Benzothiadiazole and B-Aminobutyricacid Induce Resistance to Ectropis Obliqua in Tea Plants (Camellia Sinensis (L.) O. Kuntz)

Abstract
:1. Introduction
2. Results
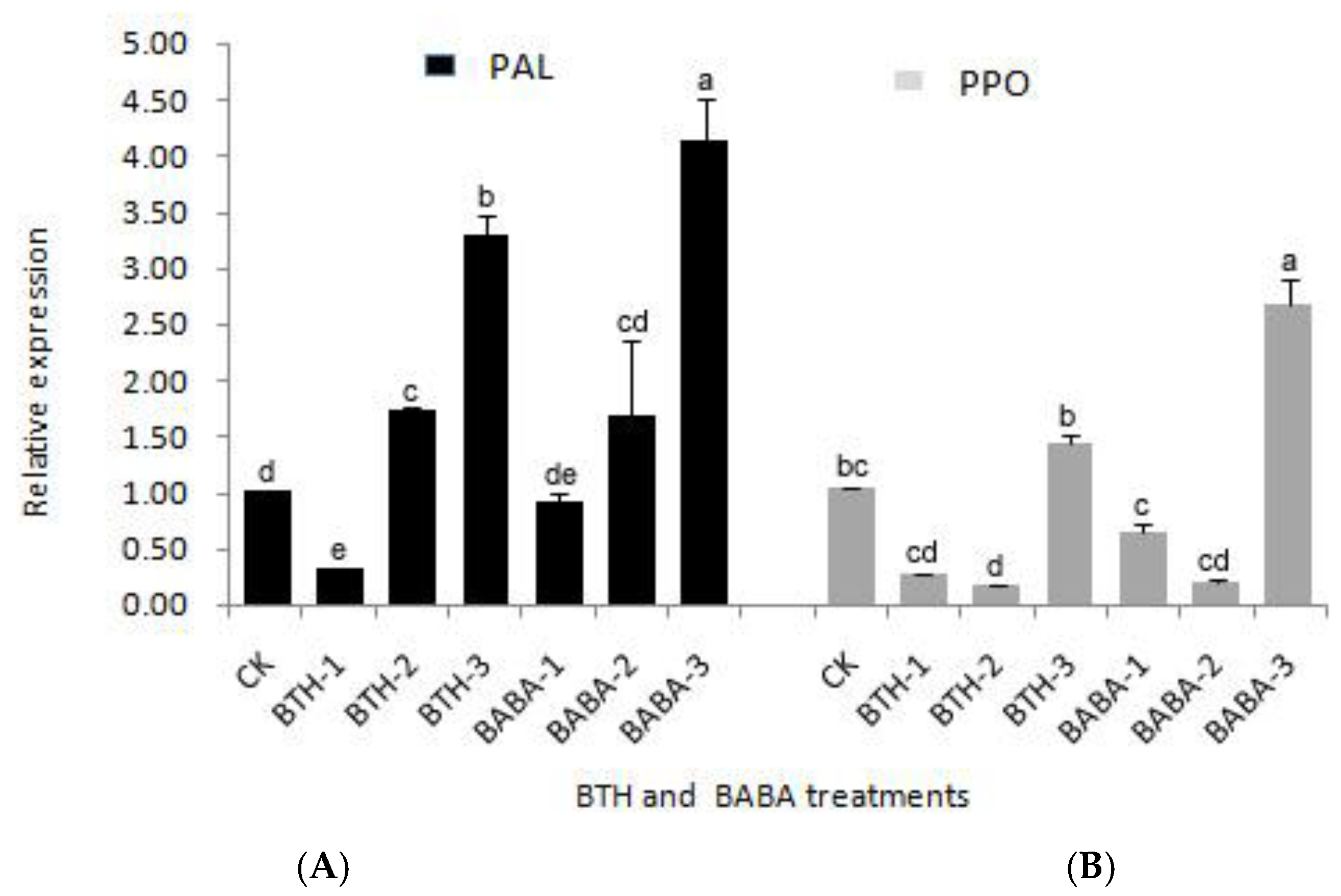
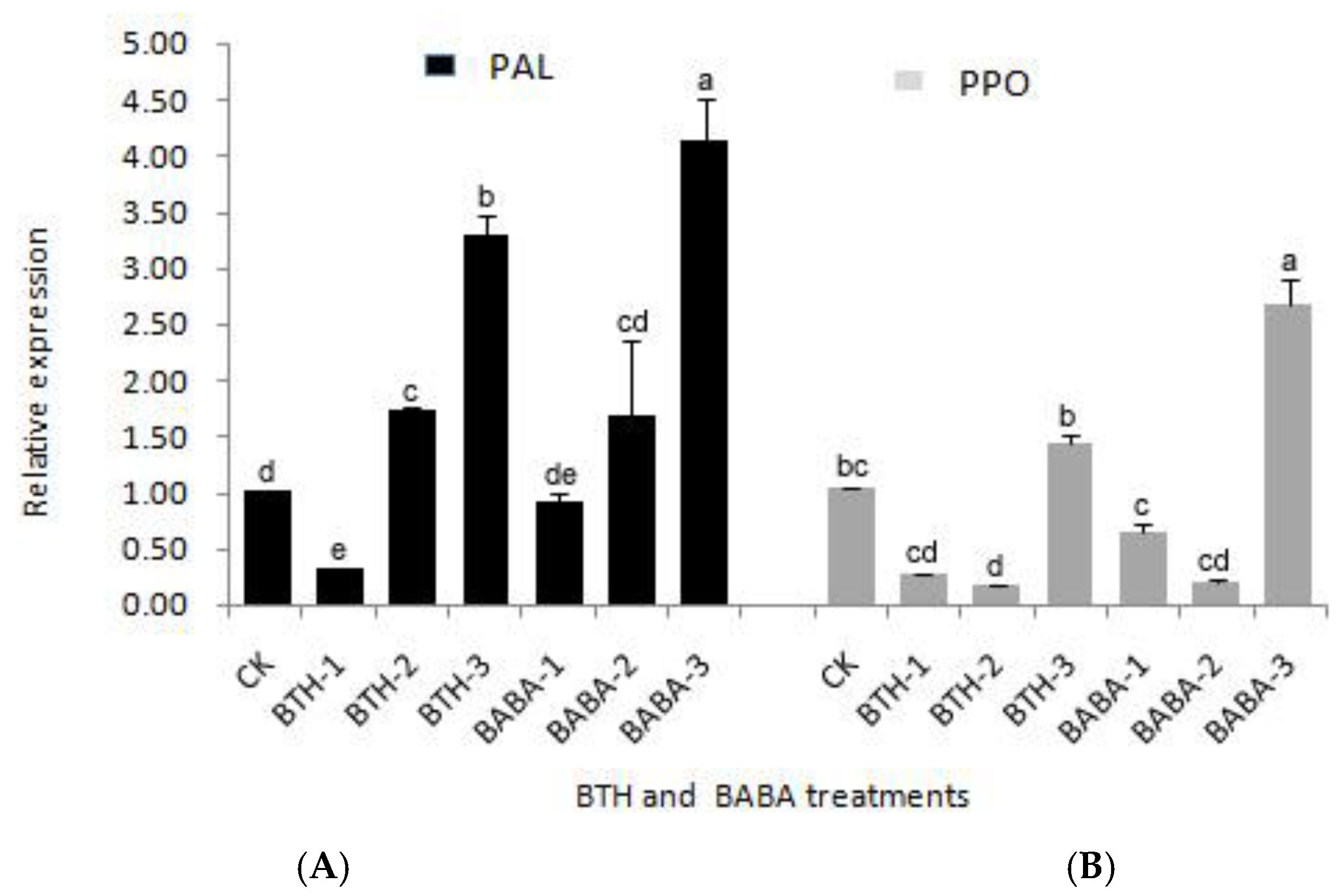
2.1. Differential Expression Analysis of PPO and PAL Genes
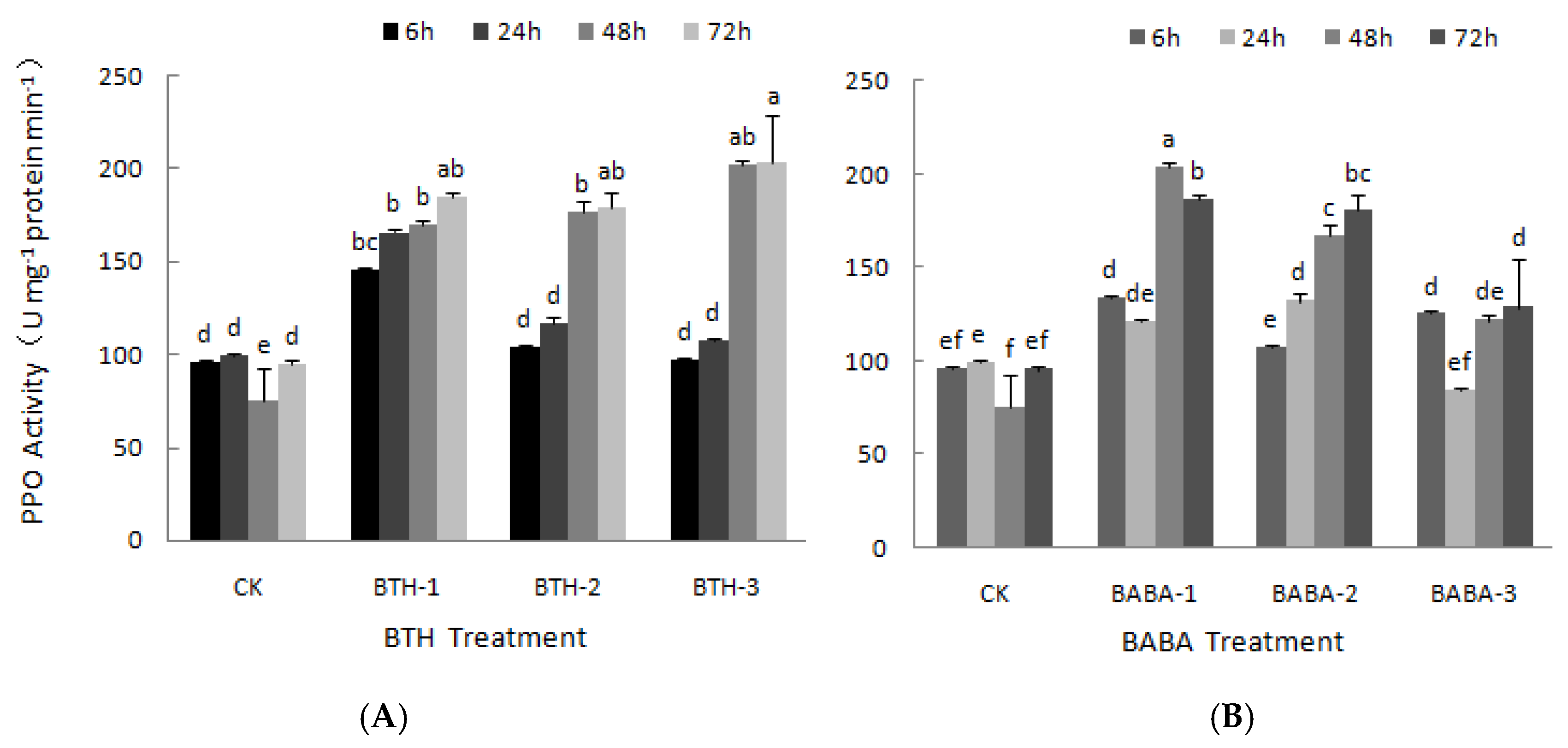
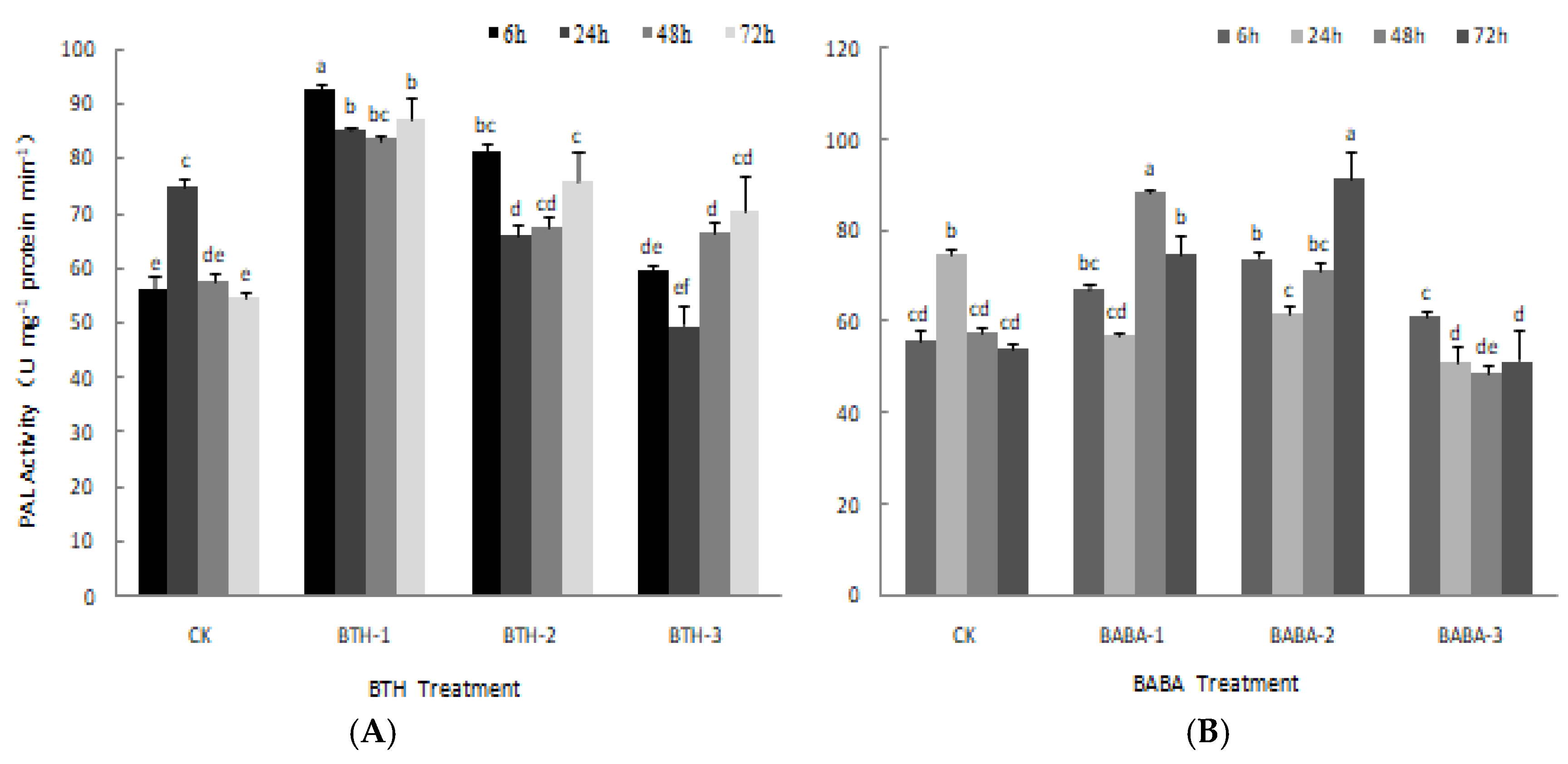
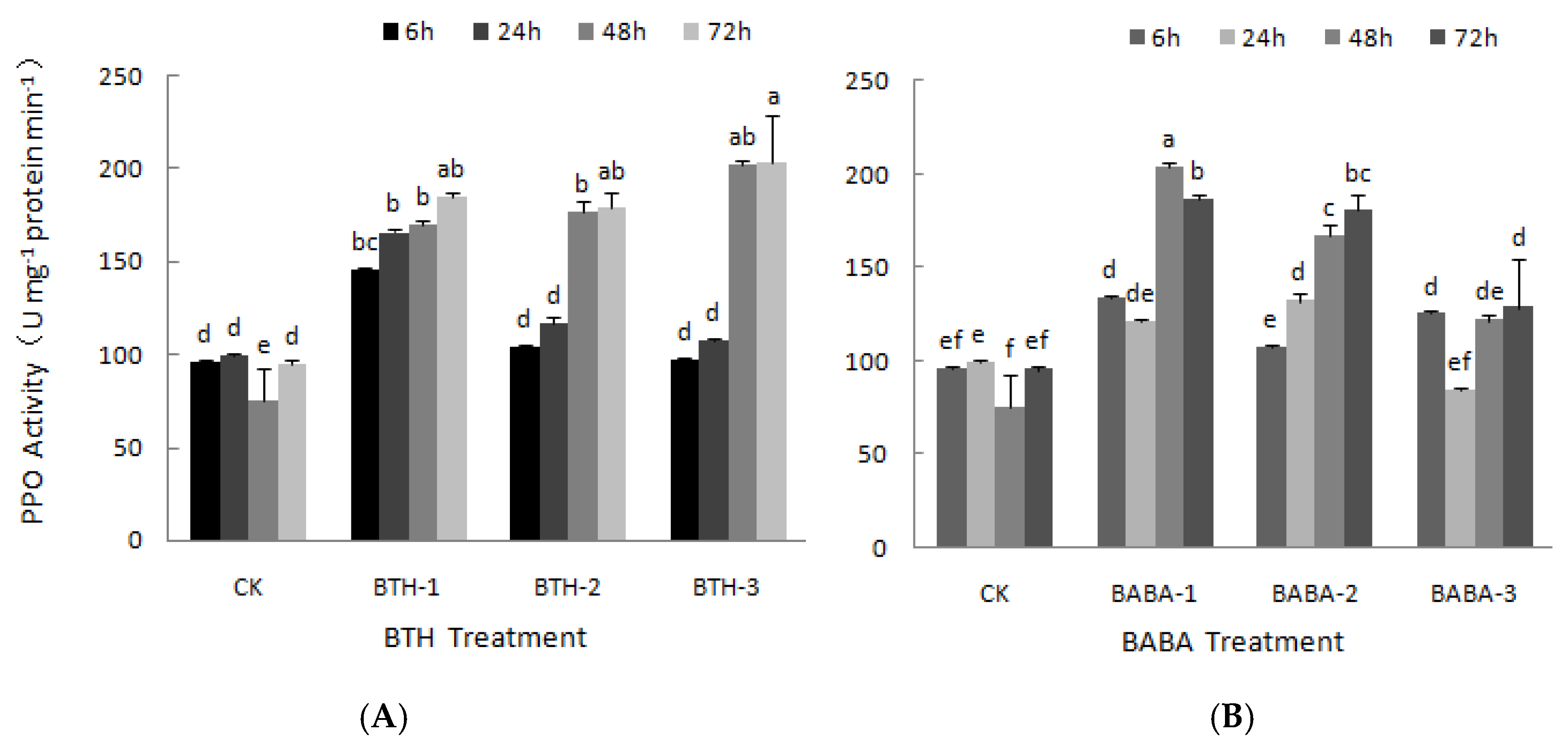
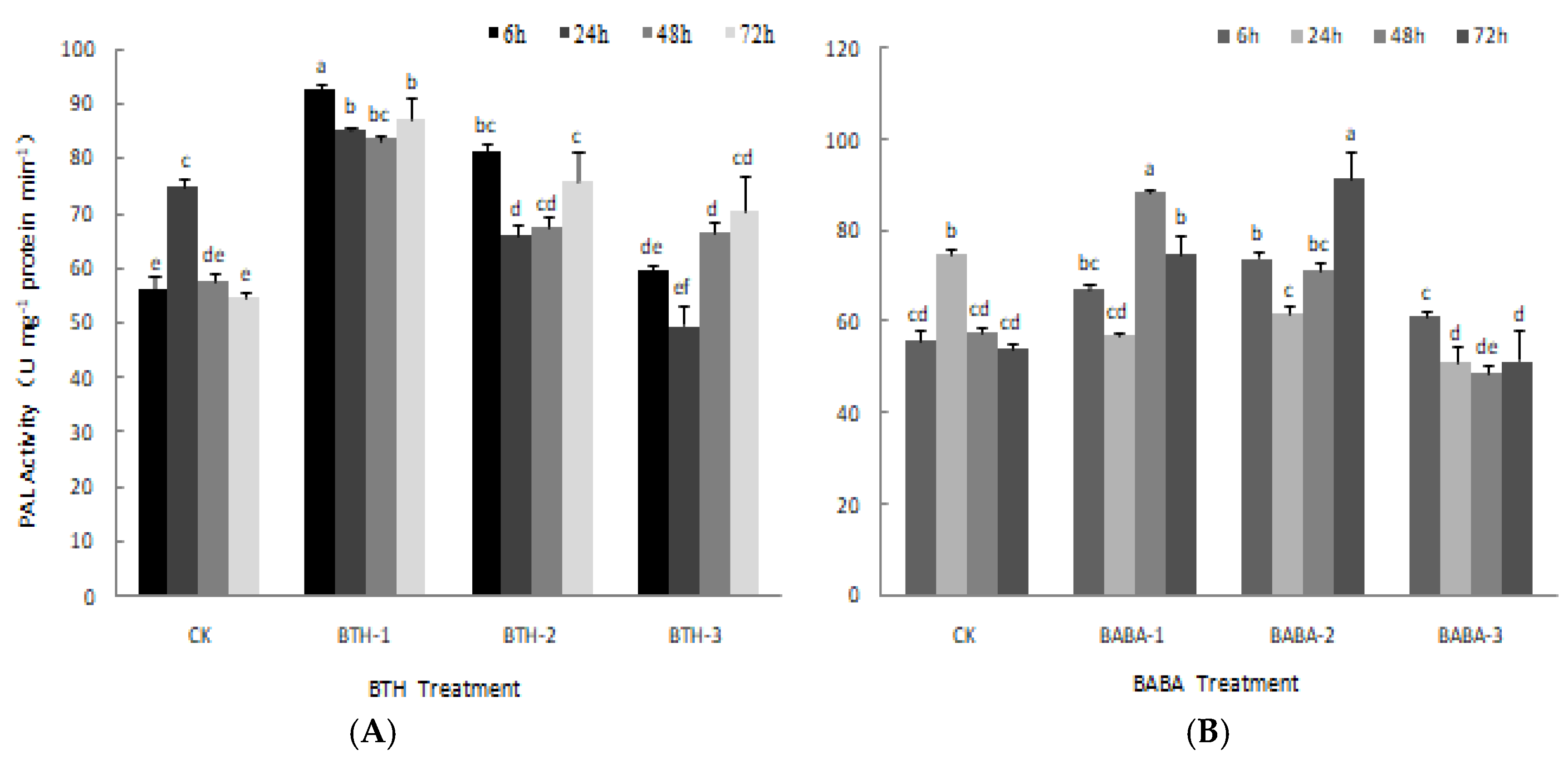
2.2. Effect of BTH and BABA on PPO and PAL Activities of Tea Plant
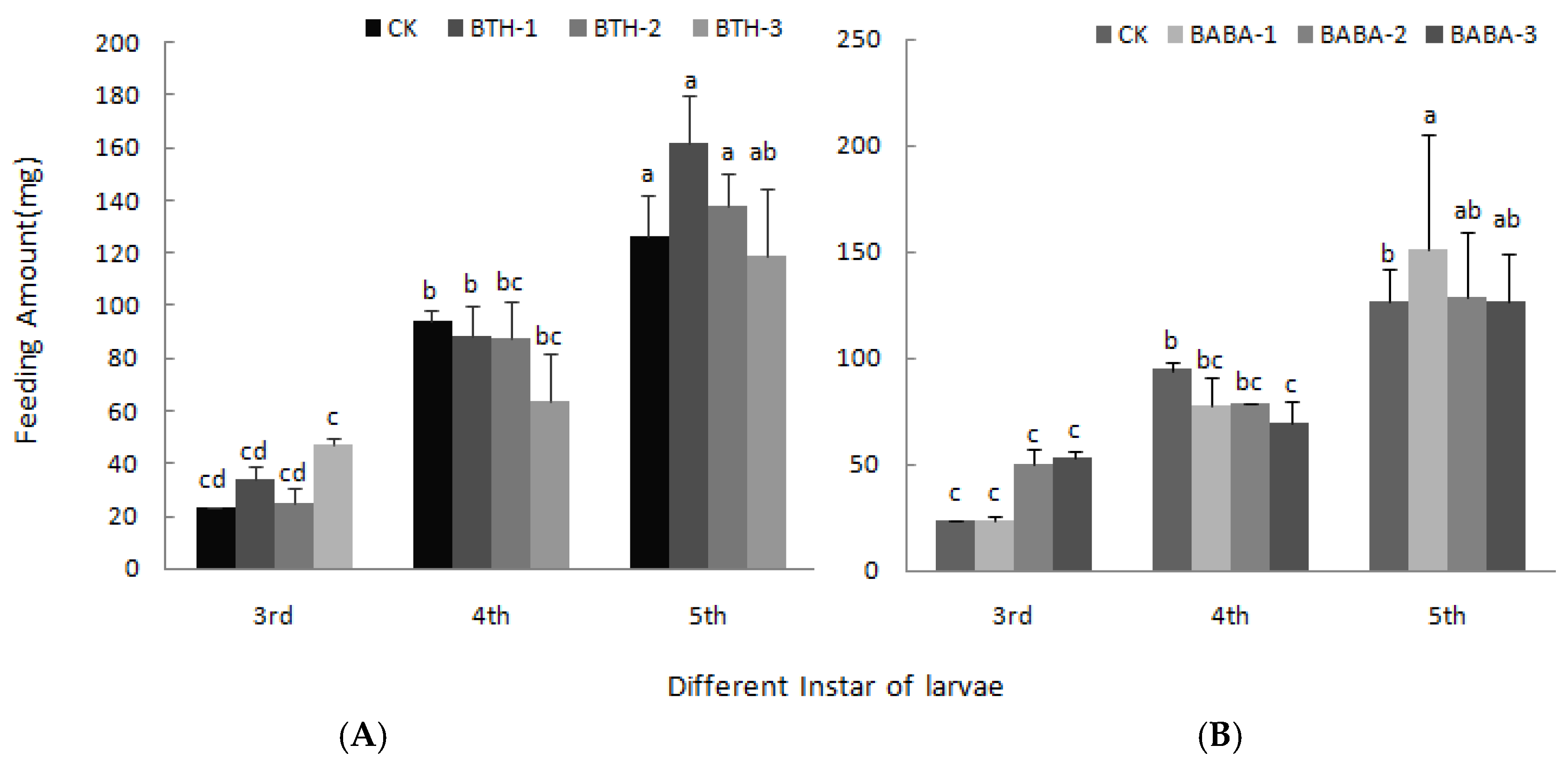
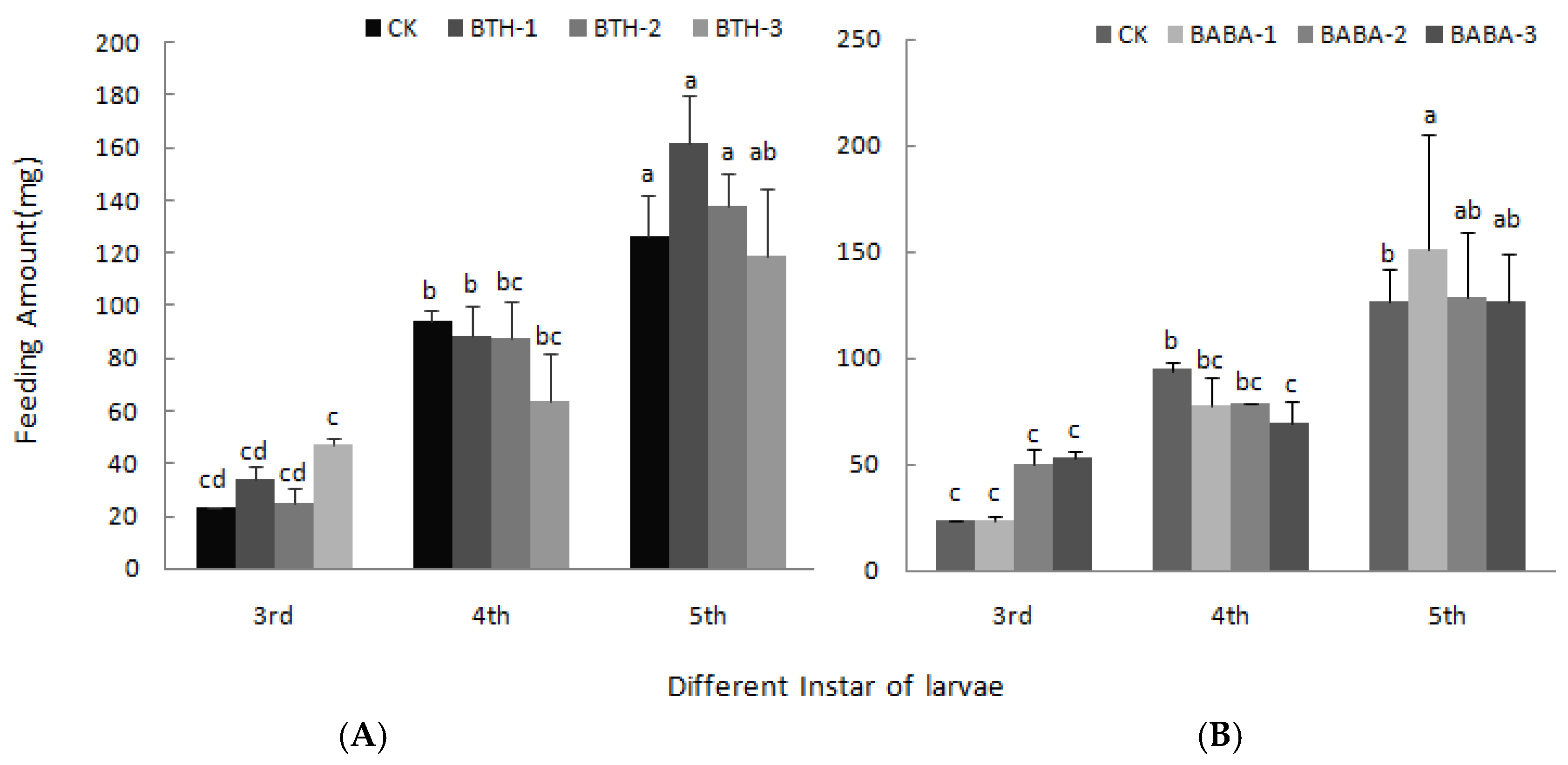
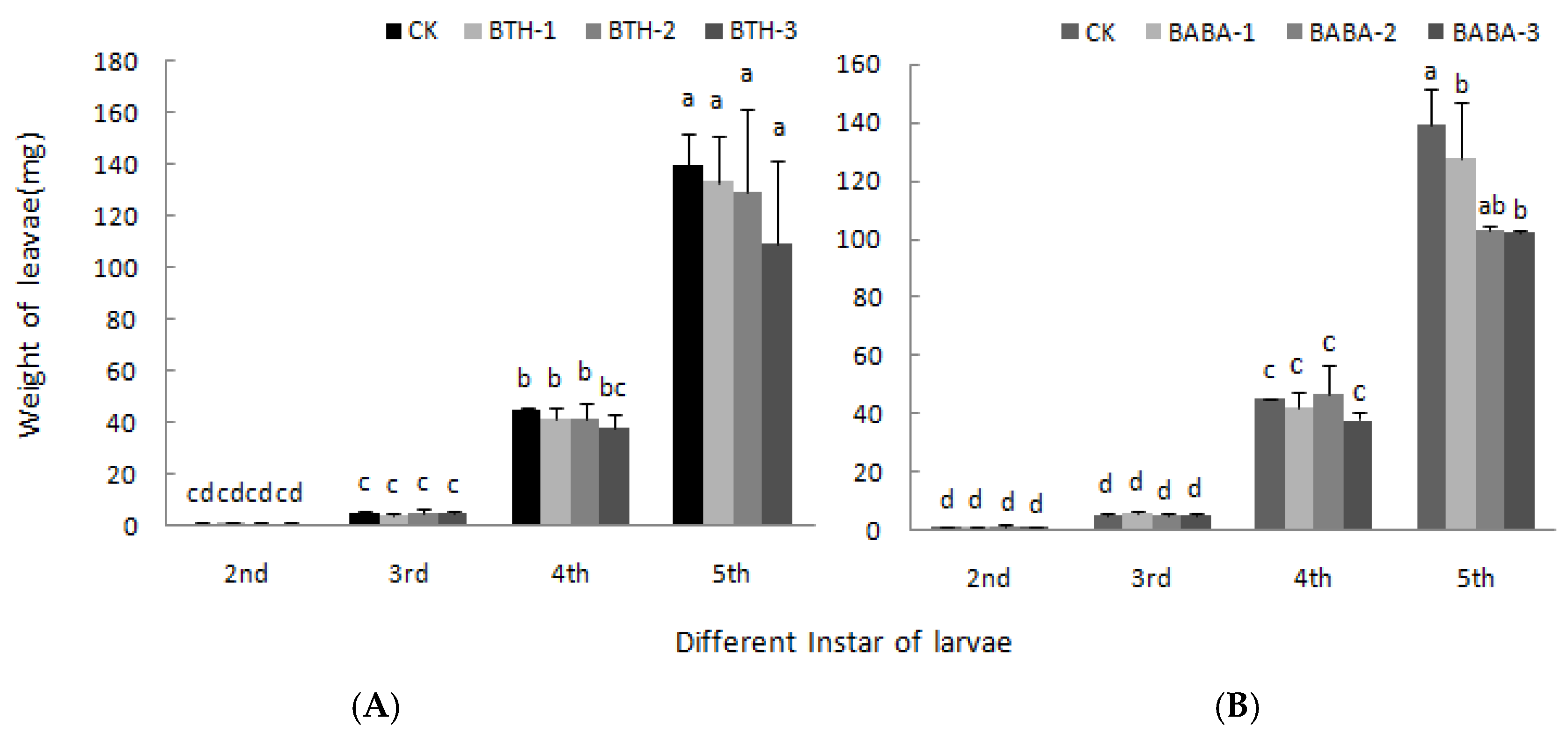
2.3. Effect of BTH and BABA on Growth of Geometrid Larvae
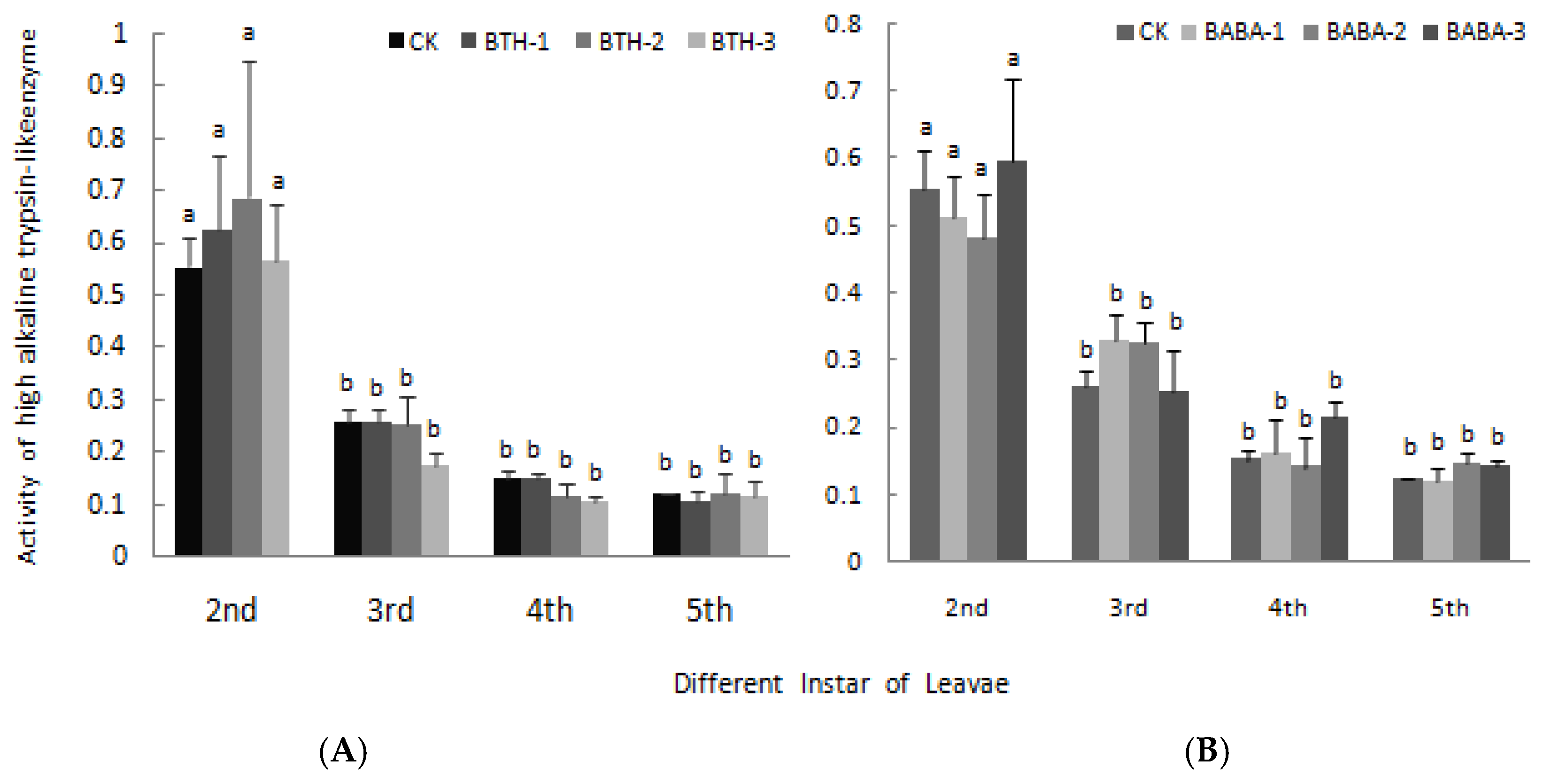
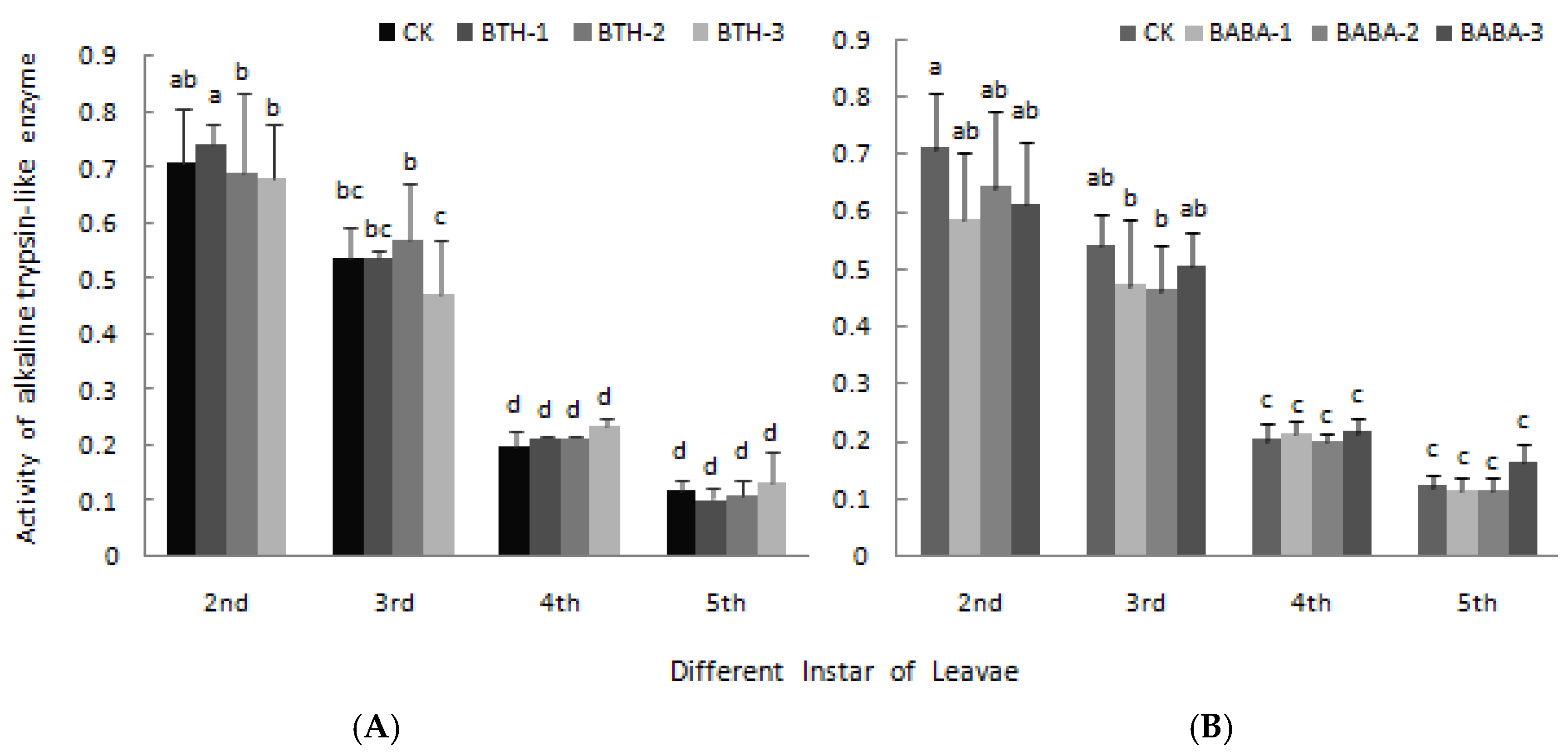
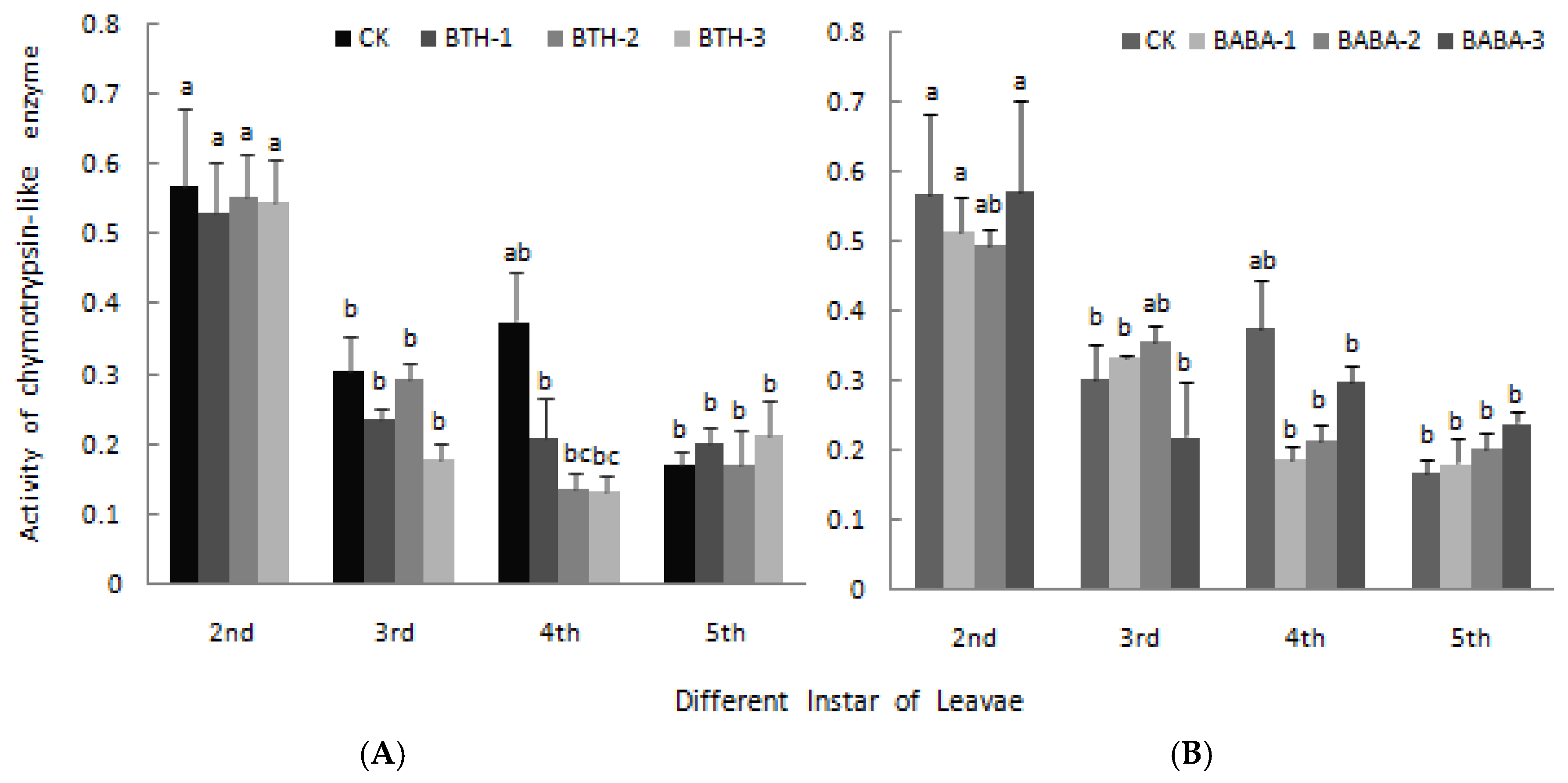
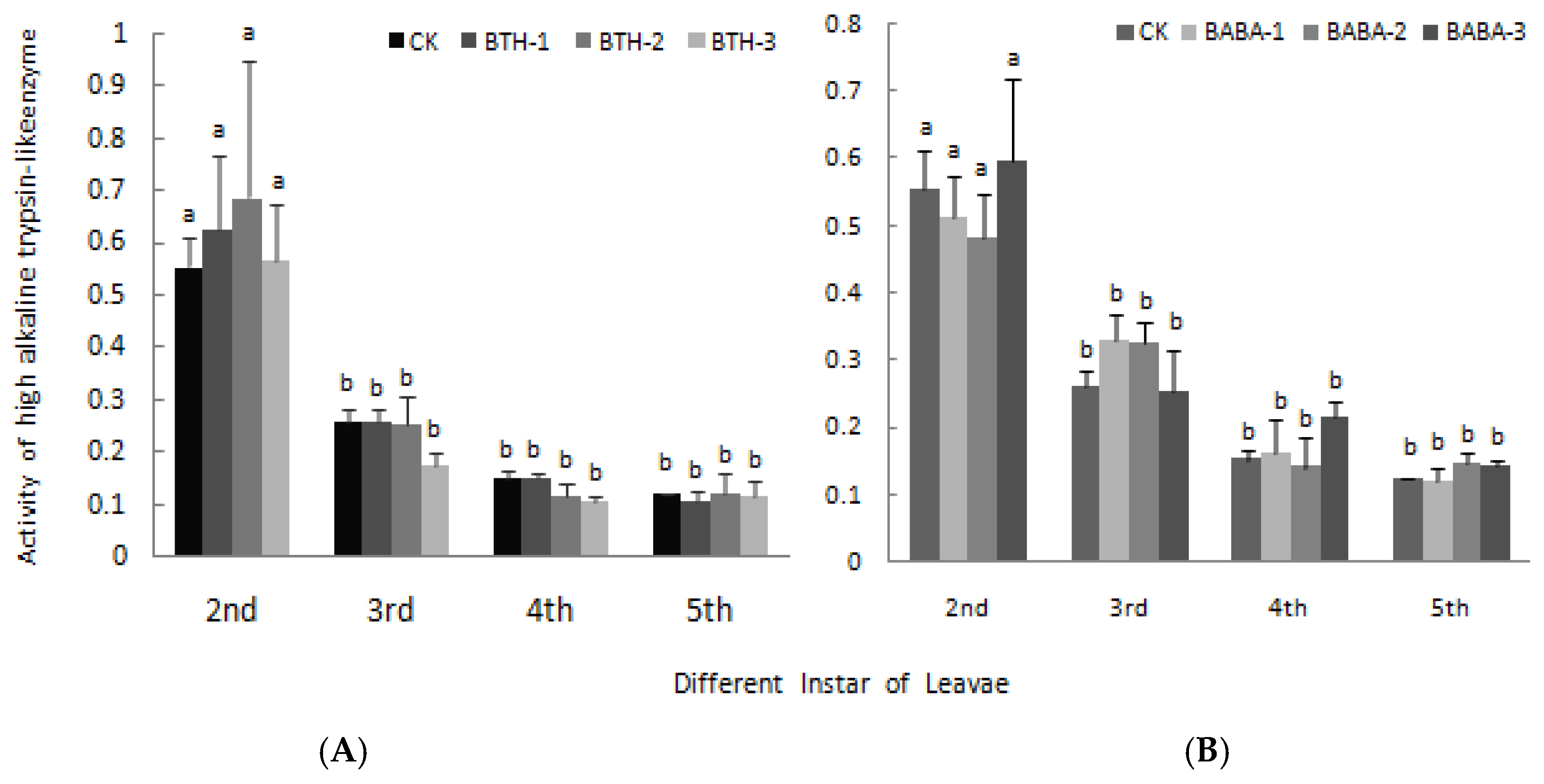
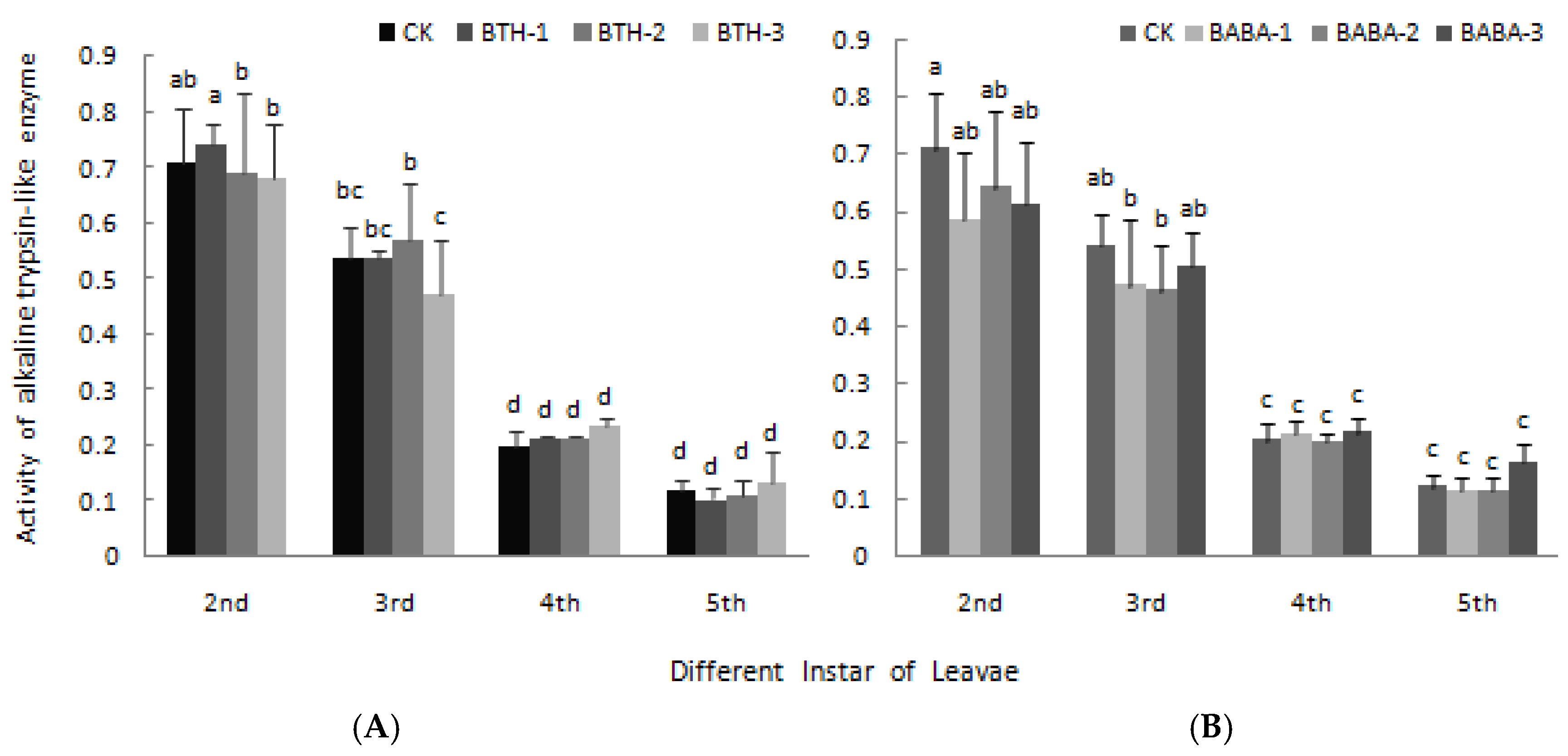
2.4. Effect of BTH and BABA on Midgut Proteinase Activity of Geometrid Larvae
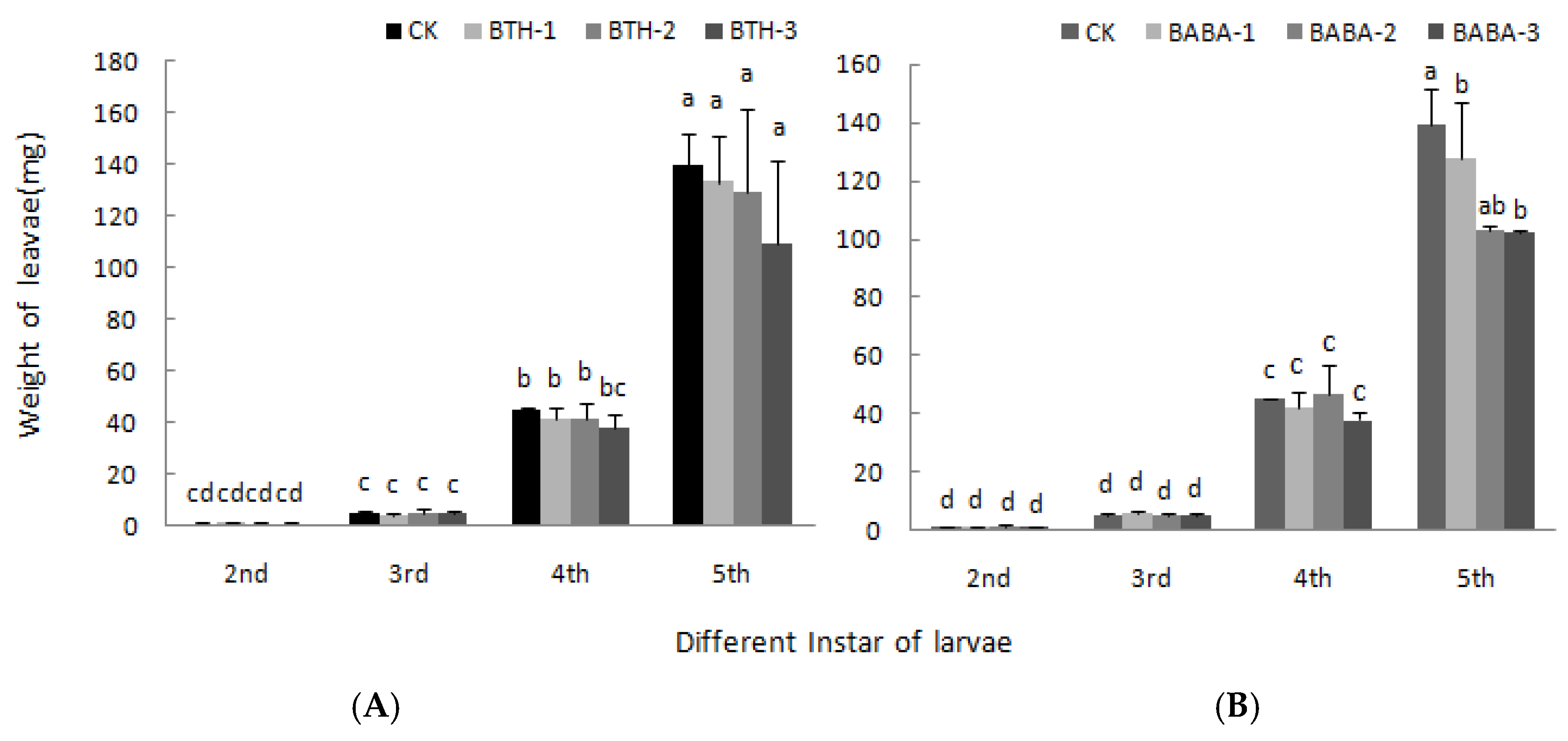
2.5. Effect of BTH and BABA on Geometrid Larvae Development
3. Discussion
4. Materials and Methods
4.1. Plant and Chemicals
4.2. Growth Environment of Ectropis obliqua
4.3. Analysis of PPO and PAL Activities
4.4. Biological Activity of Geometrid Larvae
4.5. Analysis of Midgut Proteinase Activities of Geometrid Larvae
4.6. Analysis of PAL and PPO Genes Expression
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Li, R.; Hu, Y.; Yang, Y.; Wang, W.; Shen, J. Changes of insect-resistance relative physiology indexes induced by low molecular compounds in tea plant. Southwest China J. Agric. Sci. 2016, 29, 1600–1606. [Google Scholar]
- Xin, Z.; Li, X.; Li, J.; Chen, Z.; Sun, X. Application of chemical elicitor (Z)-3-hexenol enhances direct and indirect plant defenses against tea geometrid Ectropis obliqua. BioControl 2016, 61, 1–12. [Google Scholar] [CrossRef]
