Isolation of β-1,3-Glucanase-Producing Microorganisms from Poria cocos Cultivation Soil via Molecular Biology

The Key Laboratory of Industrial Biotechnology, Ministry of Education, School of Biotechnology, Jiangnan University, 1800 Lihu Road, Wuxi 214122, China
*
Author to whom correspondence should be addressed.
Molecules 2018, 23(7), 1555; https://doi.org/10.3390/molecules23071555
Submission received: 17 May 2018
/
Revised: 25 June 2018
/
Accepted: 26 June 2018
/
Published: 27 June 2018
Abstract
:β-1,3-Glucanase is considered as a useful enzymatic tool for β-1,3-glucan degradation to produce (1→3)-linked β-glucan oligosaccharides with pharmacological activity properties. To validly isolate β-1,3-glucanase-producing microorganisms, the soil of Wolfiporia extensa, considered an environment rich in β-1,3-glucan-degrading microorganisms, was subjected to high throughput sequencing. The results demonstrated that the genera Streptomyces (1.90%) and Arthrobacter (0.78%) belonging to the order Actinomycetales (8.64%) in the phylum Actinobacteria (18.64%) were observed in soil for P. cocos cultivation (FTL1). Actinomycetes were considered as the candidates for isolation of glucan-degrading microorganisms. Out of 58 isolates, only 11 exhibited β-1,3-glucan-degrading activity. The isolate SYBCQL belonging to the genus Kitasatospora with β-1,3-glucan-degrading activity was found and reported for the first time and the isolate SYBC17 displayed the highest yield (1.02 U/mg) among the isolates. To check the β-1,3-glucanase contribution to β-1,3-glucan-degrading activity, two genes, 17-W and 17-Q, encoding β-1,3-glucanase in SYBC17 and one gene QLK1 in SYBCQL were cloned and expressed for verification at the molecular level. Our findings collectively showed that the isolates able to secrete β-1,3-glucanase could be obtained with the assistance of high-throughput sequencing and genes expression analysis. These methods provided technical support for isolating β-1,3-glucanase-producing microorganisms.
1. Introduction
The glycoside hydrolase β-1,3-glucanase, extensively distributed among plants, fungi, and bacteria, acts on 1,3-β-glucosidic bonds of structural β-1,3-glucans to hydrolyze or transfer glycosides [1,2]. Based on the hydrolysis position, β-1,3-glucanases are divided into endo-type (E.C. 3.2.1.39) and exo-type (E.C 3.2.1.58) categories. β-1,3-Glucans are the main cell wall component.β-1,3-Glucanase is able to hydrolyze these β-1,3-glucans, as a defense against fungal pathogens in plants, as well as to suppress fungal growth in fermentation technology [3,4]. Moreover, β-1,3-glucans can be degraded into (1→3)-linked β-d-glucan oligosaccharides with immunomodulating and antitumor properties by β-1,3-glucanase [5,6]. The exo-β-1,3-glucanase gene encodes an immune-dominant protein as a cell wall remodeler, which is a candidate for immunotherapy and can be used in diagnostic tests [7].
In bacteria, numerous recombinant and wild enzymes have been characterized from different sources, e.g., Streptomyces [8], Arthrobacter sp. [9,10], Cellulosimicrobium cellulans [11], Nocardiopsis sp. [12], Paenibacillus sp. [13], Thermotoga neapolitana [14], Bacillus circulan [15]. However, no reports have been made on β-1,3-glucanase selected from the genus Kitasatospora. β-1,3-Glucanase originating from bacteria are mainly classified into glycosyl hydrolase family 16, according to the putative amino acid sequences and secondary structures. The methods of isolating bacteria able to secrete β-1,3-glucanase were based on the lysing of cell walls of higher fungi or degrading polysaccharides [16,17]. Glucanase-producing microorganisms belonging to pathogenic bacteria were often selected in these way, and other isolates able to secrete β-1,3-glucanase were covered or their growth was inhibited. Although the methods of isolating strains with glucanase activity have been described, detailed data on the diversity of glucanase-producing microorganisms in any specific environment, which would be beneficial to screen the isolates able to secrete β-1,3-glucanase, are unavailable.
P. cocos, a fungus consisting of 90% β-glucan and 10% terpenes by dry weight, grows around the roots of pine trees. Linear 1,3-β-d-glucans with some glucose branches as the main component were extracted from P. cocos sclerotium [18]. Plant growth influences the diversity of rhizosphere microorganisms [19]. The soil for P. cocos cultivation is more likely to harbor diverse populations of bacteria with β-1,3-glucanase for polysaccharides hydrolysis.
Recent advancements have been made in high-throughput sequencing, which can provide sequencing depths to cover various bacterial ecosystems, e.g., soil, sediment, animal, insect gut, marine or freshwater [20,21,22,23,24]. The combination of high-throughput sequencing and culture-dependent techniques was conducted to characterize the microbial communities colonizing ancient Buddhist statues [25] as well as to construct a novel bioassay for N-acyl homoserine lactone-degrading microorganisms [26].
The aims of this study were to: (1) analyze the diversity and structure of microbial communities by using the method of high-throughput sequencing, (2) isolate and identify glucan-degrading actinomycetes, (3) determinate β-1,3-glucan-degrading activity, and (4) clone and express the genes encoding β-1,3-glucanase.
2. Results and Discussion
2.1. Bacterial Community Analysis
By the Miseq sequencing analysis of soil samples, 103,094 raw reads were generated. After trimming, denoising, and filtering chimeras, 102,005 effective sequences remained with the average length of 450 bp. Subsequently, the remaining sequences were clustered by 3% dissimilarity, and 18,446 bacterial operational taxonomic units (OTUs) were identified. The numbers of OTUs, Good’s coverage, Chao1 parameters, and Shannon indexes are collected in Table 1. The OTUs obtained from soil samples are displayed using rarefaction curves (Figure S1).
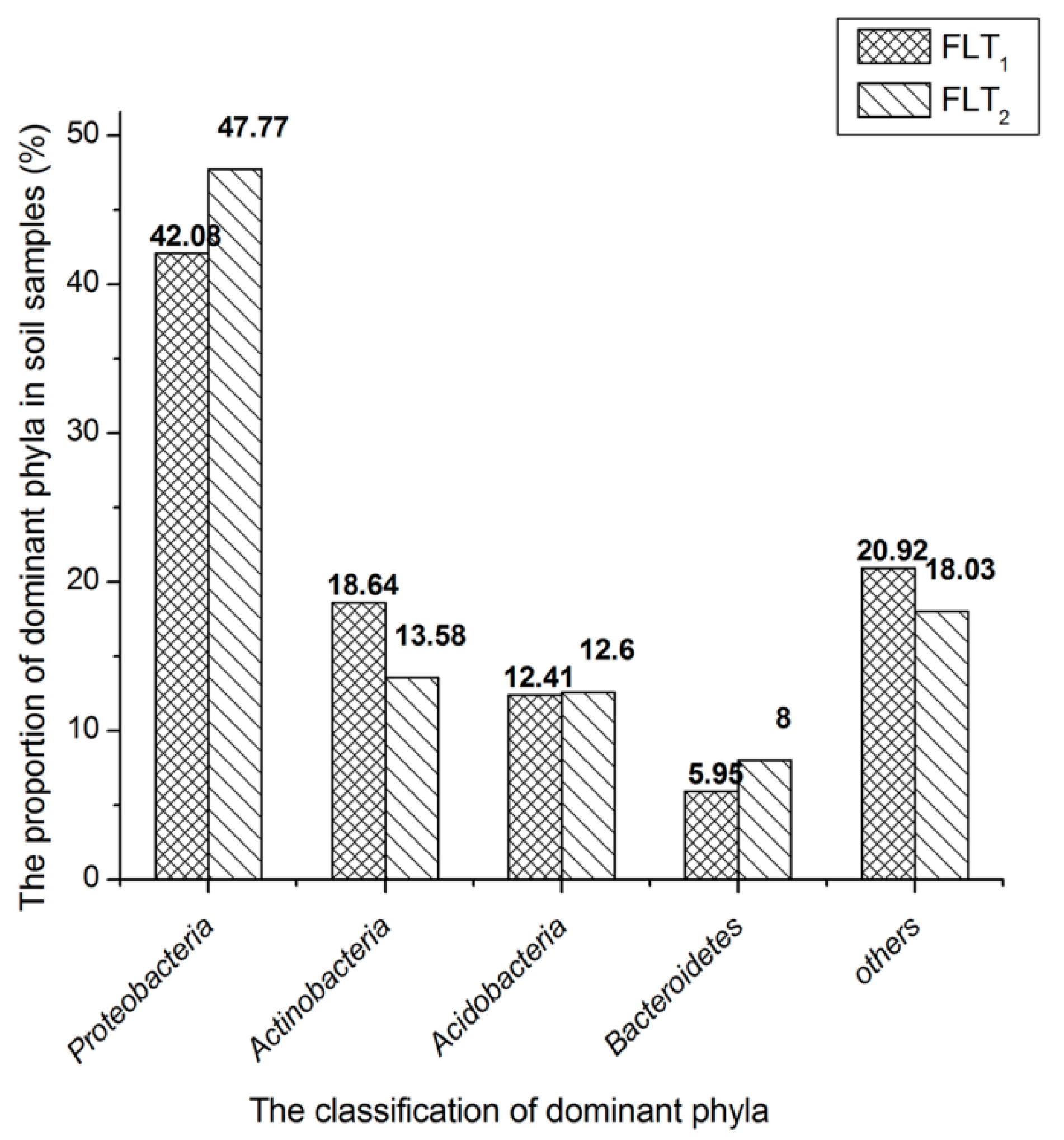
The classification results at the taxa phylum level are depicted in Figure 1. Proteobacteria (42.08%), Actinobacteria (18.64%), Acidobacteria (12.41%) and Bacteroidetes (5.95%) are considered as the dominant phyla as they contained over 5% of high-quality sequences in FLT1. The high-quality sequences not less than 1% were also classified into other subdominant phyla (Table S1). Eleven of abovementioned groups accounted for 96.13%. The remaining 16 phyla, in which effective sequences occurred at <1% abundance of the high-quality sequences, were defined as rare phyla in FLT1. Euryarchaeota and Thaumarchaeota belonging to the archaea domain accounted for 0.04% and 0.01% of the total high-quality sequences among FLT1, respectively (Table S1). The dominant phyla of FLT1 and FLT2 were compared. The composition of dominant phyla in FLT2 was similar to those of FLT1. The phylum Actinobacteria in FLT1 was over 5.06% of high-quality sequences those in FLT2. The abundances of other dominant phyla in FLT2 were greater than those in FLT1. Root exudates containing various primary and secondary plant metabolites have distinct influences on insect herbivores, nematodes, and microbes underground, in addition to deterring competing plants [27]. Thus, the microbial communities in FTL1 and FTL2 might illustrate the relationship of the abundances of dominant phyla with its use of the main metabolites produced by P. cocos. The abundance of Actinobacteria is more likely to relate with β-1,3-glucan produced by P. cocos. It is reasonable to consider that bacteria belonging to the phylum Actinobacteria probably have ability to secrete β-1,3-glucanase for β-1,3-glucan degradation. Certainly, it cannot be excluded that other dominant phyla can found microorganisms with β-1,3-glucanase activity.
