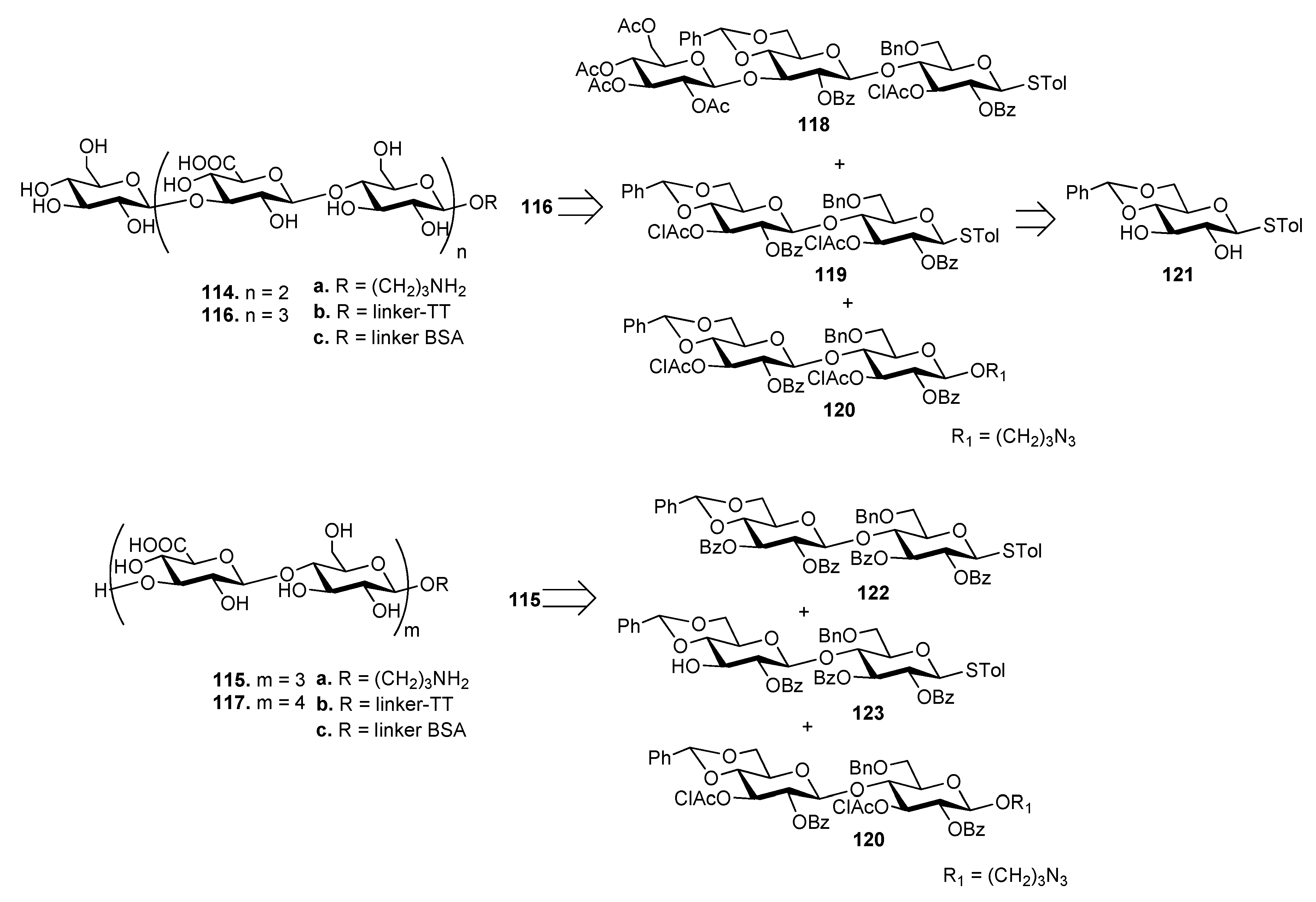
Figure 14.
Repeating unit of ST4 CPS and synthesis of ST4 CPS fragments in the pyruvalated form and the nonpyruvalated variant, as reported in [84].
Figure 14.
Repeating unit of ST4 CPS and synthesis of ST4 CPS fragments in the pyruvalated form and the nonpyruvalated variant, as reported in [84].
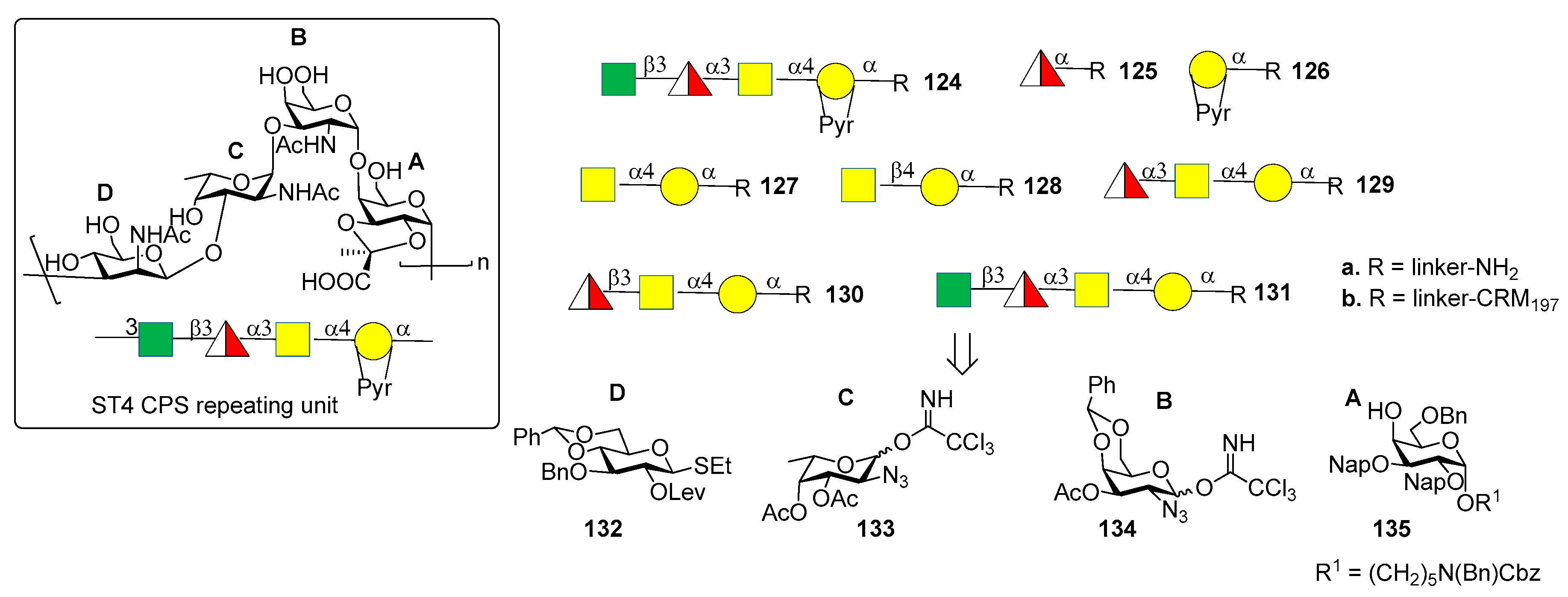
Figure 15.
Repeating unit of ST5 CPS and synthetic glycan fragments reported in [89].
Figure 15.
Repeating unit of ST5 CPS and synthetic glycan fragments reported in [89].
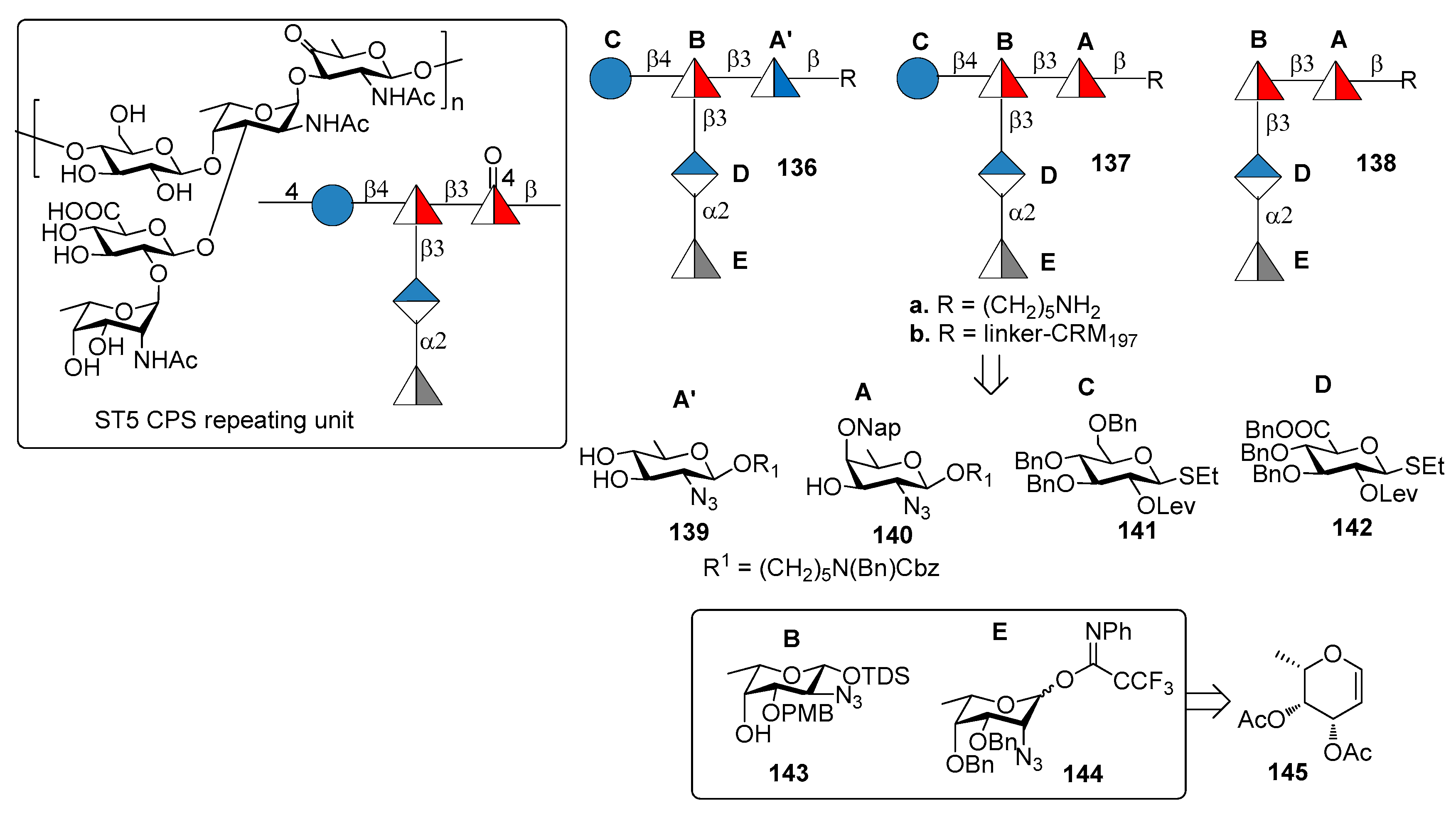
Figure 16.
Repeating unit of Streptococcus pneumoniae 8 CPS and synthetic ST8 glycan fragments reported in [92].
Figure 16.
Repeating unit of Streptococcus pneumoniae 8 CPS and synthetic ST8 glycan fragments reported in [92].

Figure 17.
Repeating unit of S. pneumoniae type 14 (ST14) and S. pneumoniae type 19F (ST19F), and their synthetic fragment conjugated to gold nanoparticles reported in Reference [96] and in Reference [98].

Figure 18.
Repeating unit of the cell wall PS of GAS and synthetic fragments reported in [104].
Figure 18.
Repeating unit of the cell wall PS of GAS and synthetic fragments reported in [104].
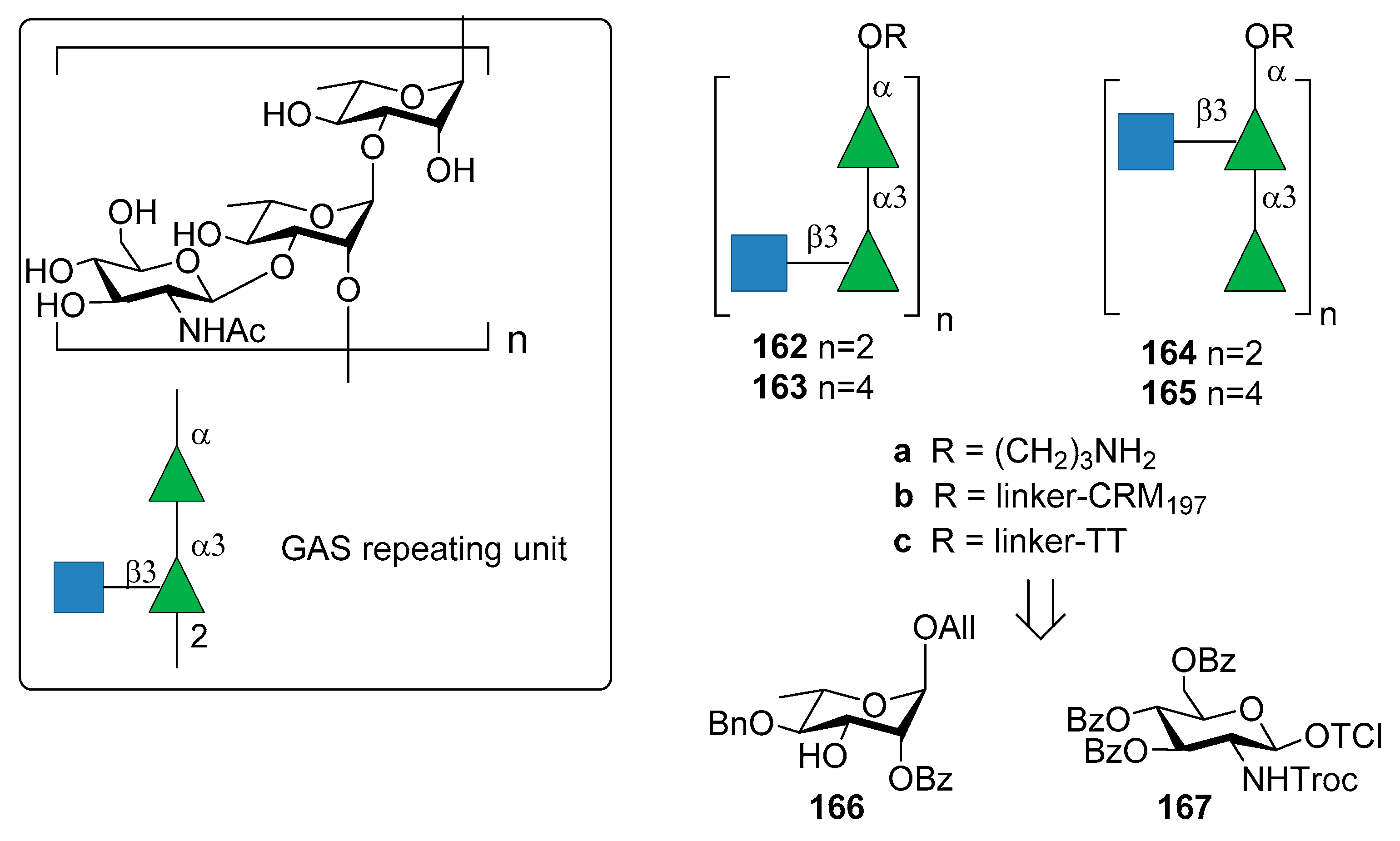
Figure 19.
GBS PSIII repeating unit.
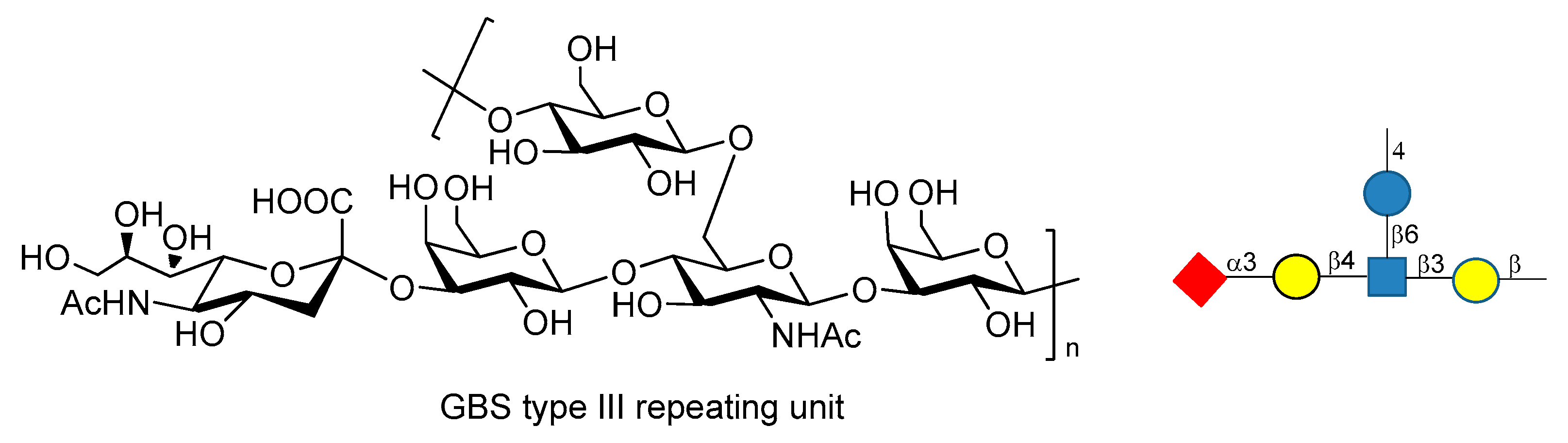
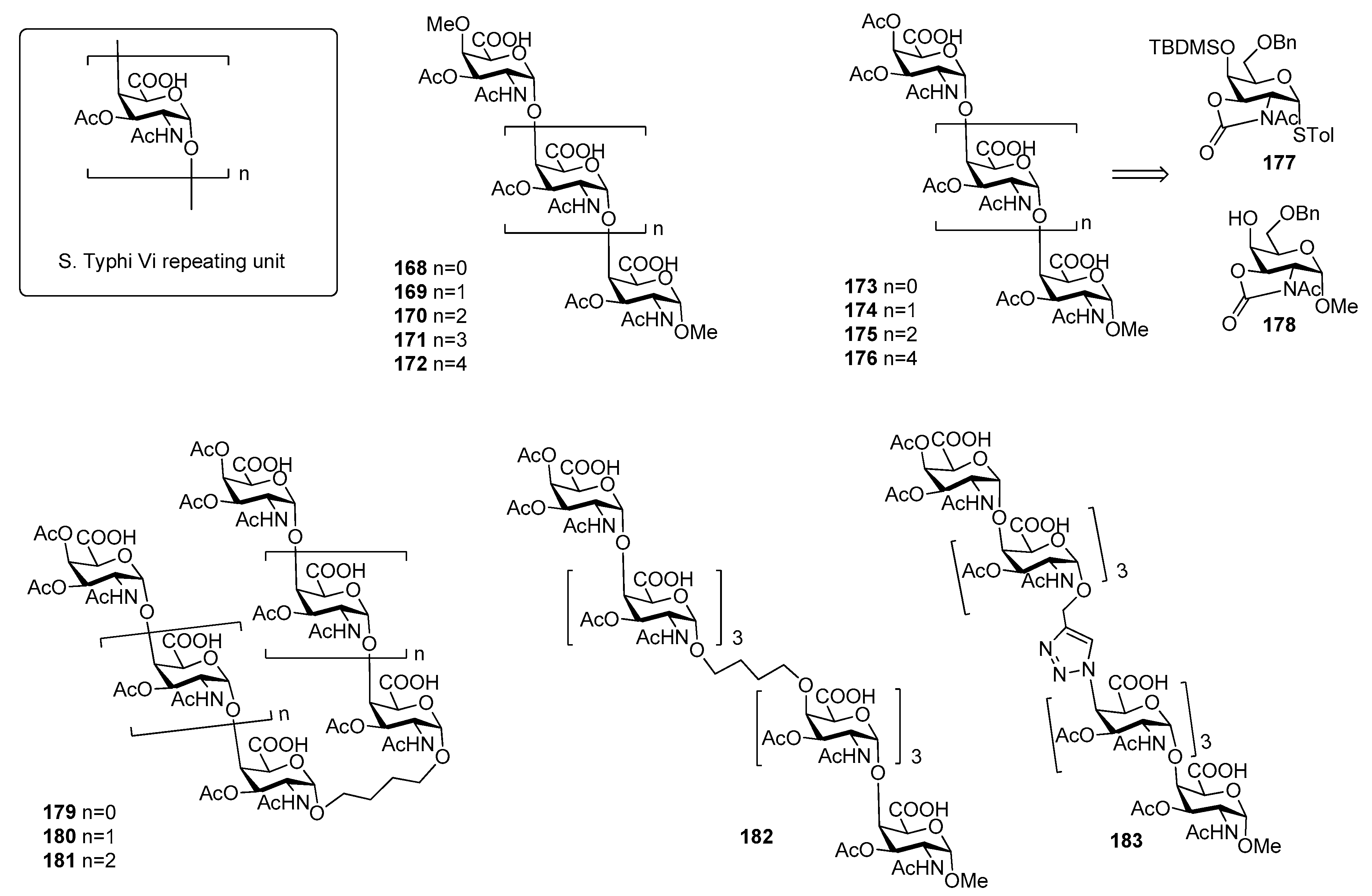
Figure 21.
