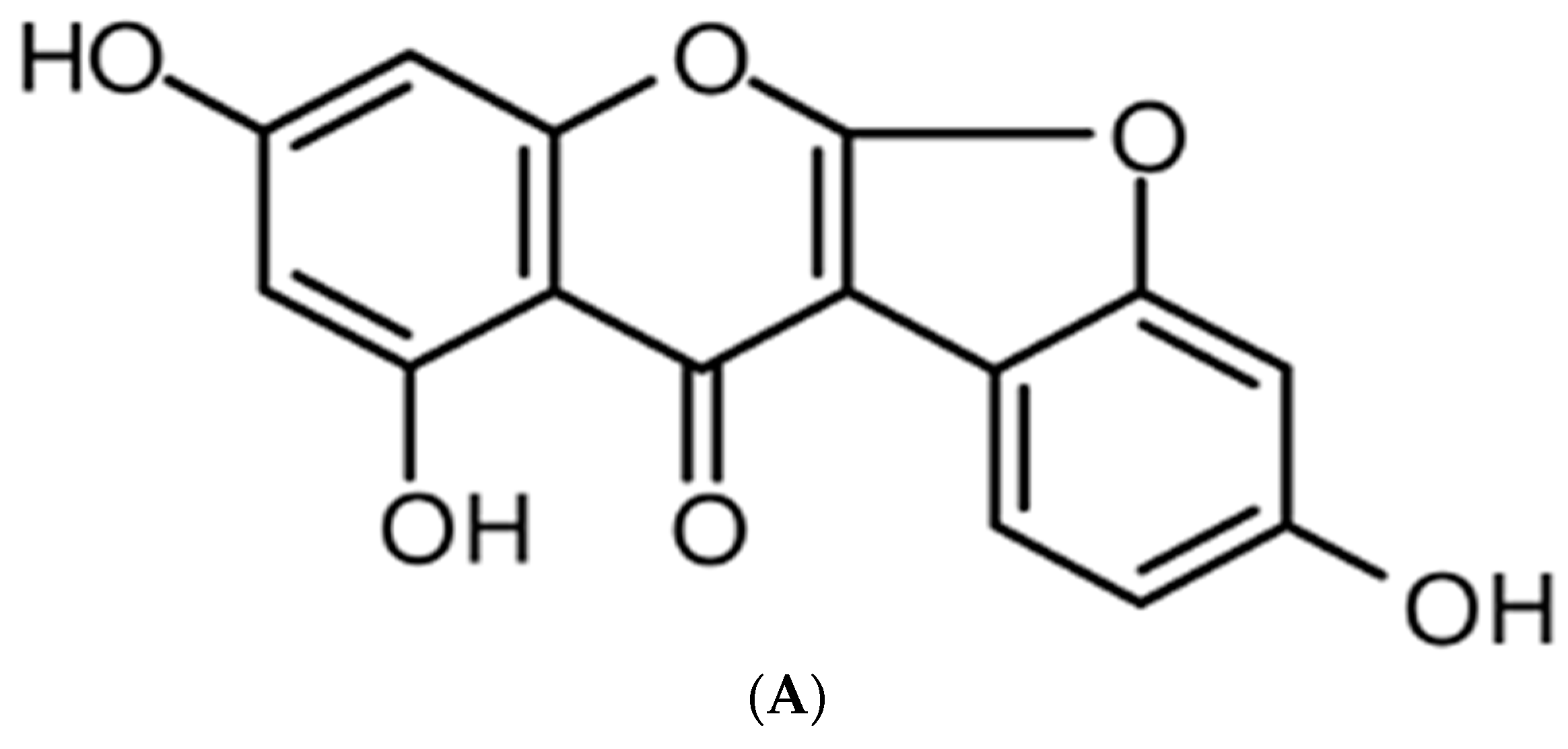
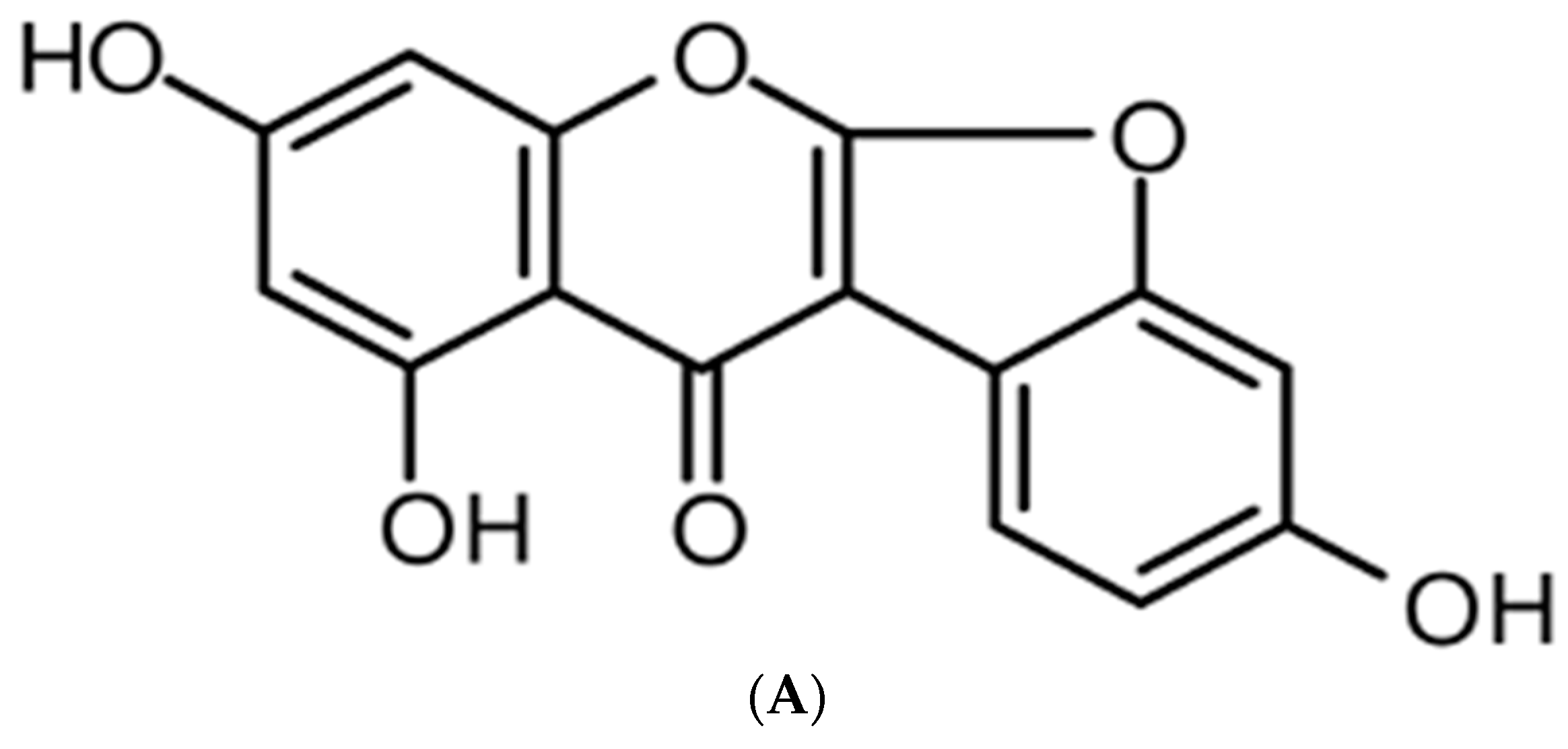
Anti-Inflammatory Effect of Lupinalbin A Isolated from Apios americana on Lipopolysaccharide-Treated RAW264.7 Cells


 and
and
Abstract
:1. Introduction
2. Results
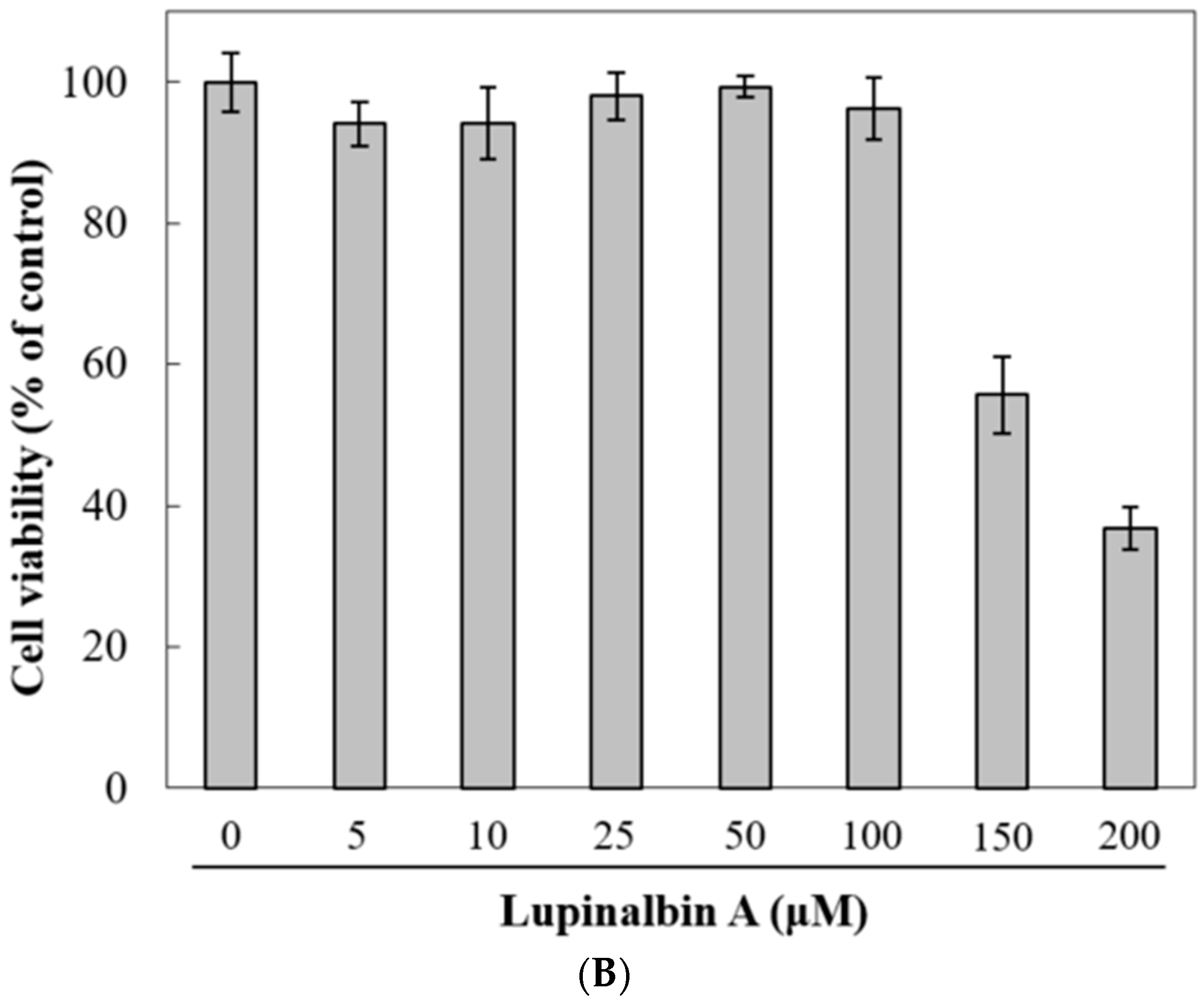
2.1. Effect of Lupinalbin A on Cell Cytotoxicity
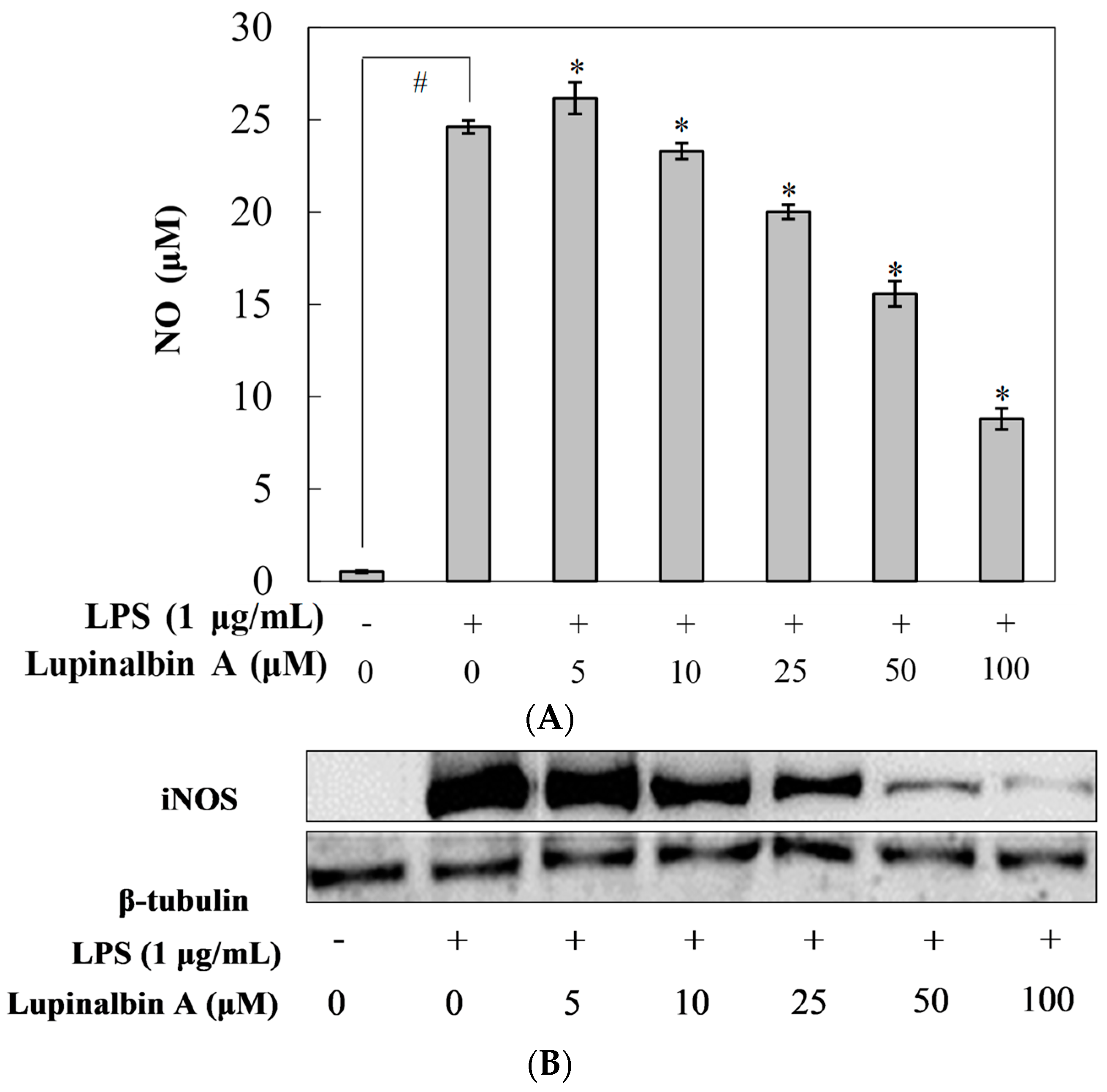
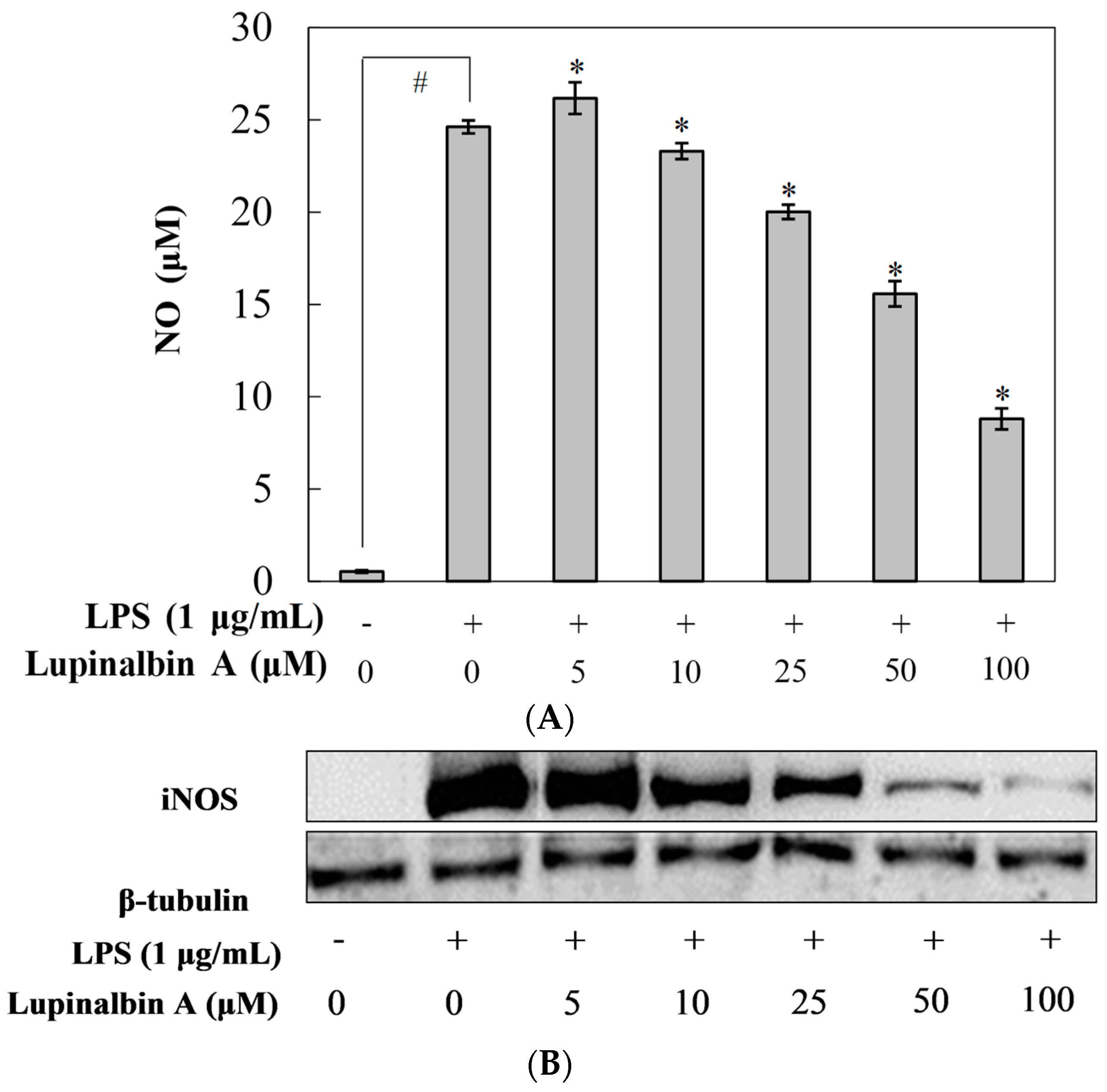
2.2. Effect of Lupinalbin A on LPS-Induced NO Production in RAW264.7 Cells
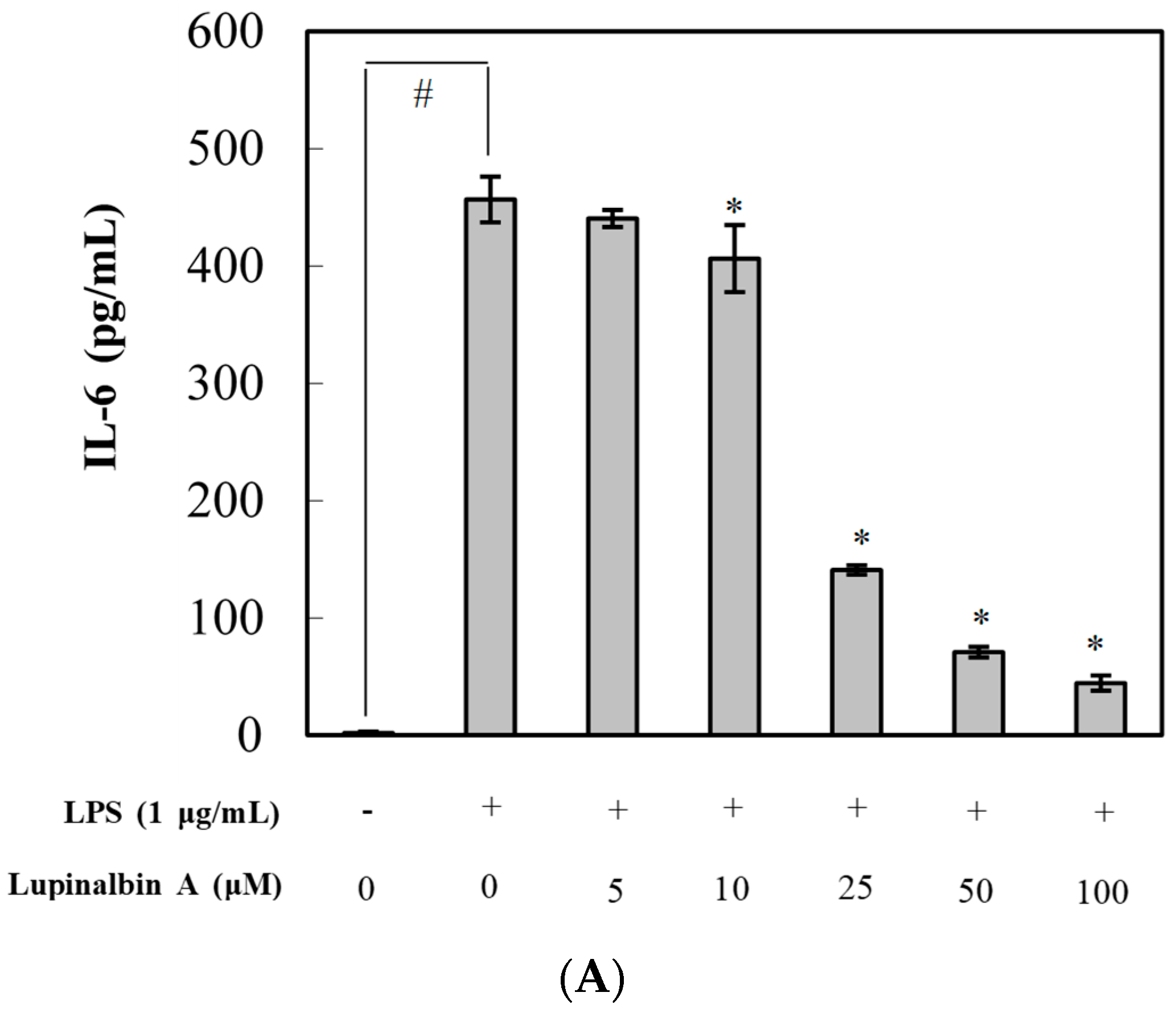
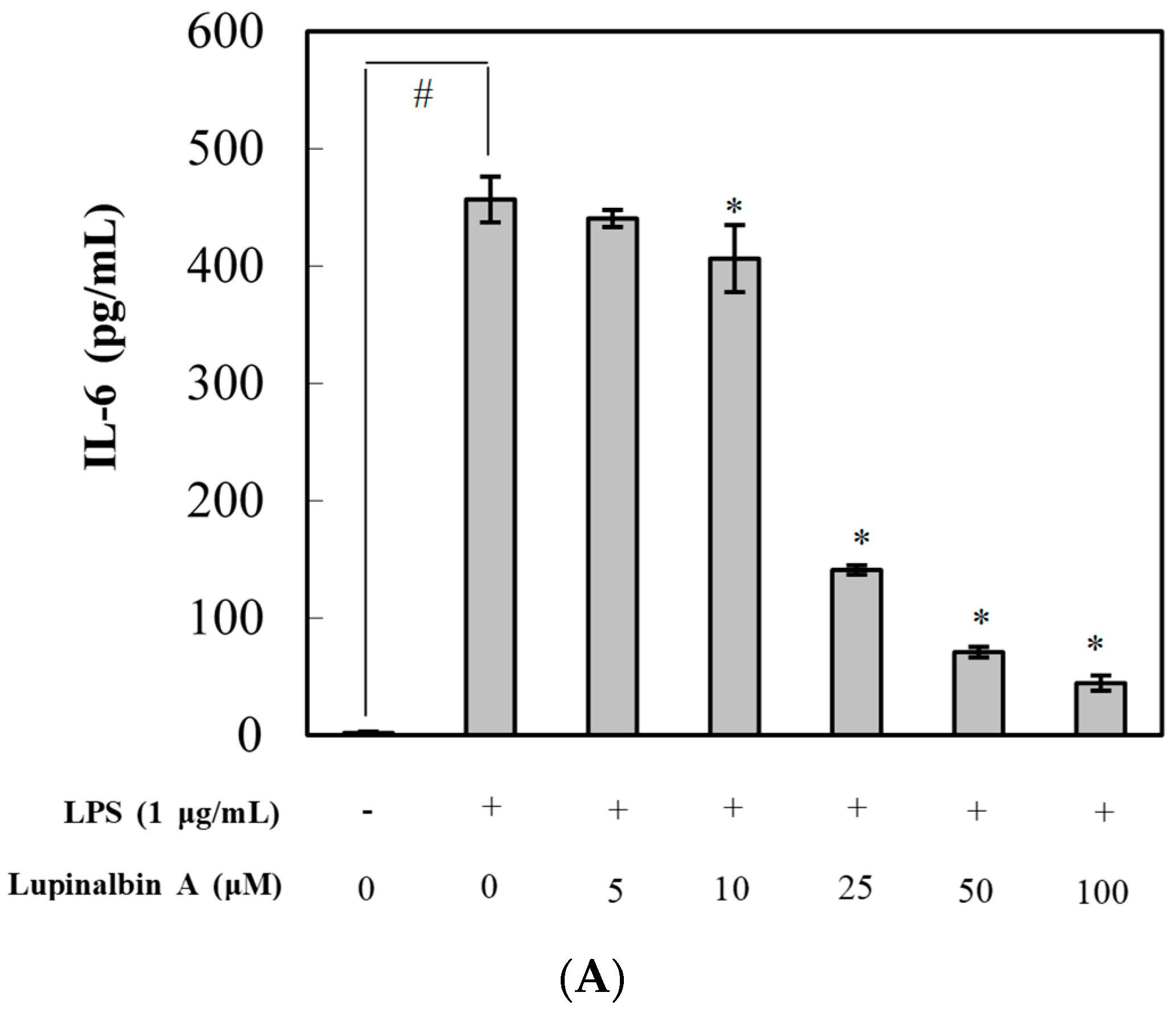
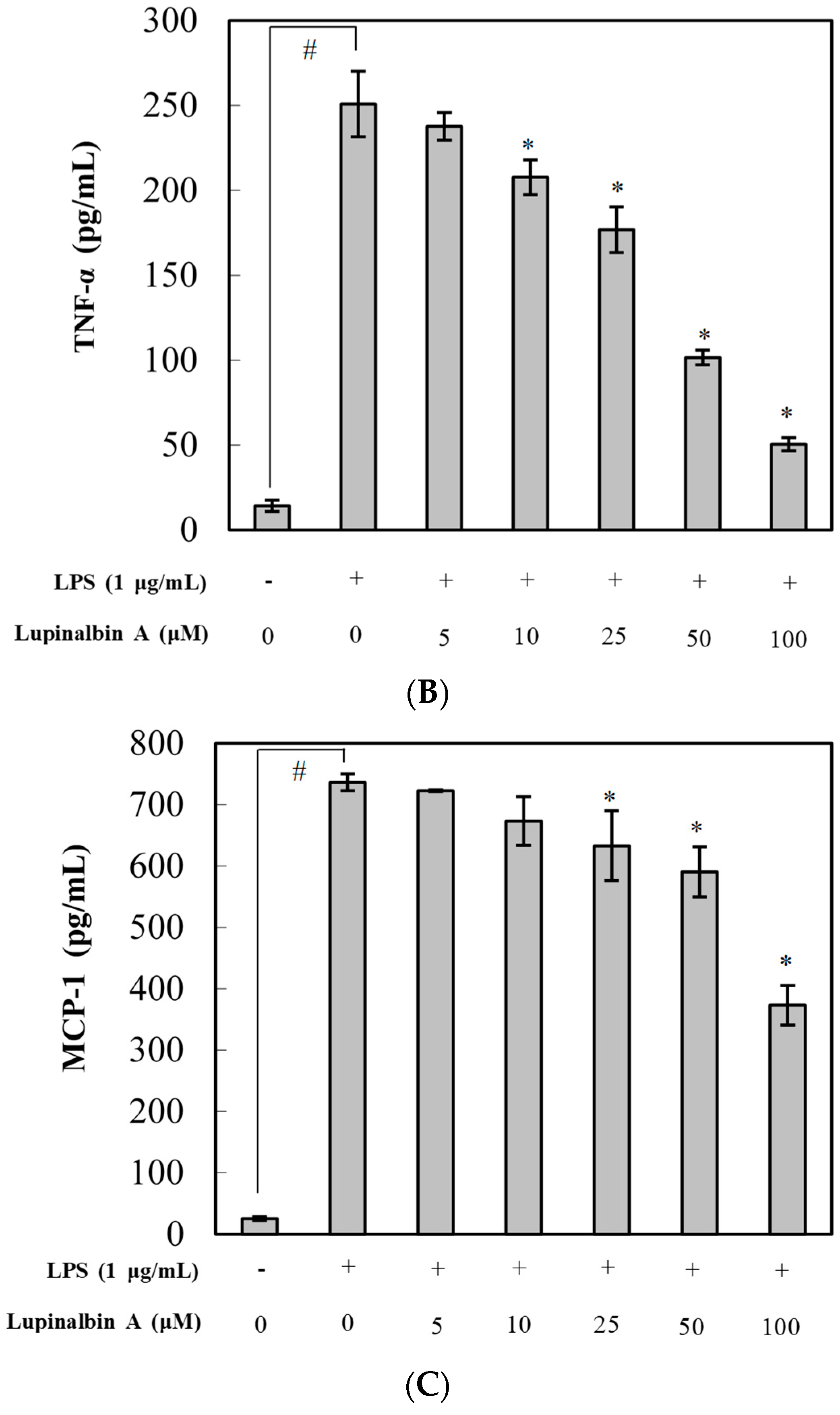
2.3. Effect of Lupinalbin A on Production of LPS-Induced Pro-Inflammatory Cytokines in RAW264.7 Cells
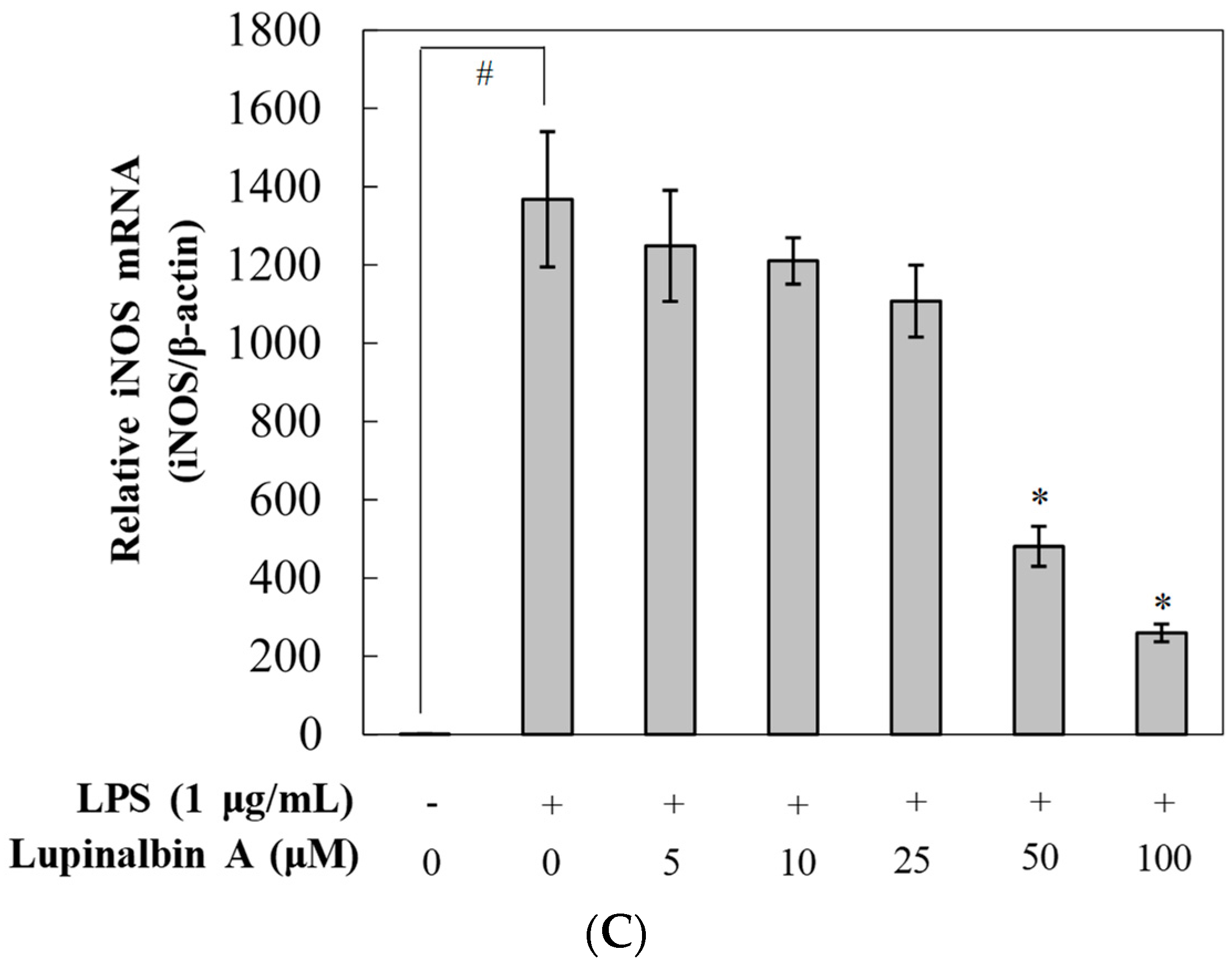
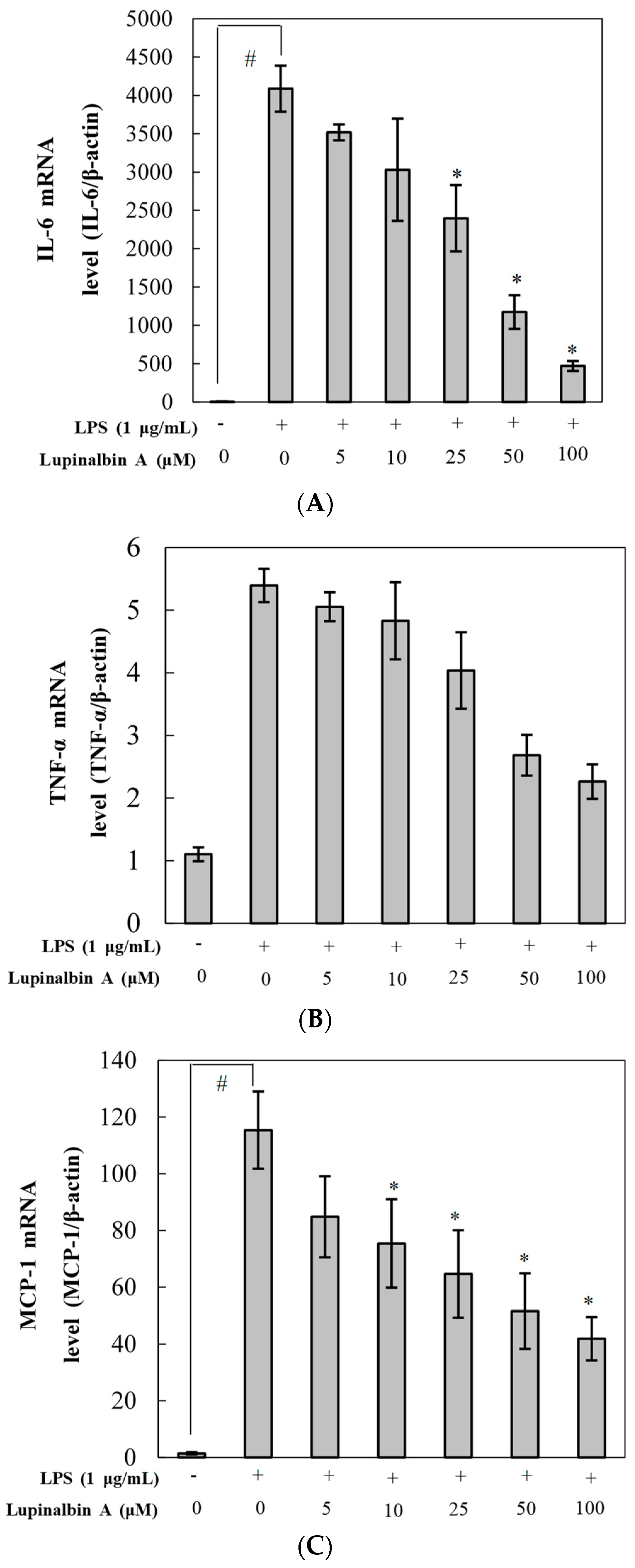
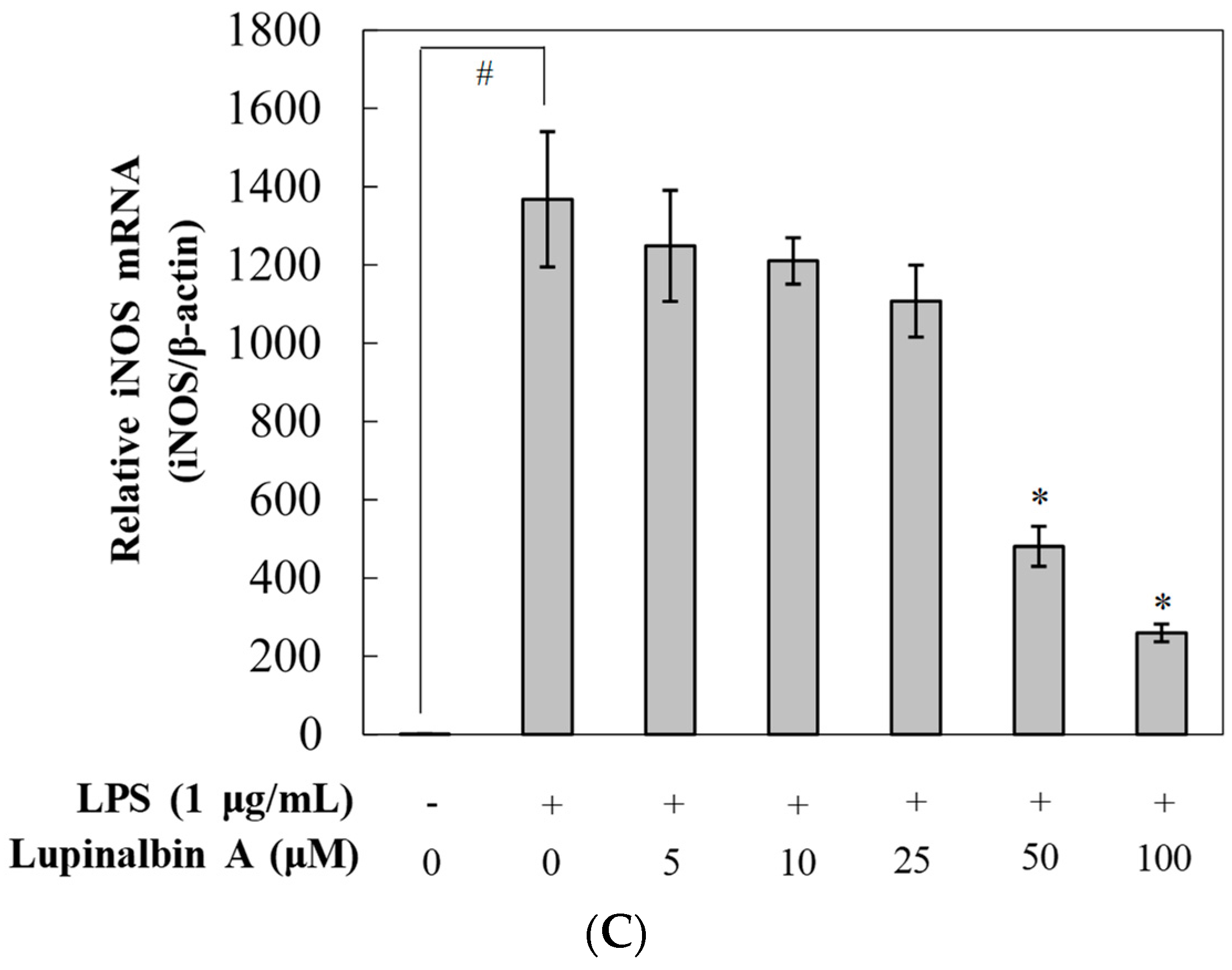
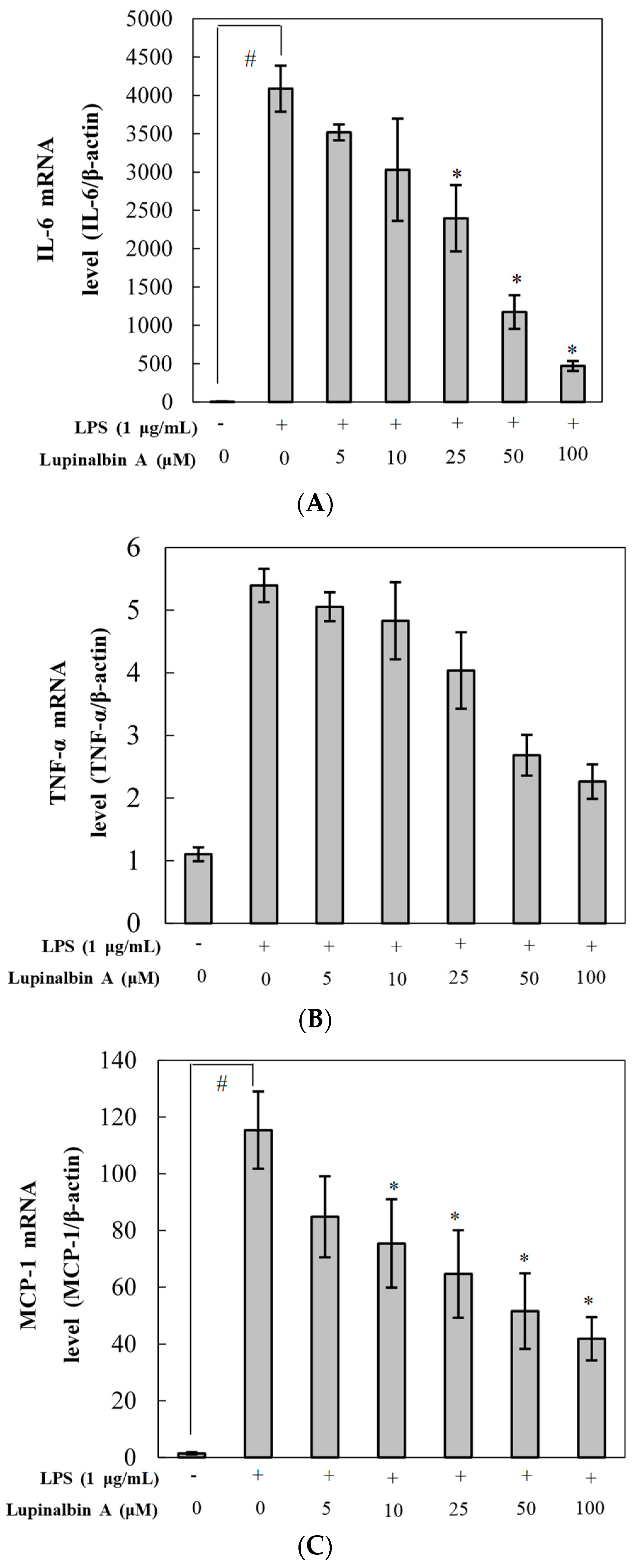
2.4. Effect of Lupinalbin A on LPS-Induced Expression of IL-6, TNF-α, and MCP-1 mRNA
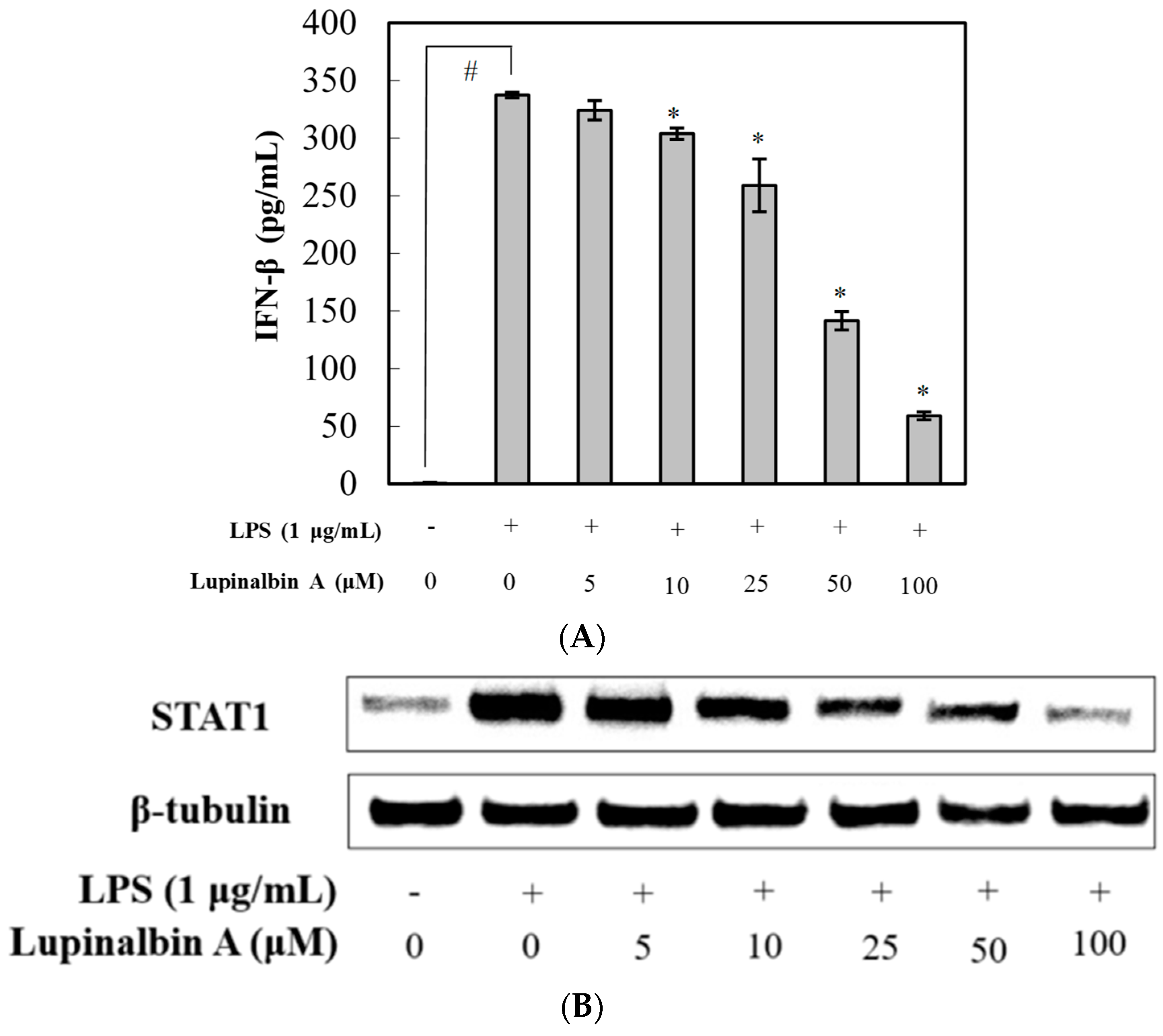
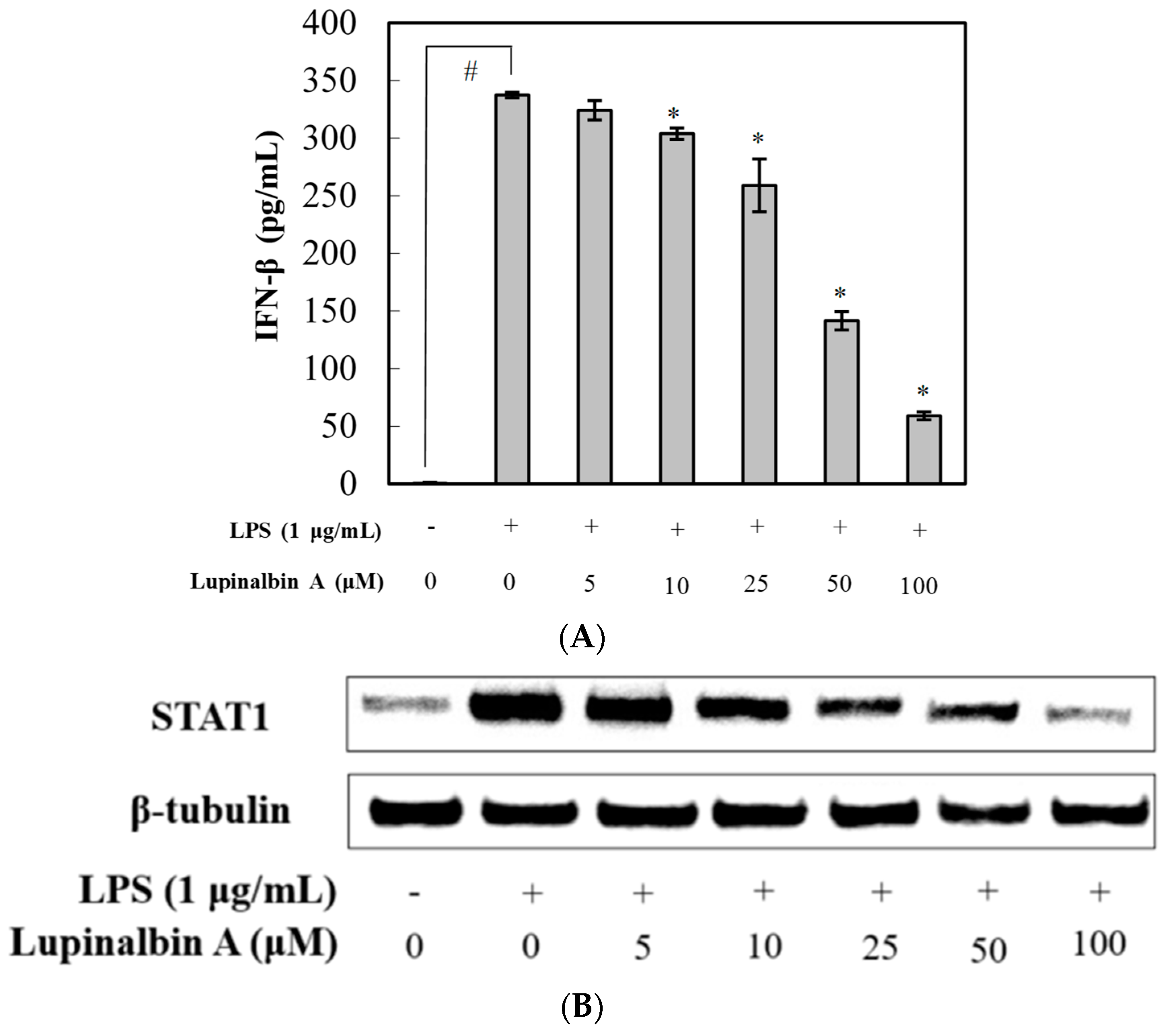
2.5. Effect of Lupinalbin A on LPS-Induced IFN-β Production and STAT1 Activation in RAW264.7 Cells
3. Discussion
4. Material and Methods
4.1. General Experimental Procedures
4.2. Chemicals
4.3. Plant Materials
4.4. Extraction, Fractionation, and Isolation of A. americana
4.5. Cell Culture
4.6. Cytotoxicity Assay
4.7. Measurement of NO Production