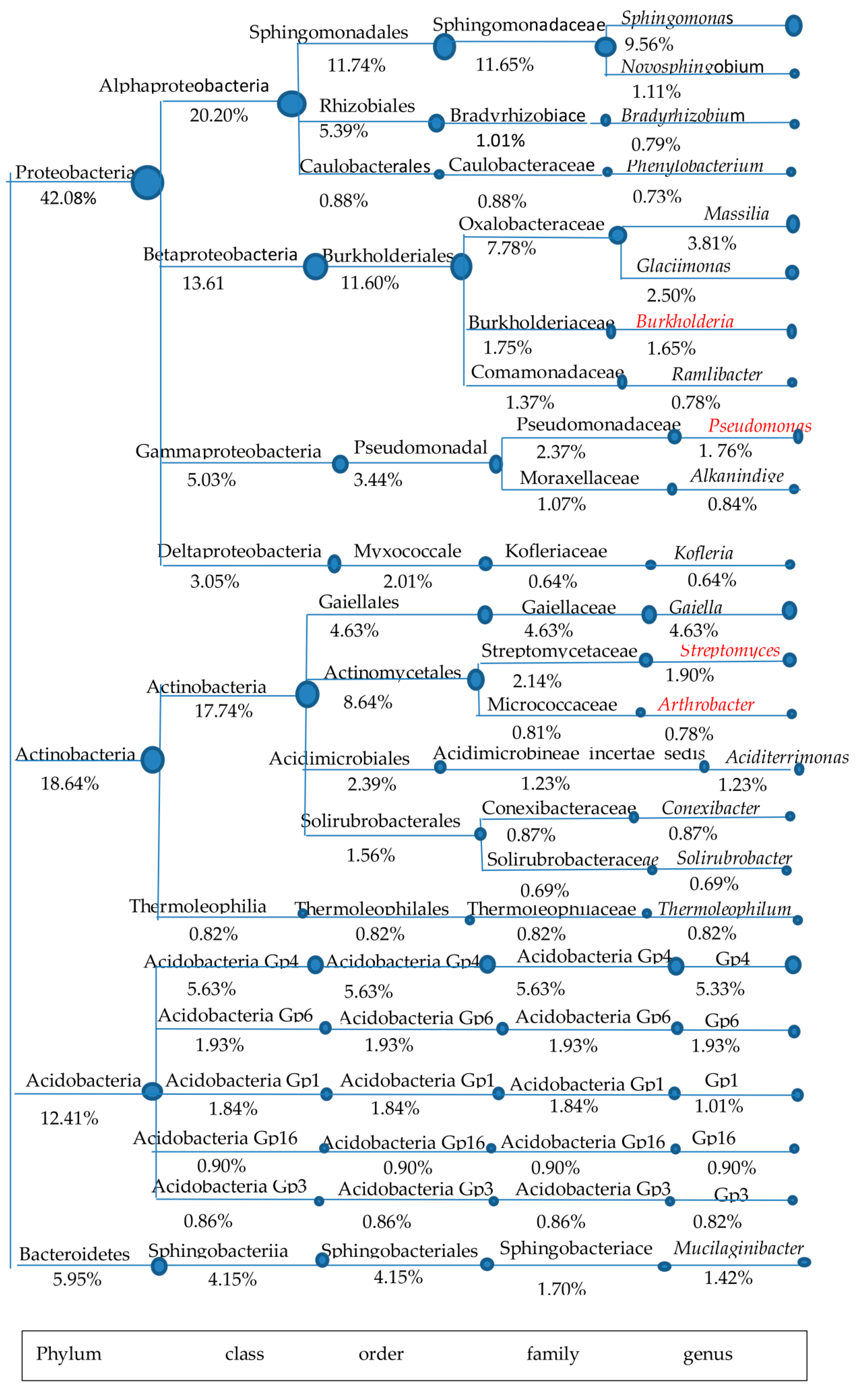
The soil for P. cocos cultivation (FLT1) was used as the candidate sample for isolating β-1,3-glucanase-producing microorganisms. To understand the diversity and structure of β-1,3-glucanase-producing microorganisms in FLT1, a combination of literature research and data analysis among dominant phyla was conducted in detail (Figure 2). Although Proteobacteria is the main dominant phylum that exists in FLT1, few reports refer to β-1,3-glucanase-producing microorganisms belonging to the phylum Proteobacteria from FLT1. Indeed, detailed data about microorganisms able to secrete glycoside hydrolases are available for this phylum, e.g., Sphingomonas [28,29], Sphingomonadaceae [30], Burkholderia [31], Pseudomonas [32].
Only the glycoside hydrolases selected from the genera Pseudomonas and Burkholderia that act on β-1,3-glucan have been exhaustively reviewed. The genus Burkholderia is worthless to isolate for its pathogenicity. Rare organisms able to degrade β-1,3-glucan were observed in the phyla Acidobacteria and Bacteroidetes from FLT1, except a characterized β-glucosidase from Mucilaginibacter sp. Strain QM49 [33]. The genera Arthrobacter and Streptomyces, accounting for 0.78% and 1.90% of the total high-quality sequences, were observed in the phylum Actinobacteria. It was noteworthy that both of the genera belong to the order Actinomycetales, which have been extensively reported as a source of β-1,3-glucanase [34,35,36]. FLT1 widely harbored the order Actinomycetales (8.64%) when it was compared with others especially the genus Pseudomonas (1.76%) belonging to the order Pseudomonadales (3.44%). Moreover, the remarkable presence of Actinobacteria was found in FLT1 but few that of microorganisms were observed in FTL2. A member of the order Actinomycetales is often called an actinomycete. It is well known that actinomycetes have unrivalled capacity to produce over two-thirds of natural antifungal metabolites [37]. Actinomycetes of the genus Streptomyces is well known as the largest genus of Actinobacteria, with properties of biological control. Overall, FLT1 probably harbored a number of actinomycetes able to secrete β-1,3-glucanase. Actinomycetes can be classified into probiotics. Thus, the soil for P. cocos cultivation was used as the candidate sample for isolating β-1,3-glucanase-producing actinomycetes.
2.2. Identification of Glucan-Degrading Microorganisms
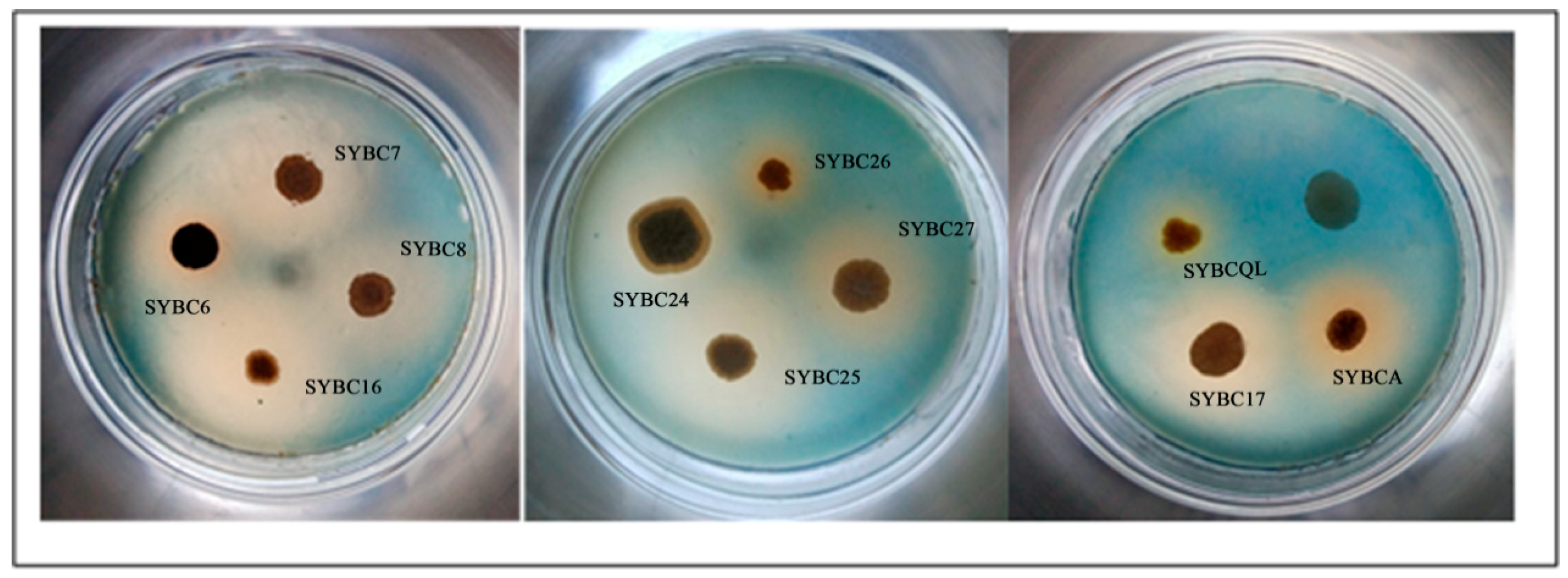
Colonies of actinomycetes were visible after the dilution of soil cultured on yeast casamino acids extract and dextrose agar (YCED). The isolates with β-1,3-glucan-degrading activity were screened among preferred actinomycetes. Out of 58 actinomycetes, only 11 among formed a clear halo around the colony after inoculation, indicating that they were able to degrade glucan (Figure 3). A positively relation can be found between the size of clear halo and the enzyme activity. The size of clear halo in SYBC26 and SYBCQL were obviously smaller than others. Thus both of them are weak to degrade β-1,3-glucan.
Ten isolates were identified to the genus Streptomyces, with the exception of SYBCQL belonging to the genus Kitasatospora, via 16S rRNA gene analysis. Each of the 16S rRNA gene sequences from isolates were aligned and submitted to the GenBank database, and all the sequences showed a high identity match (99%) to sequences obtained from the GenBank database (Table 2). Four isolates were closer to S. cellostaticus and S. capoamus. Two isolates showed high similarities to S. cinerochromogenes and S. coelescens. Three isolates were homologous to S. indiaensis, S. viridochromogenes, and K. phosalacinea, respectively, while another was homologous to S. olivogriseus and S. filipinensis. Phylogenetic analysis verified the taxonomic affiliations searched by BLAST alignment (Figure 4). The genus Streptomyces as a main member of actinomycetes was abundant in FLT1 based on high-throughput sequencing.
The results of identification were in agreement with the analysis of microbial communities at the genus level in FTL1. The genus Kitasatospora, is homologous to the genus Streptomyces, belonging to the order Actinomycetales among the phylum Actinobacteria [38]. Thus, SYBCQL able to degrade β-1,3-glucan was isolated from FLT1 under the same conditions.
