Therapeutic Potential of Oridonin and Its Analogs: From Anticancer and Antiinflammation to Neuroprotection


1
Chemical Biology Program, Department of Pharmacology and Toxicology, University of Texas Medical Branch, Galveston, TX 77555, USA
2
Department of Clinical Cancer Prevention, Division of Cancer Prevention and Population Sciences, The University of Texas MD Anderson Cancer Center, Houston, TX 77030, USA
*
Author to whom correspondence should be addressed.
Molecules 2018, 23(2), 474; https://doi.org/10.3390/molecules23020474
Submission received: 10 January 2018
/
Revised: 13 February 2018
/
Accepted: 21 February 2018
/
Published: 22 February 2018
(This article belongs to the Special Issue Neuroprotective Agents)
Abstract
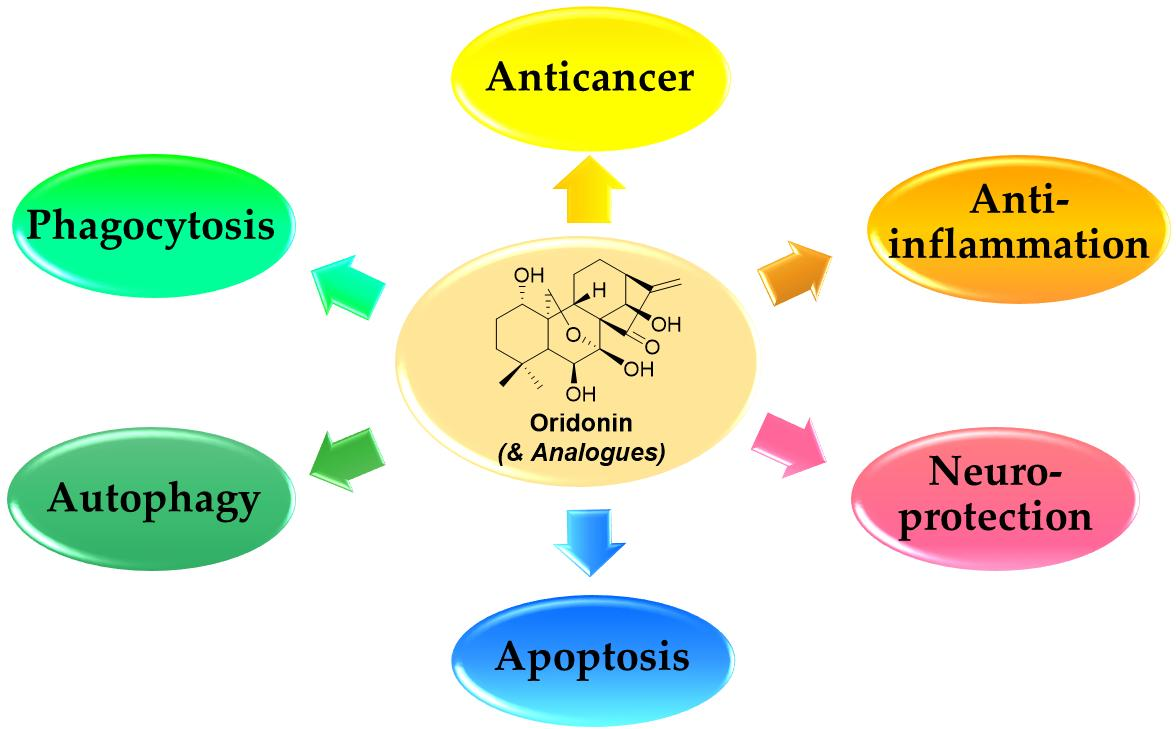
:Oridonin, a diterpenoid natural product commonly used in East Asian herbal medicine, is garnering increased attention in the biomedical community due to its extensive biological activities that include antitumor, anti-inflammatory, antimicrobial, hepatic fibrosis prevention, and neurological effects. Over the past decade, significant progress has been made in structure activity relationship and mechanism of action studies of oridonin for the treatment of cancer and other diseases. This review provides a brief summary on oridonin and its analogs in cancer drug discovery and antiinflammation and highlights its emerging therapeutic potential in neuroprotection applications.
1. Introduction
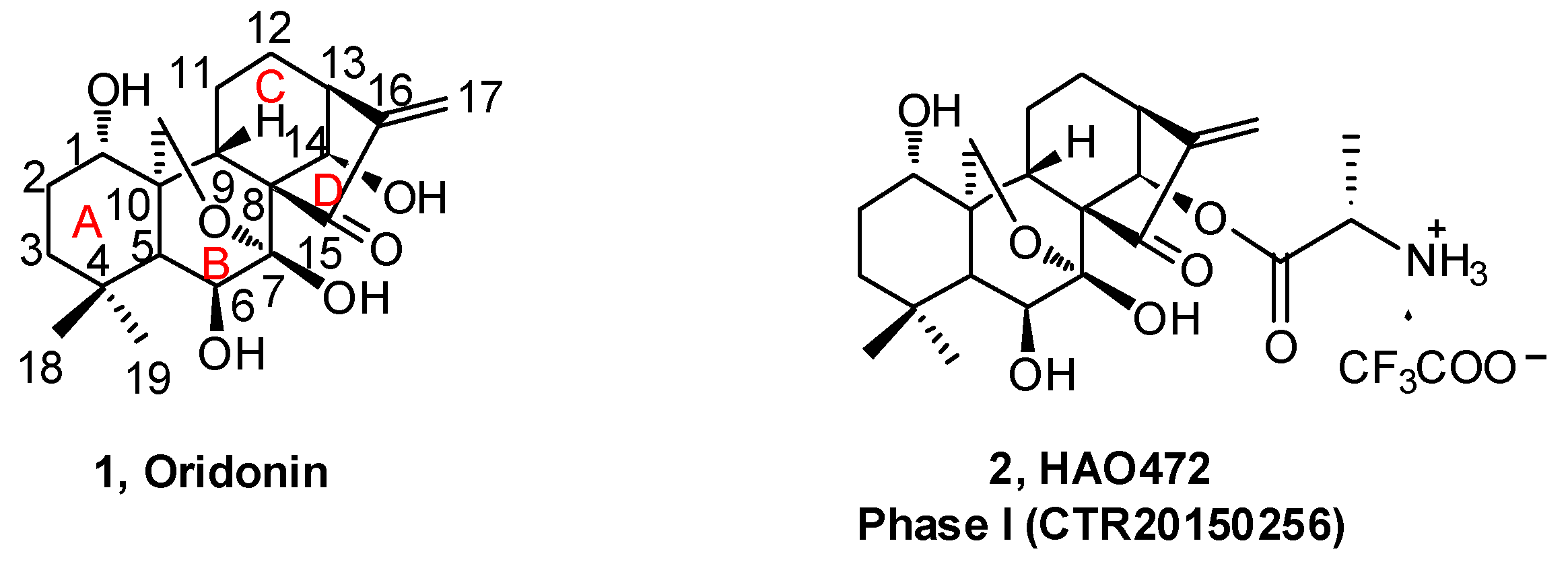
Natural products derived from animals, plants, and microbes have played an important role in the treatment of human diseases since the dawn of medicine. Natural products have high chemical diversity, bio-compatible characteristics, and other molecular properties that make them advantageous as lead scaffolds for drug discovery [1,2,3,4]. A detailed analysis of first-in-class drugs approved by the US Food and Drug Administration (FDA) from 1999 to 2013 revealed 31 (28%) of those drugs (112) were natural products or their derivatives [5]. Rabdosia rubescens, a herbal plant also known as Donglingcao, was used in East Asian traditional medicine for the treatment of inflammation and cancer [6]. Oridonin (1, Figure 1), an ent-kaurane diterpenoid isolated from Rabdosia rubenscens, was first identified as an antitumor compound in 1967 [7,8]. It has since attracted considerable attention due to its various pharmacological and physiological properties including antitumor [9,10,11,12,13,14,15,16,17], anti-inflammatory [18,19,20], antimicrobial [21], and hepatic fibrosis prevention actions [22,23,24,25], and its effects in the central nervous system (CNS) [26,27,28]. Over the past decade, significant progress has been made in structural optimization and mechanism of action studies of oridonin for the treatment of cancer and other diseases [29,30,31]. For example, Hengrui Medicine Co. Ltd. (Lianyungang, China) was recently given approval to advance HAO472 [32] (2, l-alanine-(14-oridonin)ester trifluoroacetate, Figure 1) into a Phase I human clinical trial (CTR20150246; www.chinadrugtrails.org.cn) in China for the treatment of acute myelogenous leukemia. In this review, we will first provide a summary of oridonin, its analogs, and their promising potential in cancer drug discovery and antiinflammation, and second, we will highlight its emerging therapeutic potential for neuroprotection. To our knowledge, this will be the first review article in the field that discusses the oridonin class of compounds as potential neurotherapeutics.
2. Oridonin and Its Analogs for Cancer Drug Discovery
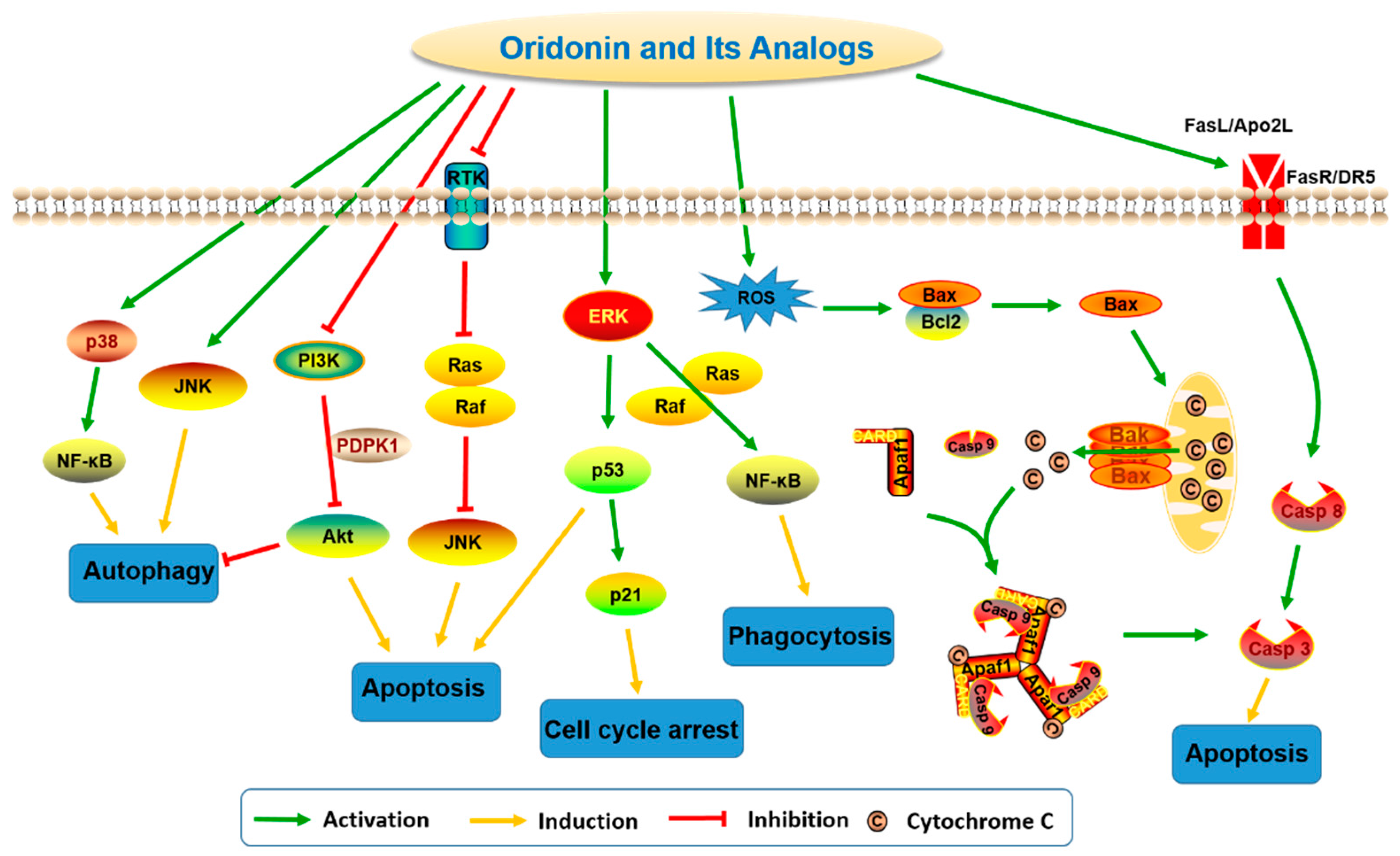
The antitumor activity of oridonin has been widely investigated and evidence suggests that oridonin may effectively inhibit the proliferation of multiple cancer cell types, including human breast cancer [9], gastric cancer [10], leukemia [11], gallbladder cancer [12], cervical carcinoma [13], and hepatocellular carcinoma [14]. Previous studies have revealed mechanisms by which oridonin can trigger autophagy, enhance phagocytosis, arrest cell cycle progression, and promote apoptosis by modulation of relevant signaling pathways (Figure 2) associated with the regulation of intracellular reactive oxygen species (ROS), Bcl-2/Bax, p53/p21, JNK, nuclear factor-kappa B (NF-κB), MAPK, PI3K, and fatty acid synthesis pathways [30,31,33].