4.8. Western Blot
4.9. Measurement of IL-6, MCP-1, TNF-α and IFN-β Levels
4.10. Quantitative Real-Time PCR Analysis
4.11. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| ELISA | Enzyme-linked immunosorbent assay |
| eNOS | Endothelial NOS |
| FBS | Fetal bovine serum |
| iNOS | Inducible NOS |
| MCP | Monocyte chemoattractant protein |
| IFN | Interferon |
| LPS | Lipopolysaccharide |
| NO | Nitric oxide |
| NOS | NO synthase |
| nNOS | Neuronal NOS |
| TNF | Tumor necrosis factor |
References
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Ridker, P.M.; Maseri, A. Inflammation and atherosclerosis. Circulation 2002, 105, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Manzi, S.; Wasko, M.C. Inflammation-mediated rheumatic diseases and atherosclerosis. Ann. Rheum. Dis. 2000, 59, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Marletta, M.A. Nitric oxide synthase structure and mechanism. J. Biol. Chem. 1993, 268, 12231–12234. [Google Scholar] [PubMed]
- Rossol, M.; Heine, H.; Meusch, U.; Quandt, D.; Klein, C.; Sweet, M.J.; Hauschildt, S. LPS-induced cytokine production in human monocytes and macrophages. Crit. Rev. Immunol. 2011, 31, 379–446. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Takeuchi, O.; Fujita, T.; Inoue, J.; Muhlradt, P.F.; Sato, S. Lipopolysaccharide stimulates the MyD88-independent pathway and results in activation of IFN-regulatory factor 3 and the expression of subset of lipopolysaccharide-inducible genes. J. Immunol. 2001, 167, 5887–5894. [Google Scholar] [CrossRef] [PubMed]