2.3. Enzyme Activity Assay
The isolates that formed a clear halo around the colony (Figure 3) were determined to have the ability to degrade glucan in an exhausted culturing medium (Table 3). The isolates able to degrade β-1,3-glucan were feeble, as compared with that of Streptomyces rutgersensis [39] and Streptomyces torulosus PCPOK-0324 [16]. SYBC17 showed the highest yield of glucan-degrading activity (1.02 U/mg) among all isolates obtained from actinomycetes. Although the specific activity of SYBCQL was lower than that of others, the genus Kitasatospora with β-1,3-glucan-degrading activity was found and reported for the first time. In general, β-glucosidases participate in β-1,3-glucan degradation along with β-1,3-glucanases. Thus SYBCQL and SYBC17 with β-1,3-glucanases activity for β-1,3-glucan degradation were further verify at the molecular level.
2.4. Gene Clone and Analysis
One gene encoding β-1,3-glucanase was amplified from the genomic DNA of SYBCQL and named QLK1, encoding the deduced protein QLK1. Based on the genomic DNA of SYBC17, two β-1,3-glucanase genes were found and named 17-W, and 17-Q. Both correspond to the deduced proteins 17-W and 17-Q. PCR products were checked by 1% agarose gel electrophoresis and sequenced after TA cloning.
The residues 1–37 of QLK1 and the residues 1–30 of 17-W was identified as N-terminal signal peptides, according to SignaIP analysis. The mature protein QLK1 consisted of 391 residues with a deduced molecular mass of 40.4 kDa. The mature protein 17-W contained 389 residues and its deduced molecular mass was the same as QLK1. Meanwhile, 17-Q without an N-terminus leader sequence encoded a mature protein with a deduced molecular mass of 48.1 kDa (Figure 5). Each of the putative amino acid sequences has a catalytic domain similar to GH 16, based on align the protein sequences from GenBank database. QLK1 and 17-W, with a potential carbohydrate-binding domain (CBM), similarly belong to the regions of CBM 13 from Streptomyces at the C-terminus sequence. The functional domains of QLK1 and 17-W were found to be similar to β-1,3-glucanase from Streptomyces sp. S27 [35]. A glycine-rich region was observed between the functional domains of GH 16 and CBM 13 in QLK1 and 17-W. The region was also found in the linker structure in β-1,3-glucanase from Streptomyces sp. S27 [35] and S. sioyaensis [8]. The C-terminus domain of 17-Q was grouped into CBM family 6, found in several xylanases, rather than CBM family 13. The functional domains of 17-Q was similar to β-1,3-glucanase from S. sioyaensis [8].
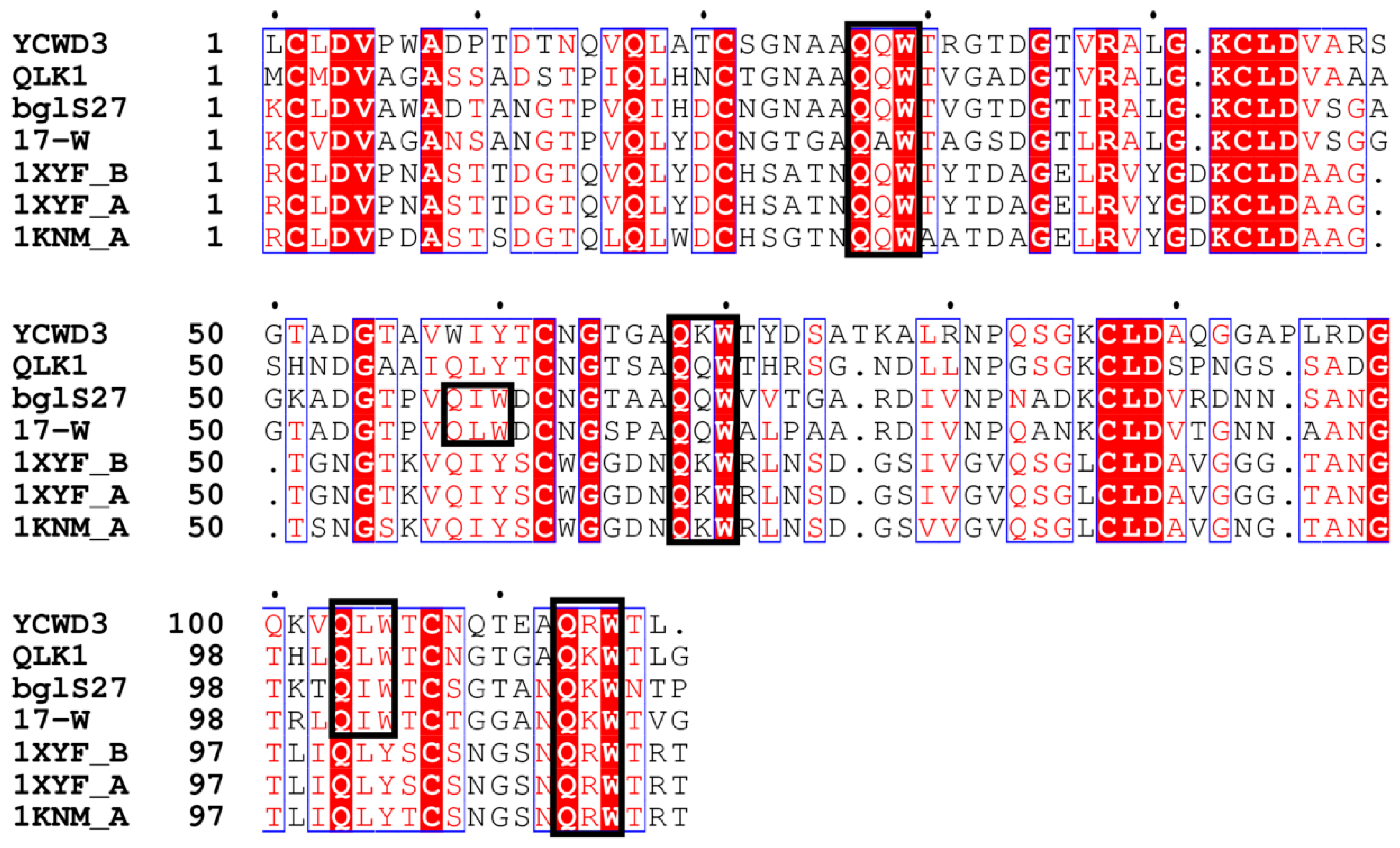
The CBMs of QLK1 and 17-W exhibited a structure like that of the ricin B-chain classified in CBM 13 members. The ricin B lectin domain is composed of three homologous regions as the QXW (Gln-X-Trp) repeats (Figure 6) [40]. There is a hypothesis that Gln works in substrate binding and Trp help to form the hydro phobic core [41]. The CBM of 17-Q is homogeneous to the CBMs belonging to family 6. Generally, members of CBM family 6 bind to xylan by connecting with the xylanase domain. For instance, the CBM of xylanase A from Clostridium stercorarium has been suggested to bind xylan and act as an important role in xylan hydrolysis [42]. Family 6 CBMs containing multiple distinct ligand binding sites present a unique ligand binding surface to recognize the non-reducing end of β-1,3-linked-glucans [43]. The CBM of S. sioyaensis β-1,3-glucanase is probably considered as an extra ordinary CBM classified into family 6, based on its binding preference, especially due to its unwilling binding to xylan (Figure 7) [8]. The ligand binding sites of 17-Q are similar to those found in the CBM of S. sioyaensis β-1,3-glucanase and probably have the same binding preference.
A highly consensus catalytic center for the hydrolysis of glycosidic bonds has been observed in GH family 16 [44,45]. A Met residue was observed in the catalytic motif of endo-β-1,3-glucanases but not in endo-β-1,3-1,4-glucanases [11]. As shown in Figure 5, a specific consensus motif with putative catalytic residues is found among these β-1,3-glucanases. BglF is completely inactive when the mutants of the deduced catalytic residues Glu123Gln and Glu128Gln are created [12]. Thus, the putative catalytic residues are crucial among these hydrolases. Besides, Glu128 protonates the glycosidic oxygen of the scissile bond by acting as a general acid [44].
The open reading frames (ORF) were aligned by using the online tool BLAST. QLK1 was 64% identical to the putative secreted hydrolase from S. coelicolor A3(2). Furthermore, QLK1 was homogeneous to β-1,3-glucanase from Nocardiopsis sp. strain F96 (63%) and Streptomyces sp. S27 (64%). 17-W was 84% identical to the putative secreted hydrolase from S. coelicolor A3(2) and showed identity with β-1,3-glucanase from Streptomyces sp. S27 (69%) and Nocardiopsis sp. strain F96 (53%). 17-Q showed identity with the putative secreted glucosidase from S. coelicolor A3(2) (80%) and endo-β-1,3-glucanase from S. sioyaensis (79%) and was 57% identical to Arthrobacter sp.NHB-10. The results indicated that QLK1 and 17-W were closer to the putative secreted hydrolase from S. coelicolor A3(2) and was significantly different from 17-Q. While 17-Q was similar to the putative secreted glucosidase from S. coelicolor A3(2) and endo-β-1,3-glucanase from S. sioyaensis (Figure 8 and Table 4). Interestingly, high homology was observed between the ORFS and those of never identified β-1,3-glucanase according to phylogenetic analysis.
2.5. Expression and Purification of Recombinant Enzymes
Recombinant enzymes were expressed with IPTG (isopropyl-β-d-1-thiogalacto-pyranoside) induction in E. coli BL21 (DE3) cells. Crude extracts in soluble form were purified using Ni+ affinity chromatography and desalting chromatography was used to remove the excess salt. SDS-PAGE analysis confirmed that QLK1, 17-W and 17-Q were overexpressed successfully, with high purity of the enzymes. The presence of molecular masses was close to the theoretical masses according to the deduced amino acid sequences of the enzymes (Figure 9). The purified recombinants QLK1, 17-W and 17-Q had specific activities of 65.82 U/mg, 132.90 U/mg, and 14.70 U/mg, respectively (Table 5). 17-Q showed the highest yield among all purified recombinant enzymes. All the recombinant enzymes displayed a several times higher level of β-1,3-glucanase activity than wild-type. These results suggested that SYBCQL and SYBC17 with β-1,3-glucanases activity were successfully confirmed in molecular level.
3. Materials and Methods
3.1. Materials
SYBCQL and SYBC17 were used as genetic DNA sources. E. coli DH5α and E. coli BL21 (DE3) were purchased from TaKaRa (Dalian, China) and used as hosts for genes cloning and expression. The plasmid pUC19 and pCold II vector were bought from TaKaRa and used for constructing recombinant plasmid. Luria-Bertani (LB) medium with 50 μg/mL ampicillin was used in recombinant plasmid amplification. The genomic DNA extraction kit, LA Taq DNA polymerase with GC buffer, PCR clean-up kit and other DNA-modifying enzymes were bought from TaKaRa. Laminarin with an average BR of 98% was purchased from Shanghai Yuanye Bio-Technology Company (Shanghai, China). The powder of fruiting bodies of the Basidiomycete P. cocos was provided by Johncan International Company (Hangzhou, China). High-throughput sequencing was performed by Shanghai Shenggong Company (Shanghai, China). Other chemicals were all of analytical grade and commercially available.