- Gao, X.H.; Wan, X.C.; Yang, Y.Q.; Zhen, G.Y.; Shen, Q. Studies on the biological habits of Ectropis obliqua Prout. Plant Prot. 2007, 33, 110–113. [Google Scholar]
- Lu, Z.; Zhao, H.; Qiao, L. The Laboratory Test of Thirteen Insecticides against Ectropis Oblique Hypulina Wehrli. Pestic. Sci. Adm. 2012, 6, 22. [Google Scholar]
- Ye, G.-Y.; Xiao, Q.; Chen, M.; Chen, X.-X.; Yuan, Z.-J.; Stanley, D.W.; Hu, C. Tea: Biological control of insect and mite pests in China. Biol. Control 2014, 68, 73–91. [Google Scholar] [CrossRef]
- Goodspeed, D.; Chehab, E.W.; Minvenditti, A.; Braam, J.; Covington, M.F. Arabidopsis synchronizes jasmonate-mediated defense with insect circadian behavior. Proc. Natl. Acad. Sci. USA 2012, 109, 4674–4677. [Google Scholar] [CrossRef] [PubMed]
- Goellner, K.; Conrath, U. Priming: It’s all the world to induced disease resistance. Eur. J. Plant Pathol. 2008, 121, 233–242. [Google Scholar] [CrossRef]
- Gang, R.C.; Yan, C.; Fang, F.; Chao, D.C. Time and dosage effects of an endophytic fungal elicitor on the volatile oil production and physiology of Atractylodes lancea suspension cells. J. Med. Plants Res. 2012, 6, 5369–5376. [Google Scholar] [CrossRef]
- Elwakeil, N.E.; Volkmar, C.; Sallam, A.A. Jasmonic acid induces resistance to economically important insect pests in winter wheat. Pest Manag. Sci. 2010, 66, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Shi, H.; Chen, L.; Wang, X.; Lü, B.; Zhang, S.; Liang, Y.; Liu, R.; Qian, J.; Sun, W. Harpin-induced expression and transgenic overexpression of the phloem protein gene AtPP2-A1 in Arabidopsis repress phloem feeding of the green peach aphid Myzus persicae. BMC Plant Biol. 2011, 11, 11. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Song, F.F.; Xie, J.C. Resistance of jasmonic acid-mediated cotton seedlings against the relative growth rate of Helicoverpa Armigera. J. Northwest A F Univ. 2013, 5, 13. [Google Scholar]