- Piras, V.; Selvarajoo, K. Beyond MyD88 and TRIF pathways in Toll-like receptor signaling. Front. Immunol. 2014, 5, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, M.; Muroi, M.; Tanamoto, K.; Suzuki, T.; Azuma, H.; Ikeda, H. Molecular mechanisms of macrophage activation and deactivation by lipopolysaccharide: Roles of the receptor complex. Pharmacol. Ther. 2003, 100, 171–194. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.J.; Filla, M.B.; Fultz, M.J.; Vogel, S.N.; Russell, S.W.; Murphy, W.J. Autocrine/paracrine IFN-αβ mediates the lipopolysaccharide-induced activation of transcription factor Stat1α in mouse macrophages: Pivotal role of Stat1α in induction of the inducible nitric oxide synthase gene. J. Immunol. 1998, 161, 4803–4810. [Google Scholar] [PubMed]
- Nara, K.; Nihei, K.I.; Ogasawara, Y.; Koga, H.; Kato, Y. Novel isoflavone diglycoside in groundnut (Apios americana Medik). Food Chem. 2011, 124, 703–710. [Google Scholar] [CrossRef]
- Wilson, P.W.; Gorny, J.R.; Blackmon, W.J.; Rwynolds, B.D. Fatty acids on the American groundnut (Apios americana). J. Food Sci. 1986, 51, 1387–1388. [Google Scholar] [CrossRef]