In a human prostate cell line, DU-145, oridonin upregulates p53 and Bax and downregulates Bcl-2 expression in a dose-dependent manner [34]. In Hela cells, oridonin-induced autophagy is negatively regulated by Ras but positively regulated by p38 and JNK MAPKs [13]. Additionally, in HepG2 cells, oridonin was reported to increase the expression levels of p-JNK, p-p38, p-p53, and p21 and elevate the level of cyclin B1/p-Cdc2 (Tyr15) complex, which results in G2/M cell cycle arrest and apoptosis through MAPK and p53 pathways [35]. Oridonin also induces apoptosis via inhibiting PTK-mediated Ras-Raf-JNK [36] and PI3K-Akt [37] survival pathways in L929 and cervical carcinoma Hela cells, respectively. Moreover, oridonin can trigger apoptosis through activating both classic extrinsic pathways, such as Fas/FasL and Apo2L/DR5-mediated signaling pathways, and mitochondrial-mediated intrinsic pathways in several cancer cells [38,39,40,41]. In U937 cells, oridonin was reported to activate NF-κB via Ras/Raf1/ERK signaling pathway-dependent IκBα degradation and subsequently regulate oridonin-enhanced phagocytosis [42]. In colorectal cancer cells, oridonin has been shown to increase the intracellular hydrogen peroxide level and reduce the glutathione content in a dose-dependent manner [43]. It was also reported that oridonin induces a rapid and significant generation of ROS in L929 cells and subsequently upregulates the expression of phospho-p53 and increases expression ratio of Bax/Bcl-2 [44]. Modulations of these pathways in different cell models may explain the broad range of anticancer activities of oridonin.
Although oridonin has a unique, relatively safe, and extensive anticancer profile, its clinical development for cancer therapy has historically been hindered by its moderate potency, limited aqueous solubility, and poor bioavailability. To overcome these obstacles and yield oridonin analogs with increased druglikeness, iterative medicinal chemistry efforts have been made by multiple research groups and a portion of that work is discussed herein. Over the past several years, our group has synthesized a series of oridonin derivatives primarily focused on A-ring system modifications.
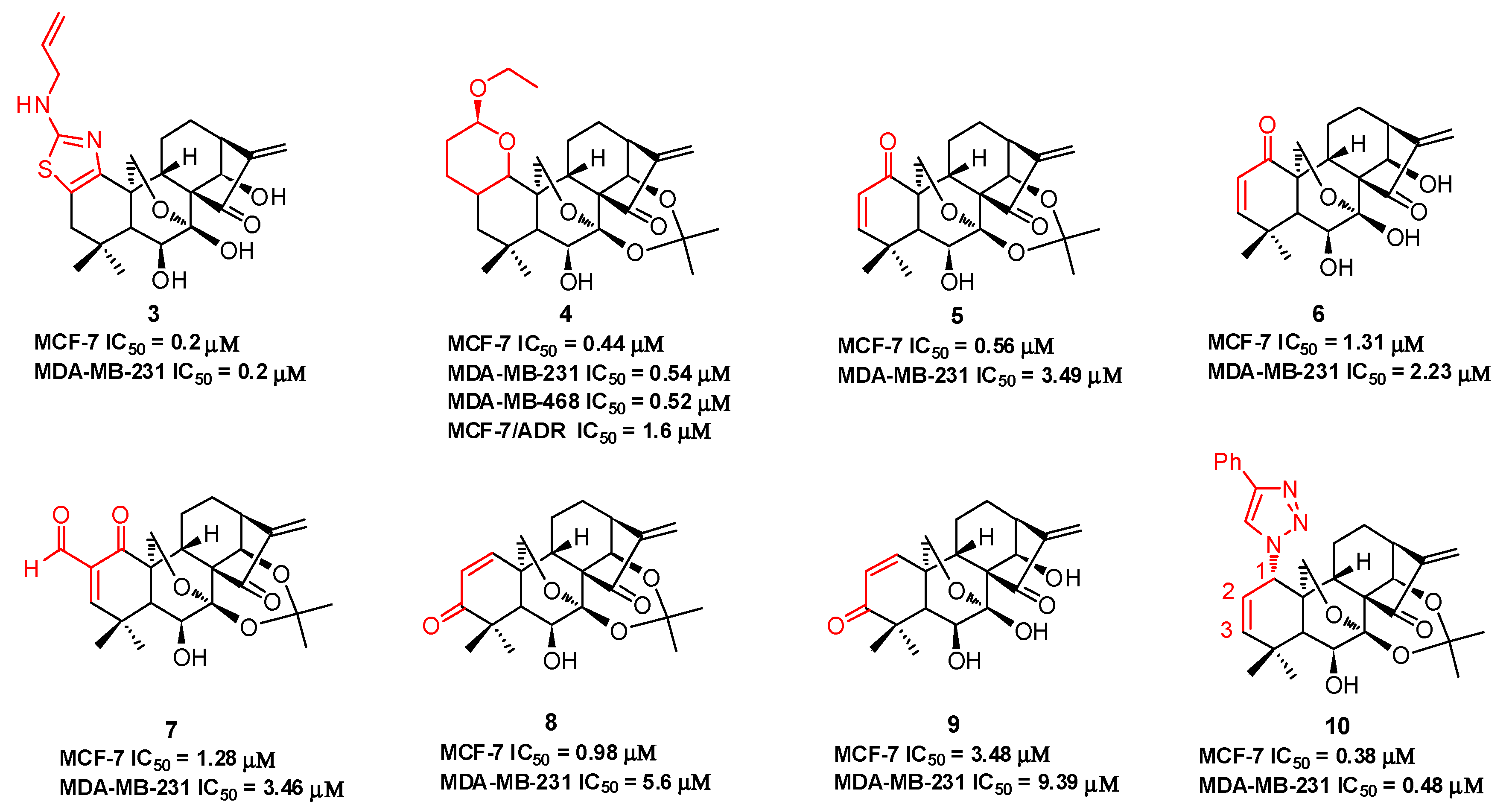
Compound 3 (Figure 3), with an N-allyl substituted thiazole moiety, exhibits potent antiproliferative activities against human breast cancer MCF-7 (IC50 = 0.2 μM) and MDA-MB-231 (IC50 = 0.2 μM) cells, which are approximately 33-fold and 147-fold more potent than oridonin, respectively [45]. The aqueous solubility of 3 has been significantly improved with a saturated concentration of 42.4 mg/mL, which is approximately 32-fold better than that of oridonin (1.29 mg/mL). Moreover, 3 significantly suppresses MDA-MB-231 xenograft tumor growth in vivo (5 mg/kg, ip, tumor growth inhibition >66%), while oridonin shows no significant efficacy at the same dose. Among the dihydropyran-fused derivatives, 4 shows the highest inhibition potency against MCF-7 (IC50 = 0.44 μM), MDA-MB-231 (IC50 = 0.54 μM), and MDA-MB-468 (IC50 = 0.52 μM) cell lines and an improved ability to overcome chemoresistance in a MCF-7/ADR cell line (IC50 = 1.6 μM) [46]. Our group also synthesized a series of dienone derivatives of oridonin with additional α,β-unsaturated ketone system diversely installed in the A-ring [47]. Dienone analogues 5–9 display significant antiproliferative effects relative to oridonin against MCF-7 and MDA-MB-231 cells with low micromolar to submicromolar potency. Compared to oridonin, 8 shows lower toxicity in normal mammary epithelial cells and increased antitumor efficacy at a dose of 5.0 mg/kg (ip, growth inhibition >55%) with no significant loss of body weight in an MDA-MB-231 xenograft tumor model. Our group developed efficient and concise synthetic approaches to rapidly and diversely introduce azide functionalities at the C-1, C-2, or C-3 positions of oridonin in a highly regio- and stereospecific manner. Subsequent functionalization of these azides through click chemistry yielded triazole derivatives. These derivatives with 1,2,3-triazole installed in the A-ring system exhibit significantly improved activities against breast cancer cells compared to oridonin. Among them, 1-triazole derivative 10 displays the most potent inhibitory activities against MCF-7 (IC50 = 0.38 μM) and MDA-MB-231 (IC50 = 0.48 μM) cell lines [48].
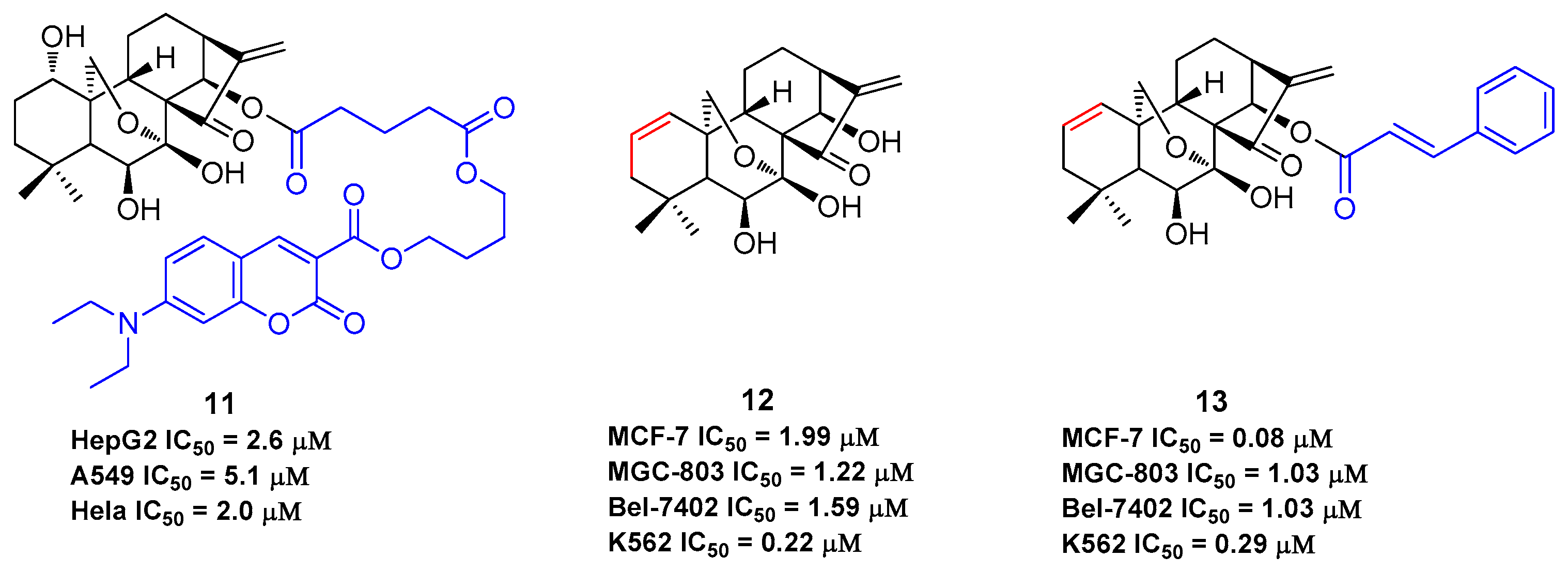
The B-ring is inert due to the low reactivity of the 7-hydroxy group and the hydrogen bond of 6-hydroxy group with the 15-carbonyl group. The α,β-unsaturated ketone in the D-ring is the active pharmacophore of oridonin, and studies have shown that reduction or opening will significantly reduce the antiproliferative effect of oridonin [49,50,51]. Alternatively, the esterification of the hydroxyl in the C-ring is an efficient way to enhance the antiproliferative activity of oridonin other than the modifications on A-ring. HAO472 (Figure 2) was designed with an alanine ester trifluoroacetate at the C-14 position to improve its aqueous solubility (i.e., 165 mg/mL). HAO472 was said to maintain the anticancer activities of oridonin (data not disclosed), while also being less likely to cause vascular injury [32,52]. Thus, in China, HAO472 has been advanced into Phase I human clinical trials for the treatment of acute myelogenous leukemia (80–320 mg/d, iv, CTR20150246). Xu and colleagues designed and synthesized a fluorescent oridonin probe 11 (Figure 4) using a linker to connect the 14-hydroxyl group of oridonin with a coumarin moiety [49]. When tested, 11 exhibited more potent antiproliferative activities compared to oridonin in HepG2 (IC50 = 2.6 μM), A549 (IC50 = 5.1 μM) and Hela (IC50 = 2.0 μM) cell lines. 11 was used to confirm that a mitochondrial pathway is involved in oridonin-mediated apoptosis and that cytochrome C plays an important role in the oridonin-mediated apoptotic process. Compound 13, bearing 1-ene and a trans-cinnamic acid moiety on the 14-position designed and synthesized from compound 12, is 200-fold (IC50 = 0.08 μM) more potent than oridonin against MCF-7 cancer cells [53]. 13 significantly decreased tumor volume and reduced tumor weight by 69.8% at a dose of 20 mg/kg/day (iv) in an MCF-7 breast cancer xenograft nude mice model, which was greater than that of the positive control, cyclophosphamide (64.6%).