- Gui, L.Y.; Chen, Z.M.; Liu, S.S. Effect of exogenous methyl jasmonate-induced tea volatiles on host-selection behavior of insects. J. Tea Sci. 2004, 24, 166–171. [Google Scholar]
- Gui, L.; Chen, Z.; Liu, S. Effect of exogenous MJA treatment of tea plants on the growth of geometrid larvae. Zhongguo Nongye Kexue 2005, 38, 302–307. [Google Scholar]
- Jin, M.; Baoyu, H. Probing behavior of the tea green leafhopper on different tea plant cultivars. Acta Ecol. Sin. 2007, 27, 3973–3982. [Google Scholar] [CrossRef]
- Yan, J.X.; Chi, D.F.; Zhang, Y.Q.; Pang, H.Y. Effects of JA and SA on the growth and development as well as defensive enzyme activity of Rosa rugosa. J. Beijing For. Univ. 2013, 35, 128–136. [Google Scholar]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Gonzales-Vigil, E.; Howe, G.A. Action of plant defensive enzymes in the insect midgut. In Induced Plant Resistance to Herbivory; Springer: Berlin, Germany, 2008; pp. 271–284. [Google Scholar]
- Constabel, C.P.; Barbehenn, R. Defensive roles of polyphenol oxidase in plants. In Induced Plant Resistance to Herbivory; Springer: Berlin, Germany, 2008; pp. 253–270. [Google Scholar]
- Agrawal, A.A.; Conner, J.K.; Johnson, M.T.J.; Wallsgrove, R. Ecological genetics of an induced plant defense against herbivores: additive genetic variance and costs of phenotypic plasticity. Evolution 2002, 56, 2206–2213. [Google Scholar] [CrossRef] [PubMed]
- Pajot, E.; Le Corre, D.; Silue, D. Phytogard® and DL-β-amino butyric acid (BABA) induce resistance to downy mildew (Bremia lactucae) in lettuce (Lactuca sativa L). Eur. J. Plant Pathol. 2001, 107, 861–869. [Google Scholar] [CrossRef]