- Wilson, P.W.; Pichardo, F.J.; Liuzzo, J.A.; Blackmon, W.J.; Rwynolds, B.D. Amino acids on the American groundnut (Apios americana). J. Food Sci. 1987, 52, 224–225. [Google Scholar] [CrossRef]
- Iwai, K.; Kuramoto, S.; Matsue, H. Suppressing effect of Apios americana on blood pressure in SHR and its active peptide. J. Clin. Biochem. Nutr. 2008, 43, 315–318. [Google Scholar]
- Cui, J.; Kim, Y.S.; Lee, E.B.; Kim, J.K.; Kang, T.H.; Jang, H.H. Immunostimulatory activity of apios tuber extract on RAW264.7 macrophage cells. J. Bacteriol. Virol. 2016, 46, 248–257. [Google Scholar] [CrossRef]
- Iwai, K.; Matsue, H. Ingestion of Apios americana Medikus tuber suppresses blood pressure and improves plasma lipids in spontaneously hypertensive rats. Nutr. Res. 2007, 27, 218–224. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, C.; Tang, S.; Yu, X.; Kouzuma, Y.; Yonekura, M. Effect of AATI, a Bowman-Birk type inhibitor from Apios americana, on proliferation of cancer cell lines. Food Chem. 2011, 128, 909–915. [Google Scholar] [CrossRef]
- Masilamani, M.; Wei, J.; Sampson, H.A. Regulation of the immune response by soybean isoflavones. Immunol Res. 2012, 54, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Hajto, T.; Hostanska, K.; Frei, K.; Rordorf, C.; Gabius, H.J. Increased secretion of tumor necrosis factor α, interleukin 1, and interleukin 6 by human mononuclear cells exposed to β-galactoside-specific lectin from clinically applied mistletoe extract. Cancer Res. 1990, 50, 3322–3326. [Google Scholar] [PubMed]