3.2. High-Throughput Sequencing
To find the candidate isolates in FLT1, for screening β-1,3-glucanase-producing microorganisms, the microbial communities in the soil for P. cocos cultivation (FLT1) and bulk soil (FLT2) were investigated and compared.
Soil sampling was carried out in March 2017. The sphagnum and duff layers of the sampling area were removed, and P. cocos were found around the roots of pine trees using a soil knife. FLT1 was collected from the soil around P. cocos growing in a township (28°35′ N, 185°95′ E), Liu’an City, Anhui Province, China. The bulk soil sample (FLT2) was gathered approximately 2 m away from FLT1 and just under the root zone of any grasses growing on the surface (pH 7.0). The soil of the 10 cm depth layer was collected using an auger with a diameter. To remove stones and roots, both of the soil samples were timely sieved (2 mm mesh) in the field. The treated samples were then kept under a low temperature maintained by ice until molecular analysis.
High-throughput sequencing in molecular analysis and data processing were conducted as described previously [46]. After DNA extraction, PCR amplification, and pyrosequencing, the MiSeq-generated raw sequences were submitted to the DDBJ database (accession number: DRA006753). The raw MiSeq-generated sequences were further processed using the soft-ware Prinseq (PRINSEQ-lite 0.19.5) [47] and the software package of Mothur1.30 with “pre.cluster” command [48]. The available sequences were clustered into operational taxonomic units (OTUs) and the thresthod value of sequences similarity was set at 0.97. Based on the results of OTU clustering, the most abundant sequence as the representative sequence of OTU was acquired and subjected to various types of analysis.
Taxonomic assignment was accomplished by the Ribosomal Database Project (RDP) Classifier according to Bergey’s taxonomy [49]. A bootstrap cutoff of 80% was used to assign the obtained sequences to each taxonomy levels. The evolutionary relationships and abundance of the dominant phyla in FLT1 at the genus level were visualized using the ete3 (Environment for Tree Exploration) package in python.
3.3. Isolation and Identification of Glucan-Degrading Microorganisms
Actinomycetes were screened from FLT1 by serial dilution and spread-plate techniques [50]. FLT1 (5 g) was mixed with 100 mL sterile distilled water and diluted to 10−5. One hundred microliter of the different dilutions were grown on the plates containing 0.03% yeast extract, 0.03% casamino acid, 0.03% d-glucose, 0.05% K2HPO4, and 1.8% agar (w/v) in triplicates, respectively. Cyclohexamide (100 μg/mL) was added to resist fungal contamination after autoclaving. The plates were incubated at 28 °C for 1–2 weeks. Typical actinomycetes colonies were picked out according to morphological characteristics as well as microscopic examination. The morphologically distinct colonies were then purified on the original media at 28 °C for one week, stored at 4 °C. Furthermore, glucan-degrading actinomycetes were inoculated on agar plates containing the powder of fruiting bodies of Basidiomycete P. cocos (0.5%, w/v) and aniline blue (0.005%, w/v), and then formed a clear halo around the colony [51].
To identify the unknown isolates, the selected isolates were incubated in 3 mL ISP-2 medium under rotary shaking at 30 °C for 48 h. A volume of 1.5 mL culture was centrifuged at 8000× g for 1 min. The pellet was then washed once with distilled water and used to extract genomic DNA. The genomes of each isolate were extracted following by the operating instruction of genomic DNA extraction kit (www.tiandz.com). 16S rRNA gene identification of glucanase-producing bacteria were amplified by using the universal primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′) and sent to Shanghai Shengong (Shanghai, China) for sequencing. A homology search of the closest phylogenetic neighbors was conducted using the online tool BLAST.
3.4. Determination of the Enzyme Activity
The isolates were inoculated aerobically with rotary shaking at 200 rpm and 30 °C for 72 h in a medium (per liter) containing laminarin 5 g, K2HPO4 1 g, NaNO3 3 g, KCl 0.5 g, MgSO4·7H2O 0.5 g, and FeSO4·7H2O 0.5 g. The cultures were centrifuged at 10,000× g at 4 °C and the culture filtrates were harvested for activity assay. The standard activity assay for β-1,3-glucan degradation was obtained by measuring the formation of reducing sugar using a colorimetric method [16]. Culture filtrates of the strains (500 μL) were mixed with 500 μL of 0.5% (w/v) laminarin in 100 mM sodium acetate buffer (pH 5.5). The reaction was conducted at 50 °C for 60 min and terminated by heating for 5 min at 100 °C. Then 2 mL of 1% dinitrosalicylate (DNS) was added into the reaction solution and the mixture was boiled for 10 min. The mixture was placed in an ice bath and then measured at 540 nm using the spectrophotometer. According to the standard assay conditions, one unit (U) of the activity was defined as the amount of enzymes that can liberate 1 μmol of glucose in one minute. All experiments were set to repeat, with triplicates of each treatment. Protein concentrations were measured by the method of Bradford [52] using bovine serum albumin as a standard.
3.5. Cloning and Expression of β-1,3-Glucanase Genes
A genome analysis from the NCBI database was conducted, with the genomic DNA of Kitasatospora setae KM-6054 and Streptomyces griseochromogenes ATCC 14511 employed as the templates and synthetic primers (Table 6). The genome of SYBCQL and SYBC17 has been extracted by genomic DNA extraction kit. The plasmid pCold II DNA and the vector pUC19 DNA was isolated using a plasmid miniprep kit. Then the coding sequences were amplified by the polymerase chain reaction (PCR) using LA Taq DNA polymerase with GC buffer, and sequenced after TA cloning. The nucleotide sequences were deposited in the GenBank database (accession number: 17-W, MH190407; 17-Q, MH190408; QLK1, MH190409.). Subsequently PCR products were ligated with the pcold II vector after both were digested with Hind III and Xba I. The recombinant plasmid was then transformed into E. coli BL21 (DE3) competent cells.
Transformants containing the recombinant enzymes were picked from the single colony and inoculated overnight at 37 °C in ampicillin-supplemented LB. Moreover, the overnight cultured transformants (1 mL) were transferred into 50 mL of fresh LB medium with the addition of 100 μg/mL ampicillin and grown at 37 °C to a cell density of 0.6~0.8. To induce the expression of the recombinant enzymes, IPTG was then added to a final concentration of 0.4 mM and the cultivation continued for 24 h at 15 °C. Cells were harvested by centrifugation at 4 °C and 8000× g for 10 min, and resuspended in 100 mM sodium acetate buffer at pH 5.5. Cells were lysed by sonication for 15 min on ice, and cell supernatants was collected by centrifugation (8000× g, 10 min at 4 °C) for further purification.
3.6. Purification of the β-1,3-Glucanases
To purify the recombinant proteins with six histidine residues, an AKTA Avant system at 6 °C was used (GE Healthcare, Uppsala, Sweden), followed by desalting with a HisTrap TM column (GE Healthcare, Uppsala, Sweden). The cell supernatant (crude enzyme) was applied to a HisTrap HP column (GE Healthcare) equilibrated with binding buffer A (100 mM sodium acetate, 5 mM imidazole, 500 mM NaCl, pH 5.5), and was eluted with buffer B (100 mM sodium acetate, 500 mmol/L imidazole, and 500 mmol/L NaCl, pH 5.5) using an imidazole step gradient of 0% to 100% buffer B. The collected fractions with β-1,3-glucanase activity were assayed using the former described method. The purified proteins were loaded on 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The concentration of purified proteins was tested by the method described above.
3.7. Bioinformatics Analysis
In the NCBI database (https://www.ncbi.nlm.nih.gov/genome), a homology analysis was performed using the online tool BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi). DNA and protein sequence alignments were performed using the blastn and blastp programs in the NCBI database, respectively. The N-terminus signal peptide was forecasted by the SignaIP 3.0 server (http://www.cbs.dtu.dk/ services/SignaIP/). The multiple sequence alignment was performed based on ClustalW program (http://www.ebi.ac.uk/clustalW/) and embellished by ESPript (http://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi). Phylogenetic analysis was conducted by Molecular Evolutionary Genetics Analysis (MEGA) 5.0 with neighbor-joining method at a bootstrap of 1000.
4. Conclusions
In the present paper, actinomycetes with the ability to degrade β-1,3-glucan were isolated using high-throughput sequencing combined with culture-dependent techniques. Both strains SYBCQL and SYBC17 able to secrete β-1,3-glucanase for β-1,3-glucan degradation were verified at the molecular level. It was suggested that these methods could be applied to effectively isolate β-1,3-glucanase-producing microorganisms, which is useful for the screening of other metabolite-producing microorganisms from specific environment.
Supplementary Materials
The following are available online. Figure S1: Rarefaction curves based on the OTUs at the cutoff of 97% 16S rRNA sequence similarity, Table S1: Relative abundances (% of total good-quality sequences) of all phyla in each soil sample. The dominant phyla are marked in shade (>1% of good quality sequences in at least one sample), and the total abundances in each soil sample are displayed at the bottom of the latter two lines. Table S2: The abundance of taxa genus levels in soil samples. The taxa represented within the top 30 abundances at the taxa genus levels and beyond the top 30 abundances at the taxa genus levels are classified into other levels.
Author Contributions
Q.W. (Qiulan Wu) performed most of the experiments and wrote the paper. Z.G. and Y.C. provided intellectual input to the study design. X.D. and Q.W. (Qi Wang) revised the manuscript. X.L. provided all financial support for this research project.
Funding
This research was funded by the Collaborative Innovation Involving Production, Teaching and Research Funds of Jiangsu Province (BY2014023-28) and the Agricultural Support Project, Wuxi Science and Technology Development (CLE01N1310).