Retrosynthetic analysis of di- and trisaccharide fragments of S. Typhi Vi CPS, reported in Reference [147].
Figure 21.
Retrosynthetic analysis of di- and trisaccharide fragments of S. Typhi Vi CPS, reported in Reference [147].
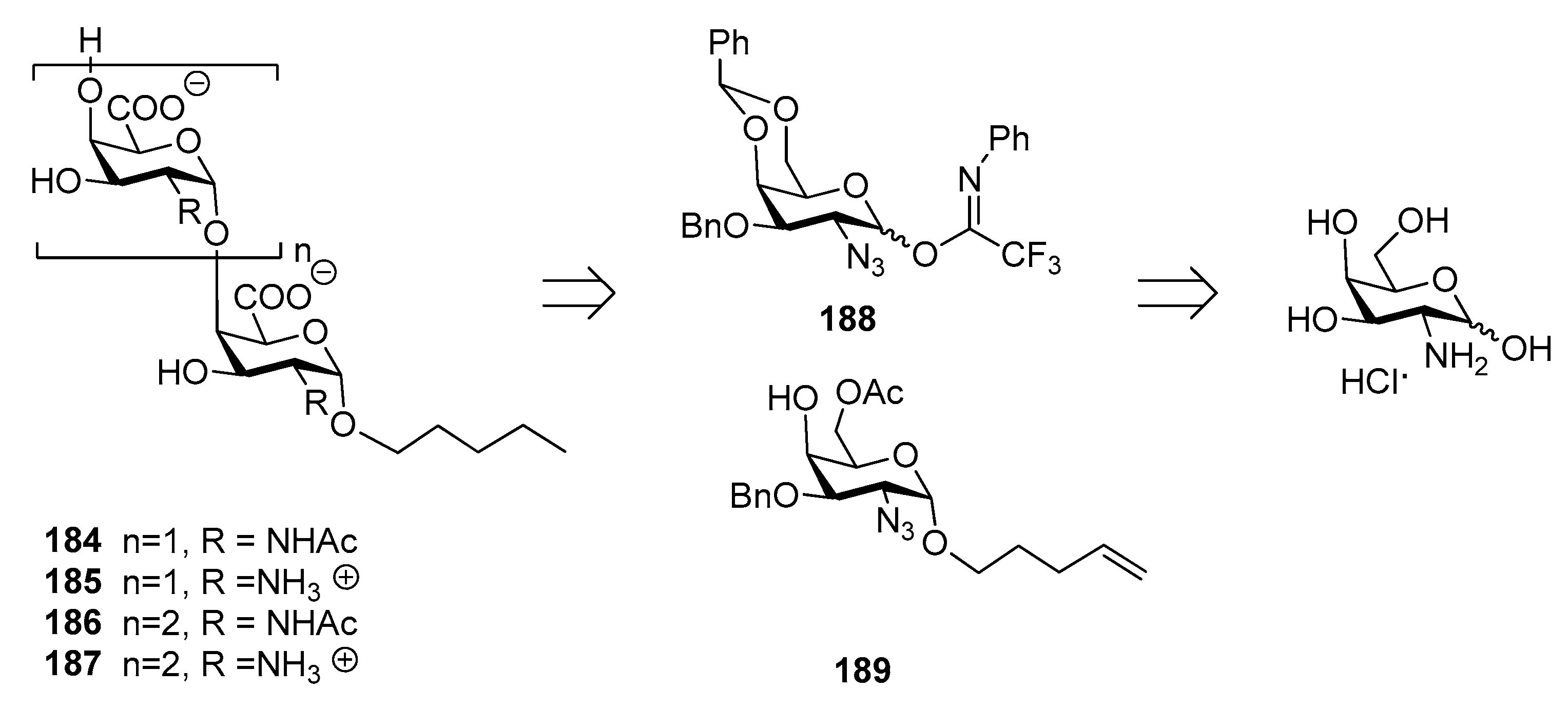
Figure 22.
Repeating unit of PsI of P. aeruginosa and synthetic fragments reported in Reference [158].
Figure 22.
Repeating unit of PsI of P. aeruginosa and synthetic fragments reported in Reference [158].
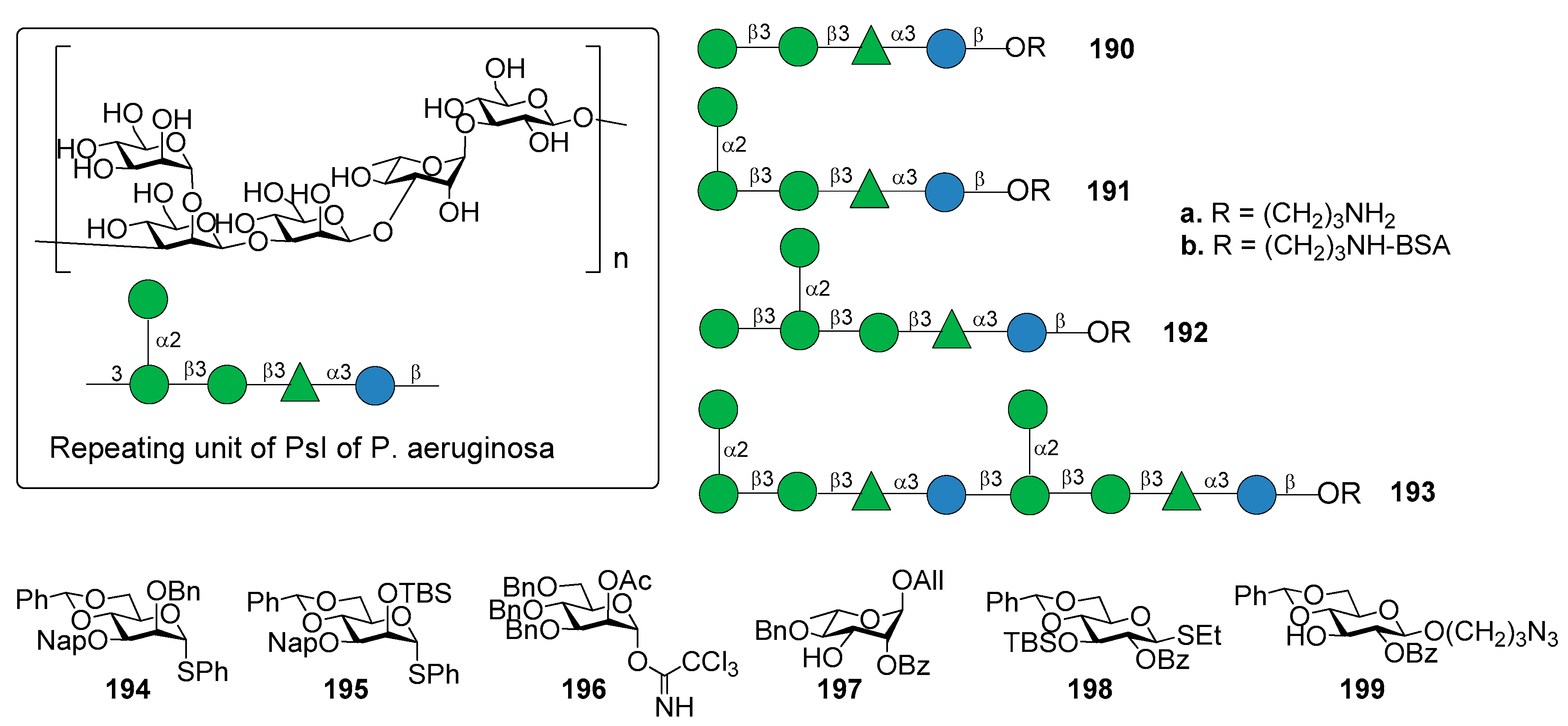
Figure 23.
Men A CPS carbocyclic analogues reported in Reference [172].
Figure 23.
Men A CPS carbocyclic analogues reported in Reference [172].

Figure 24.
Retrosynthetic strategy for the synthesis of C-phosphono analogues of MenA CPS reported in [174] and multivalent presentation of the fragments [181].
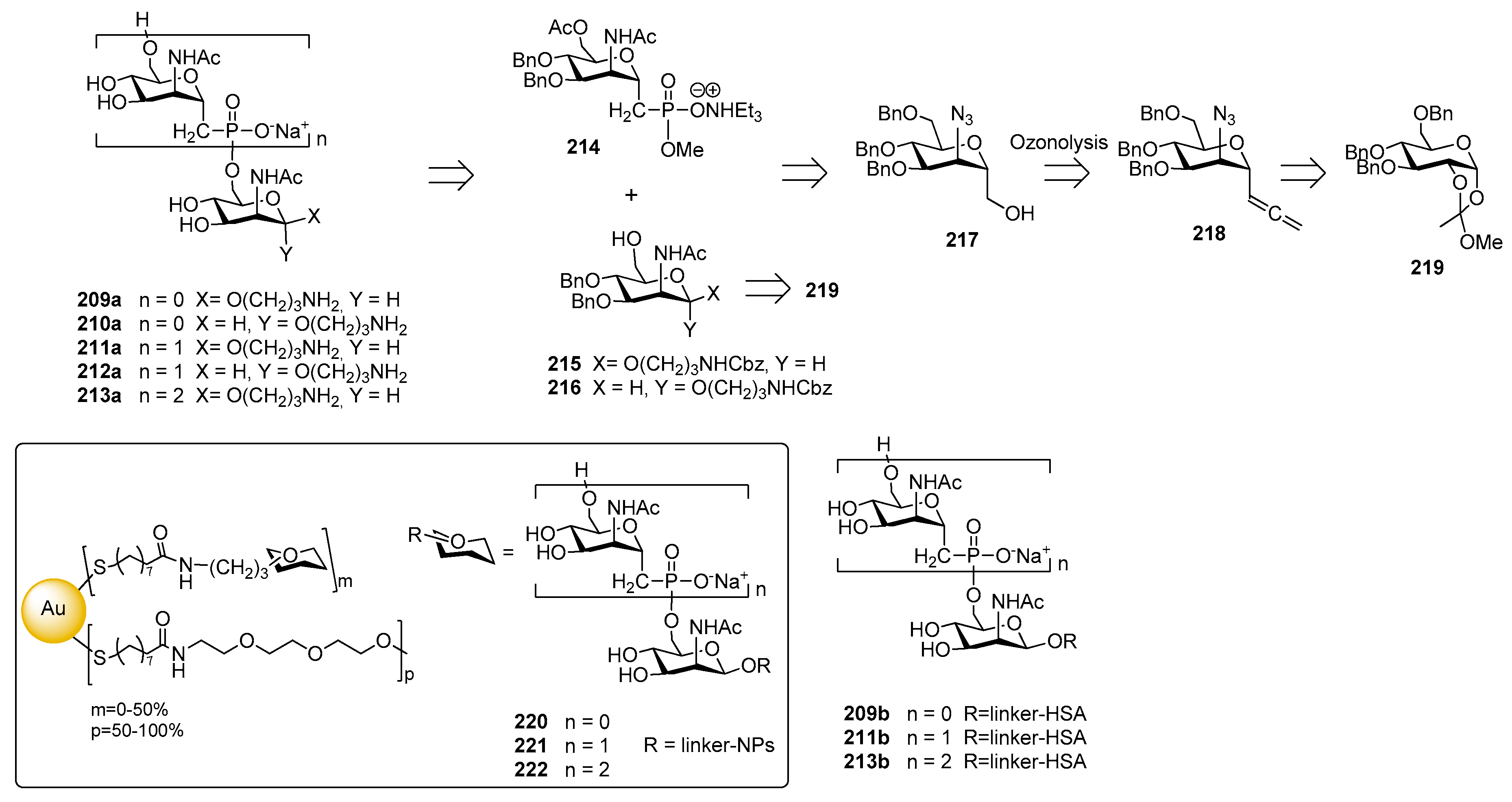
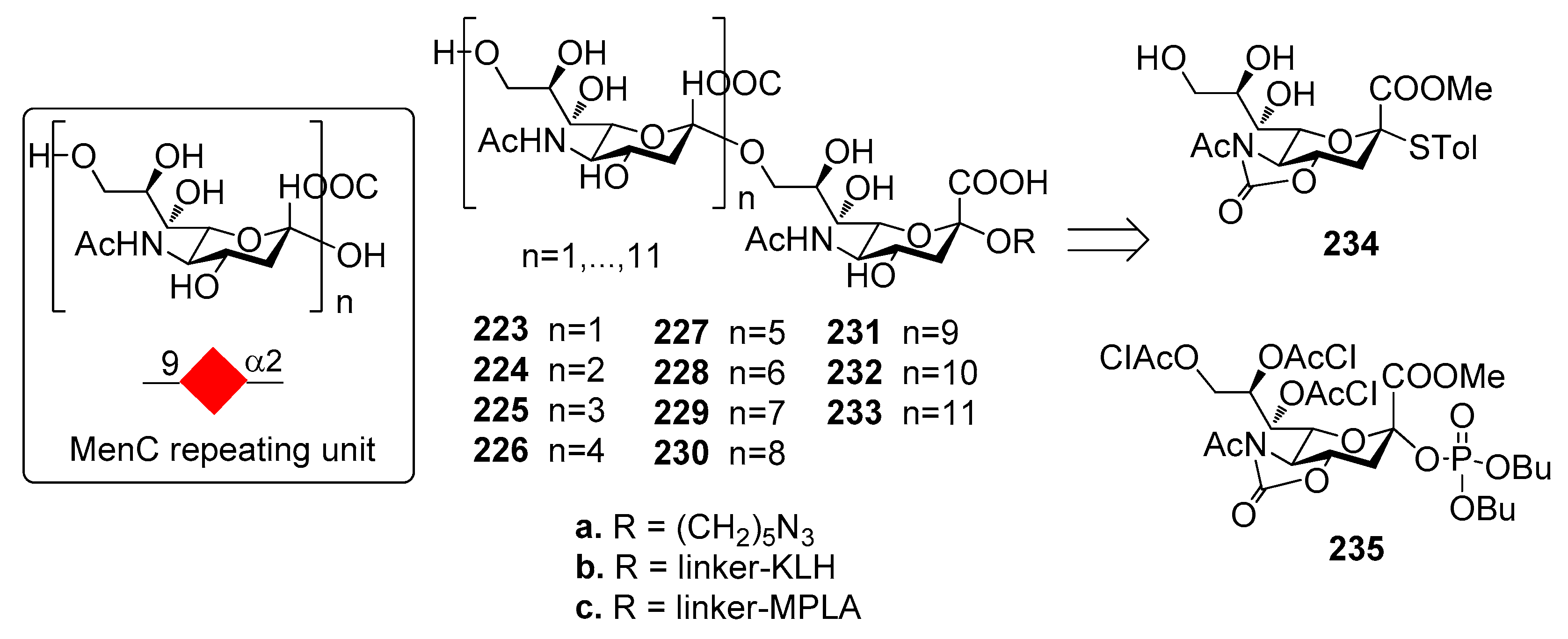
Figure 26.
Repeating unit of MenW CPS and synthetic strategy used the construction of MenW CPS fragments [194].
Figure 26.
Repeating unit of MenW CPS and synthetic strategy used the construction of MenW CPS fragments [194].
Figure 28.
Phosphatidylinositol mannosides (PIMs) and retrosynthetic route of tetraacylated phosphatidylinositol hexamannoside (Ac2PIM6) reported in [223].
Figure 28.
Phosphatidylinositol mannosides (PIMs) and retrosynthetic route of tetraacylated phosphatidylinositol hexamannoside (Ac2PIM6) reported in [223].
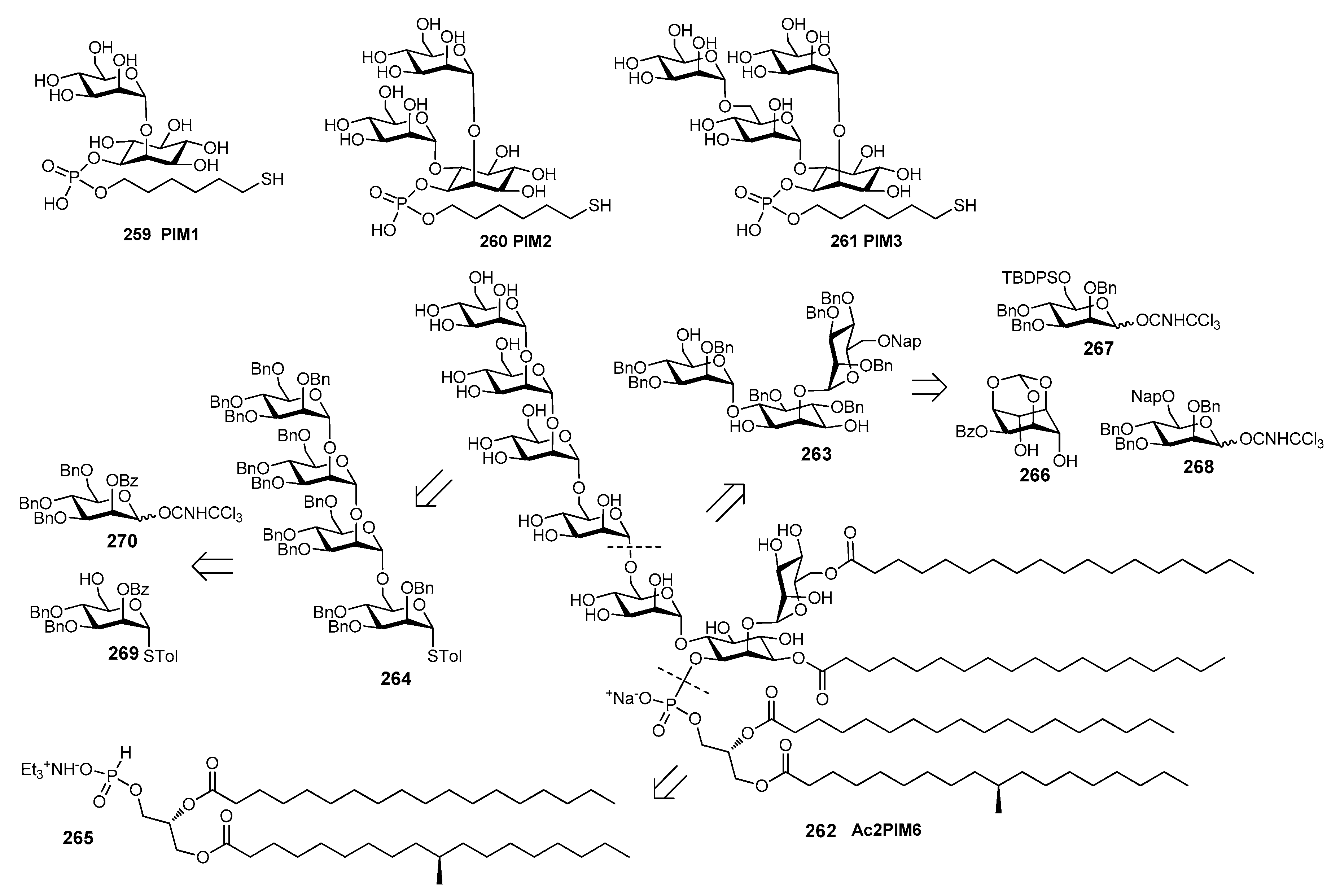
Figure 29.
LAM oligosaccharides reported in Reference [83] and retrosynthetic route to LAM tetrasaccharide coupled to a monophosphoryl lipid A (MPLA), as reported in [226].

Figure 30.
Examples of mannan, β-glucan and galactomannan components of fungal cell walls.
Figure 31.
Major components of C. Albicans cell wall: β-(1,3)-glucan and chitin (poly-β-(1,4)-N-acetylglucosamine) are the main components of inner cell wall. The outer layer is enriched with polymannans.
Figure 31.
Major components of C. Albicans cell wall: β-(1,3)-glucan and chitin (poly-β-(1,4)-N-acetylglucosamine) are the main components of inner cell wall. The outer layer is enriched with polymannans.