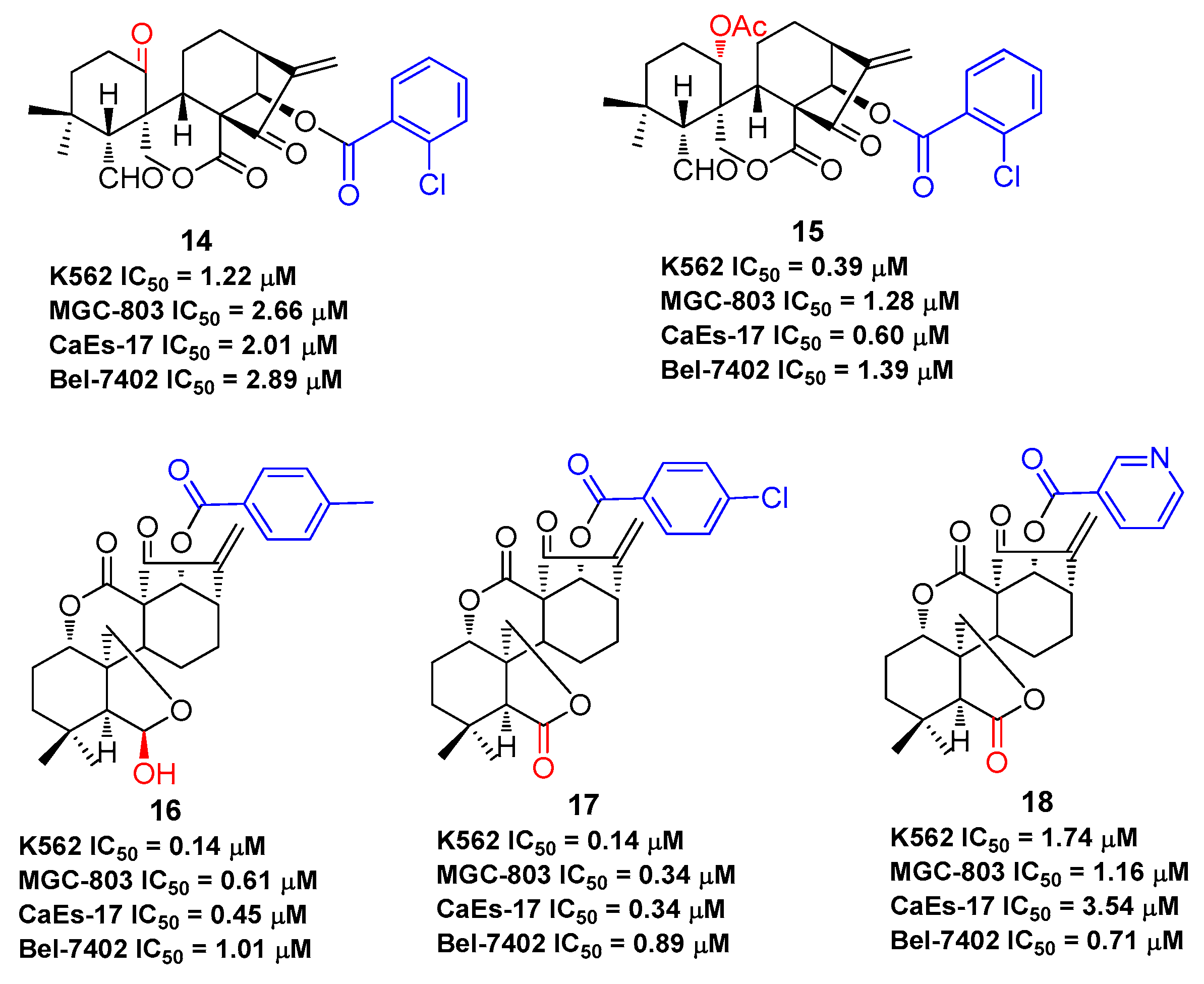
As oridonin is abundant in natural sources and is commercially available, it can also be used as an advantageous starting material to semi-synthesize other types of diterpenoid derivatives that are otherwise rare [54,55,56,57,58,59], such as spirolactone-type diterpenoid and enmein-type diterpenoid derivatives. Spirolactone-type diterpenoid derivatives 14–15 [54,55] and enmein-type diterpenoid derivatives 16–18 [56,57], synthesized from oridonin by oxidative rearrangements around C-6 and C-7 positions, showed improved antiproliferative activities against a panel of human cancer cell lines (Figure 5). Administration of salts of water-soluble compound 18 at a dose of 40 mg/kg was found to exhibit greater anti-gastric cancer effects (ip, growth inhibition = 64.8%) when compared to oridonin (ip, growth inhibition = 37.3%) in mice [57]. The synthesis of these ent-kaurane diterpenoid derivatives with simpler structures and retained bioactivities serves as another key research direction for oridonin modification and diversification. The antiproliferative activities of compounds 1–18 against various human cancer cell lines are summarized in Table 1, which indicates that oridonin is a privileged scaffold with chemical space for diverse structural optimization and drug property enhancement.
3. Antiinflammation Effects of Oridonin and Its Analogs
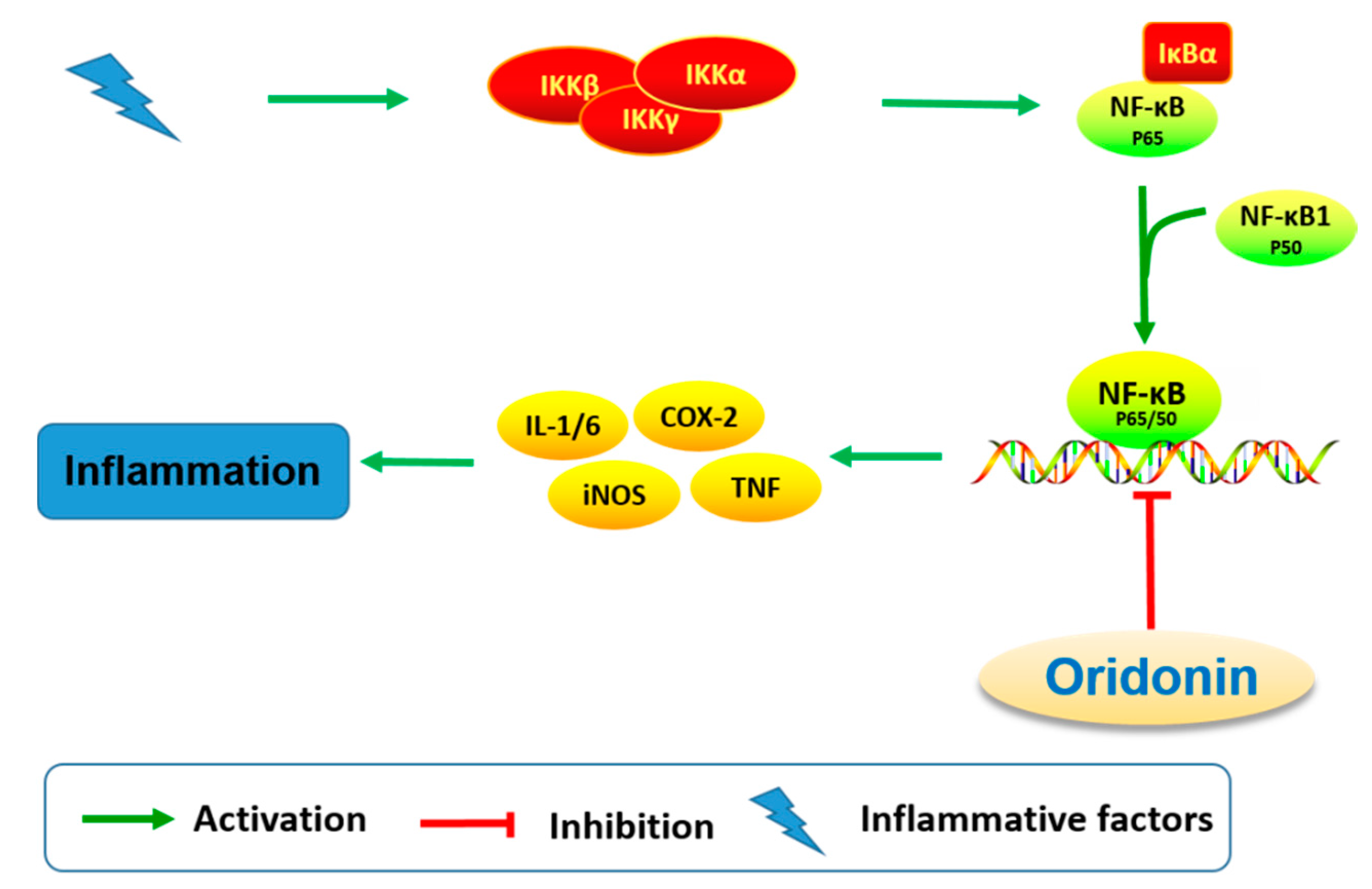
The effect on immune and pro-inflammatory mediators is another important bioactivity of oridonin (Figure 6). Studies have shown that oridonin can promote the differentiation of T cells towards CD4+/CD5+ Tregs, increase the secretion of IL-10, and modulate the Th1/Th2 balance via inducing HO-1 [19]. The effect of oridonin on intracellular tumor necrosis factor-α (TNF-α) expression was investigated and results showed that oridonin enhances endogenous pro-TNF-α expression and increases its upstream protein IκB phosphorylation [60]. Oridonin was reported to suppress the expression of inducible nitric oxide (iNOS) and cyclooxygenase-2 (COX-2) by inhibiting NF-κB DNA binding activity in HepG2, RAW264.7, and Jurkat cells [61,62]. Several groups have shown that oridonin and its water-soluble derivative (HAO472, Figure 1) might ameliorate TNBS-induced colitis by decreasing Th1/Th17 via inhibiting NF-κB signaling, subsequently reducing TNF-α, TNF-γ, IL-17A, iNOS/COX-2, and lymphocyte proliferation [52,63].
Additionally, it has been reported that oridonin and derivatives 7 (Figure 3) and 12 (Figure 4) exhibited anti-fibrogenic activities for the treatment of hepatic fibrosis [22,23,24,25]. The anti-fibrogenic effects of oridonin, 7 and 12 were investigated in the activated human LX-2 and rat HST-T6 stellate cell lines. The results showed that 7 and 12 significantly inhibited LX-2 cell proliferation in a dose- and time-dependent manner with IC50 values of 0.7 μM and 0.49 μM for 48 h, which were respectively 10-fold and 15-fold higher potency than oridonin (7.5 μM). Similar results were observed for 7 and 12 when compared to oridonin in HSC-T6 cells. However, no significant antiproliferative effects were observed on the human hepatocyte cell line C3A. These two derivatives were found to induce LX-2 cell apoptosis and S-phase cell cycle arrest and were associated with the activation of p53, p21, and cleaved caspase-3. It was also shown that 7 and 12 may mitigate endogenous production of α-SMA and ECM proteins type I collagen and fibronectin and inhibit TGF-β induced type I collagen and fibronectin production at much lower concentrations compared to oridonin. Thus, oridonin and its derivatives may hold great potential as antifibrogenic agents for the treatment of hepatic fibrosis.
4. Neuroinflammation and Neuroprotection Activities
Based on the anti-inflammatory properties of oridonin, its effects on neuroinflammation have been investigated by several research groups [27,64]. Microglia is regarded as the resident macrophage-like cell in the CNS, and can be activated by brain injury, infection, and various neuroinflammatory stimuli, consequently releasing proinflammatory and cytotoxic factors including nitric oxide (NO), TNF-α, interleukin-1β (IL-1β), interleukin-6 (IL-6), ROS, and eicosanoids [65,66,67,68,69]. Microglial activation has been observed in many neurological disorders and is noteworthy for its inflammatory and/or neurotrophic effects [69,70,71]. In LPS-activated microglia, oridonin pretreatment inhibits the release of proinflammatory mediators including NO, TNF-α, IL-1β, and IL-6 [64]. Suppression of proinflammatory mediators is accompanied by the inhibition of NF-κB DNA binding activity. Additionally, oridonin upregulates the expression of nerve nuclear growth factor (NGF), an essential neurotrophic factor for neuron survival and differentiation. These findings suggest that oridonin may have anti-neuroinflammatory and neuroregulatory effects (Figure 7) through modulation of multiple microglial pathways.
Oridonin was reported to suppress microglia and astrocyte activation in the hippocampus of the Aβ1–42 induced Alzheimer’s disease (AD) mouse model [27]. A range of activities were reported including decreasing the mRNA levels of IL-1β, IL-6, COX-2, iNOS, TNF-α, and MCP-1, upregulating the expression of IL-10, inhibiting NF-κB p65 nuclear translocation via attenuating Aβ1–42 induced IκBα phosphorylation and degradation, attenuating mitochondrial dysfunction, and reducing cognitive impairment in an Aβ1–42 induced AD mouse model [27]. In conclusion, this study has provided evidence that oridonin and its new analogues may inhibit neuroinflammation and attenuate memory deficits induced by Aβ1–42.
β-Amyloid (Aβ)-mediated synaptic dysfunction plays a critical role in the pathophysiology of AD, but the underlying mechanisms for this process remain unknown [72,73,74,75]. Xu and colleagues found that oridonin diminished synaptic dysfunction induced by Aβ1–42 in vivo and in vitro and rescued the dendritic morphological changes observed in the hippocampus of an AD mouse model. In addition, oridonin increased the expression of PSD-95 and synaptophysin and ameliorated the Aβ-induced reduction of mitochondrial activity in the synaptosomes of an AD mouse model [26]. The expression of BDNF and its receptor TrkB is ubiquitous in the brain, and the BDNF/TrkB signaling pathway has been shown to mediate the survival and differentiation of neurons, long-term potentiation, as well as plasticity [76,77]. In addition, the BDNF/TrkB pathway has been shown to play a modulatory role in learning and memory [78]. Oridonin was found to activate the BDNF/TrkB pathway and increase p-CREB expression in the hippocampus of the Aβ-induced AD mouse model, providing insight to a possible mechanism for its neuroprotective effects [26]. Additionally, in the Morris water maze test, oridonin suppressed escaping latency and searching distance and increased the number of platform crosses in the AD mouse model. These results support that oridonin can attenuate synaptic loss and promote behavioral measures in an Aβ1–42 induced AD mouse model.
5. Oridonin for Neurodegenerative Diseases
Neurodegenerative disorders are a heterogeneous group of diseases that display diverse etiologies and may impact both the CNS and the peripheral nervous system (PNS) [79,80,81]. Major neurodegenerative diseases include AD, Parkinson’s disease (PD), Huntington’s disease, multiple sclerosis, and the prion diseases. Characteristic symptoms of these diseases may include anxiety, depression, motor dysfunction, memory loss, and cognitive impairment. The causes of neurodegenerative diseases are highly diverse and may include both hereditary or environmental factors and toxic, metabolic, or infectious processes [82]. Neuronal cell damage or death is an important factor in the progression of various neurodegenerative disorders. Thus, oxidative stress, neuroinflammation, mitochondrial dysfunction, and apoptosis are major pathways responsible for neurodegeneration [83]. Several transcription factors play a role in the pathophysiology of neuronal cell damage including Nrf2, NF-κB, MAPKs, CREB, Wnt, JAK/STAT, and TLR-4, etc. [84,85,86]. Multiple therapeutic options are available that attempt to slow disease progression or control disease symptoms, such as dopaminergic treatments, acetylcholinesterase inhibitors, NMDA receptor antagonists, antipsychotic drugs, and brain stimulation [87,88,89,90]. In addition, riluzole, non-steroidal anti-inflammatory drugs, CERE-120, and caffeine A2A receptor antagonists have been used to reduce the risk of neurodegenerative diseases onset [91]. However, none of these therapies has been effective in halting the progression of neurodegenerative diseases such as AD and PD, due to their complex pathological underpinnings. The long-term use of these drugs may also produce various negative side effects. Hence, there is a need to develop safer, multi-targeted, and more effective drugs for the treatment of neurodegenerative diseases [92,93].