- Kenmochi, E.; Kabir, S.R.; Ogawa, T.; Naude, R.; Tateno, H.; Hirabayashi, J. Isolation and biochemical characterization of Apios tuber lectin. Molecules 2015, 24, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Khwaja, T.A.; Dias, C.B.; Pentecost, S. Recent studies on the anticancer activities of mistletoe (Viscum album) and its alkaloids. Oncology 1986, 43 (Suppl. 1), 42–50. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, H.Y.; Kang, S.Y.; Kim, Y.H.; Jin, C.H. Soluble epoxide hydrolase inhibitory activity of components isolated from Apios americana Medik. Molecules 2017, 22, 1432. [Google Scholar] [CrossRef] [PubMed]
- Song, S.H.; Lee, S.Y.; Cui, J.; Jang, H.H.; Kang, T.H.; Kim, J.K.; Kim, I.K.; Lee, D.K.; Choi, S.G.; Yoon, I.S.; et al. Apios americana Medik extract alleviates lung inflammation in influenza virus H1N1-and endotoxin-induced acute lung injury. J. Microbiol. Biotechnol. 2015, 25, 2146–2152. [Google Scholar]
- Takashima, M.; Nara, K.; Niki, E.; Yoshida, Y.; Hagihara, Y.; Stowe, M.; Horie, M. Evaluation of biological activities of a groundnut (Apios americana Medik) extract containing a novel isoflavone. Food Chem. 2013, 138, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Ateba, S.B.; Njamen, D.; Medjakovic, S.; Zehl, M.; Kaehlig, H.; Jungbauer, A.; Krenn, L. Lupinalbin A as the