Figure 32.
Oligomannans and glycoconjugates synthesized by the Bundle group [236].
Figure 32.
Oligomannans and glycoconjugates synthesized by the Bundle group [236].
Figure 33.
Repeating unit of Lamarin (Lam) and Lam fragments reported in [247].
Figure 33.
Repeating unit of Lamarin (Lam) and Lam fragments reported in [247].
Figure 34.
Hexa-, octa-, deca- and dodecasaccharide of β-glucan reported in Reference [248].
Figure 34.
Hexa-, octa-, deca- and dodecasaccharide of β-glucan reported in Reference [248].

Figure 35.
Structure of fragments 285a–287a and their KLH conjugates [249].
Figure 35.
Structure of fragments 285a–287a and their KLH conjugates [249].
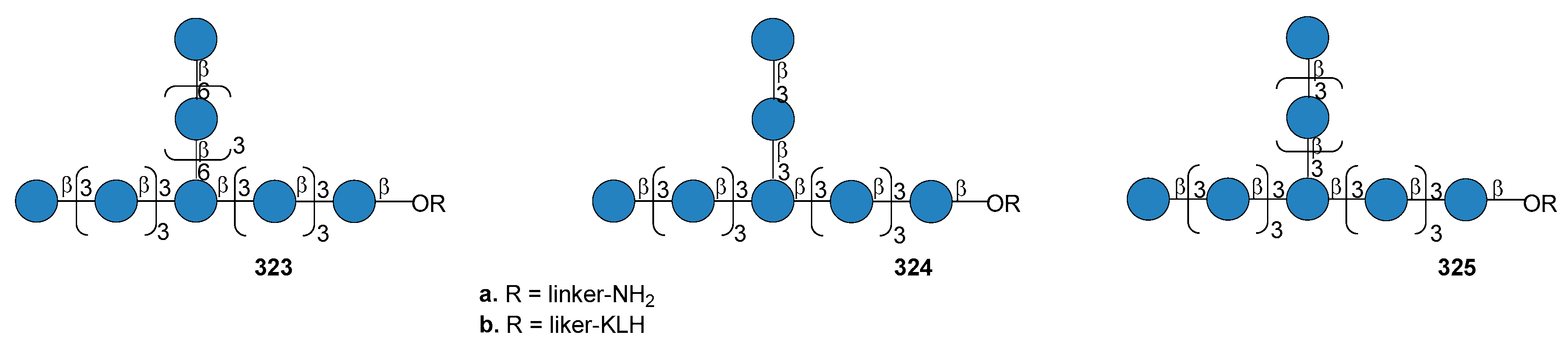
Figure 36.
Retrosynthetic analysis of pentasaccharide 326a, as reported in [250].
Figure 36.
Retrosynthetic analysis of pentasaccharide 326a, as reported in [250].

Figure 37.
A. fumigatus galactomannan structure, underlining the presence of β-(1,6)-linked Galf residues in addition to the β-(1,5)-linked Galf residues, as reported in [252].
Figure 37.
A. fumigatus galactomannan structure, underlining the presence of β-(1,6)-linked Galf residues in addition to the β-(1,5)-linked Galf residues, as reported in [252].