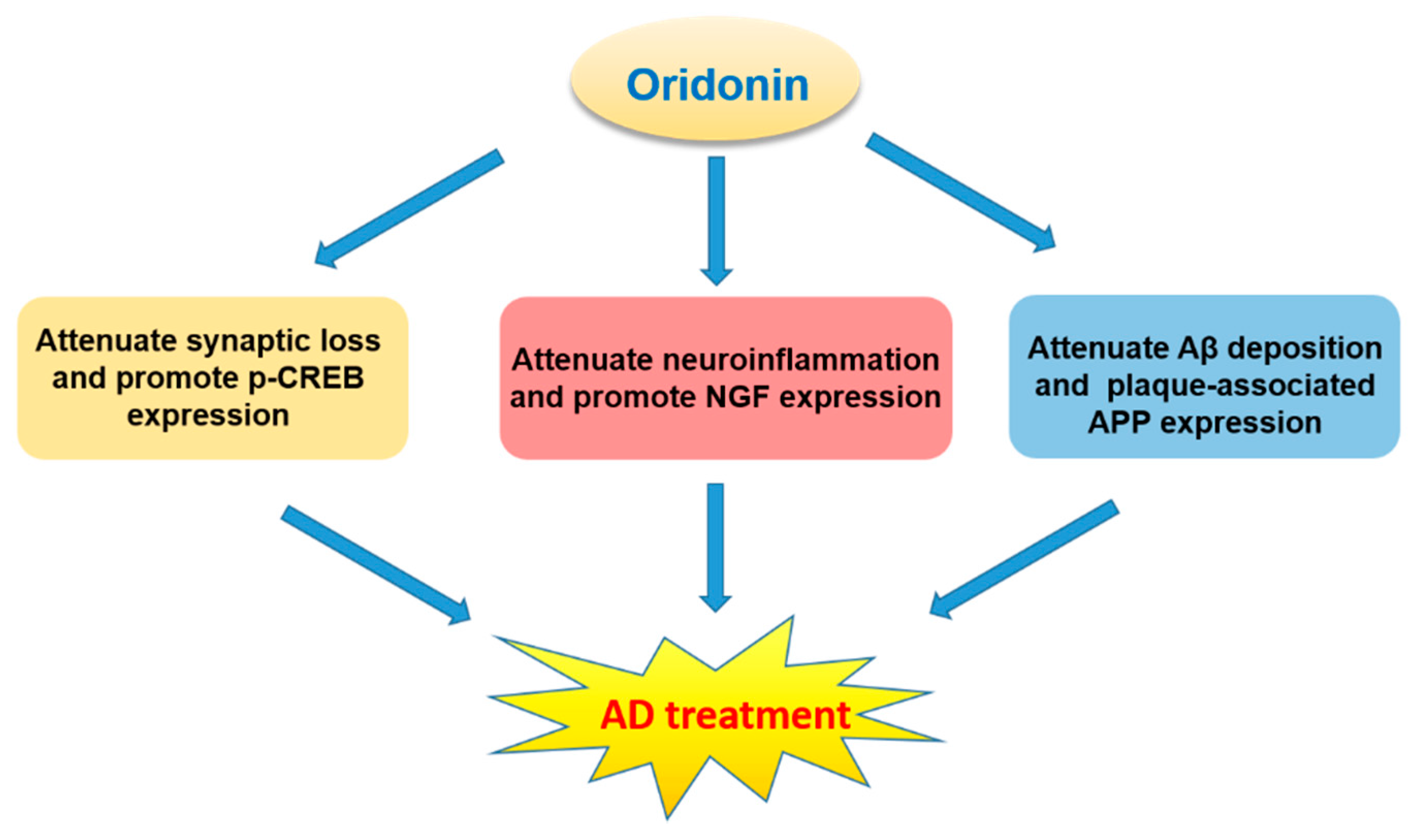
Oridonin was found to inhibit LPS-activated microglia inflammation and Aβ1–42 induced neuroinflammation, prevent synaptic loss, suppress the NF-κB pathway, and activate BDNF/TrkB/CREB and Nrf2 signaling pathways [26,27,64,94,95,96,97]. These neuroprotective effects suggest that oridonin may hold promise for the treatment of neurodegenerative diseases, especially AD (Figure 8).
AD, the primary cause of dementia, is an irreversible neurodegenerative disorder with progressive cognitive dysfunction, memory impairment, and behavioral maladaptions. The pathological features of AD are comprised of Aβ plaques (deposition of extracellular Aβ) and neurofibrillary tangles (NFTs, accumulation of intracellular hyperphosphorylated tau protein). The interesting possibility for oridonin to treat AD has already been investigated in animal models by several groups. For instance, oridonin attenuates memory and cognitive deficits in Aβ1–42 induced AD mouse models [26,27]. An oral administration of an oridonin suspension significantly attenuated Aβ aggregation, plaque-associated APP expression, and microglial activation in both the cortex and hippocampus of transgenic APP/PS1 mice at 5 months of age [28]. Further, injection of an oridonin-nanoemulsion suppressed deficits in nesting (an important affiliative behavior) and social interaction. These pathological and behavioral effects of oridonin may be due to its polypharmacology and to its modulation of multiple mechanisms/factors including reduced inflammatory activation of glial cells and immune cells, decreased Aβ deposition and APP expression directly or indirectly, as well as possible neuroprotective effects via modulating microglial function and reducing local production of proinflammatory factors. With continued research, oridonin holds potential to be developed as a therapeutic option for human AD or other neurodegenerative disorders.
6. Conclusions and Future Directions
Oridonin, a natural product commonly used in East Asian herbal medicine, has drawn increased attention in recent years due to its extensive biological activities and potential in the treatment of various diseases. Its unique, relatively safe, and remarkable anticancer pharmacological profile are noteworthy for drug discovery campaigns. A number of oridonin derivatives were designed and synthesized to pursue more potent and drug-like candidates for cancer therapy [30]. However, the exact mechanisms by which oridonin exerts these activities were inadequately understood. During the process of investigating oridonin’s mechanism of action, a variety of potential targets and signaling pathways associated with oridonin have been identified [31]. Recently, oridonin was found to ameliorate TNBS-induced colitis and inhibit HSC proliferation and fibrogenesis [25,63]. Several studies have shown that oridonin inhibits neuroinflammation, prevents synaptic loss, and regulates several targets and signaling pathways involved in the pathophysiology of neurodegenerative diseases [26,27,64]. In addition, it may ameliorate neuropathological changes and behavioral deficits in a mouse model of cerebral amyloidosis [28]. These results suggest oridonin may have the potential to treat human AD or other neurodegenerative disorders.
Oridonin displays limited aqueous solubility, low bioavailability via oral administration (F = 4.3%) or intraperitoneal injection (F = 12.6%), and high first-pass effects [45,98]. Although high lipophilicity is favorable for blood-brain barrier (BBB) permeability, limited aqueous solubility and bioavailability will decrease resultant therapeutic effects in vivo. These properties of oridonin will hamper its further clinical development as a neuroprotective agent. The accumulated SAR studies show that modifications on the A-ring system and C-14 position of oridonin may significantly improve its biological activities and aqueous solubility. Compound 3 with a thiazole fused A-ring and an additional nitrogen-containing side chain displays improved potency and aqueous solubility [45]. In addition, the introduction of hydrophilic groups (e.g., HAO472) or PEGylation at the C-14 position could also be a promising method to improve the absorption and distribution properties of oridonin derivatives [32,99,100]. Another useful strategy is the use of nanotechnology-based drug delivery approaches that may enhance drug solubility and bioavailability, improve permeability, and control drug release [101,102,103,104,105,106,107,108]. Additionally, nanostructured carriers such as nanosuspension, nanogels, and nanoparticles may prove to be an interesting strategy to afford a safe and effective delivery vehicle to overcome oral and CNS barriers [101]. In short, more attention should be directed towards the enhancement of pharmacokinetic properties in developing oridonin and its derivatives as neuroprotective agents. We believe that oridonin and its analogs have the potential to extend their application from anticancer and antiinflammation to neuroprotection, and may open new avenues to potential neurotherapeutics that can eventually benefit the patients with CNS disorders.
Acknowledgments
This work was supported by grants P30 DA028821, and R01 DA038446 from the National Institutes of Health, Breast Cancer Research Program (BCRP) Breakthrough Awards (BC160038 and BC160038P1) from the Department of Defense (DoD), Cancer Prevention Research Institute of Texas (CPRIT) award, R. A. Welch Foundation Chemistry and Biology Collaborative Grant from the Gulf Coast Consortia (GCC), and a training fellowship from the Center for Addiction Research (NIDA grant T32 DA07287), John Sealy Memorial Endowment Fund, and Institute for Translational Sciences (ITS) at UTMB.
Author Contributions
Jimin Xu and Jia Zhou designed the review. Jimin Xu and Ye Ding collected the literature. Jimin Xu, Eric A. Wold, Ye Ding, Qiang Shen and Jia Zhou wrote and corrected the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| α-SMA | α-Smooth muscle actin |
| APP | Amyloid precursor protein |
| Bax | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| BDNF | Brain-derived neurotrophic factor |
| CREB | cAMP response element-binding protein |
| ECM | Extracellular matrix |
| HSC | Hepatic stellate cell |
| ip | Intraperitoneal |
| iv | Intravenous |
| JAK | Janus kinase |
| JNK | c-Jun N-terminal kinase |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Monocyte chemotactic protein 1 |
| NMDA | N-Methyl-d-aspartic acid |
| Nrf2 | Nuclear factor erythroid-derived 2 (NFE2) related factor 2 |
| p21 | p21Cip1 protein |
| p53 | Tumor protein p53 |
| PI3K | Phosphatidylinositide 3-kinases |
| SAR | Structure-activity relationship |
| STAT | Signal transducer and activator of transcription |
| TLR-4 | Toll-like receptor 4 |
| TNBS | 2,4,6-Trinitrobenzenesulfonic acid |
| TrkB | Tropomysin receptor kinase B |
References
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, B.B.; Tiwari, V.K. Natural products: An evolving role in future drug discovery. Eur. J. Med. Chem. 2011, 46, 4769–4807. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Koehn, F.E.; Carter, G.T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Eder, J.; Sedrani, R.; Wiesmann, C. The discovery of first-in-class drugs: Origins and evolution. Nat. Rev. Drug Discov. 2014, 13, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Lu, J.; Huang, M.; Li, Y.; Chen, M.; Wu, G.; Gong, J.; Zhong, Z.; Xu, Z.; Dang, Y.; et al. Anti-cancer natural products isolated from Chinese medicinal herbs. Chin. Med. 2011, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Fujita, E.; Nagao, Y.; Kaneko, K.; Nakazawa, S.; Kuroda, H. The antitumor and antibacterial activity of the Isodon diterpenoids. Chem. Pharm. Bull. (Tokyo) 1976, 24, 2118–2127. [Google Scholar] [CrossRef] [PubMed]
- Fujita, E.; Fujita, T.; Katayama, H.; Shibuya, M. Oridonin, a new diterpenoid from Isodon species. Chem. Commun. (London) 1967, 0, 252–254. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Wang, S.; Gao, Y.; Zhang, X.; Lu, C. Oridonin phosphate-induced autophagy effectively enhances cell apoptosis of human breast cancer cells. Med. Oncol. 2015, 32, 365. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lu, X.J.; Zhang, J.; Diao, H.; Li, G.; Xu, L.; Wang, T.; Wei, J.; Meng, W.; Ma, J.L.; et al. Oridonin, a novel lysine acetyltransferases inhibitor, inhibits proliferation and induces apoptosis in gastric cancer cells through p53- and caspase-3-mediated mechanisms. Oncotarget 2016, 7, 22623–22631. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.B.; Kang, H.; Wang, L.; Gao, L.; Liu, P.; Xie, J.; Zhang, F.X.; Weng, X.Q.; Shen, Z.X.; Chen, J.; et al. Oridonin, a diterpenoid extracted from medicinal herbs, targets AML1-ETO fusion protein and shows potent antitumor activity with low adverse effects on t(8;21) leukemia in vitro and in vivo. Blood 2007, 109, 3441–3450. [Google Scholar] [CrossRef] [PubMed]
- Bao, R.; Shu, Y.; Wu, X.; Weng, H.; Ding, Q.; Cao, Y.; Li, M.; Mu, J.; Wu, W.; Ding, Q.; et al. Oridonin induces apoptosis and cell cycle arrest of gallbladder cancer cells via the mitochondrial pathway. BMC Cancer 2014, 14, 217. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Tashiro, S.; Onodera, S.; Minami, M.; Ikejima, T. Oridonin induced autophagy in human cervical carcinoma HeLa cells through Ras, JNK, and P38 regulation. J. Pharmacol. Sci. 2007, 105, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Liu, F.; Li, M. Inhibition of nuclear factor kappaB transcription activity drives a synergistic effect of cisplatin and oridonin on HepG2 human hepatocellular carcinoma cells. Anticancer Drugs 2016, 27, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yun, E.J.; Chen, W.; Ding, Y.; Wu, K.; Wang, B.; Ding, C.; Hernandez, E.; Santoyo, J.; Pong, R.C.; et al. Targeting 3-phosphoinositide-dependent protein kinase 1 associated with drug-resistant renal cell carcinoma using new oridonin analogs. Cell Death Dis. 2017, 8, e2701. [Google Scholar] [CrossRef] [PubMed]
- Ikezoe, T.; Chen, S.S.; Tong, X.J.; Heber, D.; Taguchi, H.; Koeffler, H.P. Oridonin induces growth inhibition and apoptosis of a variety of human cancer cells. Int. J. Oncol. 2003, 23, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhou, J.; Wu, K.; Huang, J.; Ding, Y.; Yun, E.J.; Wang, B.; Ding, C.; Hernandez, E.; Santoyo, J.; et al. Targeting XBP1-mediated β-catenin expression associated with bladder cancer with newly synthetic oridonin analogues. Oncotarget 2016, 7, 56842–56854. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.-M.; Lin, J.-Y. Anti-inflammatory effects of 27 selected terpenoid compounds tested through modulating Th1/Th2 cytokine secretion profiles using murine primary splenocytes. Food Chem. 2013, 141, 1104–1113. [Google Scholar] [CrossRef] [PubMed]