- Hodge, S.; Pope, T.W.; Holaschke, M.; Powell, G. The effect of β-aminobutyric acid on the growth of herbivorous insects feeding on Brassicaceae. Ann. Appl. Biol. 2006, 148, 223–229. [Google Scholar] [CrossRef]
- Nombela, G.; Pascual, S.; Aviles, M.; Guillard, E.; Muñiz, M. Benzothiadiazole induces local resistance to Bemisia tabaci (Hemiptera: Aleyrodidae) in tomato plants. J. Econ. Entomol. 2005, 98, 2266–2271. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.L.; Cai, X.M.; Ma, C.L.; Wang, G.C.; Chen, Z.M. Kinetics of polyphenol oxidase activity in tea leaves induced by exogenous application of MeJA and mechanical damage. Acta Bot. Boreal-Occident. Sin. 2011, 9, 1805–1810. [Google Scholar]
- Yang, Z.-W.; Duan, X.-N.; Jin, S.; Li, X.-W.; Chen, Z.-M.; Ren, B.-Z.; Sun, X.-L. Regurgitant derived from the tea geometrid Ectropis obliqua suppresses wound-induced polyphenol oxidases activity in tea plants. J. Chem. Ecol. 2013, 39, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Zhao, W.L.; Gao, X.W. Communication between plants: induced resistance in poplar seedlings following herbivore infestation, mechanical wounding, and volatile treatment of the neighbors. Entomol. Exp. Appl. 2013, 149, 110–117. [Google Scholar] [CrossRef]
- Lou, Y.; Hu, L.; Li, J. Herbivore-induced defenses in rice and their potential application in rice planthopper management. In Rice Planthoppers; Springer: Berlin, Germany, 2015; pp. 91–115. [Google Scholar]
- Trotel-Aziz, P.; Couderchet, M.; Vernet, G.; Aziz, A. Chitosan stimulates defense reactions in grapevine leaves and inhibits development of Botrytis cinerea. Eur. J. Plant Pathol. 2006, 114, 405–413. [Google Scholar] [CrossRef]