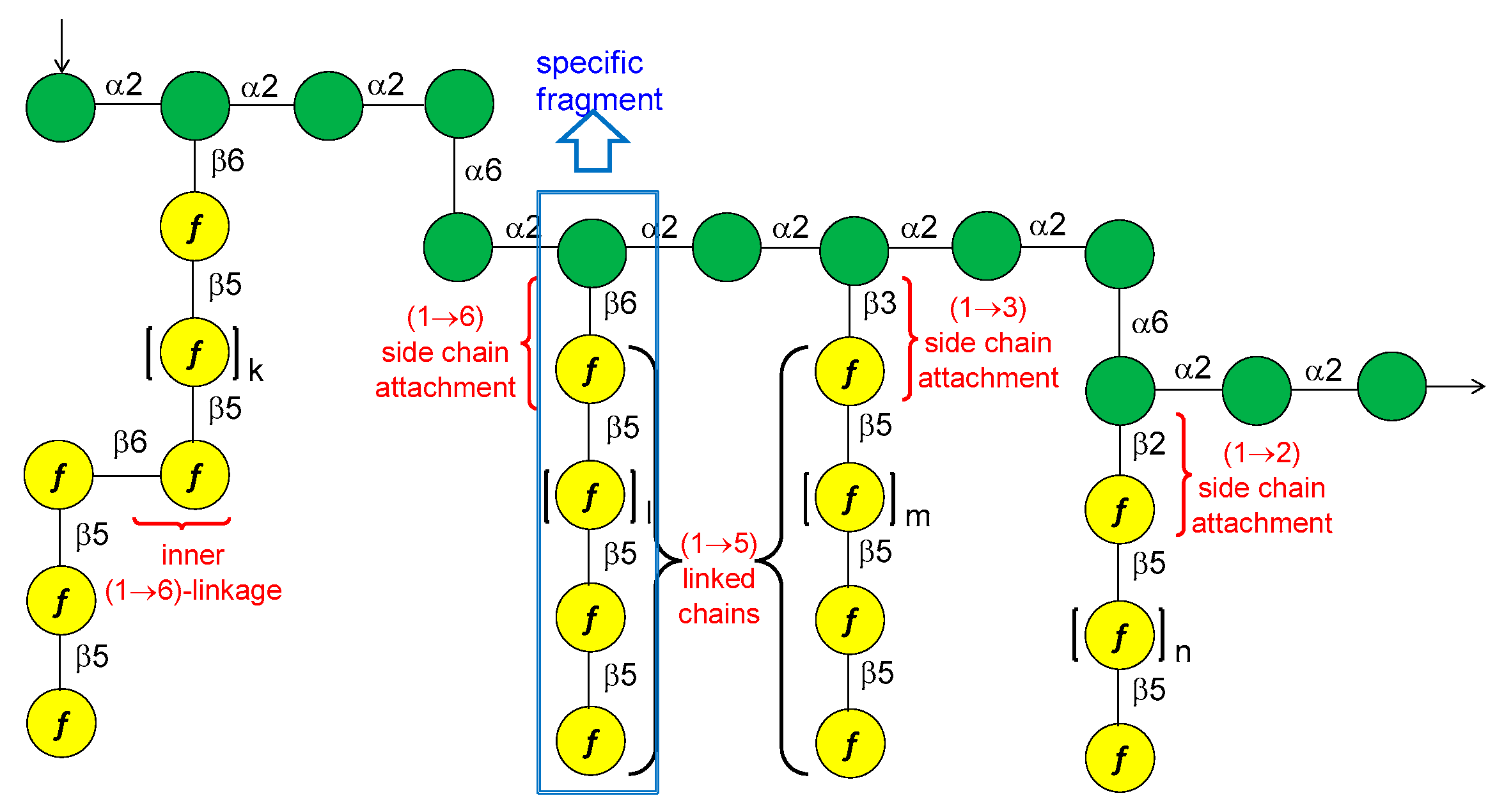

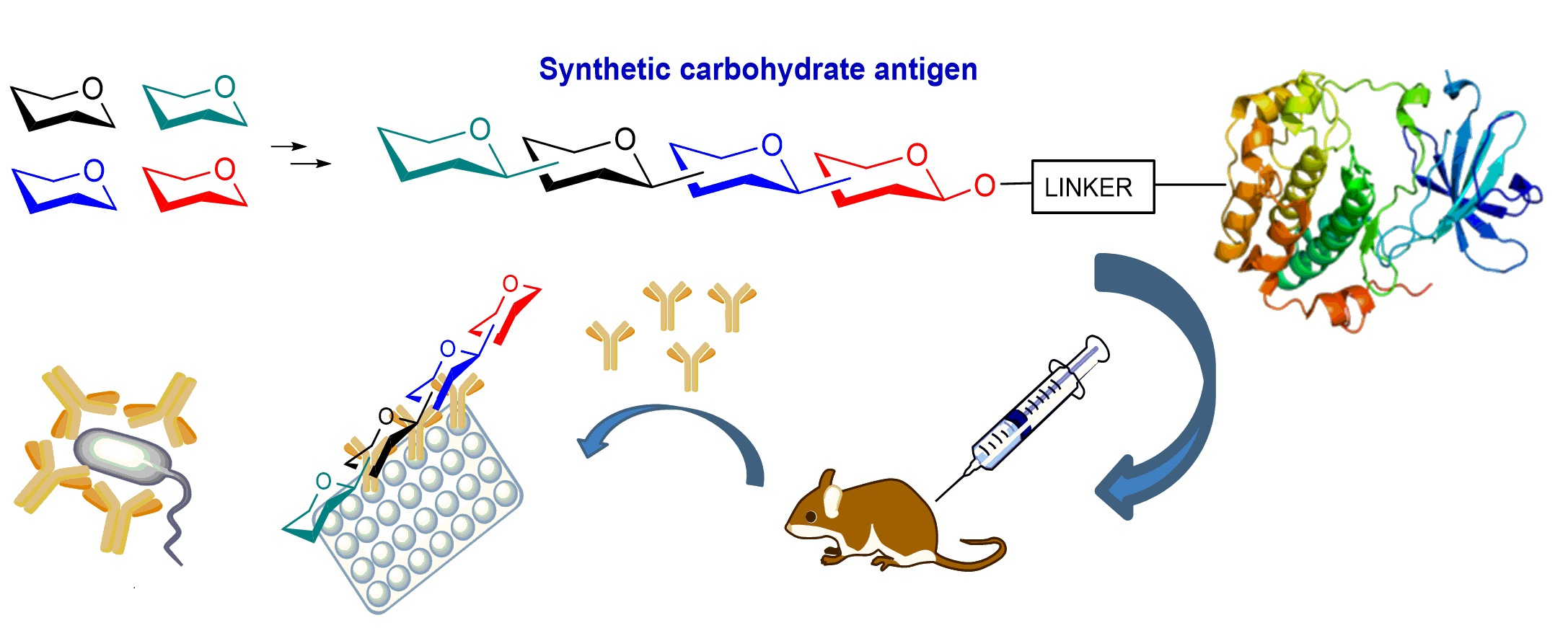{kind=link}
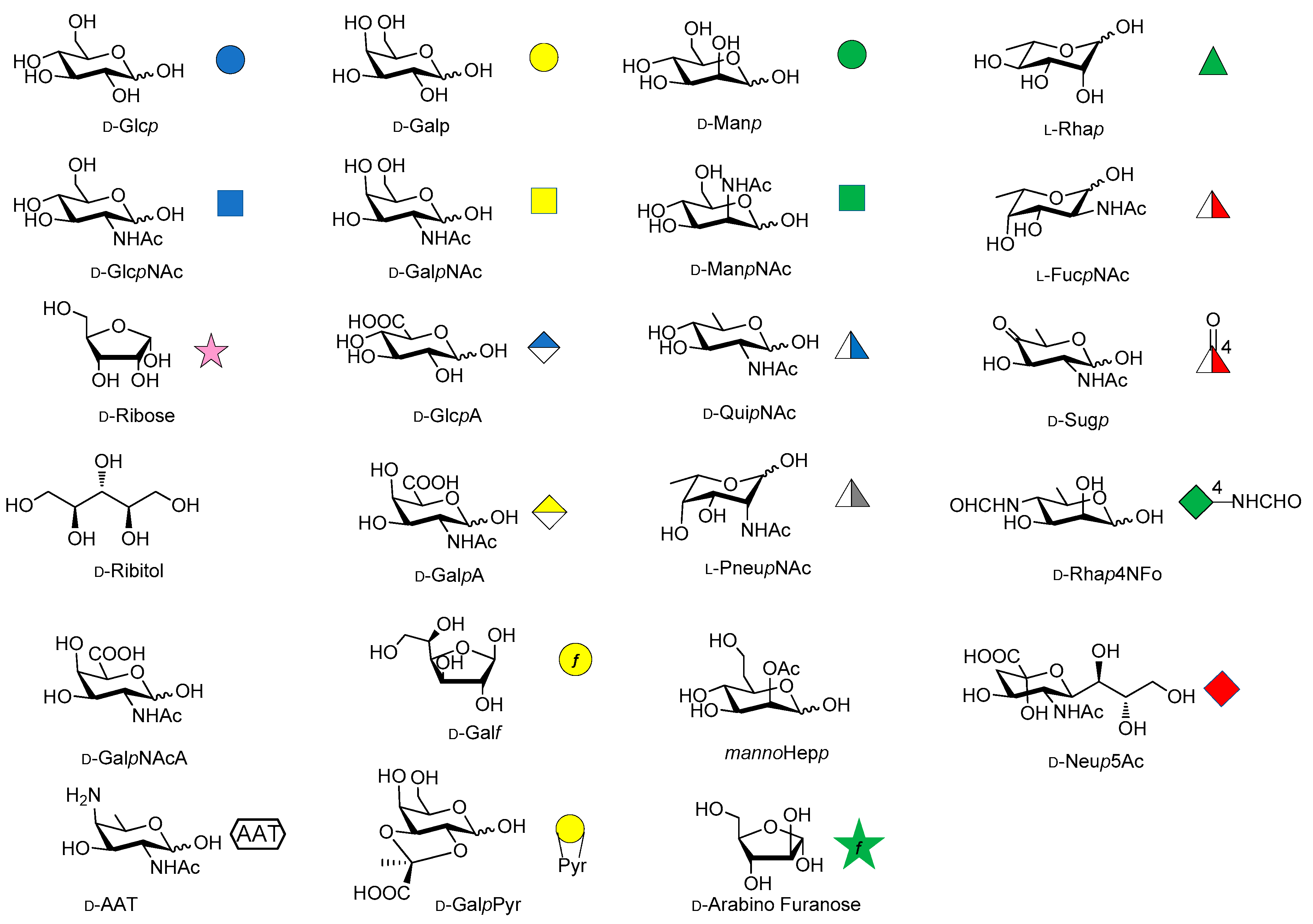{kind=link}
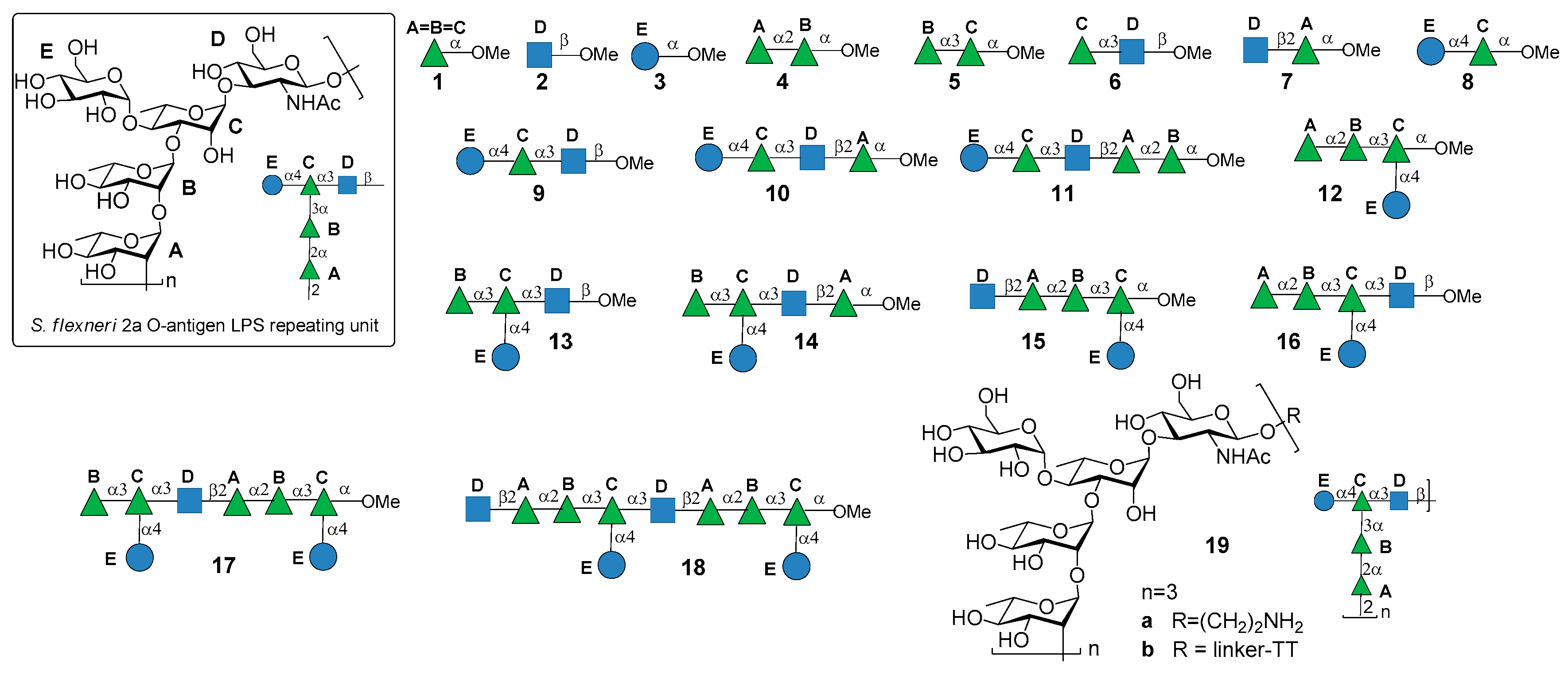{kind=link}
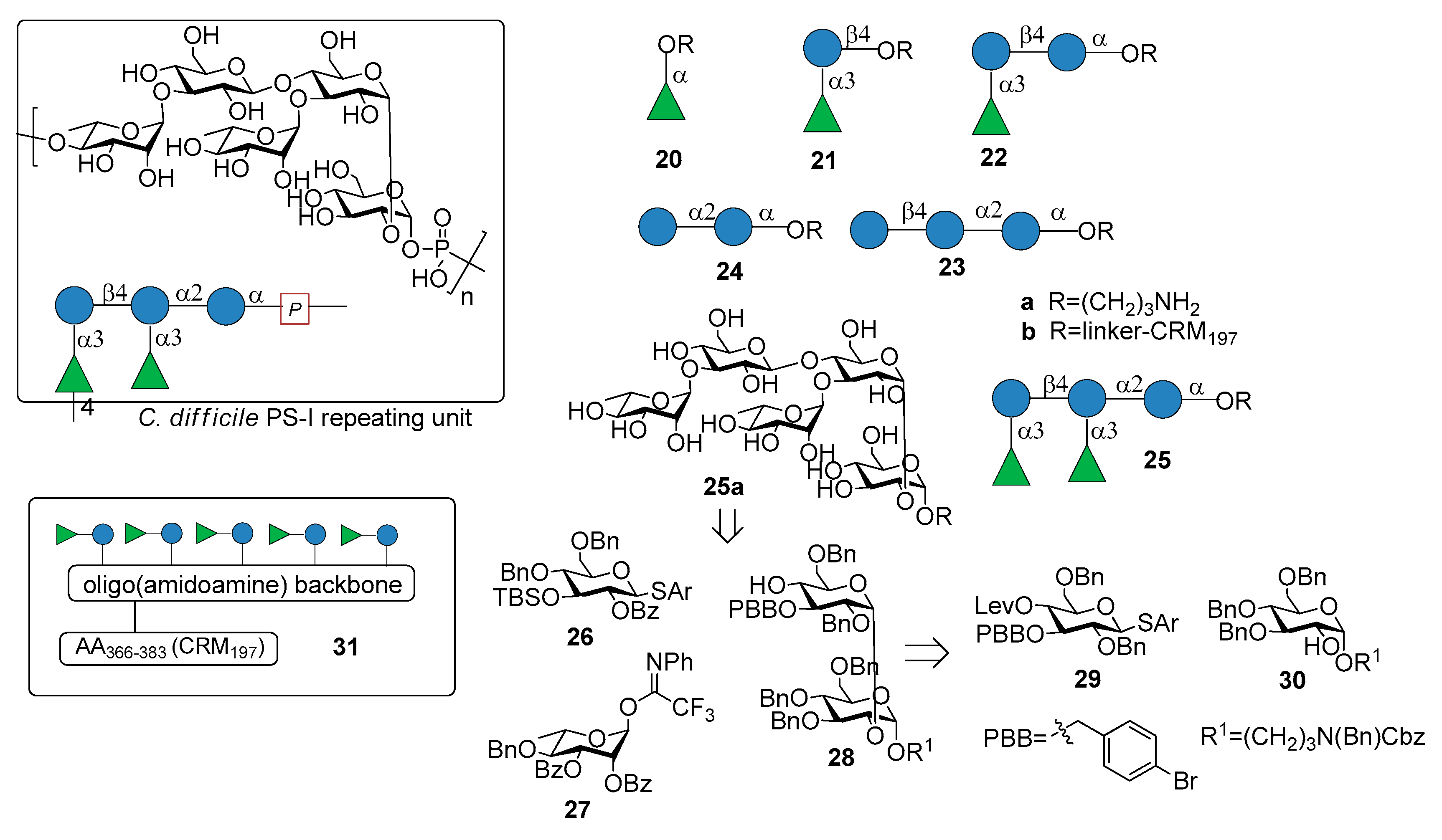{kind=link}
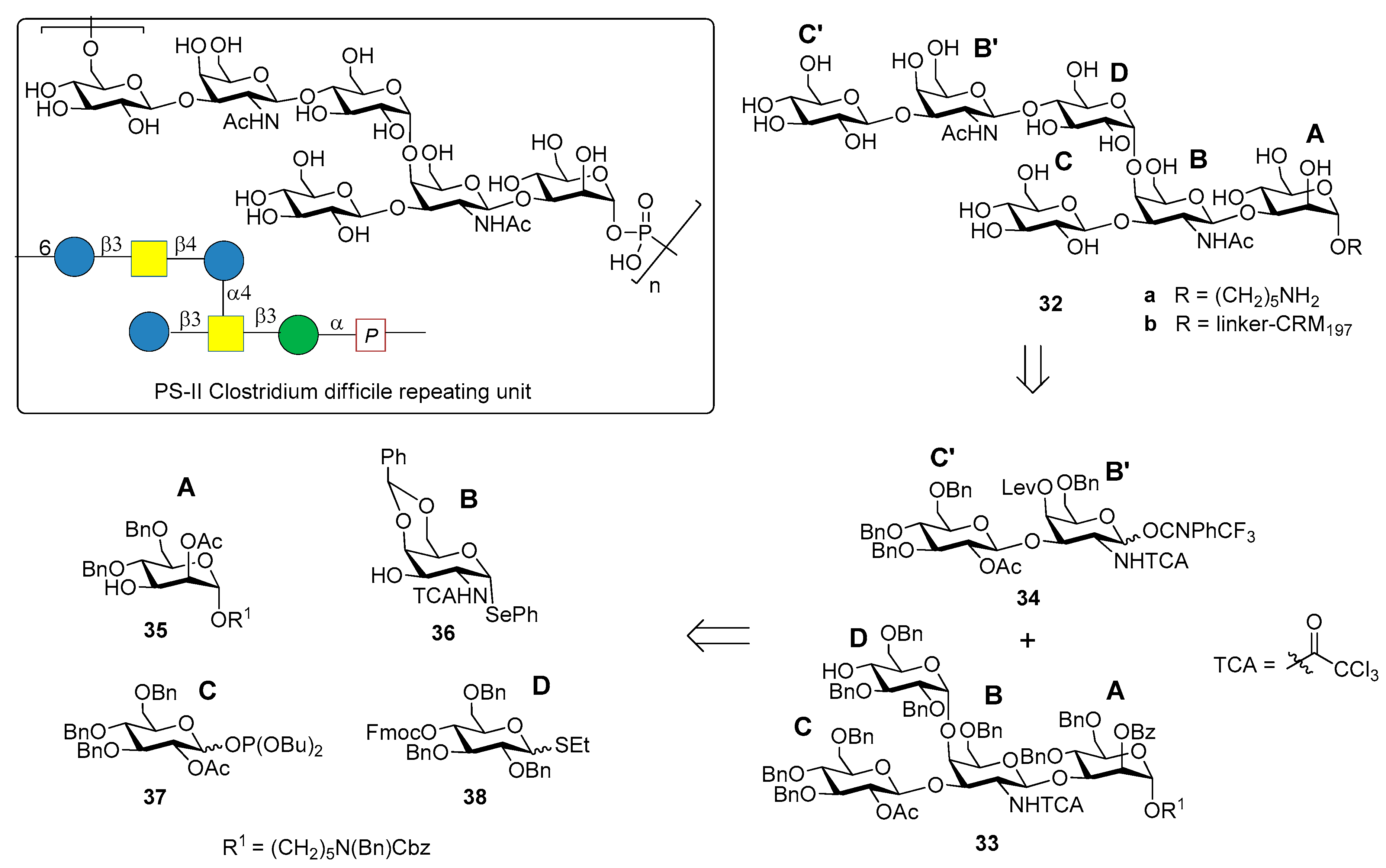{kind=link}
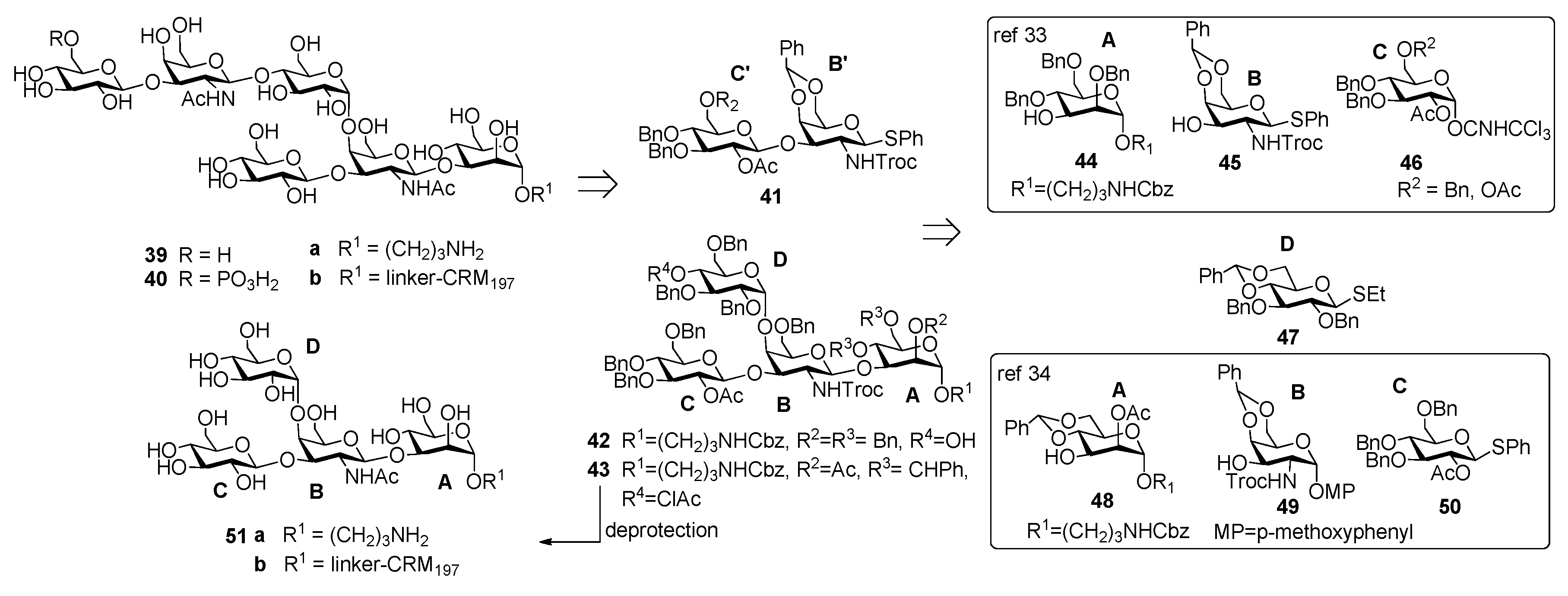{kind=link}
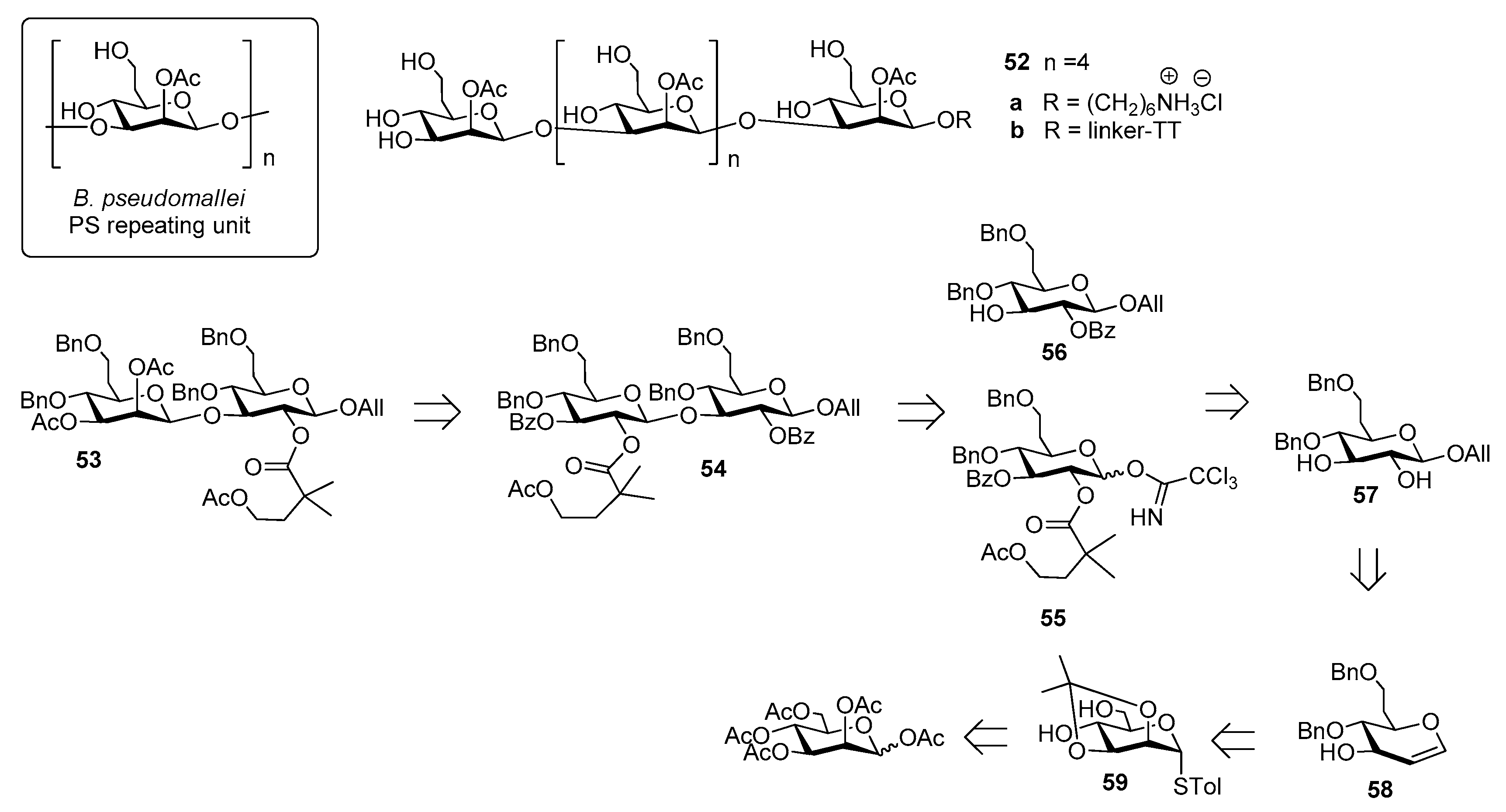{kind=link}
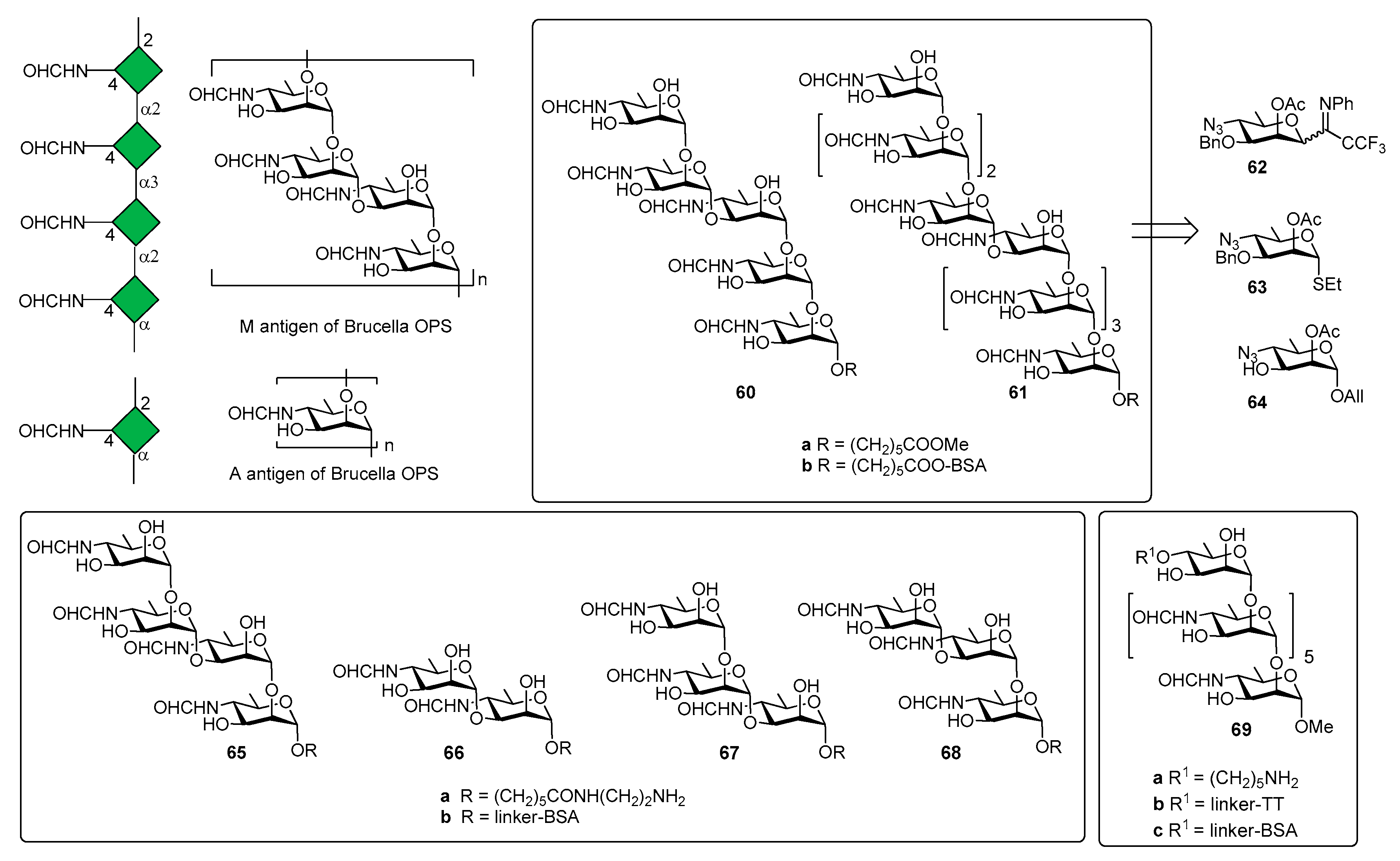{kind=link}
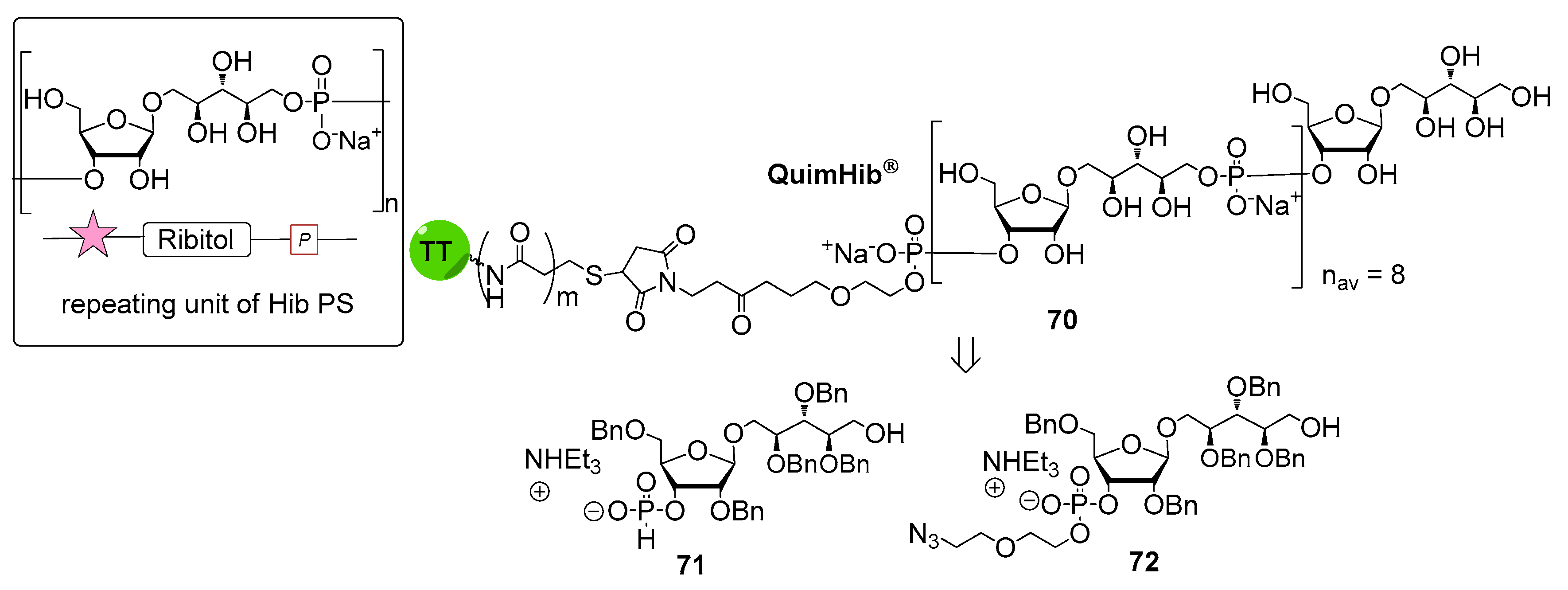{kind=link}
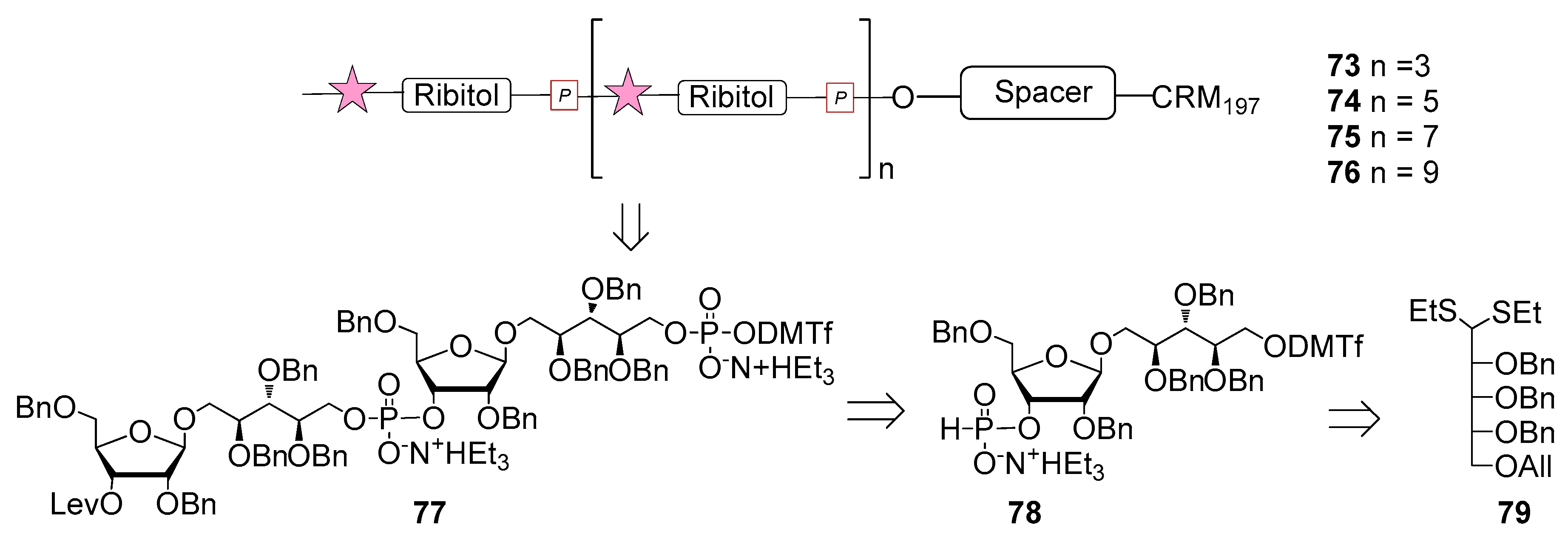{kind=link}
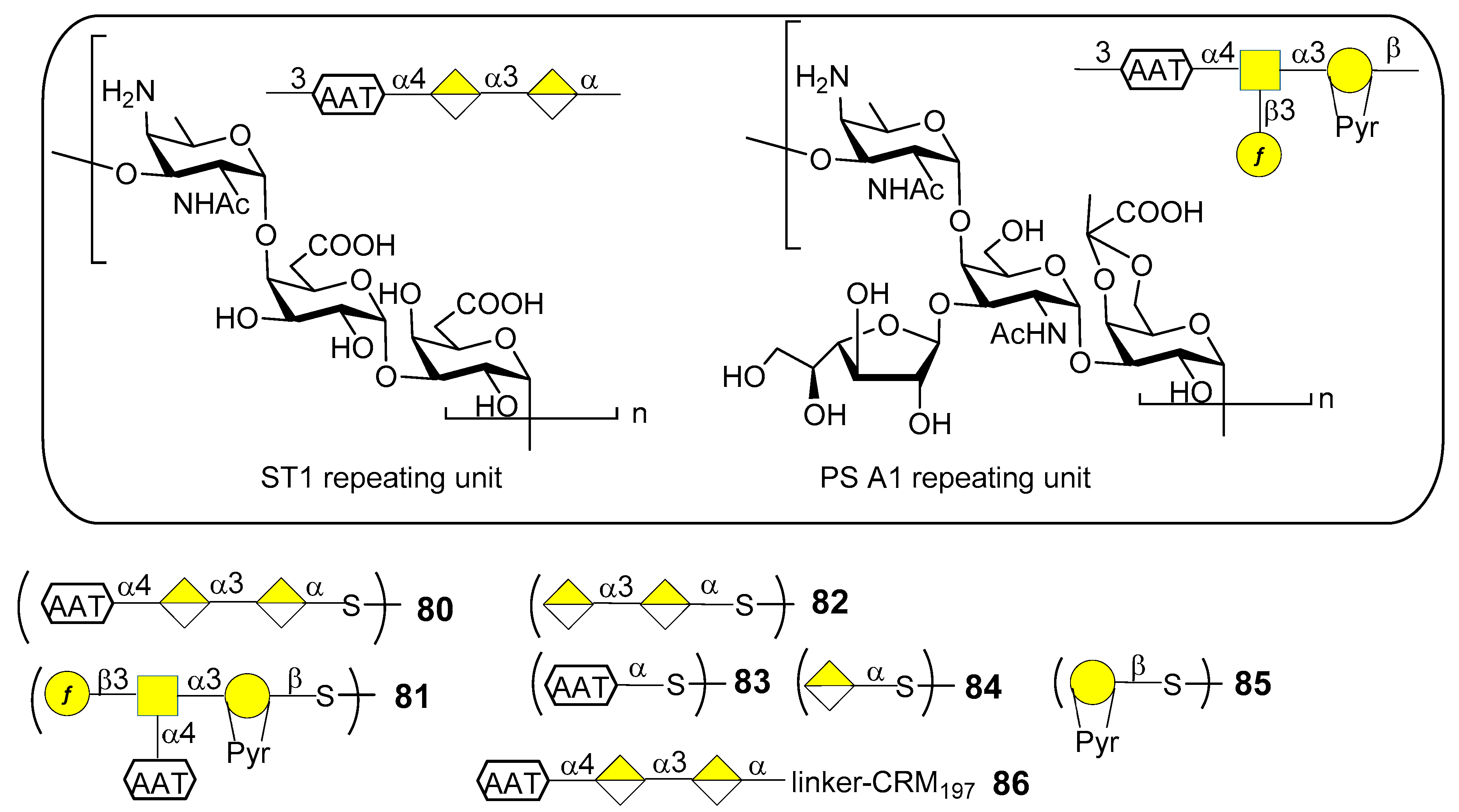{kind=link}