- Hu, A.P.; Du, J.M.; Li, J.Y.; Liu, J.W. Oridonin promotes CD4+/CD25+ Treg differentiation, modulates Th1/Th2 balance and induces HO-1 in rat splenic lymphocytes. Inflamm. Res. 2008, 57, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yang, F.; Zhang, Y.; Li, J. Studies on the cell-immunosuppressive mechanism of Oridonin from Isodon serra. Int. Immunopharmacol. 2007, 7, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Pei, L.; Li, D.; Yao, H.; Cai, H.; Yao, H.; Wu, X.; Xu, J. Synthesis and antimycobacterial evaluation of natural oridonin and its enmein-type derivatives. Fitoterapia 2014, 99, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Bohanon, F.J.; Wang, X.; Graham, B.M.; Prasai, A.; Vasudevan, S.J.; Ding, C.; Ding, Y.; Radhakrishnan, G.L.; Rastellini, C.; Zhou, J.; et al. Enhanced anti-fibrogenic effects of novel oridonin derivative CYD0692 in hepatic stellate cells. Mol. Cell. Biochem. 2015, 410, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Bohanon, F.J.; Wang, X.; Graham, B.M.; Ding, C.; Ding, Y.; Radhakrishnan, G.L.; Rastellini, C.; Zhou, J.; Radhakrishnan, R.S. Enhanced effects of novel oridonin analog CYD0682 for hepatic fibrosis. J. Surg. Res. 2015, 199, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Kuo, L.M.; Kuo, C.Y.; Lin, C.Y.; Hung, M.F.; Shen, J.J.; Hwang, T.L. Intracellular glutathione depletion by oridonin leads to apoptosis in hepatic stellate cells. Molecules 2014, 19, 3327–3344. [Google Scholar] [CrossRef] [PubMed]
- Bohanon, F.J.; Wang, X.; Ding, C.; Ding, Y.; Radhakrishnan, G.L.; Rastellini, C.; Zhou, J.; Radhakrishnan, R.S. Oridonin inhibits hepatic stellate cell proliferation and fibrogenesis. J. Surg. Res. 2014, 190, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yu, L.; Yang, H.; Li, C.; Hui, Z.; Xu, Y.; Zhu, X. Oridonin Attenuates synaptic loss and cognitive deficits in an Abeta1–42-induced mouse model of Alzheimer’s disease. PLoS ONE 2016, 11, e0151397. [Google Scholar]
- Wang, S.; Yang, H.; Yu, L.; Jin, J.; Qian, L.; Zhao, H.; Xu, Y.; Zhu, X. Oridonin attenuates Abeta1–42-induced neuroinflammation and inhibits NF-kappaB pathway. PLoS ONE 2014, 9, e104745. [Google Scholar]
- Zhang, Z.Y.; Daniels, R.; Schluesener, H.J. Oridonin ameliorates neuropathological changes and behavioural deficits in a mouse model of cerebral amyloidosis. J. Cell. Mol. Med. 2013, 17, 1566–1576. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Han, T.; Liao, J.; Hu, X.; Xu, S.; Tian, K.; Gu, X.; Cheng, K.; Li, Z.; Hua, H.; et al. Oridonin, a Promising ent-Kaurane Diterpenoid Lead Compound. Int. J. Mol. Sci. 2016, 17, 1395. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Ding, C.; Ye, N.; Liu, Z.; Wold, E.A.; Chen, H.; Wild, C.; Shen, Q.; Zhou, J. Discovery and development of natural product oridonin-inspired anticancer agents. Eur. J. Med. Chem. 2016, 122, 102–117. [Google Scholar] [CrossRef] [PubMed]
- Owona, B.A.; Schluesener, H.J. Molecular Insight in the Multifunctional Effects of oridonin. Drugs R&D 2015, 15, 233–244. [Google Scholar]
- Sun, P.; Wu, G.; Qiu, Z.; Chen, Y. Preparation of l-Alanine-(14-oridonin) Ester Trifluoroacetate for Treatment of Cancer. Patent Application CN104017000A, 3 September 2014. [Google Scholar]
- Li, C.-Y.; Wang, E.-Q.; Cheng, Y.; Bao, J.-K. Oridonin: An active diterpenoid targeting cell cycle arrest, apoptotic and autophagic pathways for cancer therapeutics. Int. J. Biochem. Cell Biol. 2011, 43, 701–704. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Gao, J.; Halicka, H.D.; Huang, X.; Traganos, F.; Darzynkiewicz, Z. The cytostatic and cytotoxic effects of oridonin (rubescenin), a diterpenoid from Rabdosia rubescens, on tumor cells of different lineage. Int. J. Oncol. 2005, 26, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ye, Y.; Chui, J.H.; Zhu, G.Y.; Li, Y.W.; Fong, D.W.; Yu, Z.L. Oridonin induces G2/M cell cycle arrest and apoptosis through MAPK and p53 signaling pathways in HepG2 cells. Oncol. Rep. 2010, 24, 647–651. [Google Scholar] [PubMed]
- Cheng, Y.; Qiu, F.; Ye, Y.-C.; Tashiro, S.-i.; Onodera, S.; Ikejima, T. Oridonin induces G2/M arrest and apoptosis via activating ERK–p53 apoptotic pathway and inhibiting PTK–Ras–Raf–JNK survival pathway in murine fibrosarcoma L929 cells. Arch. Biochem. Biophys. 2009, 490, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.Z.; Yang, Y.B.; Xu, X.D.; Shen, H.W.; Shu, Y.M.; Ren, Z.; Li, X.M.; Shen, H.M.; Zeng, H.T. Oridonin induces apoptosis via PI3K/Akt pathway in cervical carcinoma HeLa cell line. Acta Pharmacol. Sin. 2007, 28, 1819–1826. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-Q.; Mu, Z.-Q.; You, S.; Tashiro, S.-I.; Onodera, S.; Ikejima, T. Fas/FasL Signaling Allows Extracelluar-Signal Regulated Kinase to Regulate Cytochrome c Release in Oridonin-Induced Apoptotic U937 Cells. Biol. Pharm. Bull. 2006, 29, 1873–1879. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ding, Y.; Chen, C.H.; Zhou, Z.; Ding, C.; Chen, H.; Zhou, J.; Chen, C. A new oridonin analog suppresses triple-negative breast cancer cells and tumor growth via the induction of death receptor 5. Cancer Lett. 2016, 380, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-L.; Wu, L.-J.; Tashiro, S.-I.; Onodera, S.; Ikejima, T. Oridonin induced A375-S2 cell apoptosis via BAX-regulated caspase pathway activation, dependent on the cytochrome C/CASPASE-9 apoptosome. J. Asian Nat. Prod. Res. 2004, 6, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-L.; Wu, L.-J.; Zuo, H.-J.; Tashiro, S.-I.; Onodera, S.; Ikejima, T. Cytochrome c release from oridonin-treated apoptotic A375-S2 Cells is dependent on p53 and Extracellular signal-regulated kinase activation. J. Pharmacol. Sci. 2004, 96, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-Q.; You, S.; Tashiro, S.-I.; Onodera, S.; Ikejima, T. Activation of Phosphoinositide 3-kinase, protein kinase C, and extracellular signal-regulated kinase is required for oridonin-enhanced phagocytosis of apoptotic bodies in human macrophage-like U937 Cells. J. Pharmacol. Sci. 2005, 98, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.H.; Liu, F.; Wei, W.; Liu, L.B.; Xu, M.H.; Guo, Z.Y.; Li, W.; Jiang, B.; Wu, Y.L. Oridonin induces apoptosis and senescence by increasing hydrogen peroxide and glutathione depletion in colorectal cancer cells. Int. J. Mol. Med. 2012, 29, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.N.; Huang, J.; Yang, J.; Tashiro, S.; Onodera, S.; Ikejima, T. Caspase inhibition augmented oridonin-induced cell death in murine fibrosarcoma l929 by enhancing reactive oxygen species generation. J. Pharmacol. Sci. 2008, 108, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Zhang, Y.; Chen, H.; Yang, Z.; Wild, C.; Chu, L.; Liu, H.; Shen, Q.; Zhou, J. Novel nitrogen-enriched oridonin analogues with thiazole-fused A-ring: Protecting group-free synthesis, enhanced anticancer profile, and improved aqueous solubility. J. Med. Chem. 2013, 56, 5048–5058. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Wang, L.; Chen, H.; Wild, C.; Ye, N.; Ding, Y.; Wang, T.; White, M.A.; Shen, Q.; Zhou, J. ent-Kaurane-based regio- and stereoselective inverse electron demand hetero-Diels-Alder reactions: synthesis of dihydropyran-fused diterpenoids. Org. Biomol. Chem. 2014, 12, 8442–8452. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Zhang, Y.; Chen, H.; Yang, Z.; Wild, C.; Ye, N.; Ester, C.D.; Xiong, A.; White, M.A.; Shen, Q.; et al. Oridonin ring A-based diverse constructions of enone functionality: Identification of novel dienone analogues effective for highly aggressive breast cancer by inducing apoptosis. J. Med. Chem. 2013, 56, 8814–8825. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Zhang, Y.; Chen, H.; Wild, C.; Wang, T.; White, M.A.; Shen, Q.; Zhou, J. Overcoming synthetic challenges of oridonin A-ring structural diversification: Regio- and stereoselective installation of azides and 1,2,3-triazoles at the C-1, C-2, or C-3 position. Org. Lett. 2013, 15, 3718–3721. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Luo, S.; Yao, H.; Cai, H.; Miao, X.; Wu, F.; Yang, D.H.; Wu, X.; Xie, W.; Yao, H.; et al. Probing the anticancer action of oridonin with fluorescent analogues: visualizing subcellular localization to mitochondria. J. Med. Chem. 2016, 59, 5022–5034. [Google Scholar] [CrossRef] [PubMed]
- Nan, F.; Zhang, Y.; Liu, H.; Ding, J.; Meng, L.; Xu, C. Preparation of ent-Kaurane Type Diterpenoid Derivatives as Antitumor Agents. Patent Application CN102584760A, 18 July 2012. [Google Scholar]
- Huang, S.X.; Xiao, W.L.; Li, L.M.; Li, S.H.; Zhou, Y.; Ding, L.S.; Lou, L.G.; Sun, H.D. Bisrubescensins A-C: Three new dimeric ent-kauranoids isolated from Isodon rubescens. Org. Lett. 2006, 8, 1157–1160. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.Q.; Wang, H.L.; Chen, K.; Wang, S.B.; Xu, Y.; Ye, Q.; Sun, Y.W. Oridonin derivative ameliorates experimental colitis by inhibiting activated T-cells and translocation of nuclear factor-kappa B. J. Dig. Dis. 2016, 17, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Yao, H.; Luo, S.; Zhang, Y.K.; Yang, D.H.; Li, D.; Wang, G.; Hu, M.; Qiu, Y.; Wu, X.; et al. A Novel Potent Anticancer Compound Optimized from a Natural Oridonin Scaffold Induces Apoptosis and Cell Cycle Arrest through the Mitochondrial Pathway. J. Med. Chem. 2017, 60, 1449–1468. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, D.; Xu, S.; Cai, H.; Yao, H.; Zhang, Y.; Jiang, J.; Xu, J. The conversion of oridonin to spirolactone-type or enmein-type diterpenoid: Synthesis and biological evaluation of ent-6,7-seco-oridonin derivatives as novel potential anticancer agents. Eur. J. Med. Chem. 2012, 52, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Cai, H.; Jiang, B.; Liu, G.; Wang, Y.; Wang, L.; Yao, H.; Wu, X.; Sun, Y.; Xu, J. Synthesis of spirolactone-type diterpenoid derivatives from kaurene-type oridonin with improved antiproliferative effects and their apoptosis-inducing activity in human hepatoma Bel-7402 cells. Eur. J. Med. Chem. 2013, 59, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xu, S.; Cai, H.; Pei, L.; Zhang, H.; Wang, L.; Yao, H.; Wu, X.; Jiang, J.; Sun, Y.; et al. Enmein-type diterpenoid analogs from natural kaurene-type oridonin: Synthesis and their antitumor biological evaluation. Eur. J. Med. Chem. 2013, 64, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xu, S.; Cai, H.; Pei, L.; Wang, L.; Wu, X.; Yao, H.; Jiang, J.; Sun, Y.; Xu, J. Library construction and biological evaluation of enmein-type diterpenoid analogues as potential anticancer agents. ChemMedChem 2013, 8, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, Y.; Lu, W.; Nan, F.J. Synthesis and revision of stereochemistry of rubescensin S. Org. Biomol. Chem. 2011, 9, 4436–4439. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jiang, Y.; Li, Q.; Xiao, Q.; Jia, Y.; Tu, P. Formal synthesis of semiaquilegin A. Tetrahedron Lett. 2010, 51, 1121–1123. [Google Scholar] [CrossRef]