- Thakur, M.; Sohal, B.S. Role of elicitors in inducing resistance in plants against pathogen infection: A review. ISRN Biochem. 2013, 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.S. Inducible direct plant defense against insect herbivores: a review. Insect Sci. 2008, 15, 101–114. [Google Scholar] [CrossRef]
- Wang, Y.-N.; Tang, L.; Hou, Y.; Wang, P.; Yang, H.; Wei, C.-L. Differential transcriptome analysis of leaves of tea plant (Camellia sinensis) provides comprehensive insights into the defense responses to Ectropis oblique attack using RNA-Seq. Funct. Integr. Genom. 2016, 16, 383–398. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.M.; Chandrashekar, A.; Venkatesh, Y.P. Eggplant polyphenol oxidase multigene family: Cloning, phylogeny, expression analyses and immunolocalization in response to wounding. Phytochemistry 2011, 72, 2275–2287. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Ling, L. Changes of Total Phenol Content and the Activities of PPO, POD and PAL during the Browning in Phalaenopsis Explant In Vitro. Acta Hortic. Sin. 2006, 33, 671–674. [Google Scholar]
- Worrall, D.; Holroyd, G.H.; Moore, J.P.; Glowacz, M.; Croft, P.; Taylor, J.E.; Paul, N.D.; Roberts, M.R. Treating seeds with activators of plant defence generates long-lasting priming of resistance to pests and pathogens. New Phytol. 2012, 193, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Medina, A.; Flors, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.J.; Pozo, M.J.; Ton, J.; van Dam, N.M.; Conrath, U. Recognizing plant defense priming. Trends Plant Sci. 2016, 21, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Tamayo, M.C.; Rufat, M.; Bravo, J.M.; San Segundo, B. Accumulation of a maize proteinase inhibitor in response to wounding and insect feeding, and characterization of its activity toward digestive proteinases of Spodoptera littoralis larvae. Planta 2000, 211, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.