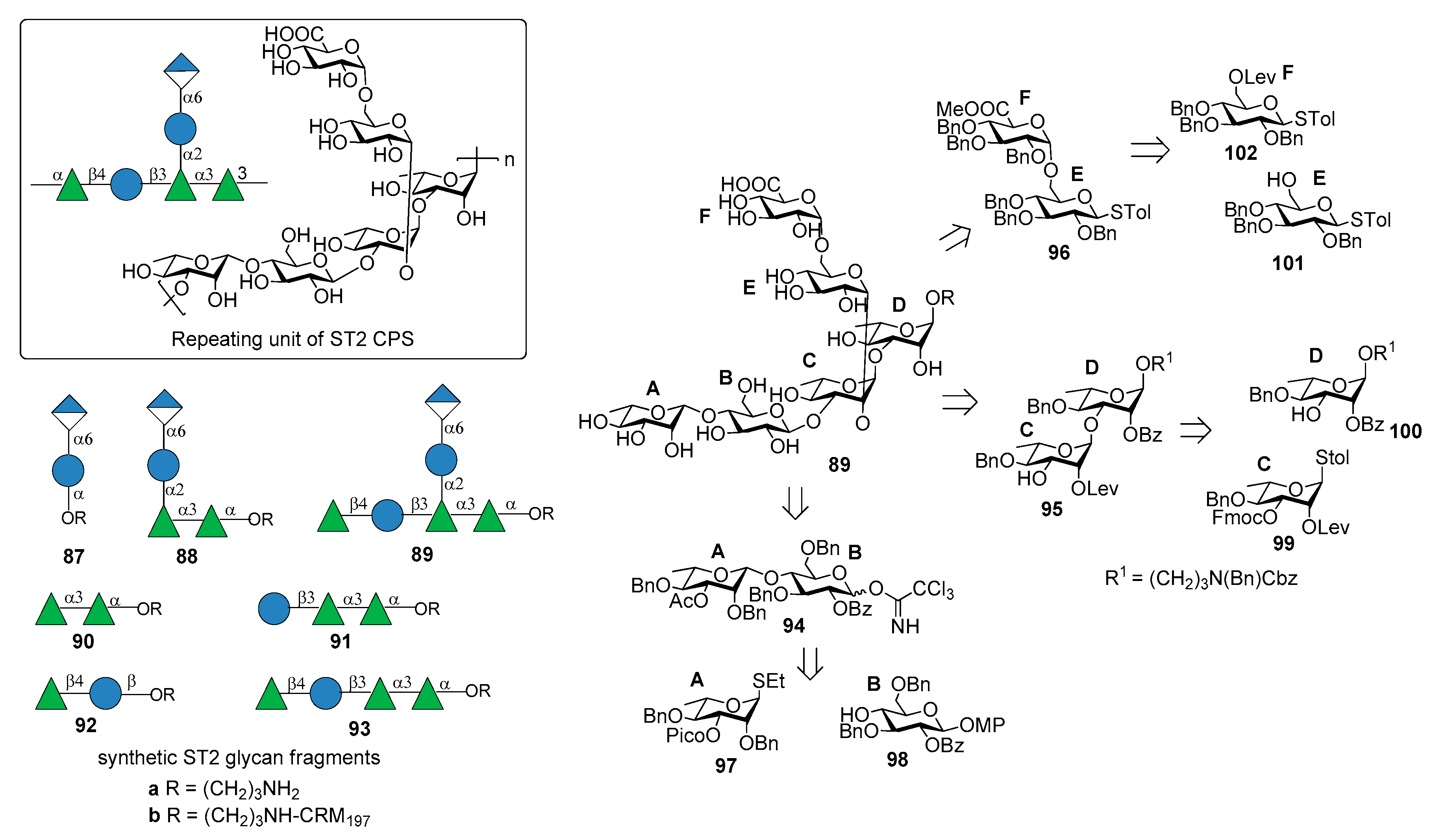{kind=link}
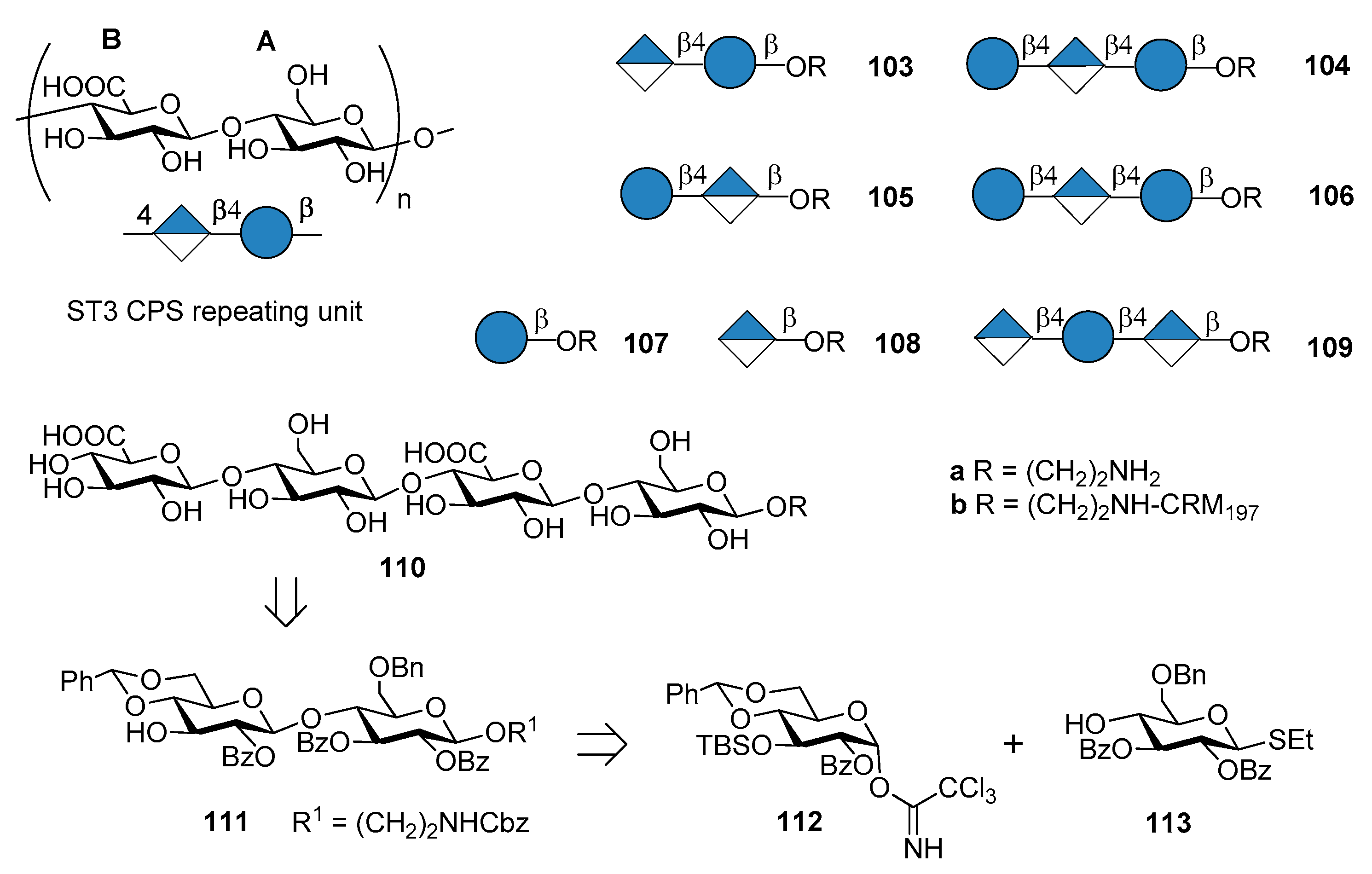{kind=link}
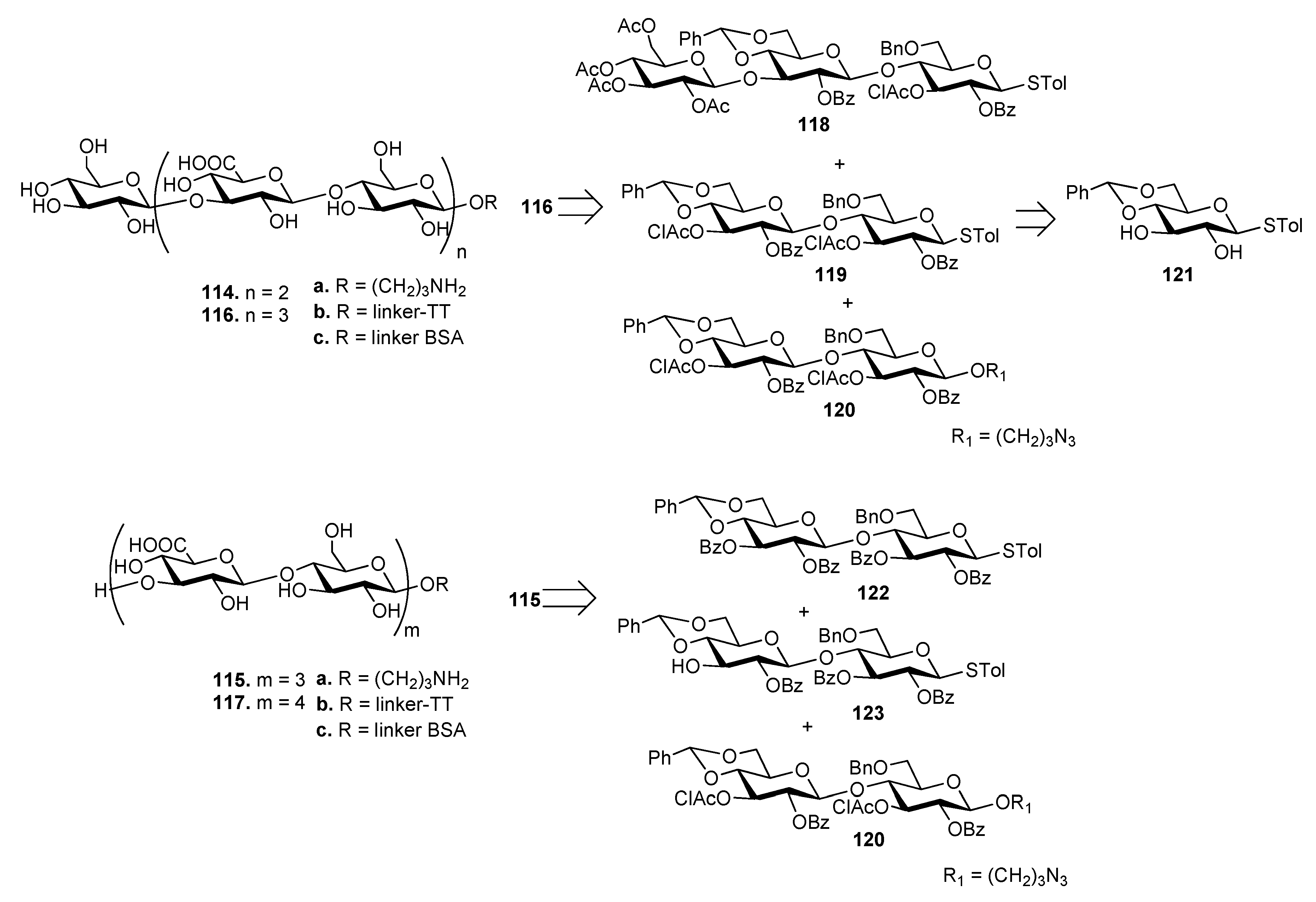{kind=link}
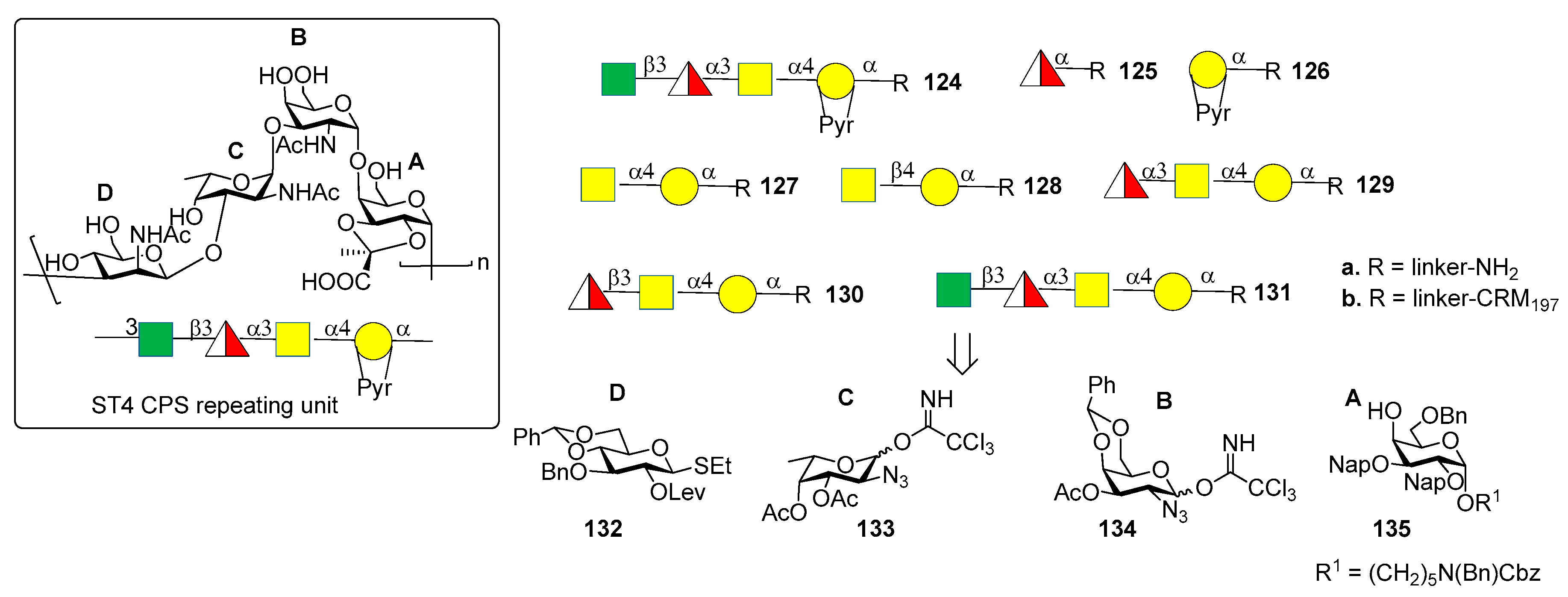{kind=link}
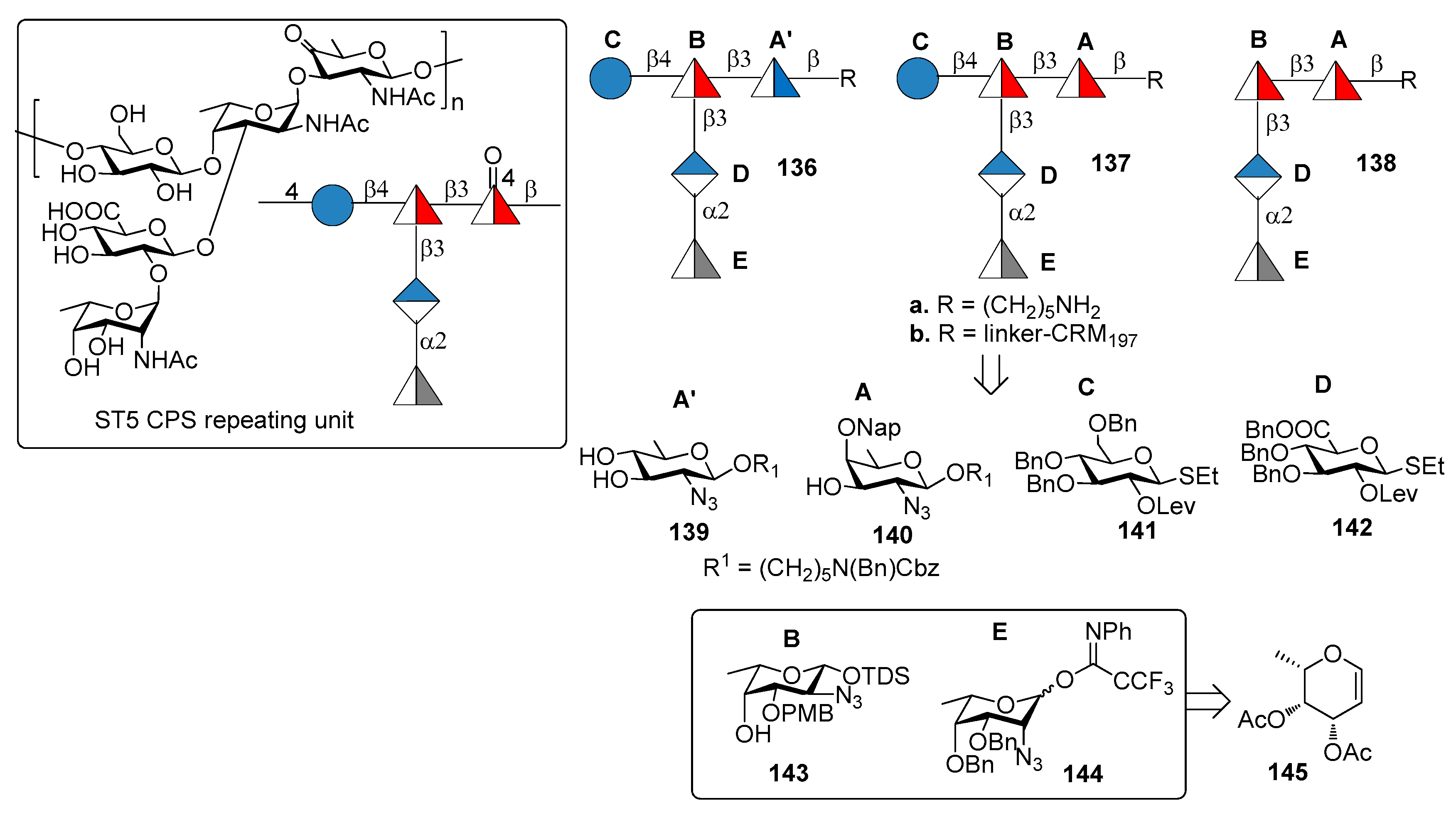{kind=link}
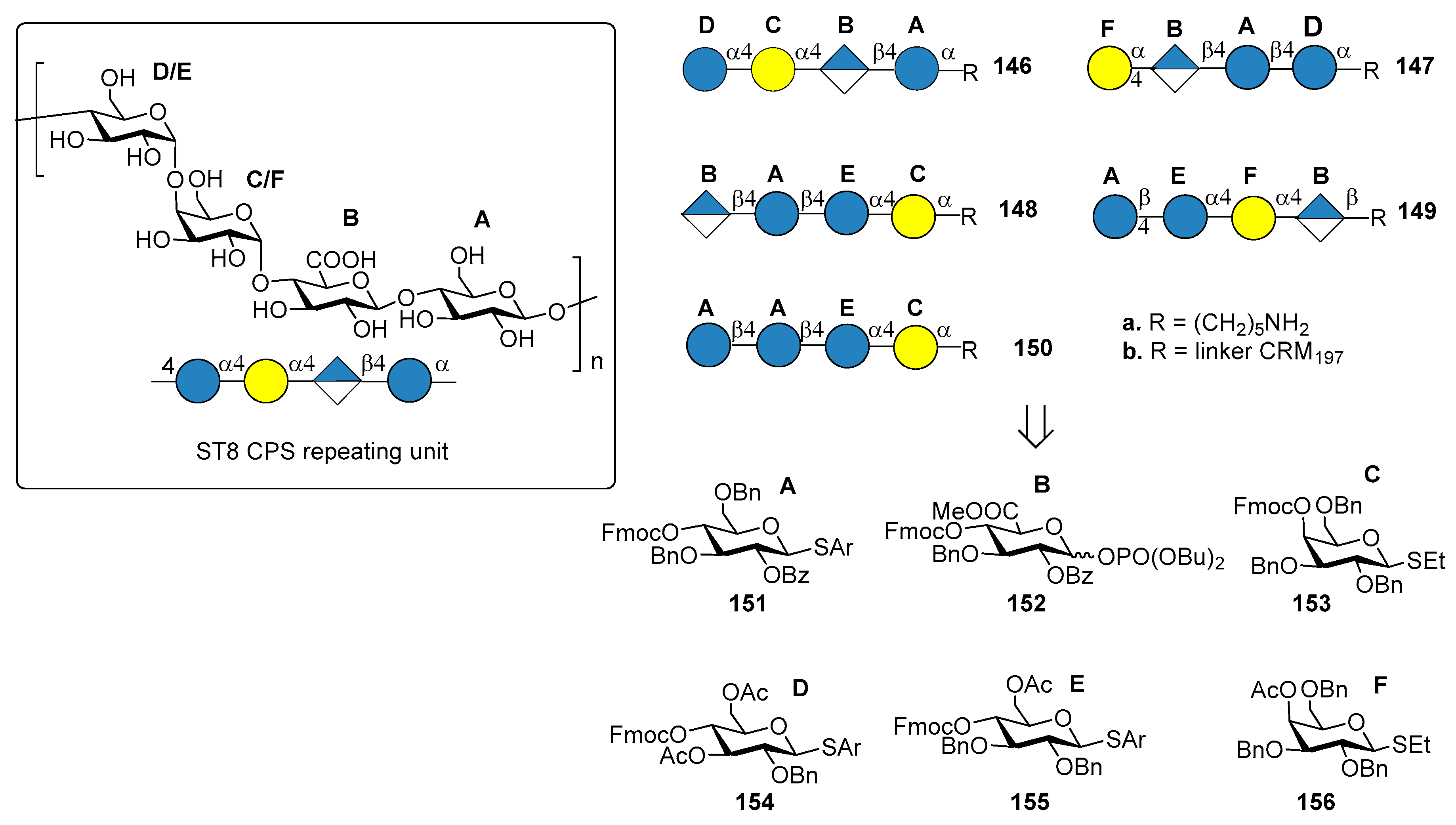{kind=link}
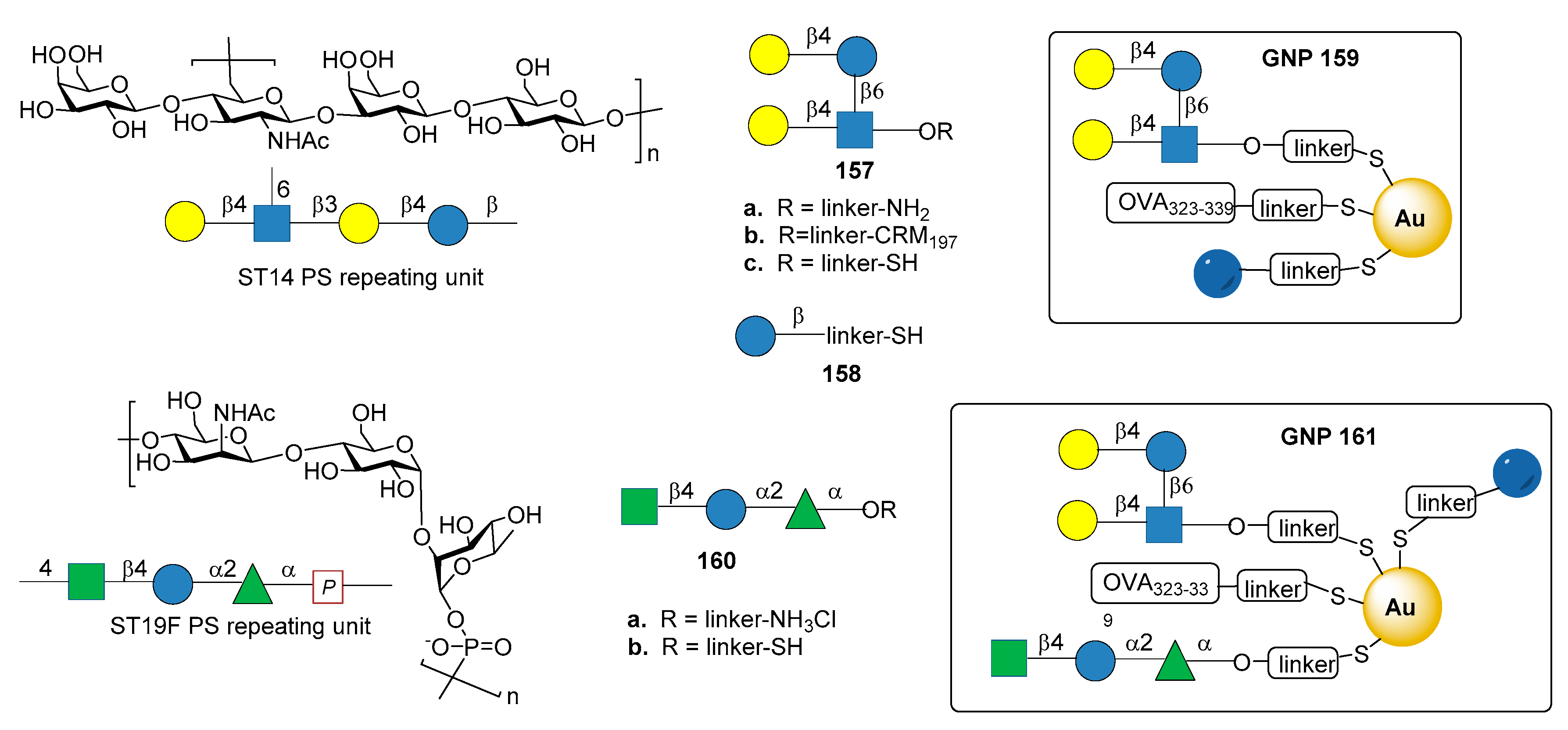{kind=link}
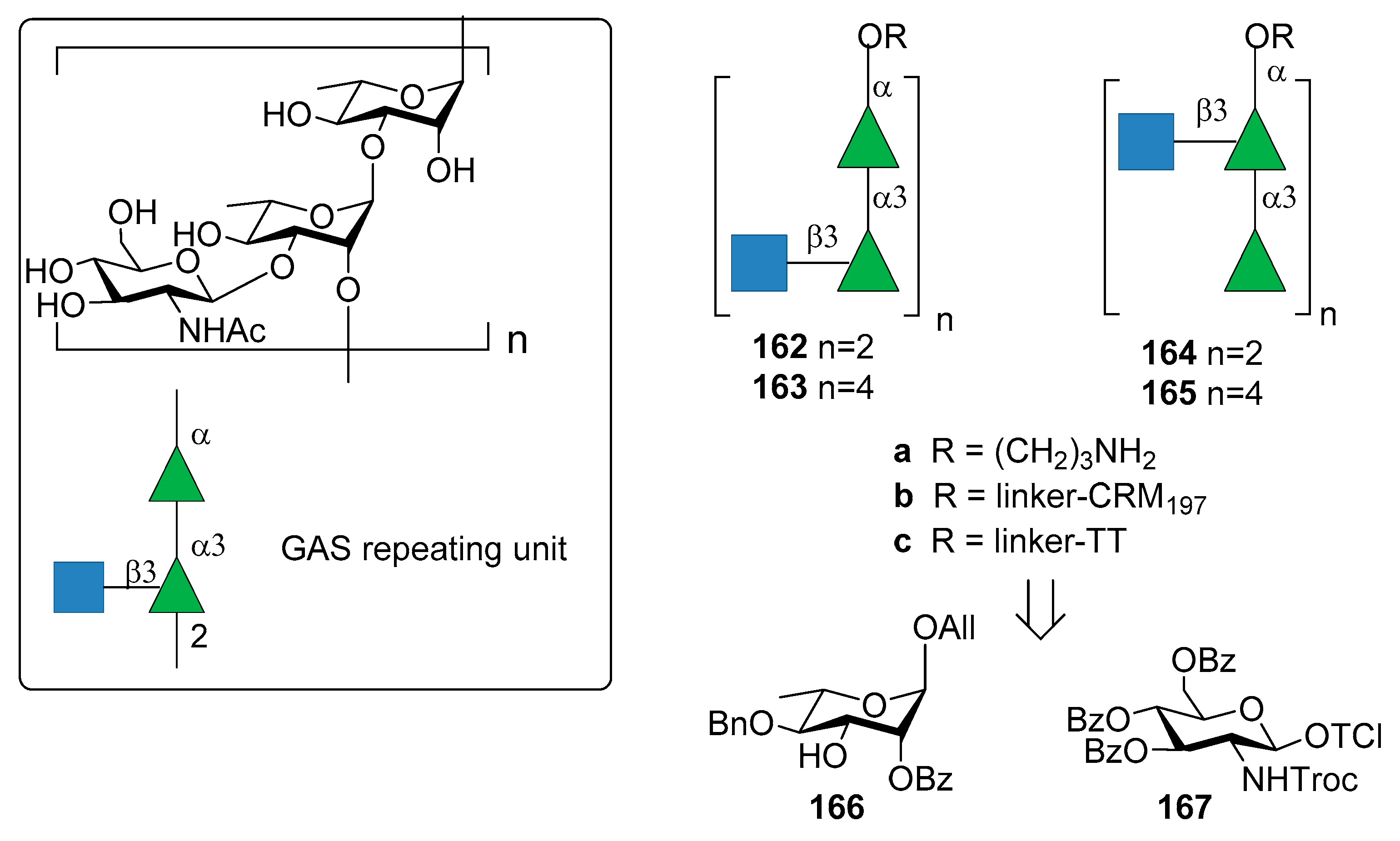{kind=link}
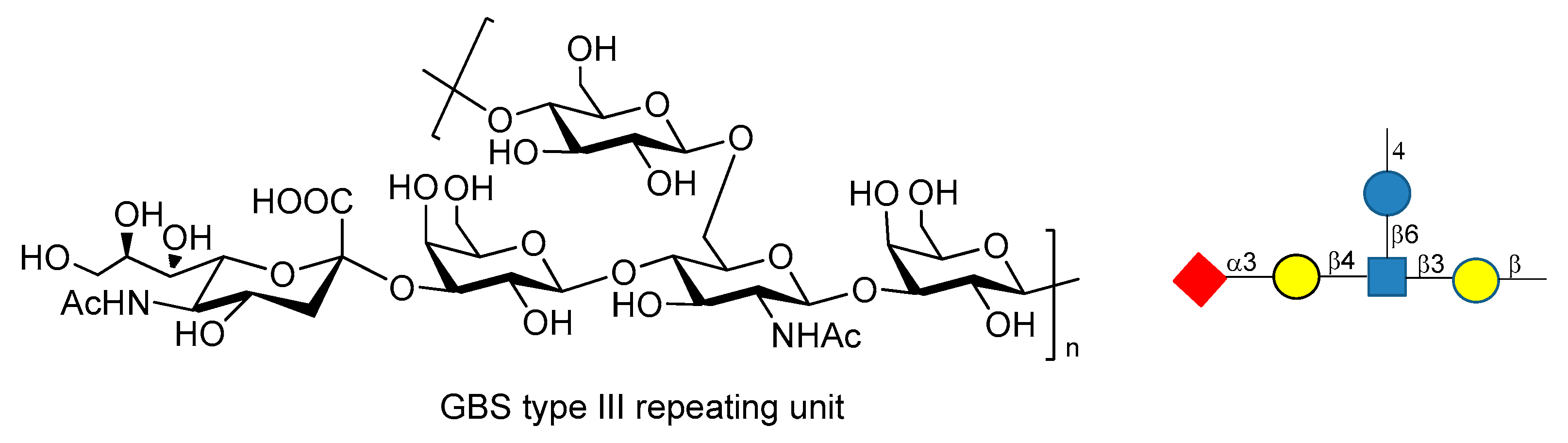{kind=link}
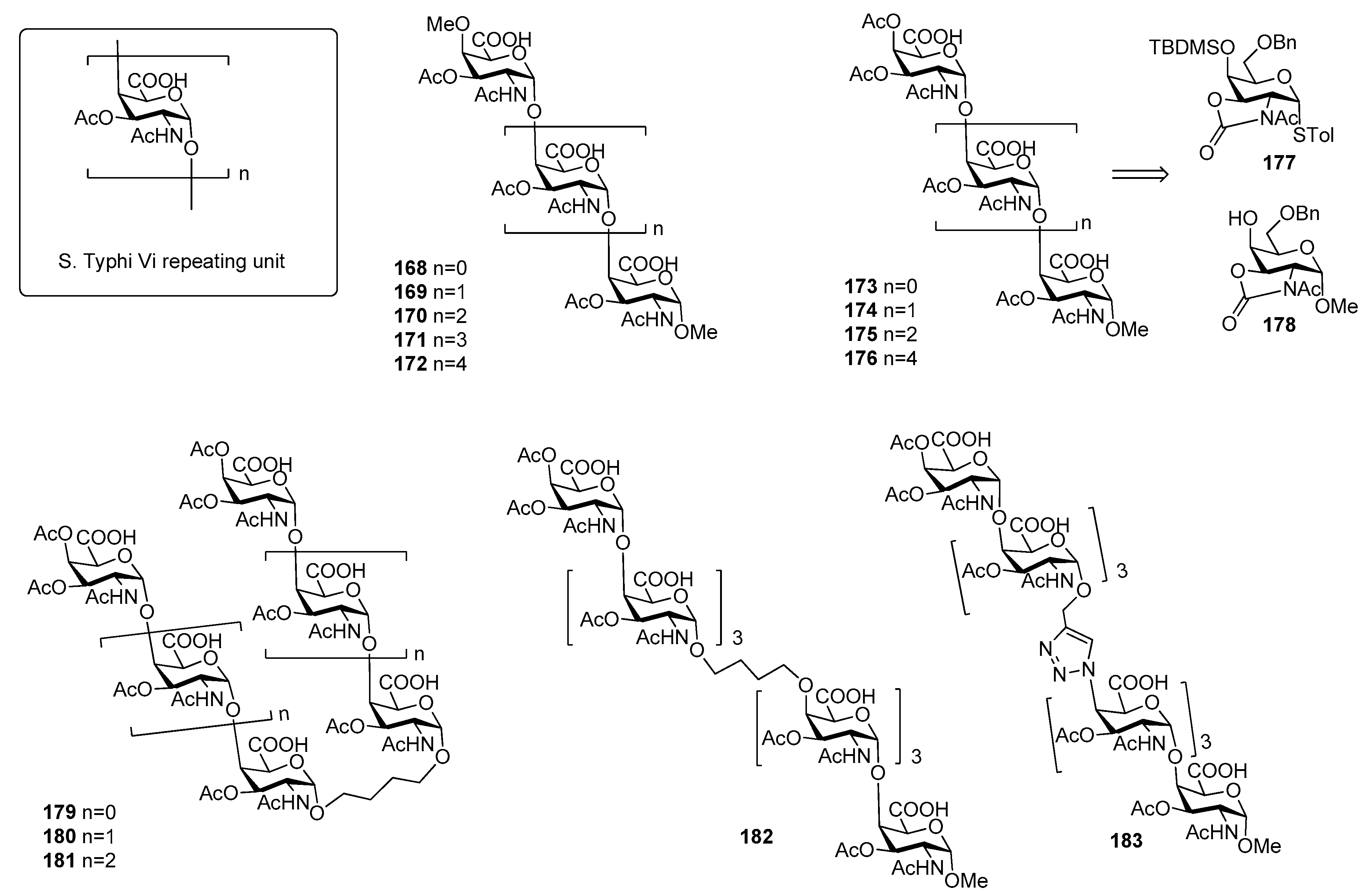{kind=link}
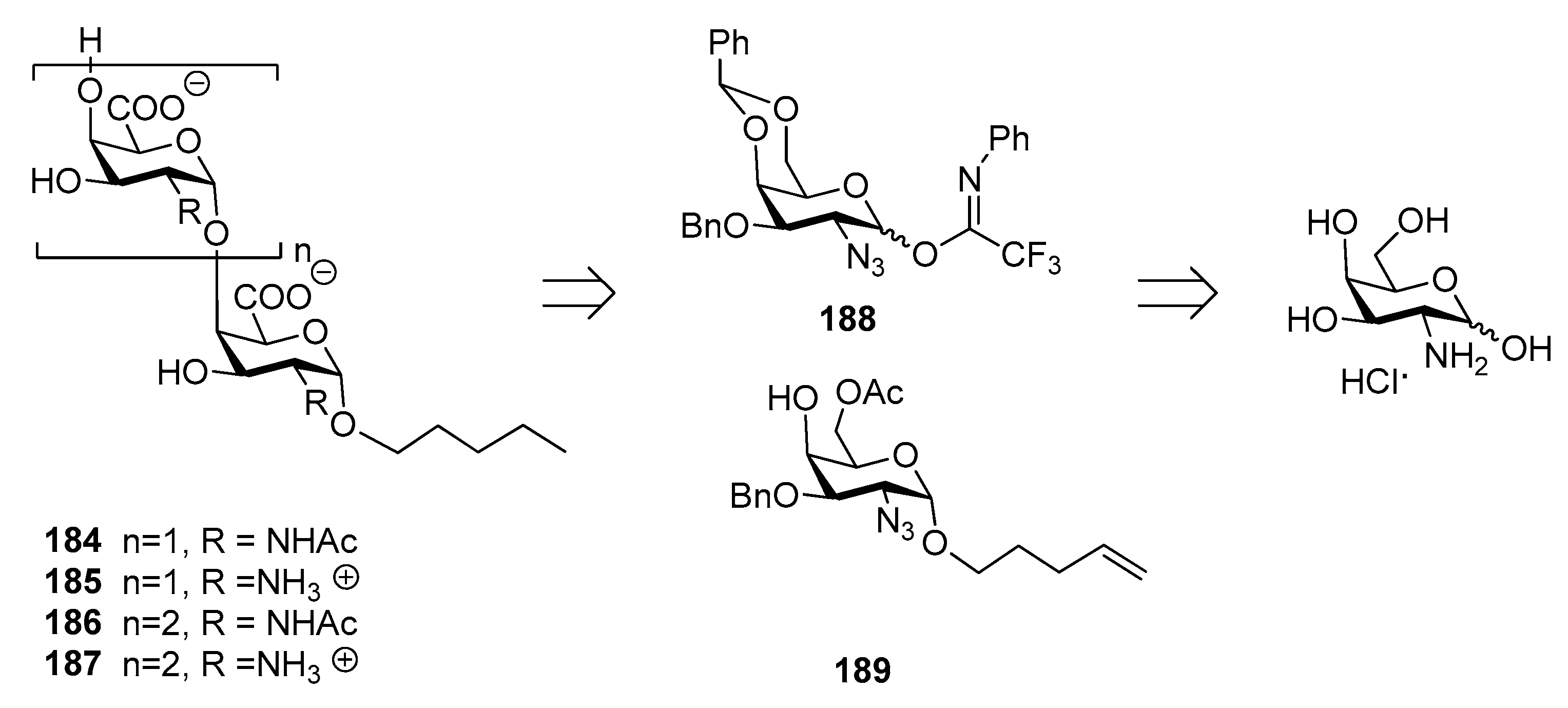{kind=link}