- Huang, J.; Wu, L.; Tashiro, S.; Onodera, S.; Ikejima, T. A comparison of the signal pathways between the TNF alpha- and oridonin-induced murine L929 fibrosarcoma cell death. Acta Med. Okayama 2005, 59, 261–270. [Google Scholar] [PubMed]
- Leung, C.H.; Grill, S.P.; Lam, W.; Han, Q.B.; Sun, H.D.; Cheng, Y.C. Novel mechanism of inhibition of nuclear factor-kappa B DNA-binding activity by diterpenoids isolated from Isodon rubescens. Mol. Pharmacol. 2005, 68, 286–297. [Google Scholar] [PubMed]
- Ikezoe, T.; Yang, Y.; Bandobashi, K.; Saito, T.; Takemoto, S.; Machida, H.; Togitani, K.; Koeffler, H.P.; Taguchi, H. Oridonin, a diterpenoid purified from Rabdosia rubescens, inhibits the proliferation of cells from lymphoid malignancies in association with blockade of the NF-kappa B signal pathways. Mol. Cancer Ther. 2005, 4, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Y.; Saas, P.; Wang, H.; Xu, Y.; Chen, K.; Zhong, J.; Yuan, Y.; Wang, Y.; Sun, Y. Oridonin’s therapeutic effect: suppressing Th1/Th17 simultaneously in a mouse model of Crohn’s disease. J. Gastroenterol. Hepatol. 2015, 30, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xue, Y.; Wang, Y.; Feng, D.; Lin, S.; Xu, L. Multiple-modulation effects of Oridonin on the production of proinflammatory cytokines and neurotrophic factors in LPS-activated microglia. Int. Immunopharmacol. 2009, 9, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Minagar, A.; Shapshak, P.; Fujimura, R.; Ownby, R.; Heyes, M.; Eisdorfer, C. The role of macrophage/microglia and astrocytes in the pathogenesis of three neurologic disorders: HIV-associated dementia, Alzheimer disease, and multiple sclerosis. J. Neurol. Sci. 2002, 202, 13–23. [Google Scholar] [CrossRef]
- Streit, W.J. Microglial response to brain injury: A brief synopsis. Toxicol. Pathol. 2000, 28, 28–30. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Scarano, F.; Baltuch, G. Microglia as mediators of inflammatory and degenerative diseases. Annu. Rev. Neurosci. 1999, 22, 219–240. [Google Scholar] [CrossRef] [PubMed]
- Kreutzberg, G.W. Microglia: A sensor for pathological events in the CNS. Trends Neurosci. 1996, 19, 312–318. [Google Scholar] [CrossRef]
- Minghetti, L.; Levi, G. Microglia as effector cells in brain damage and repair: focus on prostanoids and nitric oxide. Prog. Neurobiol. 1998, 54, 99–125. [Google Scholar] [CrossRef]
- McGeer, P.L.; Yasojima, K.; McGeer, E.G. Association of interleukin-1 beta polymorphisms with idiopathic Parkinson’s disease. Neurosci. Lett. 2002, 326, 67–69. [Google Scholar] [CrossRef]
- Le, W.; Rowe, D.; Xie, W.; Ortiz, I.; He, Y.; Appel, S.H. Microglial activation and dopaminergic cell injury: An in vitro model relevant to Parkinson’s disease. J. Neurosci. 2001, 21, 8447–8455. [Google Scholar] [PubMed]
- Masliah, E.; Mallory, M.; Alford, M.; DeTeresa, R.; Hansen, L.A.; McKeel, D.W. Jr.; Morris, J.C. Altered expression of synaptic proteins occurs early during progression of Alzheimer’s disease. Neurology 2001, 56, 127–129. [Google Scholar] [CrossRef] [PubMed]
- Sze, C.I.; Troncoso, J.C.; Kawas, C.; Mouton, P.; Price, D.L.; Martin, L.J. Loss of the presynaptic vesicle protein synaptophysin in hippocampus correlates with cognitive decline in Alzheimer disease. J. Neuropathol. Exp. Neurol. 1997, 56, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Dickson, D.W.; Crystal, H.A.; Bevona, C.; Honer, W.; Vincent, I.; Davies, P. Correlations of synaptic and pathological markers with cognition of the elderly. Neurobiol. Aging 1995, 16, 285–298. [Google Scholar] [CrossRef]
- Davies, C.A.; Mann, D.M.; Sumpter, P.Q.; Yates, P.O. A quantitative morphometric analysis of the neuronal and synaptic content of the frontal and temporal cortex in patients with Alzheimer’s disease. J. Neurol. Sci. 1987, 78, 151–164. [Google Scholar] [CrossRef]
- Chen, K.; Henry, R.A.; Hughes, S.M.; Connor, B. Creating a neurogenic environment: The role of BDNF and FGF2. Mol. Cell. Neurosci. 2007, 36, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Rivera, C.; Li, H.; Thomas-Crusells, J.; Lahtinen, H.; Viitanen, T.; Nanobashvili, A.; Kokaia, Z.; Airaksinen, M.S.; Voipio, J.; Kaila, K.; et al. BDNF-induced TrkB activation down-regulates the K+–Cl− cotransporter KCC2 and impairs neuronal Cl− extrusion. J. Cell Biol. 2002, 159, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Nabeshima, T. Brain-derived neurotrophic factor/TrkB signaling in memory processes. J. Pharmacol. Sci. 2003, 91, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Hardiman, O.; Doherty, C.P.; Elamin, M.; Bede, P. Neurodegenerative Disorders: A Clinical Guide; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Chang, R.C.-C. Advanced Understanding of Neurodegenerative Diseases; InTech: Rijeka, Croatia, 2011; p. 1. [Google Scholar]
- Chang, R.C.-C. Neurodegenerative Diseases: Processes, Prevention, Protection and Monitoring; InTech: Rijeka, Croatia, 2011. [Google Scholar]
- Losada-Barreiro, S.; Bravo-Diaz, C. Free radicals and polyphenols: The redox chemistry of neurodegenerative diseases. Eur. J. Med. Chem. 2017, 133, 379–402. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.L.; Yang, C.M. Role of redox signaling in neuroinflammation and neurodegenerative diseases. Biomed. Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Sowndhararajan, K.; Kim, S. Neuroprotective and cognitive enhancement potentials of Angelica gigas Nakai root: A review. Sci. Pharm. 2017, 85, 21. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, C.Z.; Ding, C.; Wild, C.; Copits, B.; Swanson, G.T.; Johnson, K.M.; Zhou, J. A combined bioinformatics and chemoinformatics approach for developing asymmetric bivalent AMPA receptor positive allosteric modulators as neuroprotective agents. ChemMedChem 2013, 8, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Ansari, N.; Khodagholi, F. Natural products as promising drug candidates for the treatment of Alzheimer’s disease: Molecular mechanism aspect. Curr. Neuropharmacol. 2013, 11, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Okun, M.S. Deep-brain stimulation—Entering the era of human neural-network modulation. N. Engl. J. Med. 2014, 371, 1369–1373. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, Y. Recent research progress in and future perspective on treatment of Parkinson’s disease. Integr. Med. Int. 2014, 1, 67–79. [Google Scholar] [CrossRef]
- Doody, R.; Ferris, S.; Salloway, S.; Sun, Y.; Goldman, R.; Watkins, W.; Xu, Y.; Murthy, A. Donepezil treatment of patients with MCI A 48-week randomized, placebo-controlled trial. Neurology 2009, 72, 1555–1561. [Google Scholar] [CrossRef] [PubMed]
- Grossberg, G.T. Diagnosis and treatment of Alzheimer’s disease. J. Clin. Psychiatry 2003, 64 (Suppl. 9), 3–6. [Google Scholar] [PubMed]
- Chen, X.; Pan, W. The treatment strategies for neurodegenerative diseases by integrative medicine. Integr. Med. Int. 2014, 1, 223–225. [Google Scholar] [CrossRef]
- Fang, J.; Pang, X.; Yan, R.; Lian, W.; Li, C.; Wang, Q.; Liu, A.-L.; Du, G.-H. Discovery of neuroprotective compounds by machine learning approaches. RSC Adv. 2016, 6, 9857–9871. [Google Scholar] [CrossRef]
- Fang, J.; Li, Y.; Liu, R.; Pang, X.; Li, C.; Yang, R.; He, Y.; Lian, W.; Liu, A.L.; Du, G.H. Discovery of multitarget-directed ligands against Alzheimer’s disease through systematic prediction of chemical-protein interactions. J. Chem. Inf. Model. 2015, 55, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Role of Nrf2 in Oxidative Stress and Toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Villeneuve, N.F.; Wang, X.J.; Sun, Z.; Chen, W.; Li, J.; Lou, H.; Wong, P.K.; Zhang, D.D. Oridonin confers protection against arsenic-induced toxicity through activation of the Nrf2-mediated defensive response. Environ. Health Perspect. 2008, 116, 1154–1161. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Li, C.Y.; Lu, J.M.; Tian, R.B.; Wei, J. Allicin ameliorates cognitive deficits ageing-induced learning and memory deficits through enhancing of Nrf2 antioxidant signaling pathways. Neurosci. Lett. 2012, 514, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Sun, J.; Zhang, T.T.; Ma, B.; Cui, S.M.; Chen, D.W.; He, Z.G. Pharmacokinetic behaviors and oral bioavailability of oridonin in rat plasma. Acta Pharmacol. Sin. 2006, 27, 1642–1646. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zhang, D.; Zhao, Z.; Jia, L.; Zheng, D.; Liu, G.; Hao, L.; Zhang, Q.; Tian, X.; Li, C.; et al. Synthesis, characterization, in vitro and in vivo evaluation of PEGylated oridonin conjugates. Int. J. Pharm. 2013, 456, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Pasut, G.; Veronese, F.M. PEG conjugates in clinical development or use as anticancer agents: An overview. Adv. Drug Del. Rev. 2009, 61, 1177–1188. [Google Scholar] [CrossRef] [PubMed]
- Ganta, S.; Deshpande, D.; Korde, A.; Amiji, M. A review of multifunctional nanoemulsion systems to overcome oral and CNS drug delivery barriers. Mol. Membr. Biol. 2010, 27, 260–273. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Duan, C.; Zhang, D.; Jia, L.; Liu, G.; Liu, Y.; Wang, F.; Li, C.; Guo, H.; Zhang, Q. Galactosylated chitosan nanoparticles for hepatocyte-targeted delivery of oridonin. Int. J. Pharm. 2012, 436, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.M.; Cullis, P.R. Liposomal drug delivery systems: From concept to clinical applications. Adv. Drug Del. Rev. 2013, 65, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Zhang, D.; Xu, X.; Feng, F.; Ren, G.; Chu, Q.; Zhang, Q.; Tian, K. Oridonin nanosuspension was more effective than free oridonin on G2/M cell cycle arrest and apoptosis in the human pancreatic cancer PANC-1 cell line. Int. J. Nanomed. 2012, 7, 1793–1804. [Google Scholar]
- Jia, L.; Shen, J.; Zhang, D.; Duan, C.; Liu, G.; Zheng, D.; Tian, X.; Liu, Y.; Zhang, Q. In vitro and in vivo evaluation of oridonin-loaded long circulating nanostructured lipid carriers. Int. J. Biol. Macromol. 2012, 50, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liu, Y.; Feng, N.; Xu, J. Preparation and evaluation of self-microemulsifying drug delivery system of oridonin. Int. J. Pharm. 2008, 355, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, P.; Feng, N.; Zhang, X.; Wu, S.; Zhao, J. Optimization and in situ intestinal absorption of self-microemulsifying drug delivery system of oridonin. Int. J. Pharm. 2009, 365, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Zhang, D.; Chen, M.; Duan, C.; Dai, W.; Jia, L.; Zhao, W. Studies on pharmacokinetics and tissue distribution of oridonin nanosuspensions. Int. J. Pharm. 2008, 355, 321–327. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds 3–10 and 12 are available from the authors. |
Figure 1.
The structures of oridonin and HAO472.
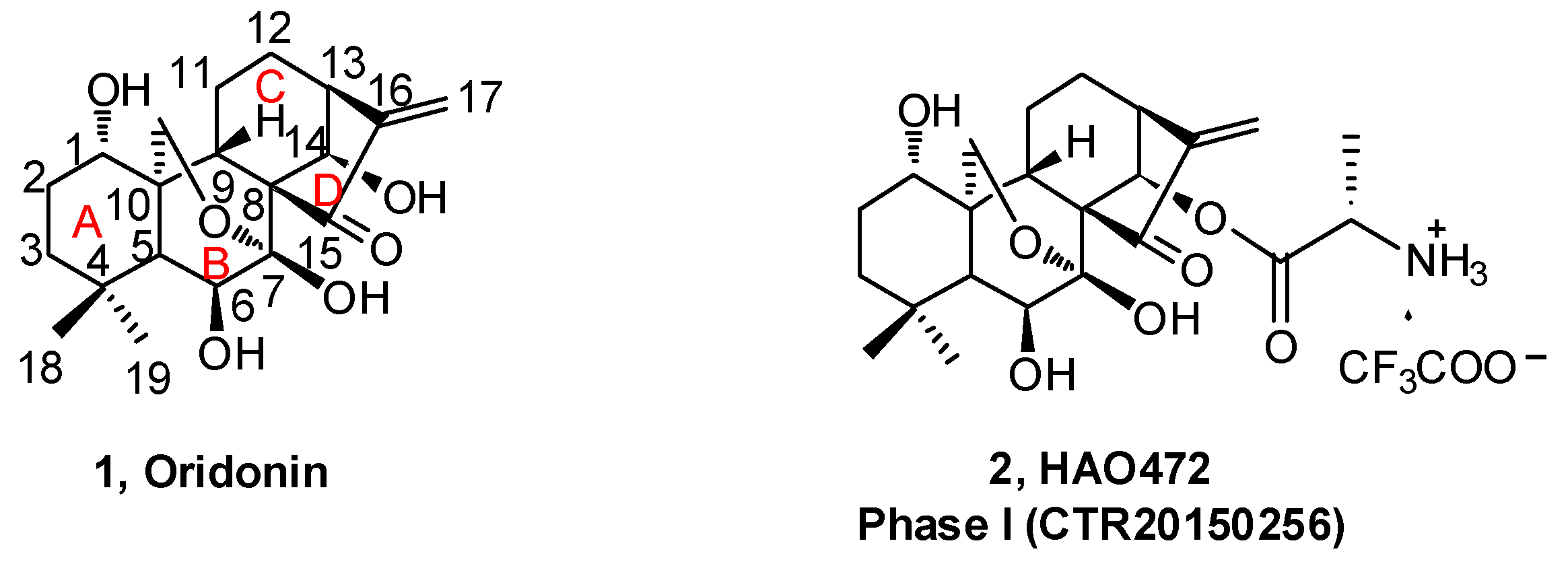
Figure 2.