-Y.; Wu, G.; Wan, F.-H. Activities of digestive and detoxification enzymes in multiple generations of beet armyworm, Spodoptera exigua (Hübner), in response to transgenic Bt cotton. J. Pest Sci. 2010, 83, 453–460. [Google Scholar] [CrossRef]
- Zhao, A.; Zhan, E.; Sun, C.; Liu, T.; Li, Y. Effects of Cry1Ac toxin on proteases and carboxylesterase activities in the larvael midgut of Plutella xylostella. J. Plant Prot. 2017, 44, 713–720. [Google Scholar]
- Macedo, M.L.; das Graças Machado Freire, M. Insect digestive enzymes as a target for pest control. Invertebr. Surviv. J. 2011, 8, 190–198. [Google Scholar]
- Fürstenberg-Hägg, J.; Zagrobelny, M.; Bak, S. Plant defense against insect herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.-H.; Zhang, M.; Zhao, H.; Zhang, Y.; Wang, X.-X.; Guo, S.-S.; Zhang, Z.-F.; Liu, T.-X. Deciphering the mechanism of β-aminobutyric acid-induced resistance in wheat to the grain aphid, Sitobion avenae. PLoS ONE 2014, 9, e91768. [Google Scholar] [CrossRef] [PubMed]
- Ton, J.; Jakab, G.; Toquin, V.; Flors, V.; Iavicoli, A.; Maeder, M.N.; Métraux, J.-P.; Mauch-Mani, B. Dissecting the β-aminobutyric acid–induced priming phenomenon in Arabidopsis. Plant Cell 2005, 17, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Ton, J.; Mauch-Mani, B. β-amino-butyric acid-induced resistance against necrotrophic pathogens is based on ABA-dependent priming for callose. Plant J. 2004, 38, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Will, T.; van Bel, A.J.E. Physical and chemical interactions between aphids and plants. J. Exp. Bot. 2006, 57, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Tjallingii, W.F. Salivary secretions by aphids interacting with proteins of phloem wound responses. J. Exp. Bot. 2006, 57, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Zimmerli, L.; Métraux, J.