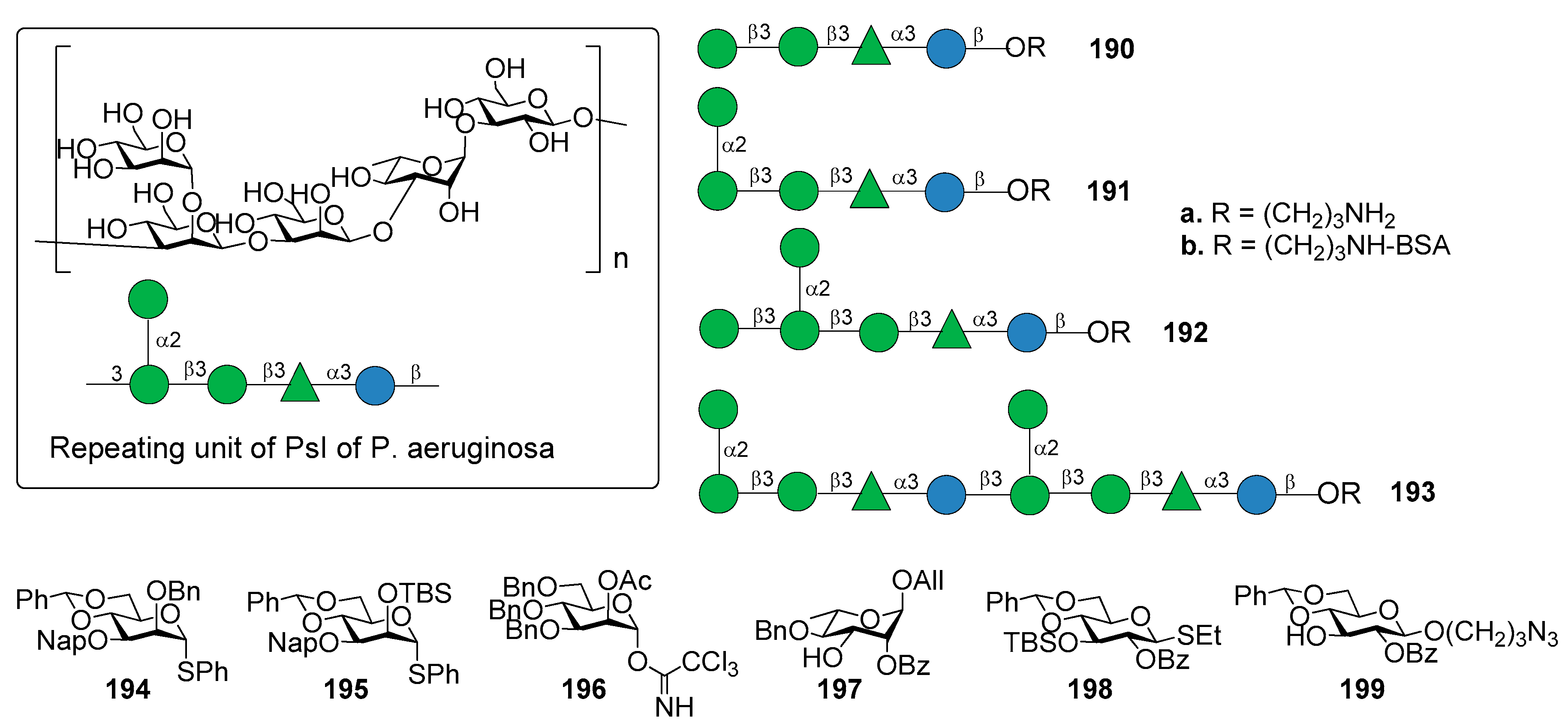{kind=link}
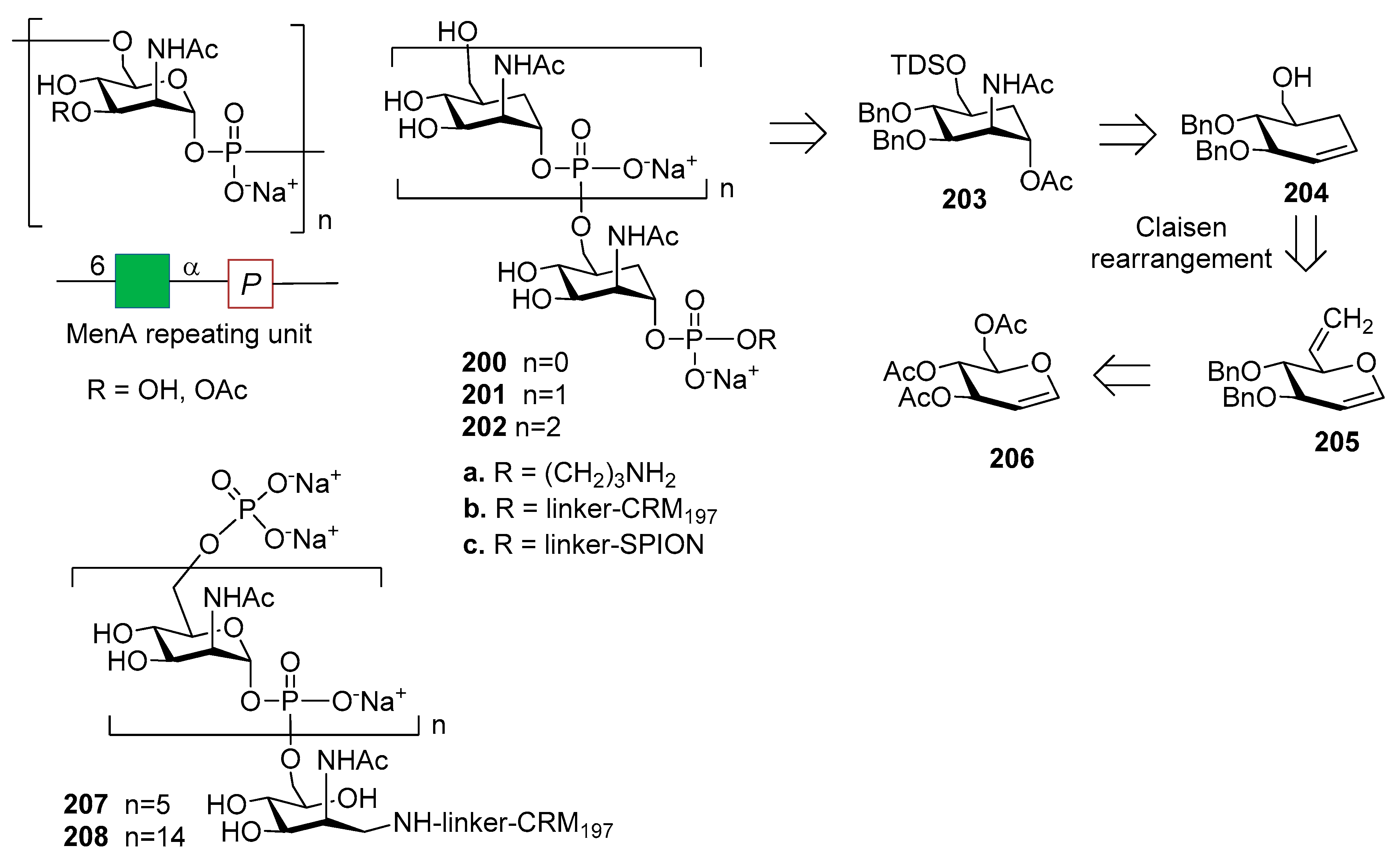{kind=link}
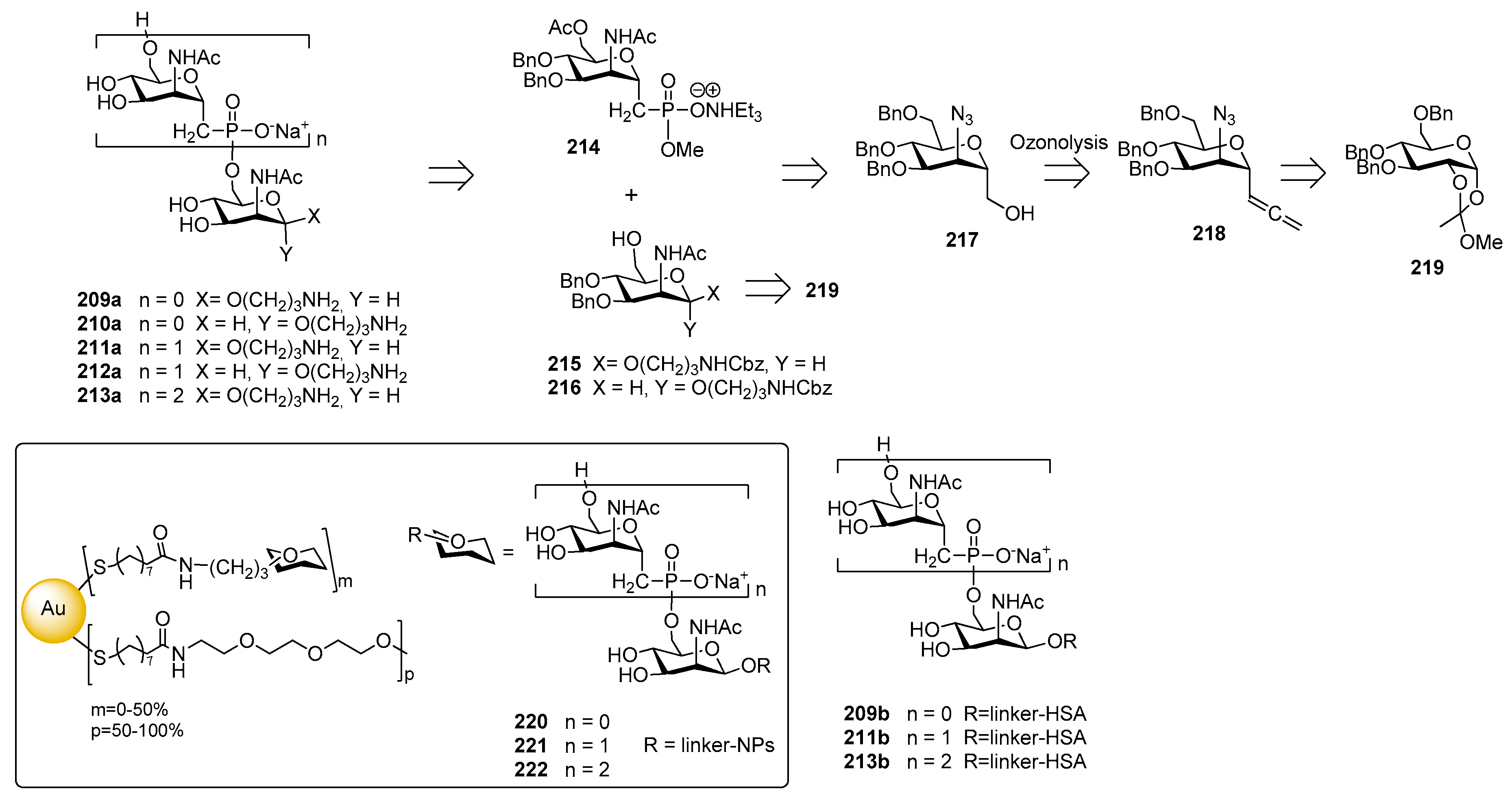{kind=link}
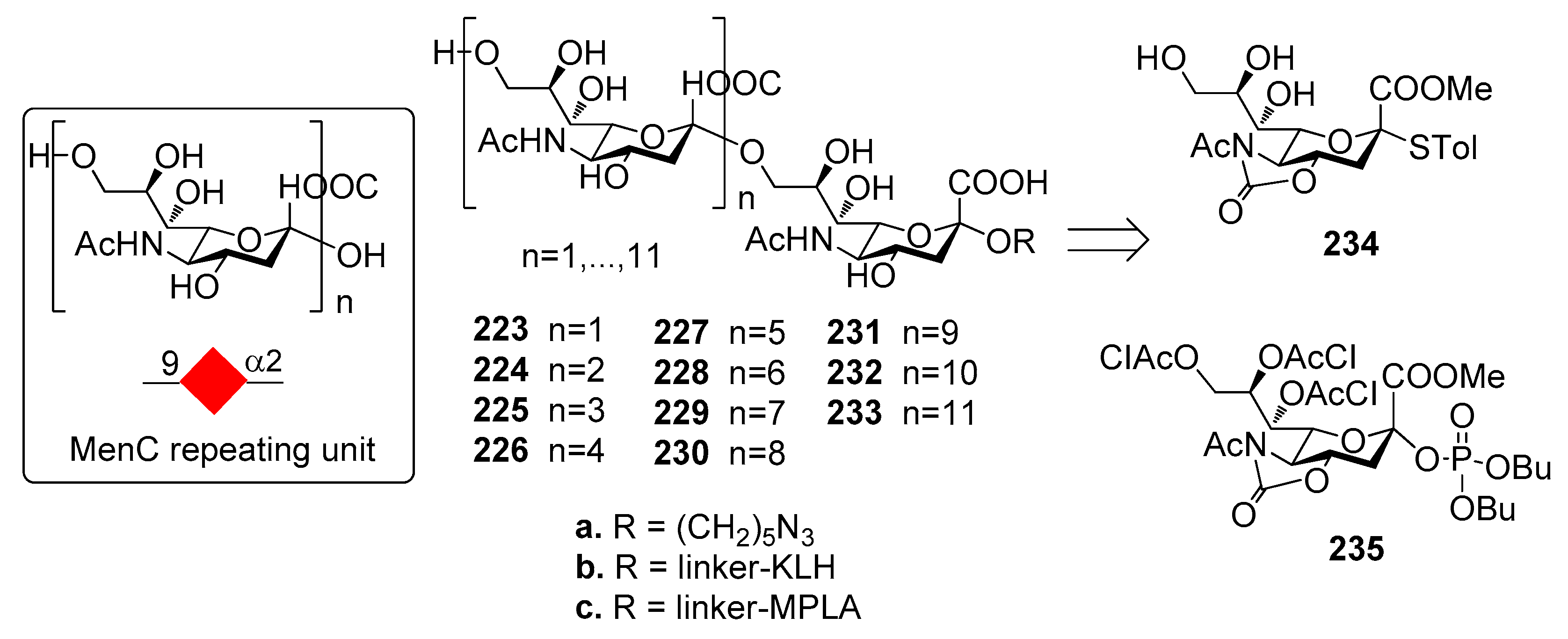{kind=link}
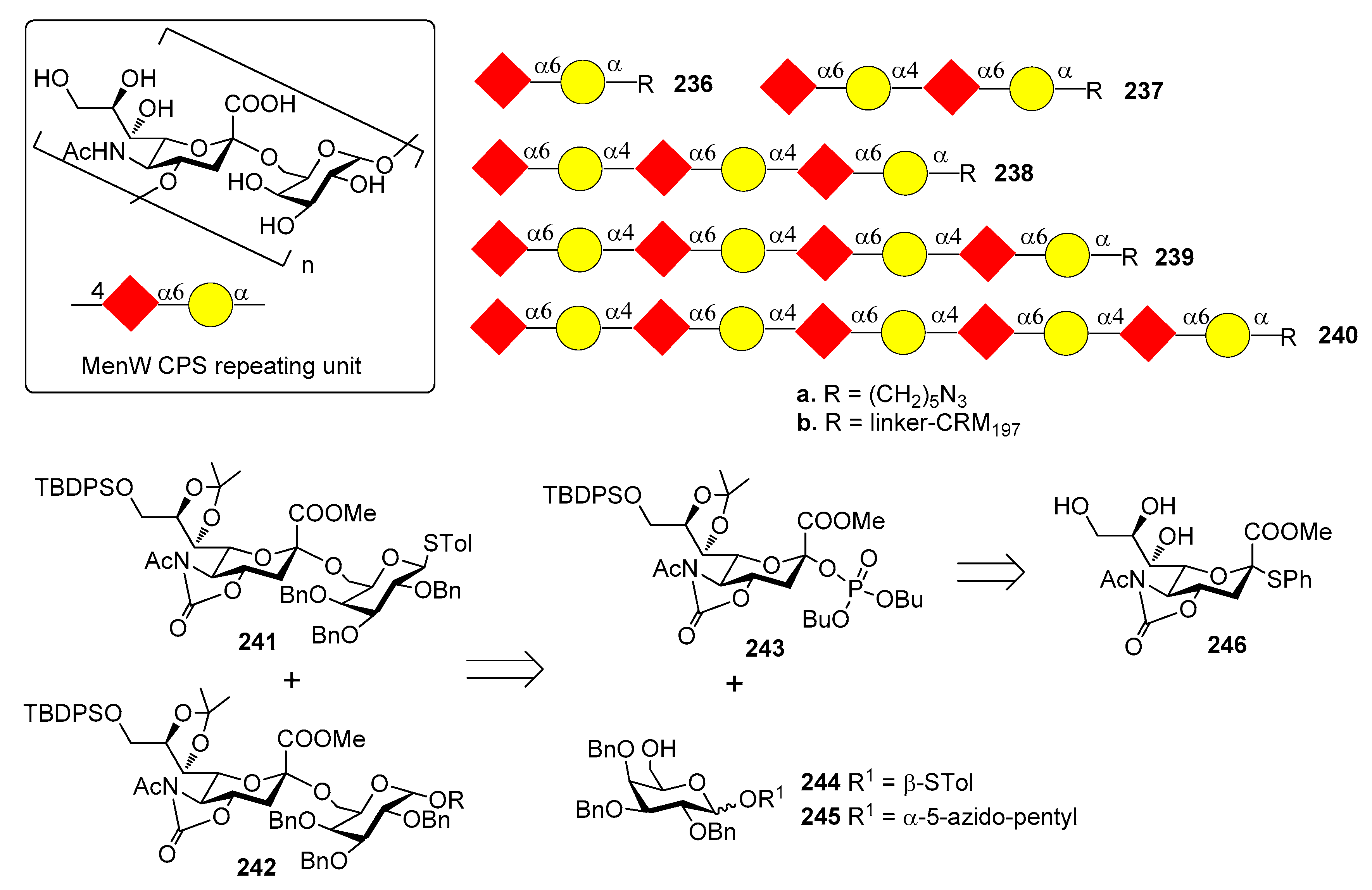{kind=link}
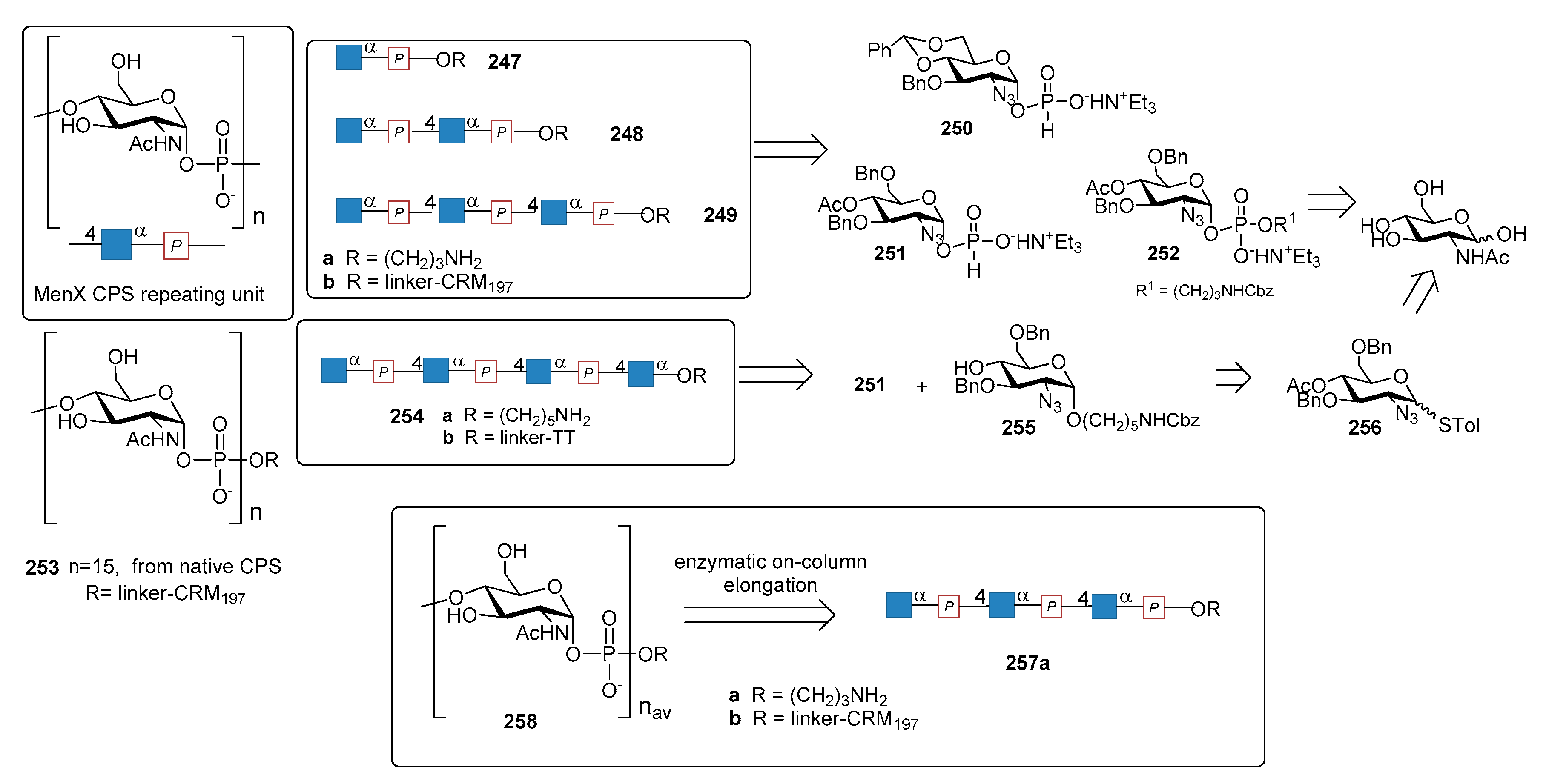{kind=link}
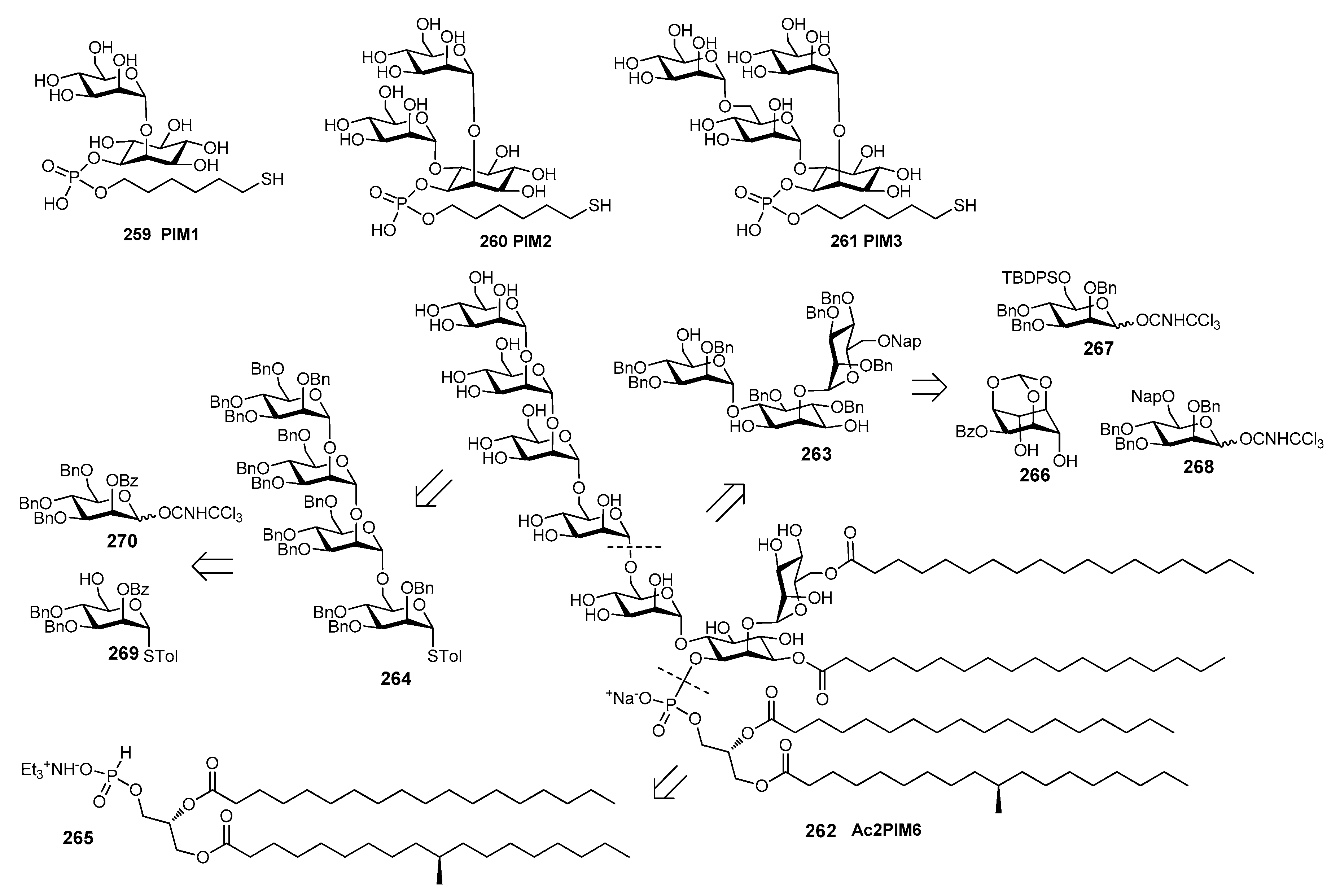{kind=link}
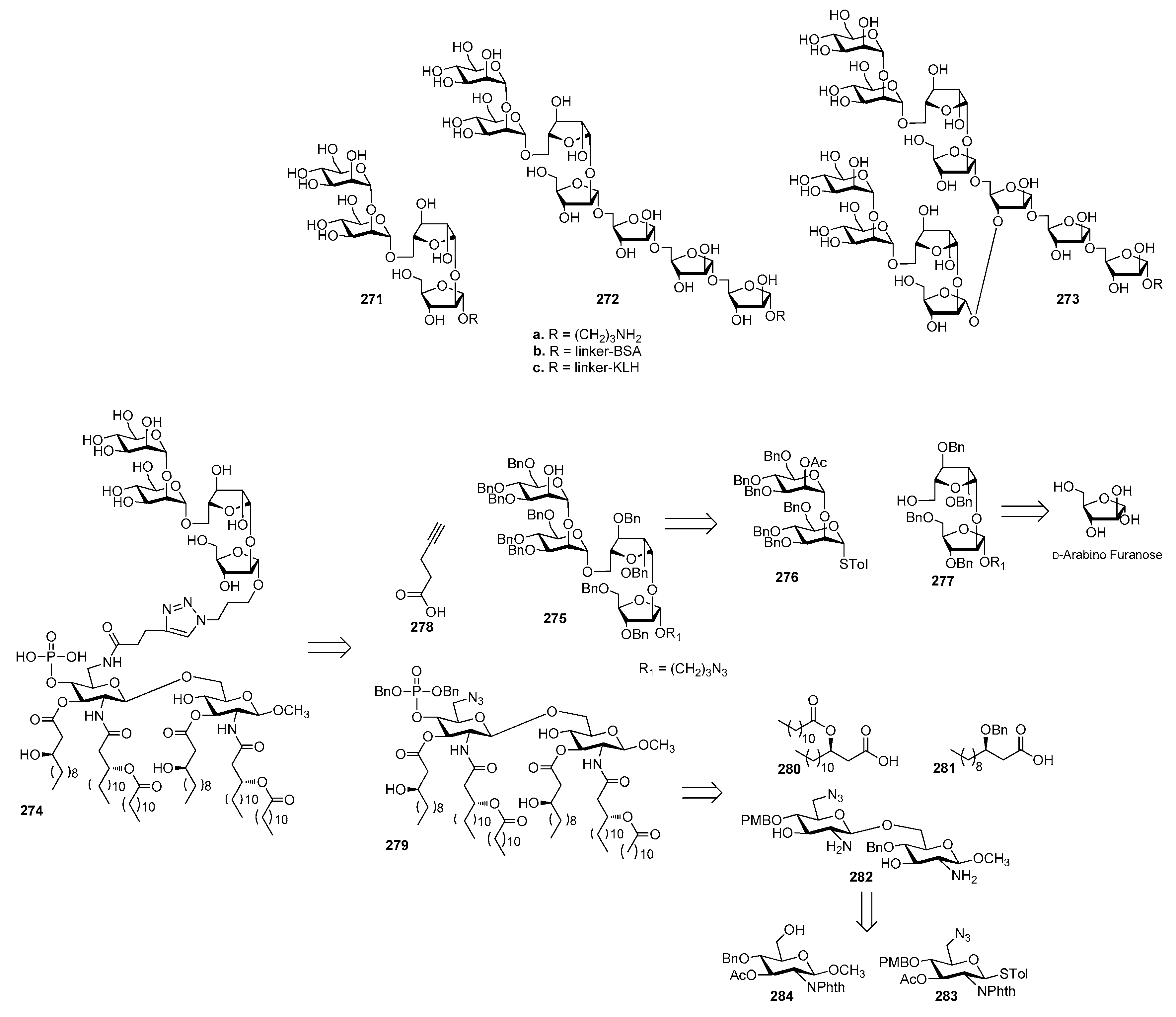{kind=link}
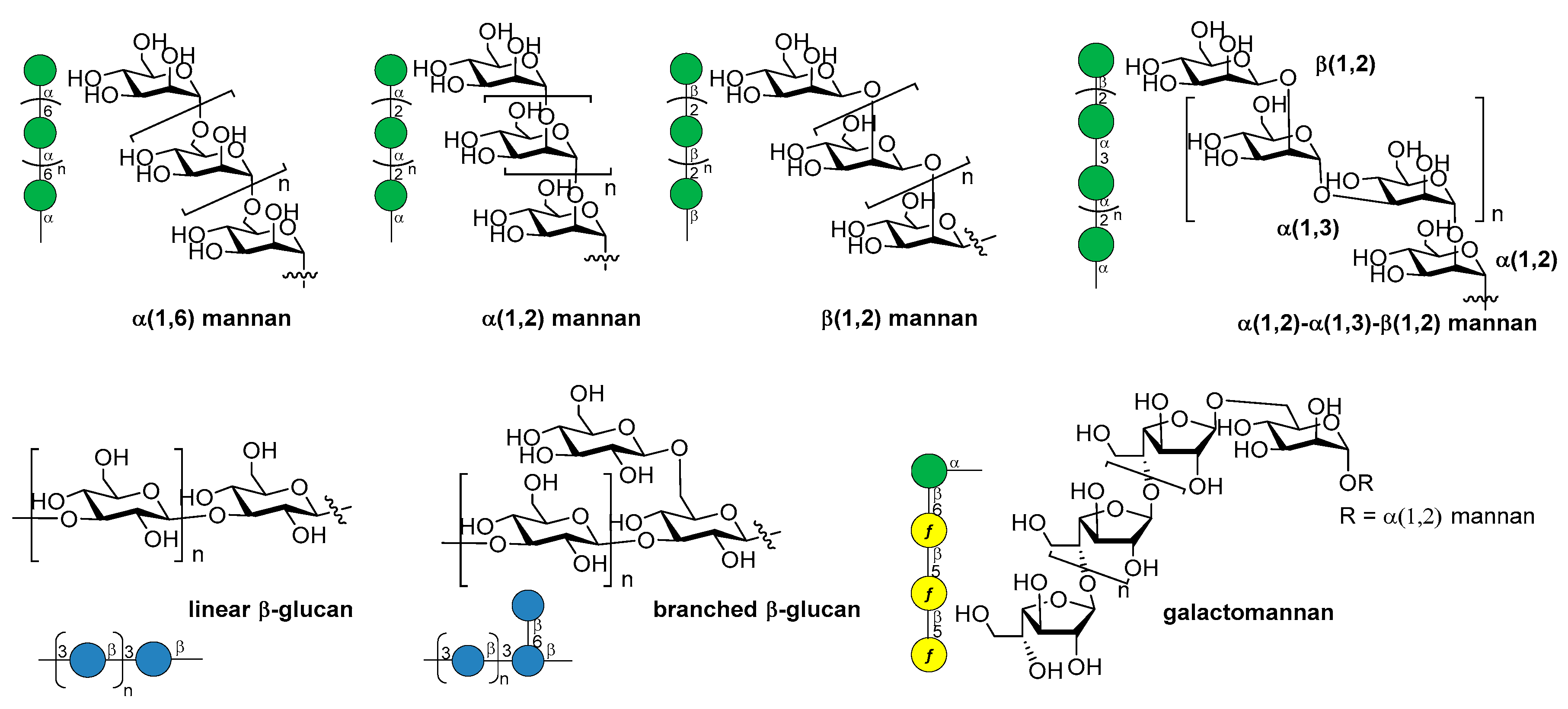{kind=link}
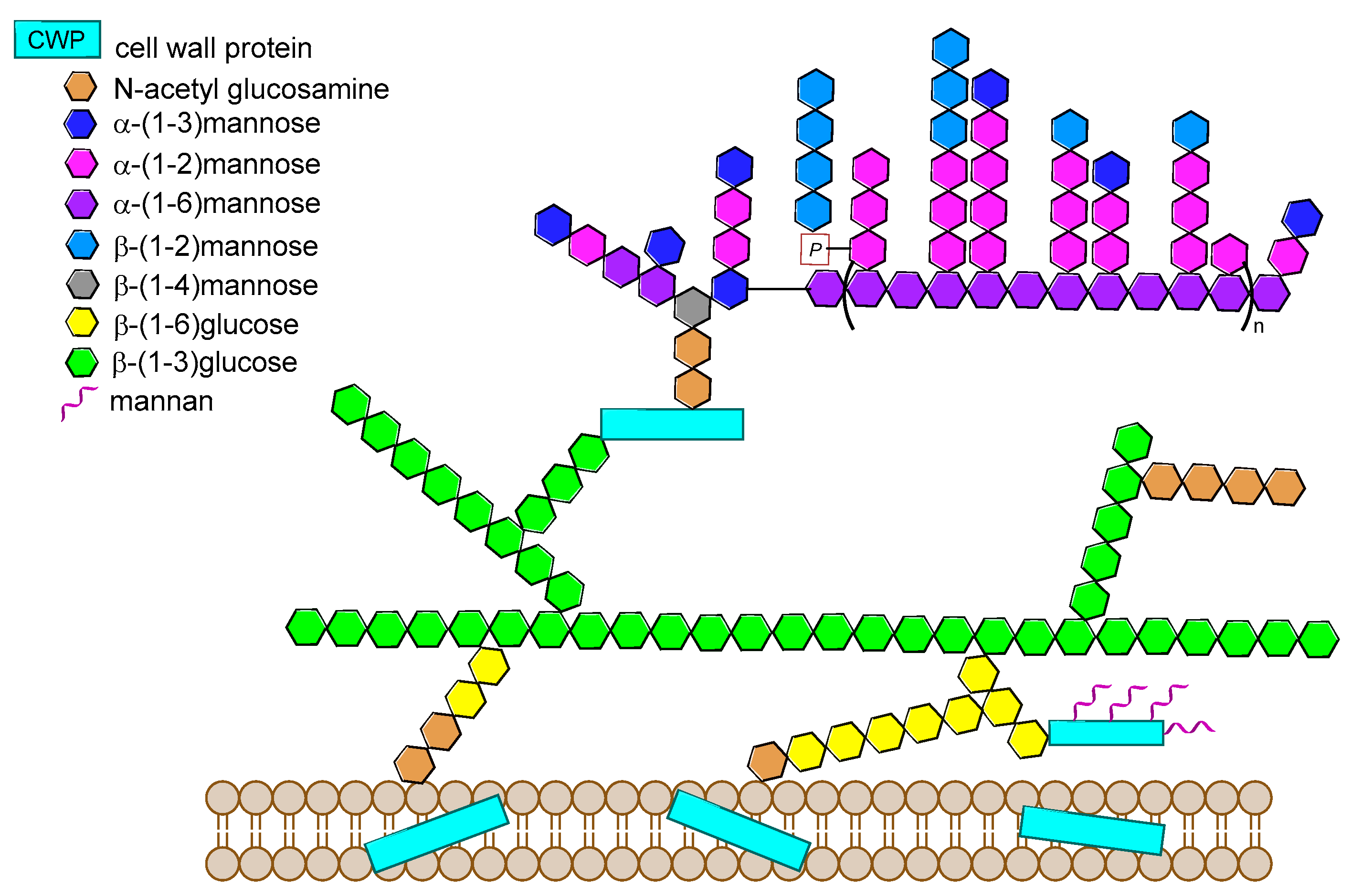{kind=link}
{kind=link}
{kind=link}