Acknowledgments
The work was financially supported by the Collaborative Innovation Involving Production, Teaching and Research Funds of Jiangsu Province (BY2014023-28) and the Agricultural Support Project, Wuxi Science and Technology Development (CLE01N1310).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kobayashi, T.; Uchimura, K.; Kubota, T.; Nunoura, T.; Deguchi, S. Biochemical and genetic characterization of β-1,3 glucanase from a deep subseafloor Laceyella putida. Appl. Microbiol. Biotechnol. 2016, 100, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Kusaykin, M.I.; Belik, A.A.; Kovalchuk, S.N.; Dmitrenok, P.S.; Rasskazov, V.A.; Isakov, V.V.; Zvyagintseva, T.N. A new recombinant endo-1,3-β-d-glucanase from the marine bacterium Formosa algae KMM 3553: enzyme characteristics and transglycosylation products analysis. World J. Microbiol. Biotechnol. 2017, 33, 40. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, S.B.; Gupta, A. Studies on application of β-1,3 glucanase in the degradation of glucans produced by Botrytis cinerea and inhibition of fungal growth. Biocatal. Agric. Biotechnol. 2016, 7, 45–47. [Google Scholar] [CrossRef]
- Lee, S.Y. Biocontrol of anthracnose in pepper using chitinase, β-1,3 glucanase, and 2-furancarboxaldehyde produced by Streptomyces cavourensis SY224. J. Microbiol. Biotechnol. 2012, 22, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Cheng, L.; Meng, Y.; Li, S.; Zhao, X.; Du, Y.; Yin, H. Cellulosimicrobium cellulans strain E4-5 enzymatic hydrolysis of curdlan for production of (1→3)-linked β-d-glucan oligosaccharides. Carbohydr. Polym. 2015, 134, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Hida, T.H.; Ishibashi, K.; Miura, N.N.; Adachi, Y.; Shirasu, Y.; Ohno, N. Cytokine induction by a linear 1,3-glucan, curdlan-oligo, in mouse leukocytes in vitro. Inflamm. Res. 2009, 58, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, T.C.; Weiblen, C.; de Azevedo, M.I.; de Avila Botton, S.; Robe, L.J.; Pereira, D.I.; Monteiro, D.U.; Lorensetti, D.M.; Santurio, J.M. Microevolutionary analyses of Pythium insidiosum isolates of Brazil and Thailand based on exo-1,3-β-glucanase gene. Infect. Genet. Evol. 2017, 48, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.Y.; Cheng, C.W.; Huang, J.W.; Meng, M. Isolation and biochemical characterization of an endo-1,3-β-glucanase from Streptomyces sioyaensis containing a C-terminal family 6 carbohydrate-binding module that binds to β-1,3-glucan. Microbiology 2002, 148, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Kang, Y.-N.; Ban, M.; Oda, M.; Kobayashi, R.; Ohnishi, M.; Mikami, B. Crystallization and preliminary crystallographic analysis of endo-1,3-beta-glucanase from Arthrobacter sp. Acta Cryst. 2005, 61, 68–70. [Google Scholar]
- Okazaki, K.; Nishimura, N.; Matsuoka, F.; Hayakawa, S. Cloning and characterization of the gene encoding endo-β-1,3-glucanase from Arthrobacter sp. NHB-10. Biosci. Biotechnol. Biochem. 2007, 71, 1568–1571. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, Y.; Oda, M. Molecular characterization of endo-1,3-β-glucanase from Cellulosimicrobium cellulans: effects of carbohydrate-binding module on enzymatic function and stability. BBA 2011, 1814, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Masuda, S.; Endo, K.; Koizumi, N.; Hayami, T.; Fukazawa, T.; Yatsunami, R.; Fukui, T.; Nakamura, S. Molecular identification of a novel β-1,3-glucanase from alkaliphilic Nocardiopsis sp. strain F96. Extremophiles 2006, 10, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.Y.; Meng, M. Biochemical characterization and antifungal activity of an endo-1,3-β-glucanase of Paenibacillus sp. isolated from garden soil. Appl. Microbiol. Biotechnol. 2003, 61, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Zverlov, V.V.; Volkov, I.Y.; Velikodvorskaya, T.V.; Schwarz, W.H. Highly thermostable endo-1,3-β-glucanase (laminarinase) LamA from Thermotoga neapolitana: Nucleotide sequence of the gene and characterization of the recombinant gene product. Microbiology 1997, 143, 1701–1708. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Taki, J.; Yamamoto, M.; Aono, R. Cloning and structural analysis of bglm gene coding for the fungal cell wall-lytic β-1,3-glucan-hydrolase bglm of Bacillus circulans IAM1165. Biosci. Biotechnol. Biochem. 2014, 66, 1246–1255. [Google Scholar] [CrossRef]
- Park, J.K.; Kim, J.D.; Park, Y.I.; Kim, S.K. Purification and characterization of a 1,3-β-d-glucanase from Streptomyces torulosus PCPOK-0324. Carbohydr. Polym. 2012, 87, 1641–1648. [Google Scholar] [CrossRef]
- Mallikharjuna Rao, K.L.N.; Siva Raju, K.; Ravisankar, H. Cultural conditions on the production of extracellular enzymes by Trichoderma isolates from tobacco rhizosphere. Braz. J. Microbiol. 2016, 47, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, M.; Ruan, D.; Shashkov, A.S.; Kilcoyne, M.; Savage, A.V.; Zhang, L. Chemical components and molecular mass of six polysaccharides isolated from the sclerotium of Poria cocos. Carbohydr. Res. 2004, 339, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Marschner, P. Plant-Microbe Interactions in the Rhizosphere and nutrient cycling. In Soil Biology; Marschner, P., Rengel, Z., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 10, pp. 60–182. ISBN 978-3-540-68027-7. [Google Scholar]
- Tourna, M.; Freitag, T.E.; Nicol, G.W.; Prosser, J.I. Growth, activity and temperature responses of ammonia-oxidizing archaea and bacteria in soil microcosms. Environ. Microbiol. 2008, 10, 1357–1364. [Google Scholar] [CrossRef] [PubMed]
- Quaiser, A.; Zivanovic, Y.; Moreira, D.; Lopez-Garcia, P. Comparative metagenomics of bathypelagic plankton and bottom sediment from the Sea of Marmara. ISME J. 2011, 5, 285–304. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Hong, I.P.; Bok, J.I.; Kim, B.Y.; Song, J.; Weon, H.Y. Pyrosequencing analysis of the bacterial communities in the guts of honey bees Apis cerana and Apis mellifera in Korea. J. Microbiol. 2012, 50, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Cydzik-Kwiatkowska, A.; Zielinska, M. Bacterial communities in full-scale wastewater treatment systems. World J. Microbiol. Biotechnol. 2016, 32, 66. [Google Scholar] [CrossRef] [PubMed]
- Highlander, S.K. High throughput sequencing methods for microbiome profiling: Application to food animal systems. Anim. Health Res. Rev. 2012, 13, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, B.; Wang, L.; Ge, Q. Distribution and diversity of bacteria and fungi colonizing ancient buddhist statues analyzed by high-throughput sequencing. Int. Biodeter. Biodegr. 2017, 117, 245–254. [Google Scholar] [CrossRef]
- Liu, P.; Gao, Y.; Huang, W.; Shao, Z.; Shi, J.; Liu, Z. A novel bioassay for high-throughput screening microorganisms with N-acyl homoserine lactone degrading activity. Appl. Biochem. Biotechnol. 2012, 167, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, N.M.; Bouwmeester, H.J. Metabolomics in the rhizosphere: Tapping into belowground chemical communication. Trends Plant Sci. 2016, 21, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, W.; Ochiai, A.; Momma, K.; Itoh, T.; Mikami, B.; Maruyama, Y.; Murata, K. Crystal structure of the glycosidase family 73 peptidoglycan hydrolase FlgJ. Biochem. Biophys. Res. Commun. 2009, 381, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhou, J.; Gao, Y.; Guan, Y.; Li, J.; Tang, X.; Xu, B.; Ding, J.; Huang, Z. Molecular and biochemical characterizations of a new low-temperature active mannanase. Folia Microbiol. 2015, 60, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Aylward, F.O.; McDonald, B.R.; Adams, S.M.; Valenzuela, A.; Schmidt, R.A.; Goodwin, L.A.; Woyke, T.; Currie, C.R.; Suen, G.; Poulsen, M. Comparison of 26 sphingomonad genomes reveals diverse environmental adaptations and biodegradative capabilities. Appl. Environ. Microbiol. 2013, 79, 3724–3733. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.H.; Ye, J.R.; Liu, H.; Xu, X.L.; Wu, X.Q. Isolation and characterization of a new Burkholderia pyrrocinia strain JK-SH007 as a potential biocontrol agent. World J. Microbiol. Biotechnol. 2011, 27, 2203–2215. [Google Scholar] [CrossRef]
- Kitamura, E.; Kamei, Y. Molecular cloning of the gene encoding beta-1,3(4)-glucanase A from a marine bacterium, Pseudomonas sp. PE2, an essential enzyme for the degradation of Pythium porphyrae cell walls. Appl. Microbiol. Biotechnol. 2006, 71, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.H.; Liu, Q.M.; Kim, J.K.; Sung, B.H.; Kim, S.G.; Kim, S.C.; Im, W.T. Identification and characterization of a mucilaginibacter sp. strainqm49 β-glucosidase and its use in the production of the pharmaceutically active minor ginsenosides (s)-rh1 and (s)-rg2. Appl. Environ. Microbiol. 2013, 79, 5788–5798. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.M.; Liu, S.W.; Hsu, M.T.; Hung, C.L.; Lai, C.C.; Cheng, W.C.; Wang, H.J.; Li, Y.K.; Wang, W.C. Structure, mechanistic action, and essential residues of a GH-64 enzyme, laminaripentaose-producing beta-1,3-glucanase. J. Biol. Chem. 2009, 284, 26708–26715. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Yao, G.; Yang, P.; Li, N.; Luo, H.; Bai, Y.; Wang, Y.; Yao, B. Cloning, characterization, and antifungal activity of an endo-1,3-β-d-glucanase from Streptomyces sp. S27. Appl. Microbiol. Biotechnol. 2010, 85, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, P. Revisiting the Cellulosimicrobium cellulans yeast-lytic beta-1,3-glucanases toolbox: A review. Microb. Cell Fact. 2006, 5, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doumbou, C.L.; Hamby Salove, M.K.; Crawford, D.L.; Beaulieu, C. Actinomycetes, promising tools to control plant diseases and to promote plant growth. Phytoprotection 2001, 82, 85. [Google Scholar] [CrossRef] [Green Version]
- Girard, G.; Traag, B.A.; Sangal, V.; Mascini, N.; Hoskisson, P.A.; Goodfellow, M.; van Wezel, G.P. A novel taxonomic marker that discriminates between morphologically complex actinomycetes. Open Biol. 2013, 3, 130073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javmen, A.; Grigiskis, S.; Rudenkov, M.; Mauricas, M. Purification and partial characterization of a novel β-1,3-endoglucanase from Streptomyces rutgersensis. Protein J. 2013, 32, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Hazes, B. The (QxW)3 domain: A flexible lectin scaffold. Protein Sci. 1996, 5, 1490–1501. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, Z.; Kuno, A.; Kaneko, S.; Yoshida, S.; Kobayashi, H.; Kusakabe, I.; Mizuno, H. Crystal structure of streptomyces olivaceoviridis e-86 β-xylanase containing xylan-binding domain. J. Mol. Biol. 2000, 300, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.L.; Sakka, K.; Karita, H.; Kimura, T.; Ohmiya, K. Adsorption of clostridium stercorarium xylanase a to insoluble xylan and the importance of the cbds to xylan hydrolysis. J. Ferment. Bioeng. 1998, 85, 63–68. [Google Scholar] [CrossRef]
- Van Bueren, A.L.; Morland, C.; Gilbert, H.J.; Boraston, A.B. Family 6 carbohydrate binding modules recognize the non-reducing end of β-1,3-linked glucans by presenting a unique ligand binding surface. J. Biol. Chem. 2005, 280, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.Y.; Hsiao, Y.Y.; Meng, M.; Li, T.T. The 1.5 A structure of endo-1,3-β-glucanase from Streptomyces sioyaensis: evolution of the active-site structure for β-1,3-glucan-binding specificity and hydrolysis. Acta Crystallogr. C 2008, 64, 964–970. [Google Scholar]
- Juncosa, M.; Pons, J.; Dot, T.; Querol, E.; Planas, A. Identification of active site carboxylic residues in bacizzus zicheniformis 1,3-1,4-β-d-glucan-4-glucanohydrolase by site-directed mutagenesis. J. Biol. Chem. 1994, 269, 14530–14535. [Google Scholar] [PubMed]
- Zhao, D.; Huang, R.; Zeng, J.; Yu, Z.; Liu, P.; Cheng, S.; Wu, Q.L. Pyrosequencing analysis of bacterial community and assembly in activated sludge samples from different geographic regions in China. Appl. Microbiol. Biotechnol. 2014, 98, 9119–9128. [Google Scholar] [CrossRef] [PubMed]
- Kunin, V.; Engelbrektson, A.; Ochman, H.; Hugenholtz, P. Wrinkles in the rare biosphere: pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ. Microbiol. 2010, 12, 118–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huse, S.M.; Welch, D.M.; Morrison, H.G.; Sogin, M.L. Ironing out the wrinkles in the rare biosphere through improved OTU clustering. Environ. Microbiol. 2010, 12, 1889–15898. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.T.; Davies, F.L. Use of antibiotics for selective isolation and enumeration of actinomycetes in soil. J. Genet. Microbiol. 1965, 38, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Mahasneh, A.M.; Stewart, D.J. A Medium for Detecting β-(1,3) Glucanase Activity in Bacteria. J. Appl. Microbiol. 1980, 48, 457–458. [Google Scholar]
- Bradford, M.M. Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |
Figure 1.