Oridonin regulates multi-signaling pathways related to autophagy, apoptosis, phagocytosis, and cell cycle arrest.
Figure 2.
Oridonin regulates multi-signaling pathways related to autophagy, apoptosis, phagocytosis, and cell cycle arrest.
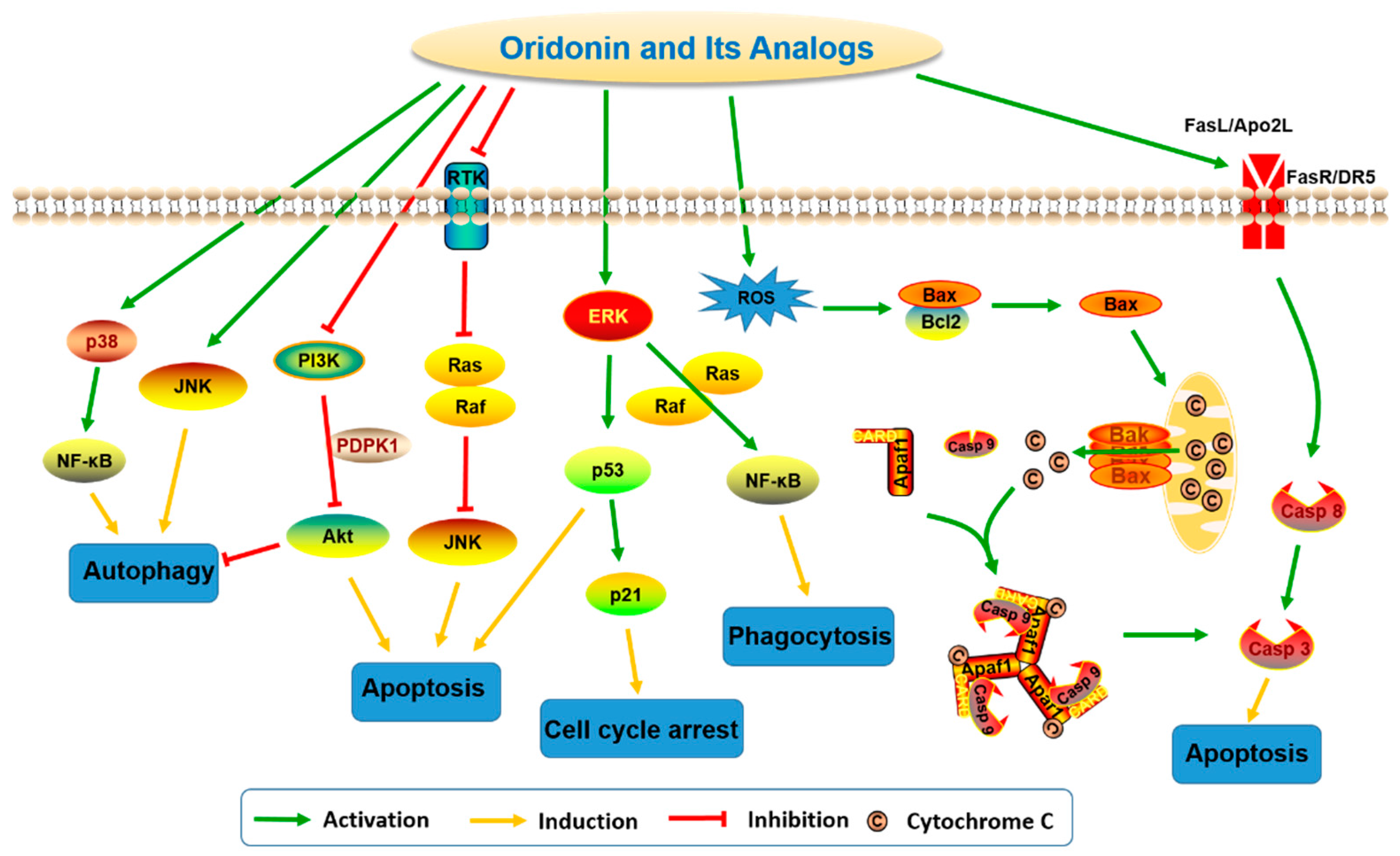
Figure 3.
Diversified modifications on A-ring of oridonin.
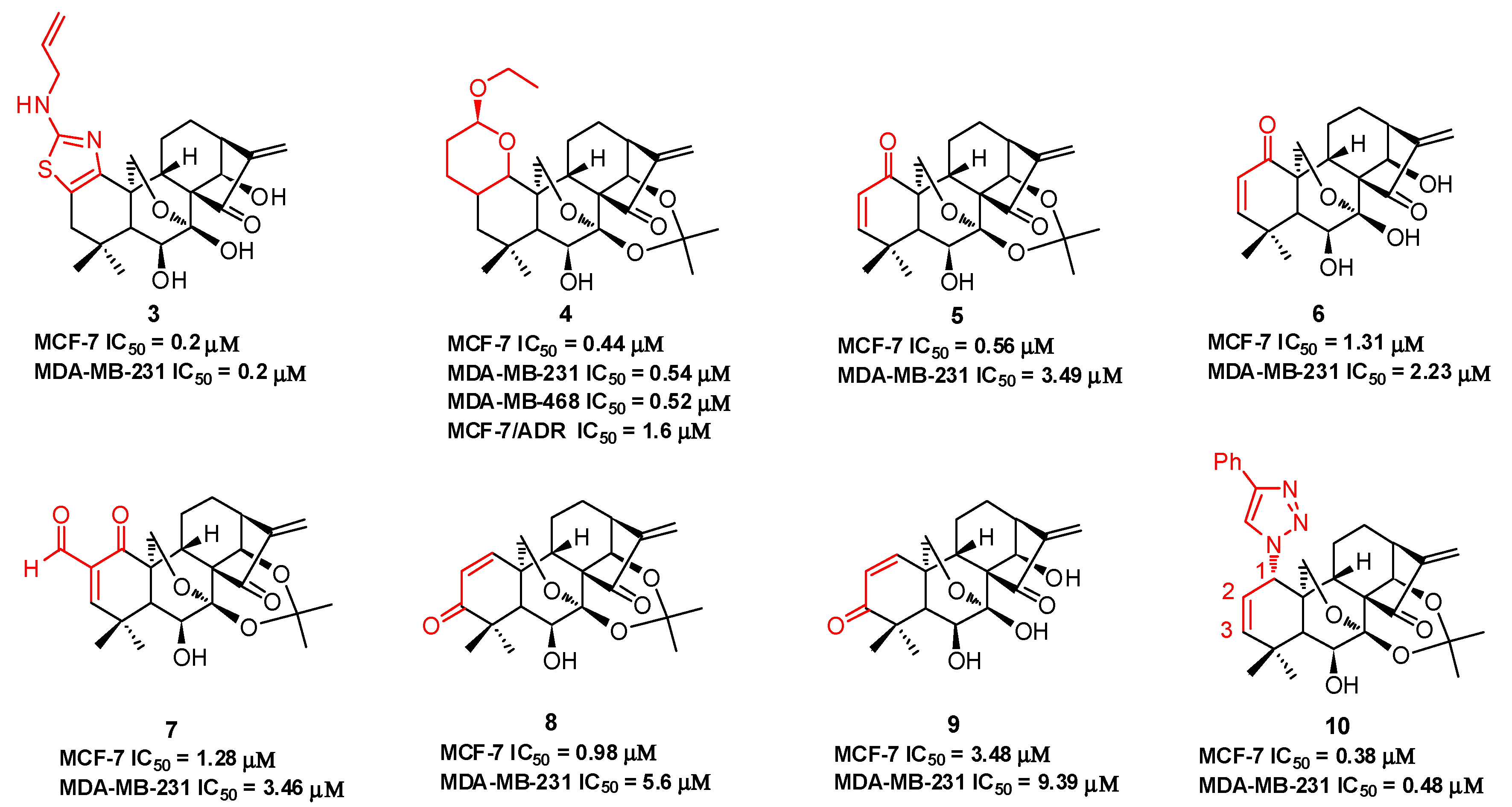
Figure 4.
The structures of C-1 and C-14 modified oridonin derivatives.
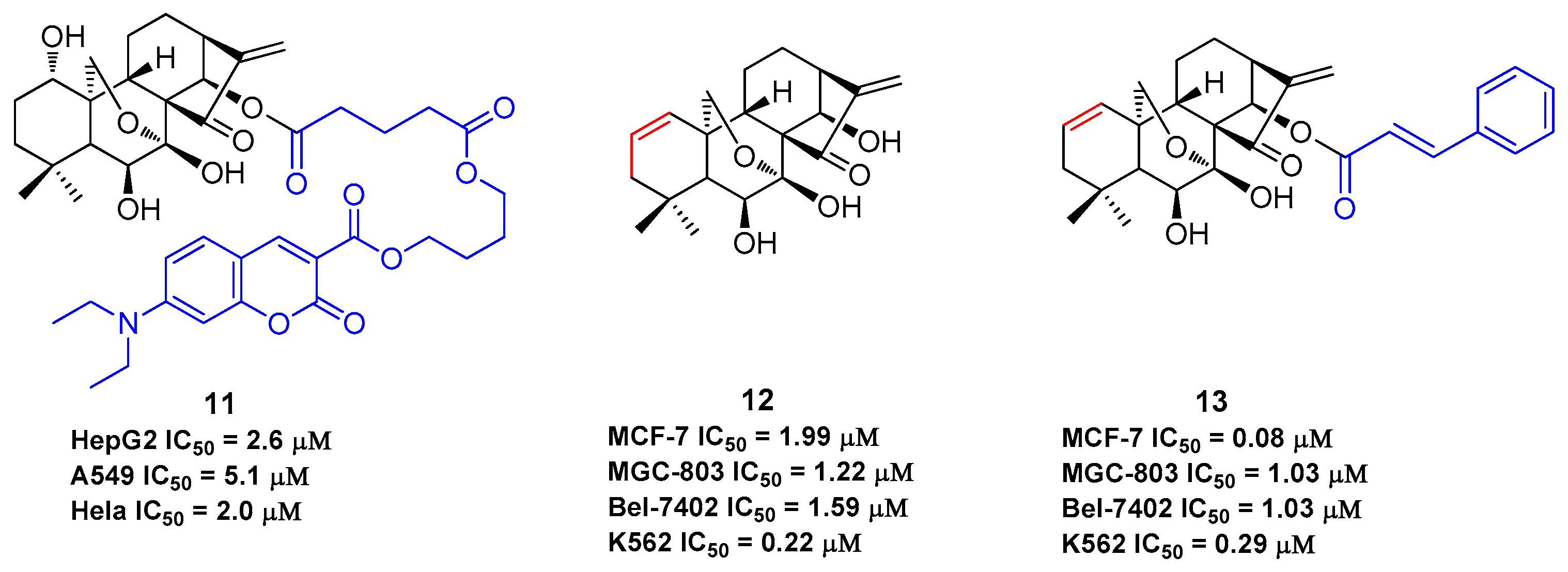
Figure 5.
Spirolactone-type and enmein-type diterpenoid derivatives generated from oridonin.