-P.; Mauch-Mani, B. β-Aminobutyric acid-induced protection of Arabidopsis against the necrotrophic fungus Botrytis cinerea. Plant Physiol. 2001, 126, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Wang, C.F. Effects of BTH on Resistance Induction of Cucumber against Cladosporium cucumerinum. J. Northwest A F Univ. 2005, 33, 69–72. [Google Scholar]
- Hukkanen, A.T.; Kokko, H.I.; Buchala, A.J.; McDougall, G.J.; Stewart, D.; Kärenlampi, S.O.; Karjalainen, R.O. Benzothiadiazole induces the accumulation of phenolics and improves resistance to powdery mildew in strawberries. J. Agric. Food Chem. 2007, 55, 1862–1870. [Google Scholar] [CrossRef] [PubMed]
- Silue, D.; Pajot, E.; Cohen, Y. Induction of resistance to downy mildew (Peronospora parasitica) in cauliflower by DL-β-amino-n-butanoic acid (BABA). Plant Pathol. 2002, 51, 97–102. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Van Strien, E.A. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Fu, J.-Y. Molecular cloning and expression analysis of a putative sesquiterpene synthase gene from tea plant (Camellia sinensis). Acta Physiol. Plant. 2013, 35, 289–293. [Google Scholar] [CrossRef]
Sample Availability: Not Available. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Survival Rate (%) | Pupation Rate (%) | Emergence Rate (%) |
|---|---|---|---|
| CK | 95.56 ± 3.85 a | 82.22 ± 3.85 a | 43.38 ± 0.60 a |
| BTH-1 | 88.89 ± 1.02 a | 75.56 ± 1.02 a | 26.32 ± 0.07 a,b |
| BTH-2 | 86.67 ± 0 a | 73.33 ± 0 b | 39.39 ± 0.52 a |
| BTH-3 | 77.78 ± 1.02 b | 64.44 ± 1.02 b | 13.56 ± 0.42 b |
| BABA-1 | 88.89 ± 1.39 a | 75.56 ± 1.39 a | 35.26 ± 0.57 a,b |
| BABA-2 | 82.22 ± 1.39 b | 68.89 ± 1.39 b | 35.48 ± 0.24 a,b |