Phylum-level microbial communities in soil samples. The taxa represented account for >5% abundance in at least one sample. Other phyla represent the taxa with their maximum abundance of <5% in any sample.
Figure 1.
Phylum-level microbial communities in soil samples. The taxa represented account for >5% abundance in at least one sample. Other phyla represent the taxa with their maximum abundance of <5% in any sample.

Figure 2.
A tree was composed of classification systems within dominant phyla at the genus level in FTL1. The abundances of taxa genera are over the top 30. The genera with bacterial β-1,3-glucanase are marked in red.
Figure 2.
A tree was composed of classification systems within dominant phyla at the genus level in FTL1. The abundances of taxa genera are over the top 30. The genera with bacterial β-1,3-glucanase are marked in red.

Figure 3.
Glucan-degrading actinomycetes formed a clear halo around the colony.

Figure 4.
Phylogenetic analysis of 16S rRNA gene sequences of the isolates from FTL1. A neighbor-joining tree was obtained from a BLAST search of 16S rRNA gene sequences of the isolates for phylogenetic inference. The bootstrap values presented at corresponding branches were evaluated using 1000 replicates.
Figure 4.
Phylogenetic analysis of 16S rRNA gene sequences of the isolates from FTL1. A neighbor-joining tree was obtained from a BLAST search of 16S rRNA gene sequences of the isolates for phylogenetic inference. The bootstrap values presented at corresponding branches were evaluated using 1000 replicates.

Figure 5.
Alignment of SYBCQL and SYBC17 enzymes with other β-1,3-glucanases of GHF16. 17-W and 17-Q, SYBC17; QLK1, SYBCQL; bgl27, Streptomyces sp. S27; Ss, Streptomyces sioyaensis. Sequences were taken from the following accession numbers: Streptomyces sp. S27 endo-β-1,3-glucanase (FJ887899); S. sioyaensis endo-β-1,3-glucanase (AF217415). The deduced amino acid signal peptides are underlined. The putative catalytic motif residues are boxed in black. The deduced location of linker regions are indicated by a wavy line.
Figure 5.
Alignment of SYBCQL and SYBC17 enzymes with other β-1,3-glucanases of GHF16. 17-W and 17-Q, SYBC17; QLK1, SYBCQL; bgl27, Streptomyces sp. S27; Ss, Streptomyces sioyaensis. Sequences were taken from the following accession numbers: Streptomyces sp. S27 endo-β-1,3-glucanase (FJ887899); S. sioyaensis endo-β-1,3-glucanase (AF217415). The deduced amino acid signal peptides are underlined. The putative catalytic motif residues are boxed in black. The deduced location of linker regions are indicated by a wavy line.

Figure 6.
Partial alignment of the C-terminal carbohydrate-binding domain (CBM) of SYBCQL and SYBC17 enzymes with several CBMs found in xylanases and endo-β-1,3-glucanases. QLK1 β-1,3-glucanase, SYBCQL; 17-W β-1,3-glucanase, SYBC17; bgl27 endo-β-1,3-d-glucanase, Streptomyces sp. S27 (FJ887899); YCWD3 β-1,3-glucanase, Arthrobacter sp. YCWD3 (D23668); 1KNM_A xylanase 10A, Streptomyces lividans (M64551); 1XYF_A Endo-β-1,4-Xylanase Chain A, Streptomyces Olivaceoviridis (PDB entry: 1XYF_A); 1XYF_B Endo-β-1,4-Xylanase Chain B, Streptomyces Olivaceoviridis (PDB entry: 1XYF_B). The Gln-X-Trp repeats are boxed in black.
Figure 6.
Partial alignment of the C-terminal carbohydrate-binding domain (CBM) of SYBCQL and SYBC17 enzymes with several CBMs found in xylanases and endo-β-1,3-glucanases. QLK1 β-1,3-glucanase, SYBCQL; 17-W β-1,3-glucanase, SYBC17; bgl27 endo-β-1,3-d-glucanase, Streptomyces sp. S27 (FJ887899); YCWD3 β-1,3-glucanase, Arthrobacter sp. YCWD3 (D23668); 1KNM_A xylanase 10A, Streptomyces lividans (M64551); 1XYF_A Endo-β-1,4-Xylanase Chain A, Streptomyces Olivaceoviridis (PDB entry: 1XYF_A); 1XYF_B Endo-β-1,4-Xylanase Chain B, Streptomyces Olivaceoviridis (PDB entry: 1XYF_B). The Gln-X-Trp repeats are boxed in black.

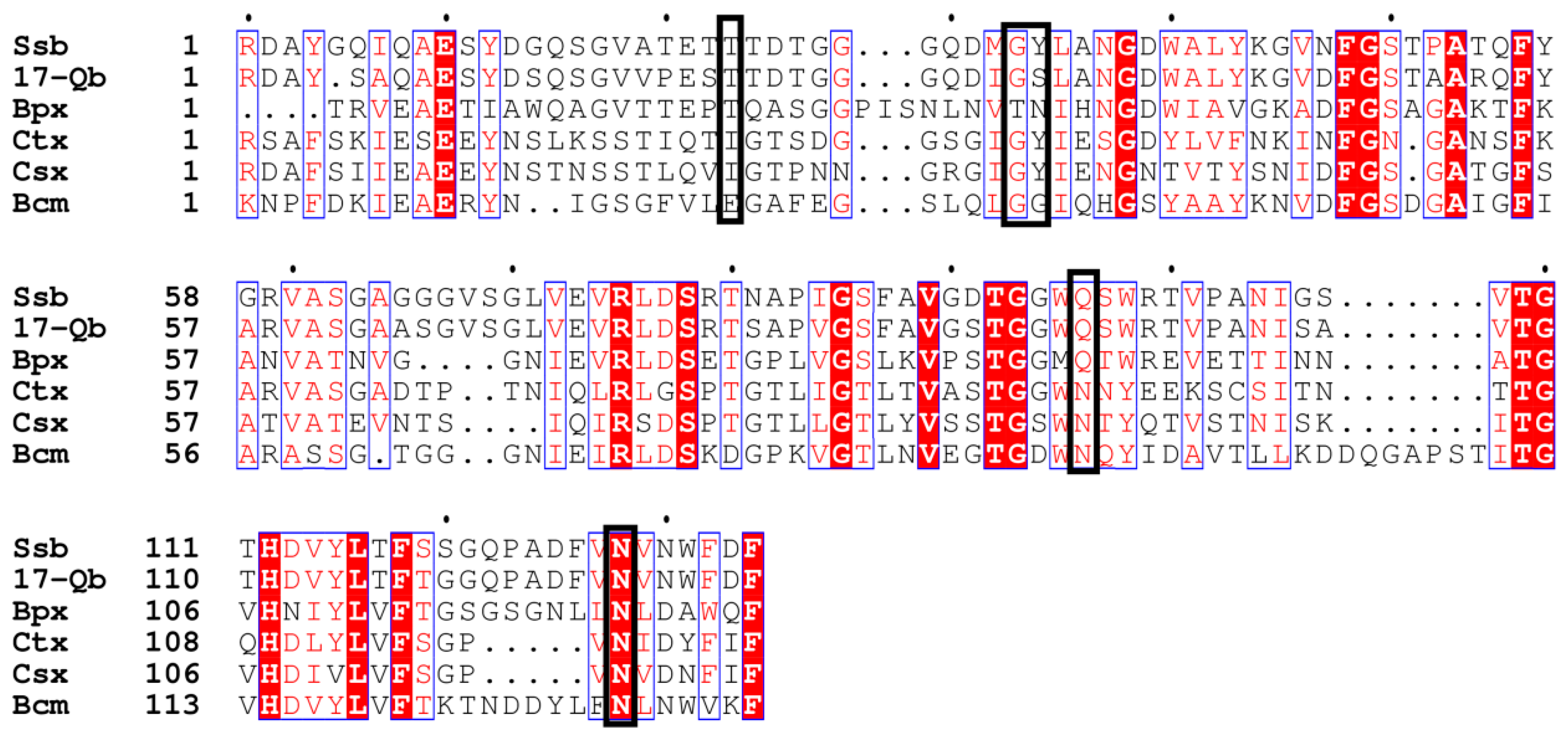
Figure 7.
Partial alignment of the C-terminal CBM of SYBC17 enzyme with several CBMs found in xylanases and endo-β-1,3-glucanases. 17-Q β-1,3-glucanase, SYBC17; Ssb endo-β-1,3-glucanase, Streptomyces sioyaensis (AF217415); Ctx xylanase A, Clostridium thermocellum F1/YS (AF04776); Csx xylanase A, Clostridium stercorarium F-9 (D13325); Bpx xylanase D, Bacillus polymyxa (X57094); Bcm α-1,6-mannanase, Bacillus circulans TN31(AB024331). The ligand binding sites are boxed in black.
Figure 7.
Partial alignment of the C-terminal CBM of SYBC17 enzyme with several CBMs found in xylanases and endo-β-1,3-glucanases. 17-Q β-1,3-glucanase, SYBC17; Ssb endo-β-1,3-glucanase, Streptomyces sioyaensis (AF217415); Ctx xylanase A, Clostridium thermocellum F1/YS (AF04776); Csx xylanase A, Clostridium stercorarium F-9 (D13325); Bpx xylanase D, Bacillus polymyxa (X57094); Bcm α-1,6-mannanase, Bacillus circulans TN31(AB024331). The ligand binding sites are boxed in black.