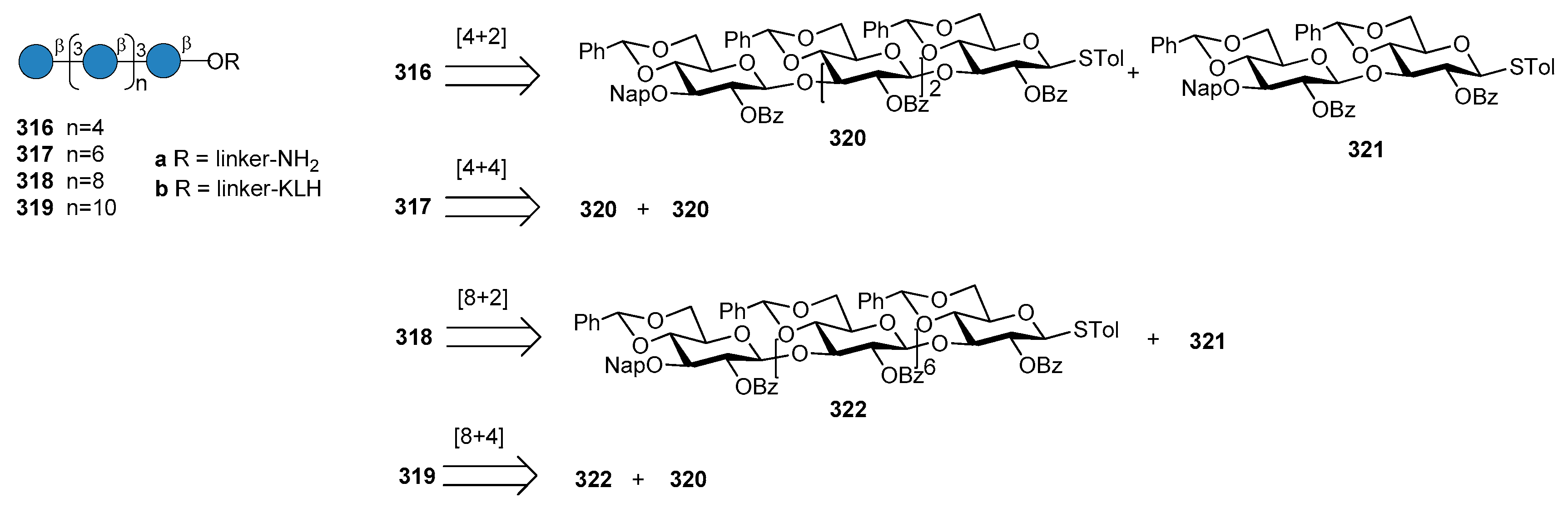{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Colombo, C.; Pitirollo, O.; Lay, L. Recent Advances in the Synthesis of Glycoconjugates for Vaccine Development. Molecules 2018, 23, 1712. https://doi.org/10.3390/molecules23071712
AMA Style
Colombo C, Pitirollo O, Lay L. Recent Advances in the Synthesis of Glycoconjugates for Vaccine Development. Molecules. 2018; 23(7):1712. https://doi.org/10.3390/molecules23071712
Chicago/Turabian StyleColombo, Cinzia, Olimpia Pitirollo, and Luigi Lay. 2018. "Recent Advances in the Synthesis of Glycoconjugates for Vaccine Development" Molecules 23, no. 7: 1712. https://doi.org/10.3390/molecules23071712