| BABA-3 | 84.44 ± 0.38 b | 71.11 ± 10.38 b | 15.15 ± 0.14 b |
| Treatment | 3rd Instar | 4th Instar | 5th Instar | Development Duration |
|---|---|---|---|---|
| CK | 2.67 ± 0.58 a | 2.00 ± 1.00 a | 2.33 ± 0.58 a,b | 15.33 ± 1.53 a |
| BTH-1 | 3.33 ± 0.58 a,b | 3.33 ± 0. 58 a,b,c | 2.67 ± 0.58 a,b | 19.00 ± 2.00 a,b |
| BTH-2 | 2.67 ± 1.15 a | 3.00 ± 1.15 a,b,c | 2.33 ± 0.58a,b | 17.33 ± 1.53 a |
| BTH-3 | 3.67 ± 0.58 a,b,c | 4.33 ± 0.58 c | 3.33 ± 0.58 b | 23.33 ± 2.52 c,d |
| BABA-1 | 2.67 ± 0.58 a | 2.67 ± 0.58 a,b | 2.00 ± 0 a | 19.00 ± 2.00 a,b |
| BABA-2 | 4.00 ± 0 b,c | 3.33 ± 0.58 a,b,c | 2.33 ± 0.58 a,b | 21.33 ± 1.15 b,c |
| BABA-3 | 4.67 ± 0.58 c | 4.00 ± 0 b,c | 4.33 ± 0.58 c | 26.33 ± 3.21 d |
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| PAL | GCGCGTTCTAACTAACTATGGG | TAGTGGATAAGACCGGCATTC |
| PPO | GGCTCTTCTTTCCGTTCC | CTAGATTCGGCTGGGTGC |
| β-actin | GCCATCTTTGATTGGAATGG | GGTGCCACAACCTTGATCTT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Yu, Y.; Li, Z.; Arkorful, E.; Yang, Y.; Liu, X.; Li, X.; Li, R. Benzothiadiazole and B-Aminobutyricacid Induce Resistance to Ectropis Obliqua in Tea Plants (Camellia Sinensis (L.) O. Kuntz). Molecules 2018, 23, 1290. https://doi.org/10.3390/molecules23061290
Li H, Yu Y, Li Z, Arkorful E, Yang Y, Liu X, Li X, Li R. Benzothiadiazole and B-Aminobutyricacid Induce Resistance to Ectropis Obliqua in Tea Plants (Camellia Sinensis (L.) O. Kuntz). Molecules. 2018; 23(6):1290. https://doi.org/10.3390/molecules23061290
Chicago/Turabian StyleLi, Huan, Ying Yu, Zhenzhen Li, Emmanuel Arkorful, Yiyang Yang, Xinqiu Liu, Xinghui Li, and Ronglin Li. 2018. "Benzothiadiazole and B-Aminobutyricacid Induce Resistance to Ectropis Obliqua in Tea Plants (Camellia Sinensis (L.) O. Kuntz)" Molecules 23, no. 6: 1290. https://doi.org/10.3390/molecules23061290