most potent estrogen receptor α- and aryl hydrocarbon receptor agonist in Eriosema laurentii de Wild (Leguminosae). BMC Complement. Altern. Med. 2014, 14, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Xu, Y.; Chang, J.; Wang, X.; Zhao, Y.; Yu, Z. Bioactives from stems and leaves of mung beans (Vigna radiata L.). J. Funct. Foods 2016, 25, 314–322. [Google Scholar] [CrossRef]
- Gagnon, H.; Tahara, S.; Bleichert, E.; Ibrahim, R.K. Separation of aglucones, glucosides and prenylated isoflavones by high-performance liquid chromatography. J. Chromatogr. 1992, 606, 255–259. [Google Scholar] [CrossRef]
- Kang, W.J.; Li, D.H.; Han, T.; Sun, L.; Fu, Y.B.; Sai, C.M.; Li, Z.L.; Hua, H.M. New chalcone and pterocarpoid derivatives from the roots of Flemingia philippinensis with antiproliferative activity and apoptosis-inducing property. Fitoterapia 2016, 112, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.F.; Liu, Y.W.; Kuo, Y.H.; Shen, C.C.; Chiou, W.F.; Chen, C.C. Flavonoids and bis-coumarin from the tubers of Apios taiwanianus. Phytochem. Lett. 2016, 15, 164–167. [Google Scholar] [CrossRef]
- Sun, F.; Li, Q.; Xu, J. Chemical composition of roots Flemingia philippinensis and their inhibitory kinetics on aromatase. Chem. Biodivers. 2017, 14, e1600193. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Lekontseva, O.; Davidge, S.T. Estrogen is a modulator of vascular inflammation. IUBMB Life 2008, 60, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Chen, J.R.; Pais, E.M.A.; Wang, T.Y.; Miao, L.; Li, L.; Li, L.Y.; Qiu, F.; Hu, L.M.; Gao, X.M.; et al. Oxyresveratrol is a phytoestrogen exerting anti-inflammatory effects through NF-κB and estrogen receptor signaling. Inflammation 2017, 40, 1285–1296. [Google Scholar] [CrossRef] [PubMed]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Kopf, M.; Bachmann, M.F.; Marsland, B.J. Averting inflammation by targeting the cytokine environment. Nat. Rev. Drug Discov. 2010, 9, 703–718. [Google Scholar] [CrossRef] [PubMed]
- Song, S.M.; Ham, Y.M.; Ko, Y.J.; Ko, E.Y.; Oh, D.J.; Kim, C.S.; Kim, D.K.; Kim, K.N.; Yoon, W.J. Anti-inflammatory activites of the products of supercritical fluid extraction from Litsea japonica fruit in RAW264.7 cells. J. Funct. Foods 2016, 22, 44–51. [Google Scholar] [CrossRef]
- Jin, C.H.; Lee, H.J.; Park, Y.D.; Choi, D.S.; Kim, D.S.; Kang, S.Y.; Seo, K.I.; Jeong, I.Y. Isoegomaketone inhibits lipopolysaccharide-induced nitric oxide production in RAW264.7 macrophages through the heme oxygenase-1 induction and inhibition of the interferon-β-STAT-1 pathway. J. Agric. Food Chem. 2010, 58, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Canesi, L.; Borghi, C.; Ciacci, C.; Fabbri, R.; Vergani, L.; Gallo, G. Bisphenol-A alters gene expression and functional parameters in molluscan hepatopancreas. Mol. Cell. Endocrinol. 2007, 276, 36–44. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′ to 3′) |