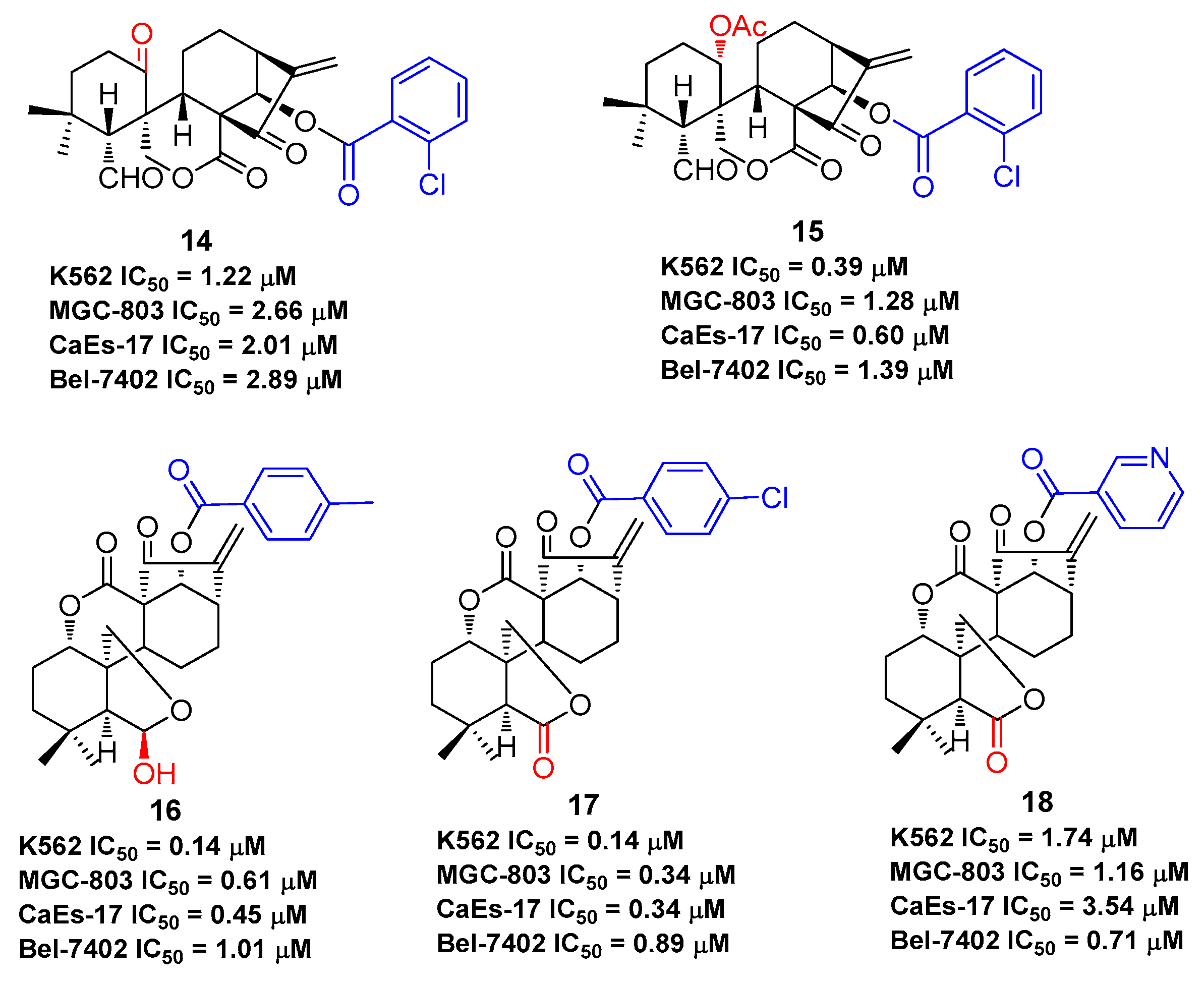
Figure 6.
Oridonin regulates signaling pathway related to inflammation.
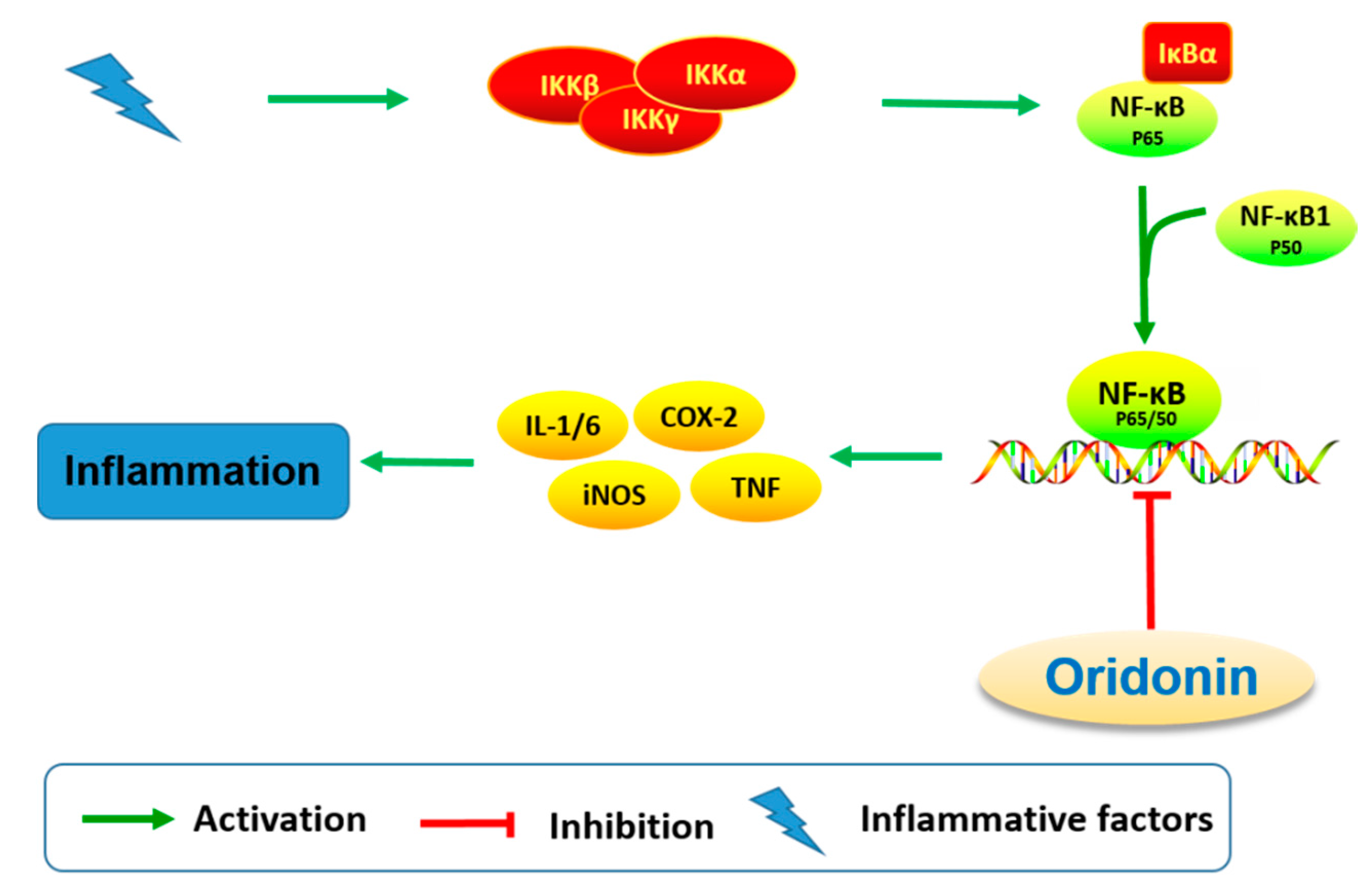
Figure 7.
Oridonin regulates signaling pathways related to anti-neuroinflammation and neuroprotection.
Figure 7.
Oridonin regulates signaling pathways related to anti-neuroinflammation and neuroprotection.
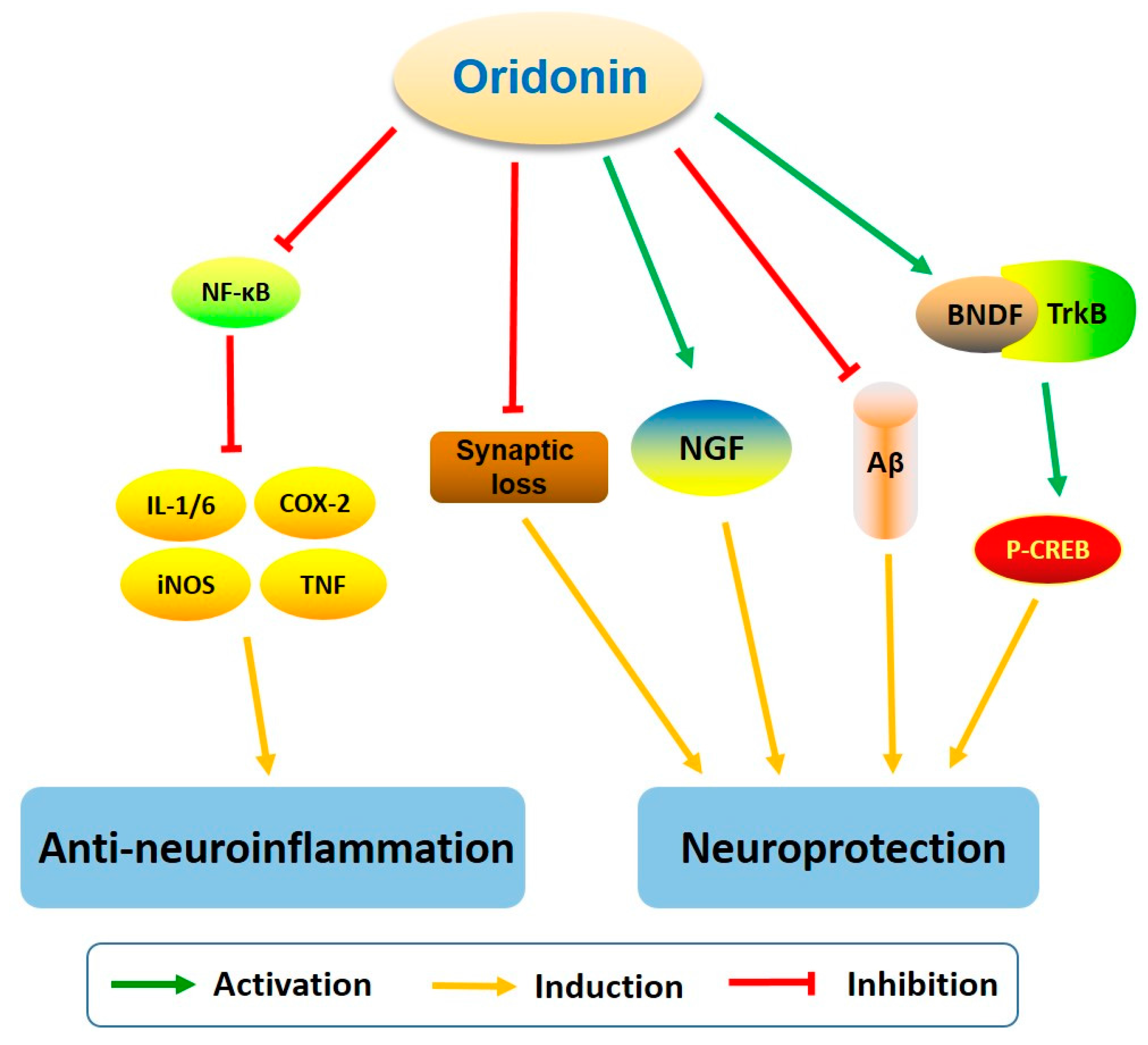
Figure 8.
The characteristics of oridonin and its potential for AD treatment.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The antiproliferative effects of compounds 1–18 against various human cancer cell lines.
| Compd. | IC50 | Ref. |
|---|---|---|
| Oridonin (1) | MCF-7: 6.6 μM; MDA-MB-231: 29.4 μM; MDA-MB-468: 5.3 μM; MCF-7/ADR: 34.8 μM; HepG2: 15.2 μM; MGC-803: 9.06 μM; Bel-7402: 5.41 μM; K562: 4.33 μM | [45,46,47,48,49,53] |
| HAO472 (2) | Not disclosed Phase I (CTR20150246) in China: leukemia | * |
| 3 | MCF-7: 0.2 μM; MDA-MB-231: 0.2 μM; AsPC1: 1.1 μM; Panc-1: 1.1 μM; DU145: 1.2 μM | [45] |
| 4 | MCF-7: 0.44 μM; MDA-MB-231: 0.54 μM; MDA-MB-468: 0.52 μM; MCF-7/ADR: 1.6 μM | [46] |
| 5 | MCF-7: 0.56 μM; MDA-MB-231: 3.49 μM; MCF-7/ADR: 5.03 μM | [47] |
| 6 | MCF-7: 1.31 μM; MDA-MB-231: 2.23 μM; MCF-7/ADR: 5.82 μM | [47] |
| 7 | MCF-7: 1.28 μM; MDA-MB-231: 3.46 μM; MCF-7/ADR: 6.55 μM | [47] |
| 8 | MCF-7: 0.98 μM; MDA-MB-231: 5.60 μM; MCF-7/ADR: 6.02 μM | [47] |
| 9 | MCF-7: 3.48 μM; MDA-MB-231: 9.39 μM | [47] |
| 10 | MCF-7: 3.48 μM; MDA-MB-231: 9.39 μM | [48] |
| 11 | HepG2: 2.6 μM; A549: 5.1 μM; Hela: 2.0 μM | [49] |
| 12 | MCF-7: 1.99 μM; MGC-803: 1.22 μM; Bel-7402: 1.59 μM; K562: 0.22 μM | [53] |
| 13 | MCF-7: 0.08 μM; MGC-803: 1.03 μM; Bel-7402: 1.03 μM; K562: 0.29 μM | [53] |
| 14 | K562: 1.22 μM; MGC-803: 2.66 μM; CaEs-17: 2.01 μM; Bel-7402: 2.89 μM | [54] |
| 15 | K562: 0.39 μM; MGC-803: 1.28 μM; CaEs-17: 0.60 μM; Bel-7402: 1.39 μM | [55] |
| 16 | K562: 0.14 μM; MGC-803: 0.61 μM; CaEs-17: 0.45 μM; Bel-7402: 1.01 μM | [56] |
| 17 | K562: 0.14 μM; MGC-803: 0.34 μM; CaEs-17: 0.34 μM; Bel-7402: 0.89 μM | [57] |
| 18 | K562: 1.74 μM; MGC-803: 1.16 μM; CaEs-17: 3.54 μM; Bel-7402: 0.71 μM | [57] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xu, J.; Wold, E.A.; Ding, Y.; Shen, Q.; Zhou, J. Therapeutic Potential of Oridonin and Its Analogs: From Anticancer and Antiinflammation to Neuroprotection. Molecules 2018, 23, 474. https://doi.org/10.3390/molecules23020474
AMA Style
Xu J, Wold EA, Ding Y, Shen Q, Zhou J. Therapeutic Potential of Oridonin and Its Analogs: From Anticancer and Antiinflammation to Neuroprotection. Molecules. 2018; 23(2):474. https://doi.org/10.3390/molecules23020474
Chicago/Turabian StyleXu, Jimin, Eric A. Wold, Ye Ding, Qiang Shen, and Jia Zhou. 2018. "Therapeutic Potential of Oridonin and Its Analogs: From Anticancer and Antiinflammation to Neuroprotection" Molecules 23, no. 2: 474. https://doi.org/10.3390/molecules23020474