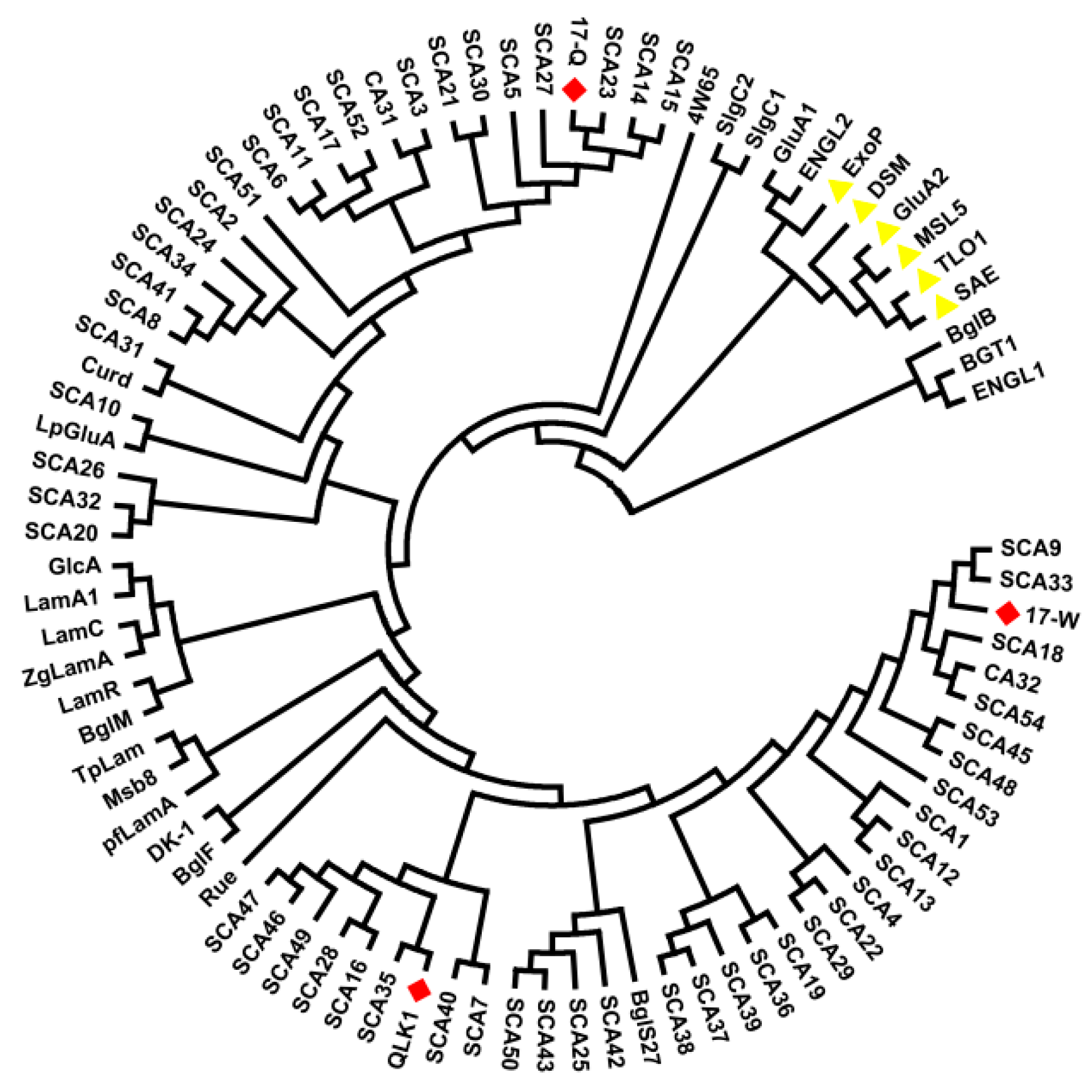
Figure 8.
Phylogenetic tree of some amino acid sequences of β-1,3-glucanase. The deduced amino acid sequences of β-1,3-glucanase from SYBCQL and SYBC17 are marked in red. Glycoside hydrolases belonging to the exo-type clade are marked in yellow and others are divided into endo-types.
Figure 8.
Phylogenetic tree of some amino acid sequences of β-1,3-glucanase. The deduced amino acid sequences of β-1,3-glucanase from SYBCQL and SYBC17 are marked in red. Glycoside hydrolases belonging to the exo-type clade are marked in yellow and others are divided into endo-types.

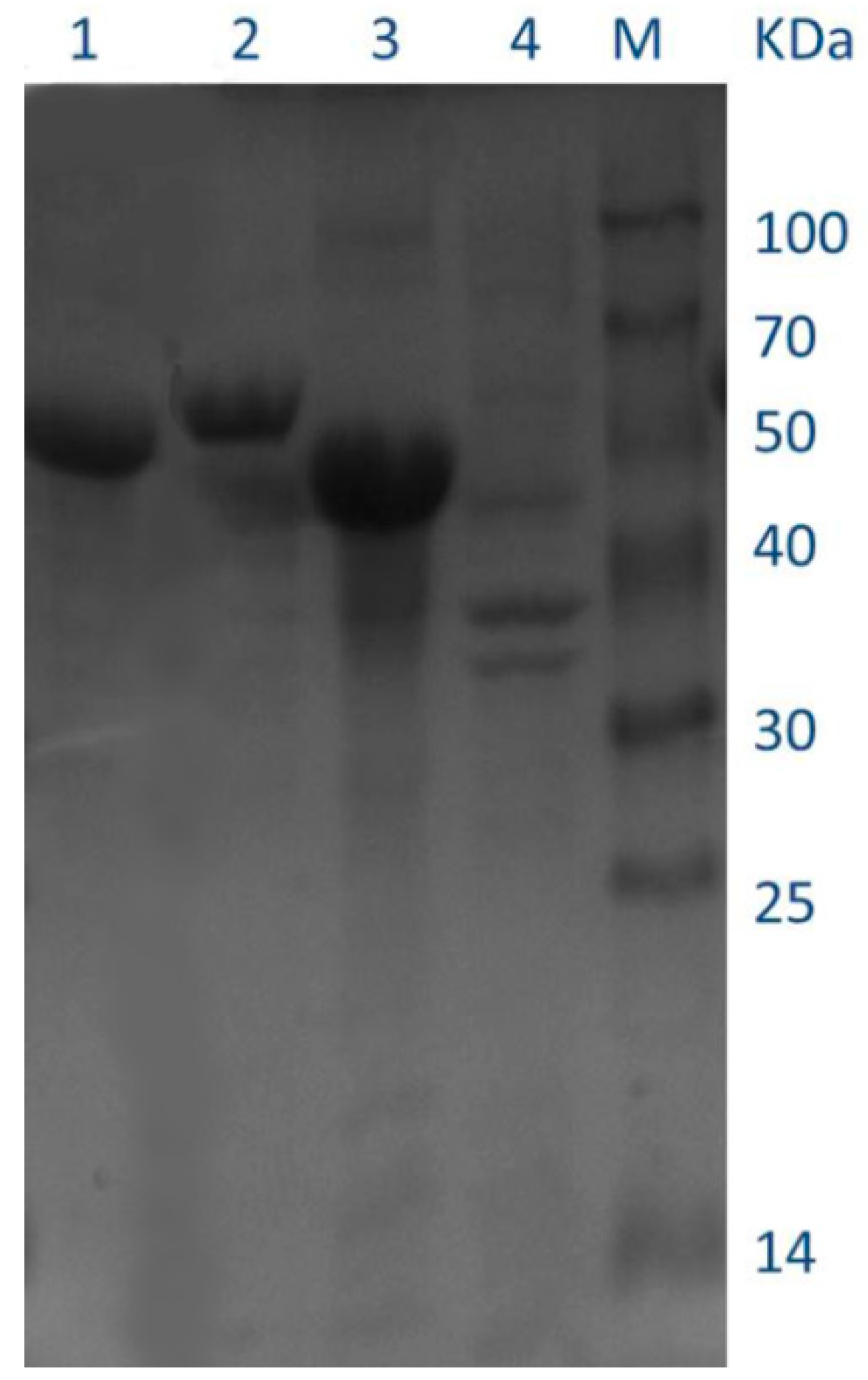
Figure 9.
SDS-PAGE analysis of the purified recombinant enzymes overexpressed in E. coli BL21 (DE3). Lanes: M, molecular mass markers; 1, purified recombinant QLK1; 2, purified recombinant 17-Q; 3, purified recombinant 17-W; 4, culture supernatant of the induced transformant harboring empty pcold II.
Figure 9.
SDS-PAGE analysis of the purified recombinant enzymes overexpressed in E. coli BL21 (DE3). Lanes: M, molecular mass markers; 1, purified recombinant QLK1; 2, purified recombinant 17-Q; 3, purified recombinant 17-W; 4, culture supernatant of the induced transformant harboring empty pcold II.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bacterial richness indices of soil samples.
| Samples | Number of Effective Sequences | Number of OTUs | Coverage (%) | Chao1 | Shannon Index |
|---|---|---|---|---|---|
| FLT1 | 44,870 | 7737 | 88.51 | 25,534.52 | 6.95 |
| FLT2 | 57,135 | 10,709 | 88.56 | 34,009.99 | 7.52 |
Table 2.
BLAST analysis of glucan-degrading microorganisms isolated from FLT1.
| Isolate Name | 16S rRNA Gene Sequence Lengh (bp) | Best BLAST Hit(s) | Accession Number | Identity (%) |
|---|---|---|---|---|
| SYBCQL | 1345 | Kitasatospora phosalacinea JKCM-G-8A Kitasatospora phosalacinea NBRC 14372 a | LC010672.1 NR_112434.1 | 99 99 |
| SYBCA | 1396 | Streptomyces cinerochromogenes MC10130 Streptomyces cinerochromogenes NBRC 13822 a Streptomyces coelescens CSSP420 Streptomyces cinerochromogenes 3CSSP87 | AB968639.1 NR_041153.1 NR_115375.1 NR_115366.1 | 99 99 99 99 |
| SYBC6 | 1418 | Streptomyces cellostaticus HUBZM22 Streptomyces capoamus JCM 4734 a | HQ853021.1 NR_040856.1 | 99 99 |
| SYBC7 | 1399 | Streptomyces cellostaticus HUBZM22 Streptomyces cellostaticus NBRC 12849 a | HQ853021.1 NR_112304.1 | 99 99 |
| SYBC8 | 1416 | Streptomyces cinerochromogenes MC10130 Streptomyces cinerochromogenes NBRC 13822 a Streptomyces coelescens CSSP420 Streptomyces cinerochromogenes 3CSSP87 | AB968639.1 NR_041153.1 NR_115375.1 NR_115366.1 | 99 99 99 99 |
| SYBC16 | 1417 | Streptomyces olivogriseus NBRC 13795 Streptomyces filipinensis NBRC 12860 a | AB184486.1 NR_041083.1 | 99 99 |
| SYBC17 | 1389 | Streptomyces cellostaticus HUBZM22 Streptomyces capoamus JCM 4734 a | HQ853021.1 NR_040856.1 | 99 99 |
| SYBC24 | 1393 | Streptomyces sp. X4-5 Streptomyces viridochromogenes NBRC 13347 a | KT581286.1 NR_112526.1 | 99 99 |
| SYBC25 | 1393 | Streptomyces cellostaticus HUBZM22 Streptomyces capoamus JCM 4734 a | HQ853021.1 NR_040856.1 | 99 99 |
| SYBC26 | 1393 | Streptomyces indiaensis LMG 19961 Streptomyces indiaensis NBRC 13964 a | AJ781344.1 NR_041155.1 | 99 99 |
| SYBC27 | 1422 | Streptomyces cellostaticus HUBZM22 Streptomyces cellostaticus NBRC 12849 a | HQ853021.1 NR_112304.1 | 99 99 |
a The best homology 16S rRNA sequences from type material based on a BLAST search of the 16S rRNA sequences among isolates for high similar inference.