|---|---|
| iNOS | Forward: TCCTACACACCAAACTGTGTGC |
| Reverse: CTCCAATCTCTGCCTATCCGTCTC | |
| IL-6 | Forward: GTTCTCTGGGAAATCGTGGAA |
| Reverse: GCAAGTCCATCATCGTTGTTC | |
| TNF-α | Forward: GCCACCACGCTCTTCTGTCTAC |
| Reverse: GGGCTACAGGCTTGTCACTCG | |
| MCP-1 | Forward: GCATCTGCCCTAAGGTCTTCA |
| Reverse: GTGGAAAAGGTAGTGGATGCATT | |
| β-actin | Forward: TGAGAGGGAAATCGTGCGTGAC |
| Reverse: GCTCGTTGCCAATAGTGATGACC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-Y.; Kim, J.H.; So, Y.; Kang, S.-Y.; Jeong, H.G.; Jin, C.H. Anti-Inflammatory Effect of Lupinalbin A Isolated from Apios americana on Lipopolysaccharide-Treated RAW264.7 Cells. Molecules 2018, 23, 583. https://doi.org/10.3390/molecules23030583
Kim H-Y, Kim JH, So Y, Kang S-Y, Jeong HG, Jin CH. Anti-Inflammatory Effect of Lupinalbin A Isolated from Apios americana on Lipopolysaccharide-Treated RAW264.7 Cells. Molecules. 2018; 23(3):583. https://doi.org/10.3390/molecules23030583
Chicago/Turabian StyleKim, Hyo-Young, Jang Hoon Kim, YangKang So, Si-Yong Kang, Hye Gwang Jeong, and Chang Hyun Jin. 2018. "Anti-Inflammatory Effect of Lupinalbin A Isolated from Apios americana on Lipopolysaccharide-Treated RAW264.7 Cells" Molecules 23, no. 3: 583. https://doi.org/10.3390/molecules23030583