Table 3.
Determination of β-1,3-glucan-degrading activity from culture filtrates of actinomycetes.
| Isolates | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) |
|---|---|---|---|
| SYBCQL | 31.22 | 3.75 | 0.12 |
| SYBCA | 26.31 | 23.15 | 0.88 |
| SYBC6 | 10.44 | 5.84 | 0.56 |
| SYBC7 | 23.82 | 13.10 | 0.55 |
| SYBC8 | 32.08 | 3.85 | 0.12 |
| SYBC16 | 24.21 | 10.65 | 0.44 |
| SYBC17 | 37.85 | 38.60 | 1.02 |
| SYBC24 | 34.21 | 6.16 | 0.18 |
| SYBC25 | 8.22 | 2.71 | 0.33 |
| SYBC26 | 10.28 | 4.52 | 0.44 |
| SYBC27 | 33.82 | 4.06 | 0.12 |
Note: The volume of culture filtrates is 50 mL.
Table 4.
The amino acid sequences of β-1,3-glucanase are used to construct the phylogenetic tree. The abbreviations of proteins correspond to positions in the phylogenetic tree of Figure 8.
Table 4.
The amino acid sequences of β-1,3-glucanase are used to construct the phylogenetic tree. The abbreviations of proteins correspond to positions in the phylogenetic tree of Figure 8.
| Organism | Protein Abbreviation | Accession No/PDB No |
|---|---|---|
| Streptomyces sioyaensis | Curd | AF217415 |
| Streptomyces matensis ATCC 23935 | LPHase | AB019428 |
| Streptomyces sp. S27 | BglS27 | FJ887899 |
| Streptomyces clavuligerus ATCC | SCA1 | EFG04651 |
| Streptomyces acidiscabies | SCA2 | WP_010357589 |
| Streptomyces coelicoflavus | SCA55 | WP_007387290 |
| Streptomyces avermitilis | SCA4 | WP_010988837 |
| Streptomyces avermitilis | SCA5 | WP_010983203 |
| Streptomyces azureus | SCA6 | GAP51072 |
| Streptomyces bingchenggensis BCW-1 | SCA7 | ADI05411 |
| Streptomyces canus | SCA8 | WP_020122288 |
| Streptomyces canus | SCA9 | WP_020117105 |
| Streptomyces cattleya | SCA10 | WP_014626989 |
| Streptomyces chartreusis | SCA11 | WP_010043060 |
| Streptomyces clavuligerus ATCC 27064 | SCA12 | EDY52285 |
| Streptomyces clavuligerus | SCA13 | WP_003957976 |
| Streptomyces collinus Tu 365 | SCA14 | AGS72893 |
| Streptomyces collinus | SCA15 | WP_020943303 |
| Streptomyces griseoaurantiacus | SCA16 | WP_006140385 |
| Streptomyces griseoflavus Tu4000 | SCA17 | EFL37893 |
| Streptomyces griseoflavus | SCA18 | WP_004921557 |
| Streptomyces griseus | SCA19 | WP_012377737 |
| Streptomyces himastatinicus | SCA20 | WP_009714916 |
| Streptomyces hokutonensis | SCA21 | WP_019069886 |
| Streptomyces hokutonensis | SCA22 | WP_019068505 |
| Streptomyces hygroscopicus | SCA23 | WP_014676131 |
| Streptomyces lincolnensis | SCA24 | ANS69291 |
| Streptomyces malaysiensis | SCA25 | ATL81267 |
| Streptomyces niveus | SCA26 | WP_023538571 |
| Streptomyces olivochromogenes | SCA27 | GAX48907 |
| Streptomyces pratensis | SCA28 | WP_014152186 |
| Streptomyces prunicolor | SCA29 | WP_019057733 |
| Streptomyces prunicolor | SCA30 | WP_019059013 |
| Streptomyces roseochromogenus | SCA31 | WP_023545390 |
| Streptomyces scopuliridis RB72 | SCA32 | PVE09807 |
| Streptomyces sp. 351MFTsu5.1 | SCA33 | WP_020134833 |
| Streptomyces sp. 351MFTsu5.1 | SCA34 | WP_020135556 |
| Streptomyces sp. AA4 | SCA35 | EFL08653 |
| Streptomyces sp. ACT-1 | SCA36 | WP_003964231 |
| Streptomyces sp. SPB074 | SCA37 | WP_008747151 |
| Streptomyces sp. SPB78 | SCA38 | EFL00779 |
| Streptomyces sp. SPB78 | SCA39 | EFK98142 |
| Streptomyces sparsogenes DSM 40356 | SCA40 | OMI34149 |
| Streptomyces sviceus | SCA41 | WP_007386005 |
| Streptomyces thermolilacinus | SCA42 | WP_023590036 |
| Streptomyces violaceusniger | SCA43 | WP_014059092 |
| Streptomyces viridochromogenes | SCA44 | WP_003994249 |
| Streptomyces viridosporus ATCC 14672 | SCA45 | EFE71345 |
| Streptomyces viridosporus ATCC 14672 | SCA46 | EFE68955 |
| Streptomyces viridosporus | SCA47 | WP_004986925 |
| Streptomyces viridosporus | SCA48 | WP_016827665 |
| Streptomyces viridosporus | SCA49 | WP_016825877 |
| Streptomyces violaceusniger Tu 4113 | SCA50 | AEM85607 |
| Streptomyces sp. SCC 2136 | SCA51 | CAF31374 |
| Streptomyces zinciresistens | SCA52 | WP_007495688 |
| Streptomyces zinciresistens | SCA53 | WP_007501949 |
| Streptomyces coelicoflavus | SCA54 | WP_007389367 |
| Streptomyces lydicus | SlgC1SlgC2 | CBA11580CBA11566 |
| Nocardiopsis sp. F96 | BglF | AB244275 |
| Arthrobacter sp. Rue61a | Rue | WP_014920770 |
| Streptomyces sp. SirexAA-E | SAE | G2NFJ9 |
| Streptomyces hygroscopicus subsp. jinggangensis TL01 | TL01 | AEY93509 |
| Arthrobacter sp. NHB-10 | GluA2 | AB289602 |
| SYBC17 | 17-W | MH190407 |
| SYBC17 | 17-Q | MH190408 |
| SYBCQL | QLK1 | MH190409 |
| Cellulosimicrobium cellulans DK-1 | DK-1 | EU589324 |
| B. circulans | GlcA | P23903 |
| T. maritima Msb8 | Msb8 | 3AZX |
| B.circulans bglM | BglM | AB078775 |
| Pseudomonas sp. PE2 | GluA1 | BAC16331 |
| Zobellia galactanivorans | ZgLamA | 4BQ1 |
| Paenibacillus sp. CCRC 17245 | LamA1 | ABJ15796 |
| Corallococcus sp. | LamC | KX583630 |
| Mycobacterium fortuitum | 4W65 | 4W65 |
| Thermotoga neapolitana | BglB | Z77856 |
| Thermotoga petrophila | TpLam | CP000702 |
| Pyrococcus furiosus | pfLamA | 2VY0 |
| Corallococcus sp. | LamC | KX583630 |
| Rhodothermus marinus ITI278 | LamR | AAC69707 |
| Aspergillus fumigatus | ENGL2 | AFUA_2G14360 |
| Aspergillus fumigatus | BGT1 | AF038596 |
| Aspergillus fumigatus | ENGL1 | AFUA_1G04260 |
| Pseudoalteromonas sp. BB1 | ExoP | DQ361032 |
| Actinosynnema mirum DSM 43827 | DSM | ACU35625 |
| Micromonospora sp. L5 | MSL5 | ADU06434 |
| Laceyella putida | LpGluA | LC060791 |
| Streptomyces coelicolor A3(2) | CA31 | NP_630740 |
| Streptomyces coelicolor A3(2) | CA32 | NP_625089 |
Table 5.
Summary of the purification of recombinant enzymes.
| Named | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) |
|---|---|---|---|
| QLK1 | 2.11 | 138.88 | 65.82 |
| 17-W | 1.31 | 174.09 | 132.90 |
| 17-Q | 2.33 | 34.26 | 14.70 |
Table 6.
Sequences of the target gene primers.
| Gene Name | Primer Name | Primer Sequences (5′-3′) | Restriction Site |
|---|---|---|---|
| 17-W | 17-WF | GCCGAAGCTTATGGCCTCCCCCCGCCTGCTCC | Hind III |
| 17-WR | GCCGTCTAGATCAGCCGACCGTCCACTTCTGGTTGGC | Xba I | |
| 17-Q | 17-QF | GCCGAAGCTTATGAGTGAAACCTCCGGCATACCCA | Hind III |
| 17-QR | GCCGTCTAGATCAGTGACCGAAGTCGAACCAGTTCAC | Xba I | |
| QLK1 | QLK1-F | GCCGAAGCTTATGGCTGCTGCCCCACGCACGCGC | Hind III |
| QLK1-R | GCCGTCTAGATCAGCCCAGCGTCCACTTCTGCGCGCC | Xba I |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, Q.; Dou, X.; Wang, Q.; Guan, Z.; Cai, Y.; Liao, X. Isolation of β-1,3-Glucanase-Producing Microorganisms from Poria cocos Cultivation Soil via Molecular Biology. Molecules 2018, 23, 1555. https://doi.org/10.3390/molecules23071555
AMA Style
Wu Q, Dou X, Wang Q, Guan Z, Cai Y, Liao X. Isolation of β-1,3-Glucanase-Producing Microorganisms from Poria cocos Cultivation Soil via Molecular Biology. Molecules. 2018; 23(7):1555. https://doi.org/10.3390/molecules23071555
Chicago/Turabian StyleWu, Qiulan, Xin Dou, Qi Wang, Zhengbing Guan, Yujie Cai, and Xiangru Liao. 2018. "Isolation of β-1,3-Glucanase-Producing Microorganisms from Poria cocos Cultivation Soil via Molecular Biology" Molecules 23, no. 7: 1555. https://doi.org/10.3390/molecules23071555